

Abstract
The development of the brain is sex-dimorphic, and as a result so are many neurological disorders. One approach for studying sex-dimorphic brain development is to measure gene expression in biological samples using RT-qPCR. However, the accuracy and consistency of this technique relies on the reference gene(s) selected. We analyzed the expression of ten reference genes in male and female samples over three stages of brain development, using popular algorithms NormFinder, GeNorm and Bestkeeper. The top ranked reference genes at each time point were further used to quantify gene expression of three sex-dimorphic genes (Wnt10b, Xist and CYP7B1). When comparing gene expression between the sexes expression at specific time points the best reference gene combinations are: Sdha/Pgk1 at E11.5, RpL38/Sdha E12.5, and Actb/RpL37 at E15.5. When studying expression across time, the ideal reference gene(s) differs with sex. For XY samples a combination of Actb/Sdha. In contrast, when studying gene expression across developmental stage with XX samples, Sdha/Gapdh were the top reference genes. Our results identify the best combination of two reference genes when studying male and female brain development, and emphasize the importance of selecting the correct reference genes for comparisons between developmental stages.
Keywords: Reference gene, Sex-dimorphic, Brain, Development, Mice, RT-qpcr
Introduction
Mammals that reproduce through sexual reproduction have intrinsic differences between male and female sexes. The most obvious morphological differences are those of reproductive organs. However, many other less apparent organs and tissues are also modulated in a sex-dimorphic manner. In particular, there exist numerous differences in the human brain between the male and female sexes. The rate of maturation of the female brain reaches its peak maturity at 10 years of age compared to the male brain, which only peaks 4 years later (Giedd et al., 1997). On average females have a higher percentage of gray matter although volume is smaller in comparison (Goldstein et al., 2001; Gur et al., 1999) neuronal cells in the hypothalamus may only be responsive to estrogen or testosterone (Arnold, 2004; Lenroot et al., 2007). Furthermore, numerous neurological diseases have sex-specific biases (Fombonne, 2009), including, schizophrenia (risk ratio 1 female:1.4 male) (Abel, Drake & Golds, 2010; Saha et al., 2005) and Autism spectrum disorder (1 female:4 male) (Mottron et al., 2015). In order to understand how such differences arise at the level of gene expression, we set out to determine the best set of reference genes to study mouse sex-dimorphic brain development during key developmental stages.
The house mouse (Mus musculus) is an excellent choice for embryonic studies of mammalian development owing to its comparatively short gestation period and accelerated life span. In particular, mice have been widely used to study sex-dimorphic brain development (Maekawa et al., 2014; Ngun et al., 2011). To acquire more insight into the molecular drives of sex-dimorphic brain development, it is necessary to study the expression of genes in the developing brain. High-throughput sequencing technologies such as RNA sequencing (RNA-seq) provide a powerful technique to study changes in gene expression (Hrdlickova, Toloue & Tian, 2017). However, data gathered from these high-throughput technologies needs to be validated to ensure accurate interpretation through repeated biological replicates.
The most commonly used method for validating the expression of a gene identified by sequencing is Reverse Transcription quantitative Polymerase Chain Reaction (RT-qPCR). RT-qPCR allows for the detection and quantification of specific cDNA fragments generated from RNA samples. However, to obtain levels of expression comparable between samples, the target gene must be normalized to the expression of at least two internal controls (termed reference genes) that are stably expressed throughout all samples. Normalization is needed to compensate for different amounts of cDNA present in the sample along with differing PCR efficiencies of primer sets. Therefore, the selection of the reference gene is important as inappropriate reference genes can bias the data and thus lead to misinterpretation of results.
Ideally, the reference gene should be present at a consistent level across all compared samples, regardless of treatment or disease state of the sample. Furthermore, the chosen reference gene should be constitutively expressed across all cell types and tissues. However despite large-scale high-throughput technologies, no such gene has been found. Therefore the most common and validated approach is to find a reference gene that is the least variable in the specific context of the study.
The most regularly used reference genes are Actb and Gapdh (Boda et al., 2009). However, a number of studies have shown that these genes are expressed differentially in the brain. Gapdh has been shown to have sex dimorphic protein levels in adulthood (Perrot-Sinal, Davis & McCarthy, 2001) and to be up regulated in neuronal apoptosis (Chen et al., 1999; Sawa et al., 1997). Actb and Gapdh expression has also been shown to vary across tissue types, among cell types and also during stages of cell proliferation and development of the brain (Sotelo-Silveira et al., 2008; Veazey & Golding, 2011).
To fill in the gap in the literature, we set out to find a set of reference genes most suitable for studies on the embryonic brain from E11.5 to E15.5 in males and females. This work will prove highly valuable unbiased study to uncover suitable reference genes for studies of embryonic brain development and illustrate the importance of the accurate selection of reference genes for RT-qPCR analyses.
Methods
All animal work was performed under the University of Otago Animal Ethics Committee number: ET13/14. Inbred C57BL/6 mice were purchased from the Hercus Taieri Resource Unit (University of Otago, Dunedin, NZ).
Sample collection
Whole embryonic brain tissue (overlying epidermis and cranial facial tissues were removed) and tail tips were collected from timed stages (E11.5 to 18.5) pregnant mothers. RNA was extracted from brain tissue using Purelink RNA mini kit (Ambion, USA) according to manufacturer’s instructions. DNA was isolated from tail tips using 0.2 mg/mL (final concentration) Proteinase K (New England Biolabs, Ipswich, MA, USA) and then added to DirectPCR Lysis reagent 102-T (Viagen Biotechnologies, Los Angeles, CA, USA). Tail tips were incubated overnight at 55°C and followed by heat inactivation of the proteinase K at 85°C for 45 min. Samples were centrifuged at 14,000 g for 1 min to pellet cell debris and 2 µL of each sample was used for sexing PCR. The RNA for each time point and sex was a pooled sample, with a minimum of three separate biological samples collected for each condition (from separate litters). Total RNA was quantified with a Nanodrop (ThermoFisher, Waltham, MA, USA) and purity assessed using the 260/230 and 260/280 ratios. A ratio of ∼2 was taken as acceptable for pure RNA.
Sexing of embryos by PCR
To sex the embryos, the Sry gene was amplified using primers listed in Table S1. Following PCR, products were run on a 2% agarose gel, a band appears at approximately 380 bp, indicating the presence of the Y-chromosome for male embryos.
DNase treatment and reverse transcription
To remove genomic DNA present in the sample, 1 µg RNA was added to 1 µL DNase and incubated at 37 °C for 40 min. A phenol/chloroform extraction was carried out followed by ethanol precipitation to purify sample. Reverse transcriptase was performed using iScript (Bio-Rad, Hercules, CA, USA) according to manufacturer’s instructions.
RT-qPCR
Oligonucleotide primers (Table S1) were designed and ordered from Integrated DNA Technologies (IDT, Newark, NJ, USA). Each reaction contained 10 µL SYBR Select Master Mix (ThermoFisher), 1.25 µL 20 pmol forward and reverse primers, 6.75 µL water and 2 µL cDNA. All reactions also included a no reverse transcription control and each reaction was carried out in triplicate. RT-qPCR was carried out in the Stratagene Mx3000p (Agilent Technologies, San Diego, CA, USA) under the following conditions: denaturing at 50°C for 2 min, annealing 96°C for 2 min, followed by 40 cycles of amplification (96°C 15 s, 60°C 15 s and 72°C 1 min). A final cycle for melt curve analysis was included for every qPCR plate (one cycle: 95°C 1 min, 55°C 30 s and 95°C 30 s). The threshold was automatically set by the MXPro program (v. 4.10), and the Ct value for each sample was calculated from the average of three technical replicates.
Data analysis
Data from RT-qPCR was then analyzed using three algorithms to calculate the most stable or best suited reference gene; NormFinder (Andersen, Jensen & Orntoft, 2004), GeNorm with SLqPCR R-based package (Hellemans et al., 2007) Bestkeeper (Pfaffl et al., 2004) and RefFinder (deltaCt method, (Silver et al., 2006; Xie et al., 2012) software). All data was analyzed according to the program instructions.
The geometric mean was calculated for each reference gene by assigning a number (1–10) for top ranked genes for each algorithm. The following equation was used to calculate the geometric mean for each data set: 3√NormFinder*GeNorm*BestKeeper*deltaCt. The reference genes were then ranked again based on their lowest geometric mean.
Analysis of sex specific gene expression was calculated using the following equation: ΔCt = 2−(Ctgene-Ctreference) with the average Ct value for the top two ranked reference genes. Statistical testing used was either the unpaired Student’s t-test when comparing two time-points or a 2-way ANOVA when comparing multiple time-points across time (Tukey’s multiple comparison test).
Results
Selection of candidate reference genes and mRNA transcript levels
To ensure accurate analysis of gene expression, ten candidate reference genes (Table 1) were investigated to determine how stable each gene is for sex and time-point of development. The candidate reference genes selected included those genes commonly used in RT-qPCR experiments (such as Gapdh and Actb) and genes were stably expressed in multiple mouse adult tissues, but had not been tested for suitability as reference genes with mouse embryo tissues (Kouadjo et al., 2007). RT-qPCR was carried out for brain tissue samples across three time points: E11.5, E12.5 and E15.5 using both male and female samples. These time points were chosen for analysis as the sex determination in mouse occurs between E11.0 and E12.0. During this 24 h period, considerable changes also occur with respect to neuronal development, in particular, the formation of the primary brain vesicles (Stiles & Jernigan, 2010). By E12.5, cellular proliferation is increased resulting in the expansion of neuronal precursors and formation of cortical layers (Finlay & Darlington, 1995). Neuronal differentiation, axonal branching and synaptogenesis is taking place around day 15.5 (summarized in Fig. 1) (Sur & Rubenstein, 2005) The raw Ct values for each reference gene across time and each sex are plotted in Fig. 2.
Table 1. Function, symbol and name of selected reference genes.
| Gene symbol | Gene description | Function |
|---|---|---|
| Gapdh | Glyceraldehyde 3-phosphate dehydrogenase | Catalyzes sixth step of glycolysis |
| Actb | Beta-actin | Formation of microfilaments in eukaryotic cells |
| Hprt1 | Hypoxanthine guanine phosphoribosyl transferase | Transferase that plays a role in the generation of purine nucleotide through the purine salvage pathway |
| Pgk1 | Phosphoglycerate kinase 1 | Part of the glycolysis pathway which catalyses the conversion of 1,3-diphogycerate to 3-phosphoglycerate |
| Sdha | Succinate dehydrogenase complex, subunit A, flavoprotein (Fp) | Citric acid cycle and the respiratory chain |
| Ppia | Peptidylprolyl isomerase A | Catalyzes cis-trans isomerization of proline imidic peptide bonds. Role in protein folding |
| RpL38 | Ribosomal protein L38 | Protein synthesis |
| RpL37 | Ribosomal protein L37 | Protein synthesis |
| Eif3f | Eukaryotic translation initiation factor 3 subunit F | Translation elongation |
| Eef2 | Eukaryotic translation elongation factor 2 | Translation elongation |
Figure 1. Brain developmental stages used in this study.

In mice gonadal sex determination occurs at ∼E11.5. At E11.5 the neural tube has formed the primary brain vesicles: Prosencephalon, Mesencephalon and Rhombencephalon. Between stages of E12.5 and E15.5, there is increased expansion of neuronal precursors and cell migration forming cortical layers. Simultaneously, neurons differentiate to allow for axonal branching and synapse formation.
Figure 2. Raw Ct values for selected reference genes from RT-qPCR using cDNA from male and female brain tissue.

(A) Raw Ct values for each reference gene are shown for female and male samples by developmental stage. Data is plotted as floating bar plot, with min and maximum values indicated and a line at the mean for each stage (E11.5, E12.5 and E15.5).
Determination of best-suited reference genes for male and female developing brain
Four algorithms: NormFinder, GeNorm, BestKeeper and comparative deltaCt were employed to determine the best-suited reference genes for either specific developmental stage or between sex over time. Female and male samples were combined to identify the most stable reference gene across time. At each developmental stage (E11.5, E12.5 and E15.5) female and male samples were compared against each reference gene to determine the best reference gene between both sexes.
The raw Ct values for each gene comparing sex-expression at each time point are shown as box and whisker plots (Fig. 2) with minimum and maximum values indicated at each time point. Ct values for male samples ranged between Ct values of 15–25.5 and female samples between 15–28 (Table S3). Across all time points, the reference gene with least amount of variability for male samples is Actb with a mean Ct value of 16.36 (±1.178 SD) while the highest is Eif3f with a mean Ct value of 30.06 (±5.08 SD). In females, Actb mean Ct value is 17.9 (±2.22 SD) and Eef2 has a mean Ct of 22.53, while this Ct value was not the highest, the SD had the highest amount of variability of ±4.54 SD.
NormFinder Excel based add-on is an algorithm created by Andersen, Jensen & Orntoft (2004) that ranks a set of candidate reference genes using stability values according to the variation in expression across samples and between groups. A low stability value represents a reference gene with the most stable expression in a given sample set. NormFinder analysis identified, that across all developmental stages of development (Fig. 3A, Table S4), the reference gene with the lowest stability value was RpL37 (1.211), the best combination of genes were Hprt1 and RpL38 (0.559) in the male embryonic brain. In female samples, Hprt1 (0.929) was the most stable gene across all time points, the best two gene combination was Sdha and RpL38 (stability value of 0.640) (Fig. 3A). NormFinder identified the lowest stability value at E11.5 to be Actb (0.309) (Fig. 3B), however Actb and Sdha (0.250) genes gave the best combined stability value (0.250) for RT-qPCR analysis. Following sex determination of the embryo, RpL38 (0.253) was the most stable reference gene at E12.5, with a combination of RpL38 and RpL37 (0.189) for use of two reference genes (Fig. 3B). RpL37 (0.145) is the most stable reference gene at E15.5 however for analyses with two reference genes then Pgk1 and Eef2 (0.430) were recommended (Table S3).
Figure 3. Comparison of gene stability values for NormFinder and BestKeeper.

(A) NormFinder stability values for male and female reference genes across all developmental stages. (B) The stability value of combined sexes calculated by NormFinder across all each developmental time point of development. (C) The best suited reference genes for male and female samples as calculated by BestKeeper shown as sdt error (±CP). (D) BestKeeper values shown as std error (±CP) across time in samples of both sexes. The lowest stability value for each sex is shown (Female, red; Male, blue; Both sexes, black).
BestKeeper software determines the ideal reference genes out of a number of candidates based on the standard error (±CP) of Ct values and combines them into an index using repeated pair-wise correlation analysis (Pfaffl et al., 2004). Across all developmental stages, the BestKeeper identified Pgk1 (SE: ±1.064) and Sdha (SE: ±1.004) as the best reference gene in the male sample, while Gapdh (SE: ±1.639) and Sdha (SE: ±1.441) to be ideal for female samples (Fig. 3C). However at specific developmental stages, the ideal reference gene differs. At E11.5 RpL38 (SE: ±0.68) is ranked the top reference gene, while Pgk1 (SE: ±0.522) and Hprt1 (SE: ±1.762) are the most stable reference genes for E12.5 and E15.5 respectively (Fig. 3D, Table S5).
The SLqPCR R-based package uses the GeNorm algorithm (Hellemans et al., 2007) which determines the most stable reference gene from a set of samples by calculating the geometric mean of each reference gene in a stepwise calculation. A low M value indicates stable expression of the gene: for homogenous samples a value below 0.5 indicates an unstable reference gene in the samples analyzed, whereas for heterogeneous tissues the mean M value below 1 is acceptable (Hellemans et al., 2007). The best stability value for the female samples across all time points was Sdha and Gapdh with a stability M value of 0.591 while the top three most stable reference genes in the male samples were Pgk1, and ActB with a mean M stability value of 0.281 (Fig. 4A, Table S6).
Figure 4. Comparison of stability values for GeNorm and deltaCt methods.

(A) Stability (M) value calculated by GeNorm across time in both male and female samples. (B) GeNorm mean stability (M) values in male and female samples across all developmental stages. (C) Average standard deviation using the deltaCt method for male, female and combined samples. (D) Average standard deviation using the deltaCt method for each time point tested. The lowest stability value for each sex is shown (Female, red; Male, blue; Both sexes, black).
At specific developmental time points, Sdha and Pgk1 (M of 0.2470) were calculated as the most stable reference genes for analysis of male and female samples at E11.5. Following sex determination in both male and female samples, Actb and Gapdh (M of 0.346) gene are the most stable (Fig. 4B). During increased neuronal proliferation at E15.5, Actb and Pgk1 (M of 0.719) gave the lowest M values, indicating these are reliable reference genes according to GeNorm analysis given at this stage the developing brain is heterogeneous (Table S6).
A forth method, termed the comparative deltaCt method (Silver et al., 2006) was also used to compare reference genes. This uses a method similar to that of GeNorm, determining the variation of gene expression between paired putative reference genes (the deltaCt) within each sample. Variable deltaCt values for each pair between multiple samples, resulting in a high standard deviation when the average deltaCt is calculated, means either gene or both are not stably expressed. Pairs at E11.5 were Pgk1 and Sdha (1.98/1.87), Actb and Pgk1 (1.98/1.82). At E15.5 there was much more variation between reference genes, best pair were Actb and Sdha (4.7) (Fig. 4D; Table S7) When considering brain development overtime, for the male samples Pgk1 and Sdha (2.1) were the best pair, whereas for female samples, Hprt1 and Sdha (2.7) had the lowest stand deviation of deltaCt values (Fig. 4C).
Geometric mean of each reference gene
Each reference gene was ranked from most stable to least stable, as determined by the four stability calculations described above, and assigned a number from 1 (most stable) to 10 (least stable) (Table S8). The geometric mean was taken for each set of rankings to calculate an overall ‘best’ reference gene for each condition (Tables 2 and 3). The recommended reference genes across all stages in both male and female samples are Sdha and RpL37 (GeoMean: 1.18 and 2.94 respectively). Across all stages of embryonic development in males, the recommended reference genes are Actb and Sdha (GeoMean: 2.73 and 1.96) while in the female brain, the most stable set of reference genes are Sdha and Gapdh (GeoMean 1.2 and 2.34).
Table 2. Geometric Mean of ranking values.
| Male | Female | All stages, male and female | ||||
|---|---|---|---|---|---|---|
| 1 | Sdha | 1.96 | Sdha | 1.20 | Sdha | 1.18 |
| 2 | Actb | 2.73 | Gapdh | 2.34 | RpL37 | 2.94 |
| 3 | Pgk1 | 2.91 | Hprt1 | 2.45 | Actb | 3.03 |
| 4 | Gapdh | 3.78 | RpL37 | 4.58 | Pgk1 | 3.10 |
| 5 | RpL37 | 4.16 | Actb | 4.60 | Hprt1 | 3.83 |
| 6 | Hprt1 | 4.64 | Pgk1 | 5.23 | RpL38 | 6 |
| 7 | Actb | 4.82 | RpL38 | 5.63 | Gapdh | 6.34 |
| 8 | Ppia | 7.95 | Ppia | 8.23 | Ppia | 7.23 |
| 9 | Eef2 | 8.27 | Eef2 | 8.73 | Eef2 | 8.30 |
| 10 | Eif3f | 10 | Eif3f | 10 | Eif3f | 10 |
Table 3. Geometric Mean of ranking values by developmental stage.
| E11.5 | E12.5 | E15.5 | ||||
|---|---|---|---|---|---|---|
| 1 | Sdha | 1.56 | RpL38 | 1.68 | RpL37 | 1.86 |
| 2 | Pgk1 | 1.86 | Sdha | 1.73 | Actb | 1.86 |
| 3 | Actb | 2.91 | Pgk1 | 3.31 | Sdha | 3.31 |
| 4 | Gapdh | 3.72 | Actb | 3.35 | Pgk1 | 3.72 |
| 5 | RpL38 | 4.3 | RpL37 | 3.89 | Hprt1 | 3.86 |
| 6 | RpL37 | 5.48 | Gapdh | 4.09 | Eef2 | 4.78 |
| 7 | Hprt1 | 5.89 | Hprt1 | 5.95 | RpL38 | 6.70 |
| 8 | Eef2 | 8.23 | Ppia | 7.44 | Gapdh | 7.02 |
| 9 | Ppia | 9.14 | Eef2 | 7.77 | Ppia | 8.20 |
| 10 | Eif3f | 9.24 | Eif3f | 10 | Eif3f | 10 |
For RT-qPCR analyses at specific developmental time points regardless of sex (Table 3), Sdha and Pgk1 (GeoMean: 1.56 and 1.86) are best suited for analysis at E11.5. Following determination of sex in the embryo at E12.5, RpL38 and Sdha with GeoMean values of 1.68 and 1.73 are recommended. Finally, RpL37 and Actb (GeoMean 1.86 and 1.86) are the best suited for analysis of E15.5 brain tissue.
In comparison to a careful sex or stage approach above, we also pooled all the data together (both sexes and all stages). Overall, this approached produced stability values that were high (indicating a great variation between samples) for GeNorm and BestKeeper packages (Tables S5 and S6). The geometric mean of ranked references genes for a pooled sample approached suggested that Sdha and RpL37 (Tables 2 and 3), despite RpL37 often being ranked a poor choice if we were just consider the stabilities values for E11.5 and E12.5 individually (Table S8 and S3).
Confirmation of sex-specific gene expression
Previous micro-array data showed sex-specific expression of a number of genes within the embryonic brain (forebrain) at E10.5 (Dewing et al., 2003). Wingless-type MMTV integration site family, member 10B (Wnt10b) was shown to be up-regulated in male embryos, with 1.7 fold increase in expression at E10.5 with respect to the female brain. Cytochrome P450,7b1 (CYP7B1) was expressed 2.1 fold higher in the male brain, whereas expression of X-inactive specific-transcript (Xist) was confirmed significantly higher in the female embryo with a fold change of 18.5 compared to male samples. However, these differences in gene expression were not studied over further time points, when neurogenesis has commenced. To extend the findings from Dewing et al. (2003), regarding sex-dimorphic expression of these genes, the top two ranked reference genes at each time point from this study are used to compare against a selection of three genes in male and female samples across embryonic brain development.
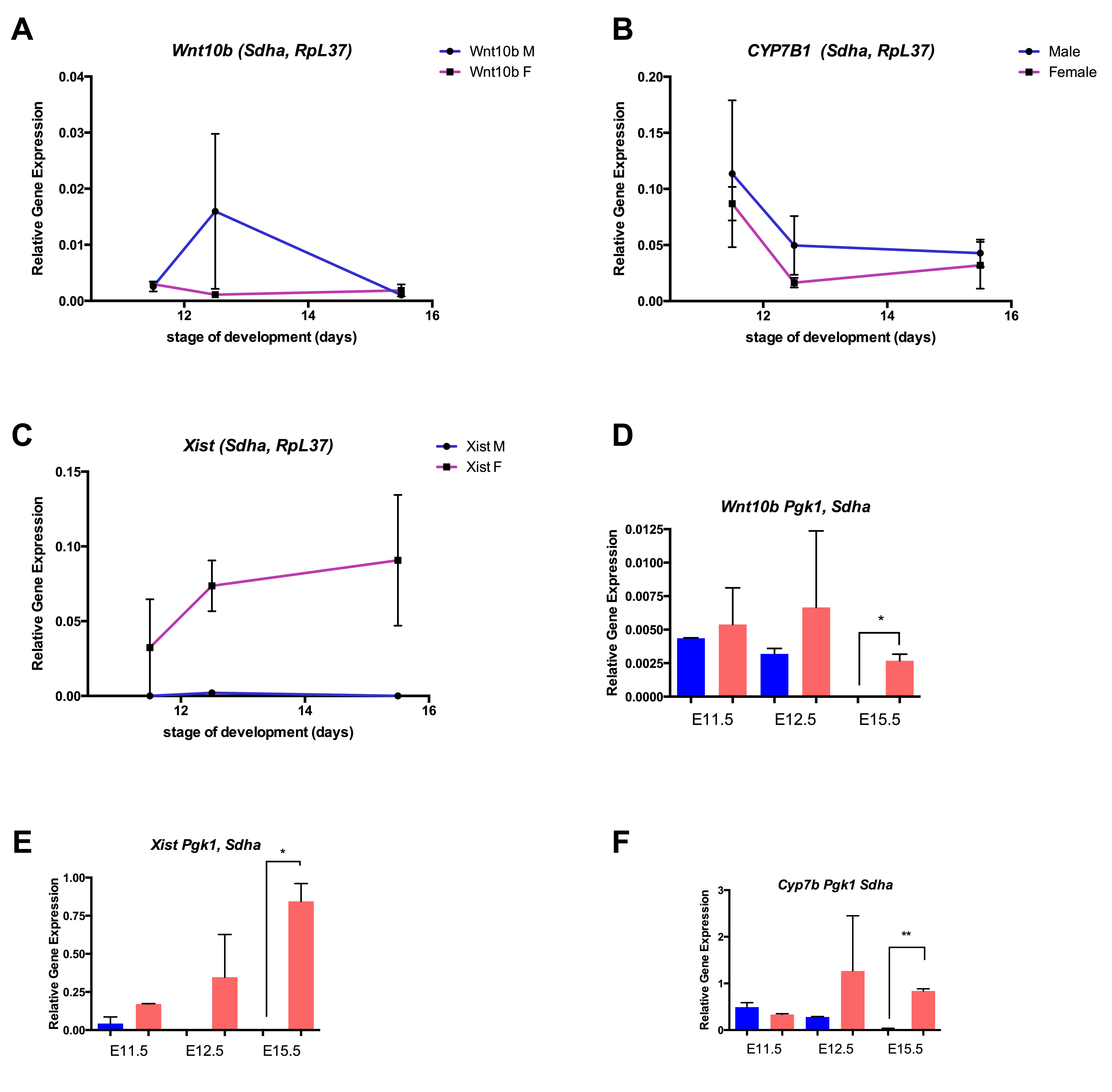
To normalize the expression of our three genes of interest, Sdha and Pgk1 oligonucleotide primers were used at E11.5, RpL38 and Sdha were used for analysis for E12.5. For analysis of E15.5, the reference genes Actb and RpL37 were used. Sex-dimorphic expression of Xist was detected at all three time points with increased expression in female brain compared to the males (Fig. 5A). CYP7B1 and Wnt10b also showed significant sex-dimorphic expression at E11.5 (P < 0.05), with higher expression observed in the male brain at this stage (Figs. 5C and 5E; 3.5-fold and 5-fold respectively). Additionally, following sex determination, Wnt10b and CYP7B1 expression was not significantly different between the sexes using these stage-specific reference genes (Figs.5C and 5E). In contrast, when pooling all data from sex and age together the top two ranked genes were RpL37 and Sdha. When using these two genes to normalize expression data there is a large variation in the expression of the three genes, especially at E12.5, between biological replicates (Figs. 5B, 5D and 5F). No significant differences were observed in gene expression between male and female samples, when using Pgk1 and Sdha as reference genes with the exception of Xist expression at E12.5 (Figs. 5B, 5D and 4F).
Figure 5. Sex specific gene expression using top ranked reference genes.

(A) Xist expression compared to the mean of the top two reference genes at each developmental time point. (B) Xist expression compared to Pgk1 and Sdha reference genes at all stages of development. (C) CYP7B1 expression compared to the mean of the top two reference genes at each developmental time stage. (D) CYP7B1 expression compared to Pgk1 and Sdha reference genes at all stages of development. (E) Wnt10b expression compared to the mean of top two reference genes at each developmental time point. (F) Wnt10b expression compared to Pgk1 and Sdha across all stages of development. Bar graphs are shown as the mean of three replicates with error bars as the mean standard error of the mean. ∗ = P < 0.05 ∗∗ = P < 0.001 ∗∗∗ = P < 0.0001 (Student’s unpaired T-test).
When studying gene expression changes over time, two slightly different sets of reference genes were ranked highest, depending upon the genotype of the sample (Table 2). We normalized gene expression at each time point for female samples (with Sdha and Gapdh) and for male samples (with Pgk1 and Sdha) to study how expression of these genes changes with respect to developmental age (Fig. 6). In comparison, when RpL37 (one of the poorer stable genes for female and male time points) and Sdha where used as reference genes for normalization of the all data points (top two genes when reference gene data was pooled Table 2), this introduced so much variation between samples, any analysis would be inconclusive (Fig. S1).
Figure 6. Expression of CYP7B1, Xist and Wnt10b across three stages of brain development.

(A) CYP7B1 mRNA expression in male (blue) and female (pink) brain tissue across developmental stages E11.5, E12.5 and E15.5 normalised to the top ranked reference genes for each sex over time. (B) Xist mRNA expression in male (blue) and female (pink) brain tissue across developmental time stages normalized to the top ranked reference genes for each sex at E11.5, E12.5 and E15.5. (C) Wnt10b mRNA expression of male (blue) and female (pink) brain tissue at developmental time stages E11.5, E12.5 and E15.5. Data is show as mean and standard error of the mean. ∗ = P < 0.05 ∗∗ = P < 0.001 Two-way ANOVA (Tukey’s multiple comparison test).
CYP7B1 mRNA expression declines between E11.5 and E12.5 in both sexes but its expression significantly increases again in the female only by E15.5 (Fig. 6A, P < 0.05). Xist transcript expression in the female brain also increased between E11.5 and E15.5 (following sex-determination window (Fig. 1)) (Fig. 6B; P < 0.001). Gene expression of the Wnt10b gene in the developing brain was low at all time-points and did not change significantly overtime in the female staged embryo samples (Fig. 6C). However, the higher expression observed in the male embryonic samples decreased also between E11.5 and E12.5, to similar expression levels found in XX samples (Fig. 6C).
Discussion
While studies using RTq-PCR have proven to be a powerful tool to study changes in gene expression, there must be a careful consideration of the type of reference genes to be used. As shown here, the reference genes used at particular developmental stages introduce much more variation into the data analysis and is problematic for drawing conclusions from RT-qPCR data. Therefore, it is essential to determine use the best reference gene pair for RT-qPCR analyses. For the comparison of sex-dimorphic gene expression at specific time points, Sdha and Pgk1 are recommended at E11.5. At E12.5, Actb and Sdha are most appropriate followed by Gapdh and Pgk1 are E15.5.
We determined the expression across time and between male and female sexes for three genes, previously identified as being expressed sex-dimorphically prior to sex-determination using the highest ranked references genes for each sex (Dewing et al., 2003). Here we identified the ideal reference genes when studying male gene expression across time, to be a combination of Actb and Sdha. In contrast, when studying gene expression across developmental stage with female samples, Sdha and Gapdh would be most appropriate. In comparison, if we used the Pgk1 gene (an X-linked gene) as the second reference for female samples, this introduced a lot of variation between biological replicates (n = 3, pooled samples from different litters) at E11.5 and E12.5. This suggests that expression of this commonly used reference gene was not stable in female embryonic brain tissues (Fig. S1) Pgk1 and Hprt are X-linked genes, subject to X inactivation during cell differentiation (Chaumeil et al., 2006; Heard, Clerc & Avner, 1997). This indicates that X-linked genes should be avoided when analyzing gene expression in XX tissues samples at early stages of brain development, particularly as our data for Xist (Fig. 6) indicates changes to the expression of X-linked genes during brain development.
CYP7B1 converts the dihydrotesosterone (DHT) metabolite 5α-androstane-3β, 17β-diol (3β-diol) to an inactive form and has been suggested to have a developmental role maintaining normal levels of estrogens and androgens in the mammalian brain (Rose et al., 1997). CYP7B1 mRNA expression has been previously detected in the hippocampus, testis and ovary (Wu et al., 1999). Knockout CYP7B1 mice have an enlarged brain (late fetal stages were examined) due to reduced apoptosis but overall brain weight normalizes after puberty for both sexes (Sugiyama et al., 2009). In female mice CYP7B1 knockout results in early onset puberty and ovarian failure (Omoto et al., 2005). A recent study found that male mutant have a reproductive behavioral defect, possibility due to altered olfactory cue sensing (Oyola et al., 2015). We found that expression of CYP7B1 is sex-dimorphic at E11.5, prior to any steroid hormone production by the developing gonads. By E15.5 the testis is producing significant levels of testosterone and DHT, expression levels of CYP7B1 remained low in the male but had increased in the female at E15.5 (Fig. 4C). Consistent with these results, previously it was found that the human CYP7B1 promoter is suppressed by DHT in human cell lines (Tang et al., 2006). In contrast, CYP7B1 expression increased with overexpression of estrogen receptors. While in females there is no prenatal estradiol production, 3β Adiol, the target of CYP7B1 activity, is produced by the immature ovary (Sugiyama et al., 2010). 3β Adiol binds to estrogen receptor β (ERβ), expression of this receptor begins at E12.5, peaks at E18.5 and is active in the absence of estrogen, possibility due to the presence of an alternative ligand such as 3β Adiol (Sugiyama et al., 2010). Together, this indicates that changes to the embryonic brain gene expression of CYP7B1 maybe regulated by androgen receptors and levels of steroid hormone metabolites during mouse development. This may produce the changes we observed in CYP7B1 expression across time in the female and males developing brain RNA samples (Figs. 4C and 5A).
The Wnt ligand, Wnt10b, was expressed at higher levels in male samples compared to female E10.5 head samples in a previous microarray study (Dewing et al., 2003). While Wnt10b knockout mice appear phenotypically normal, they exhibit a number of aging-related phenotype such as bone loss (Stevens et al., 2010). Little has been studied regarding the role of Wnt10b in early brain patterning, however research with the zebrafish model indicated that Wnt10b functions redundantly with Wnt1 in specification of the midbrain-hindbrain boundary (Lekven et al., 2003). Therefore it may play an early role in patterning of the midbrain-hindbrain boundary in mice, but any functional consequence for higher expression within the male developing brain at E10.5 (Dewing et al., 2003) and E11.5 (Fig. 4E) requires further investigation.
Xist is an essential long non-coding RNA that has a role in gene dosage compensation in XX embryos, by X chromosome in activation (XCI). During inactivation, the X chromosome that will be inactivated up-regulates Xist expression, whereas Xist gene expression from the active X chromosome is repressed (Galupa & Heard, 2015). We observed high levels of Xist expression in the female brain, compared to low or undetectable levels of Xist expression in the males between E11.5-15.5 (Fig. 4A). Regulation of Xist gene expression is complex, it is repressed by promoter bound CCCTC-binding factor (CTCF) and negatively regulated by an anti-sense RNA, Tsix. A second ncRNA transcript, Jpx, expressed from both X chromosomes, binds to CTCF and removes it from the Xist promoter region promoting expression of Xist (Sun et al., 2013). Xist gene expression (nor that of Jpx and Tsix) has not been examined in any detail overtime nor in later developing tissues. The function of Xist RNA in XCI is likely to be dosage sensitive (to Xist RNA levels) and tightly regulated. Mis-regulation (up- or down) will allow either more genes to escape X-inactivation or result in a reduction of expression of X-linked genes that are critical for brain development. Overexpression of Xist has been linked to female psychiatric conditions in humans (Ji et al., 2015).
This study has shown that the optimal reference gene(s) varies with the sex and stage of development. Therefore, when looking at gene expression either between sexes or across time for a particular sex, careful consideration should be given to which reference gene(s) are the most stable between the given samples and ideally use a combination of at least two to provide the most robust results for data analysis.
Supplemental Information
Normalized gene expression for CYP7B1 (A), Wnt10b (B) and Xist (C) using RpL37 and Sdha as combined reference genes. Normalized gene expression for Wnt10b (D), Xist (E) and CYP7B1 using Pgk1 and Sdha as combined reference genes.
{kind=link}
Name, accession number and oligonucleotide primer sequences used in this study.
Amplification efficiency for each oligonucleotide primer set.
Mean Ct values and corresponding standard deviation (SD) of each reference gene with respect to developmental stage/
Calculated NormFinder stability values, shaded values are top genes with lowest stability value.
Shaded values are top two genes with lowest std error values.
GeNorm values for each reference gene candidate.Mean M value shaded top two ranked genes.
Average standard deviation (of delta Ct values) for reference genes, sex and developmental stages.
Summary of rankings for comparing gene expression between male and female samples at each embryonic stage.
Acknowledgments
We would like to thank Simon Blanchoud for the very helpful comments on the manuscript drafts and Lisa Zondag for proofreading of the manuscript.
Funding Statement
The authors received funding from the Otago Medical Research Foundation. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Additional Information and Declarations
Competing Interests
The authors declare there are no competing interests.
Author Contributions
Tanya T. Cheung conceived and designed the experiments, performed the experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Mitchell K. Weston performed the experiments, contributed reagents/materials/analysis tools, reviewed drafts of the paper.
Megan J. Wilson conceived and designed the experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the paper, prepared figures and/or tables, reviewed drafts of the paper.
Animal Ethics
The following information was supplied relating to ethical approvals (i.e., approving body and any reference numbers):
University of Otago Animal Ethics Committee ET13/14.
Data Availability
References
- Abel, Drake & Golds (2010).Abel KM, Drake R, Goldstein JM. Sex differences in schizophrenia. International Review of Psychiatry. 2010;22(5):417–428. doi: 10.3109/09540261.2010.515205. [DOI] [PubMed] [Google Scholar]
- Andersen, Jensen & Orntoft (2004).Andersen CL, Jensen JL, Orntoft TF. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Research. 2004;64:5245–5250. doi: 10.1158/0008-5472.CAN-04-0496. [DOI] [PubMed] [Google Scholar]
- Arnold (2004).Arnold AP. Sex chromosomes and brain gender. Nature Reviews Neuroscience. 2004;5:701–708. doi: 10.1038/nrn1494. [DOI] [PubMed] [Google Scholar]
- Boda et al. (2009).Boda E, Pini A, Hoxha E, Parolisi R, Tempia F. Selection of reference genes for quantitative real-time RT-PCR studies in mouse brain. Journal of Molecular Neuroscience. 2009;37:238–253. doi: 10.1007/s12031-008-9128-9. [DOI] [PubMed] [Google Scholar]
- Chaumeil et al. (2006).Chaumeil J, Le Baccon P, Wutz A, Heard E. A novel role for Xist RNA in the formation of a repressive nuclear compartment into which genes are recruited when silenced. Genes and Development. 2006;20:2223–2237. doi: 10.1101/gad.380906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen et al. (1999).Chen RW, Saunders PA, Wei H, Li Z, Seth P, Chuang DM. Involvement of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and p53 in neuronal apoptosis: evidence that GAPDH is upregulated by p53. Journal of Neuroscience. 1999;19:9654–9662. doi: 10.1523/JNEUROSCI.19-21-09654.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dewing et al. (2003).Dewing P, Shi T, Horvath S, Vilain E. Sexually dimorphic gene expression in mouse brain precedes gonadal differentiation. Molecular Brain Research. 2003;118:82–90. doi: 10.1016/S0169-328X(03)00339-5. [DOI] [PubMed] [Google Scholar]
- Finlay & Darlington (1995).Finlay BL, Darlington RB. Linked regularities in the development and evolution of mammalian brains. Science. 1995;268(5217):1578–1584. doi: 10.1126/science.7777856. [DOI] [PubMed] [Google Scholar]
- Fombonne (2009).Fombonne E. Epidemiology of pervasive developmental disorders. Pediatric Research. 2009;65:591–598. doi: 10.1203/PDR.0b013e31819e7203. [DOI] [PubMed] [Google Scholar]
- Galupa & Heard (2015).Galupa R, Heard E. X-chromosome inactivation: new insights into cis and trans regulation. Current Opinion in Genetics & Development. 2015;31:57–66. doi: 10.1016/j.gde.2015.04.002. [DOI] [PubMed] [Google Scholar]
- Giedd et al. (1997).Giedd JN, Castellanos FX, Rajapakse JC, Vaituzis AC, Rapoport JL. Sexual dimorphism of the developing human brain. Progress in Neuro-Psychopharmacology and Biological Psychiatry. 1997;21(8):1185–1201. doi: 10.1016/S0278-5846(97)00158-9. [DOI] [PubMed] [Google Scholar]
- Goldstein et al. (2001).Goldstein JM, Seidman LJ, Horton NJ, Makris N, Kennedy DN, Caviness Jr VS, Faraone SV, Tsuang MT. Normal sexual dimorphism of the adult human brain assessed by in vivo magnetic resonance imaging. Cerebral Cortex. 2001;11:490–497. doi: 10.1093/cercor/11.6.490. [DOI] [PubMed] [Google Scholar]
- Gur et al. (1999).Gur RC, Turetsky BI, Matsui M, Yan M, Bilker W, Hughett P, Gur RE. Sex differences in brain gray and white matter in healthy young adults: correlations with cognitive performance. Journal of Neuroscience. 1999;19:4065–4072. doi: 10.1523/JNEUROSCI.19-10-04065.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heard, Clerc & Avner (1997).Heard E, Clerc P, Avner P. X-chromosome inactivation in mammals. Annual Review of Genetics. 1997;31:571–610. doi: 10.1146/annurev.genet.31.1.571. [DOI] [PubMed] [Google Scholar]
- Hellemans et al. (2007).Hellemans J, Mortier G, De Paepe A, Speleman F, Vandesompele J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biology. 2007;8 doi: 10.1186/gb-2007-8-2-r19. Article R19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hrdlickova, Toloue & Tian (2017).Hrdlickova R, Toloue M, Tian B. RNA-Seq methods for transcriptome analysis. Wiley Interdisciplinary Reviews: RNA. 2017;8(1) doi: 10.1002/wrna.1364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ji et al. (2015).Ji B, Higa KK, Kelsoe JR, Zhou X. Over-expression of XIST, the master gene for X chromosome inactivation, in females with major affective disorders. EBioMedicine. 2015;2:907–916. doi: 10.1016/j.ebiom.2015.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kouadjo et al. (2007).Kouadjo KE, Nishida Y, Cadrin-Girard JF, Yoshioka M, St-Amand J. Housekeeping and tissue-specific genes in mouse tissues. BMC Genomics. 2007;8:127. doi: 10.1186/1471-2164-8-127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lekven et al. (2003).Lekven AC, Buckles GR, Kostakis N, Moon RT. Wnt1 and wnt10b function redundantly at the zebrafish midbrain-hindbrain boundary. Developmental Biology. 2003;254:172–187. doi: 10.1016/S0012-1606(02)00044-1. [DOI] [PubMed] [Google Scholar]
- Lenroot et al. (2007).Lenroot RK, Gogtay N, Greenstein DK, Wells EM, Wallace GL, Clasen LS, Blumenthal JD, Lerch J, Zijdenbos AP, Evans AC, Thompson PM, Giedd JN. Sexual dimorphism of brain developmental trajectories during childhood and adolescence. NeuroImage. 2007;36:1065–1073. doi: 10.1016/j.neuroimage.2007.03.053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maekawa et al. (2014).Maekawa F, Tsukahara S, Kawashima T, Nohara K, Ohki-Hamazaki H. The mechanisms underlying sexual differentiation of behavior and physiology in mammals and birds: relative contributions of sex steroids and sex chromosomes. Frontiers in Neuroscience. 2014;8 doi: 10.3389/fnins.2014.00242. Article 242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mottron et al. (2015).Mottron L, Duret P, Mueller S, Moore RD, Forgeot d’Arc B, Jacquemont S, Xiong L. Sex differences in brain plasticity: a new hypothesis for sex ratio bias in autism. Molecular Autism. 2015;6 doi: 10.1186/s13229-015-0024-1. Article 33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ngun et al. (2011).Ngun TC, Ghahramani N, Sanchez FJ, Bocklandt S, Vilain E. The genetics of sex differences in brain and behavior. Frontiers in Neuroendocrinology. 2011;32:227–246. doi: 10.1016/j.yfrne.2010.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omoto et al. (2005).Omoto Y, Lathe R, Warner M, Gustafsson JA. Early onset of puberty and early ovarian failure in CYP7B1 knockout mice. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:2814–2819. doi: 10.1073/pnas.0500198102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oyola et al. (2015).Oyola MG, Zuloaga DG, Carbone D, Malysz AM, Acevedo-Rodriguez A, Handa RJ, Mani SK. CYP7B1 enzyme deletion impairs reproductive behaviors in male mice. Endocrinology. 2015;156:2150–2161. doi: 10.1210/en.2014-1786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrot-Sinal, Davis & McCarthy (2001).Perrot-Sinal TS, Davis AM, McCarthy MM. Developmental sex differences in glutamic acid decarboxylase (GAD(65)) and the housekeeping gene, GAPDH. Brain Research. 2001;922:201–208. doi: 10.1016/S0006-8993(01)03167-5. [DOI] [PubMed] [Google Scholar]
- Pfaffl et al. (2004).Pfaffl MW, Tichopad A, Prgomet C, Neuvians TP. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper–Excel-based tool using pair-wise correlations. Biotechnology Letters. 2004;26:509–515. doi: 10.1023/B:BILE.0000019559.84305.47. [DOI] [PubMed] [Google Scholar]
- Rose et al. (1997).Rose KA, Stapleton G, Dott K, Kieny MP, Best R, Schwarz M, Russell DW, Bjorkhem I, Seckl J, Lathe R. Cyp7b, a novel brain cytochrome P450, catalyzes the synthesis of neurosteroids 7alpha-hydroxy dehydroepiandrosterone and 7alpha-hydroxy pregnenolone. Proceedings of the National Academy of Sciences of the United States of America. 1997;94:4925–4930. doi: 10.1073/pnas.94.10.4925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saha et al. (2005).Saha S, Chant D, Welham J, McGrath J. A systematic review of the prevalence of schizophrenia. PLOS Medicine. 2005;2:e141. doi: 10.1371/journal.pmed.0020141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sawa et al. (1997).Sawa A, Khan AA, Hester LD, Snyder SH. Glyceraldehyde-3-phosphate dehydrogenase: nuclear translocation participates in neuronal and nonneuronal cell death. Proceedings of the National Academy of Sciences of the United States of America. 1997;94:11669–11674. doi: 10.1073/pnas.94.21.11669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silver et al. (2006).Silver N, Best S, Jiang J, Thein SL. Selection of housekeeping genes for gene expression studies in human reticulocytes using real-time PCR. BMC Molecular Biology. 2006;7:33. doi: 10.1186/1471-2199-7-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sotelo-Silveira et al. (2008).Sotelo-Silveira J, Crispino M, Puppo A, Sotelo JR, Koenig E. Myelinated axons contain beta-actin mRNA and ZBP-1 in periaxoplasmic ribosomal plaques and depend on cyclic AMP and F-actin integrity for in vitro translation. Journal of Neurochemistry. 2008;104:545–557. doi: 10.1111/j.1471-4159.2007.04999.x. [DOI] [PubMed] [Google Scholar]
- Stevens et al. (2010).Stevens JR, Miranda-Carboni GA, Singer MA, Brugger SM, Lyons KM, Lane TF. Wnt10b deficiency results in age-dependent loss of bone mass and progressive reduction of mesenchymal progenitor cells. Journal of Bone and Mineral Research. 2010;25:2138–2147. doi: 10.1002/jbmr.118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stiles & Jernigan (2010).Stiles J, Jernigan TL. The basics of brain development. Neuropsychology Review. 2010;20:327–348. doi: 10.1007/s11065-010-9148-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sugiyama et al. (2009).Sugiyama N, Andersson S, Lathe R, Fan X, Alonso-Magdalena P, Schwend T, Nalvarte I, Warner M, Gustafsson JA. Spatiotemporal dynamics of the expression of estrogen receptors in the postnatal mouse brain. Molecular Psychiatry. 2009;14:223–232. doi: 10.1038/mp.2008.118. 117. [DOI] [PubMed] [Google Scholar]
- Sugiyama et al. (2010).Sugiyama N, Barros RP, Warner M, Gustafsson JA. ERbeta: recent understanding of estrogen signaling. Trends in Endocrinology & Metabolism. 2010;21:545–552. doi: 10.1016/j.tem.2010.05.001. [DOI] [PubMed] [Google Scholar]
- Sun et al. (2013).Sun S, Del Rosario BC, Szanto A, Ogawa Y, Jeon Y, Lee JT. Jpx RNA activates Xist by evicting CTCF. Cell. 2013;153:1537–1551. doi: 10.1016/j.cell.2013.05.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sur & Rubenstein (2005).Sur M, Rubenstein JL. Patterning and plasticity of the cerebral cortex. Science. 2005;310(5749):805–810. doi: 10.1126/science.1112070. [DOI] [PubMed] [Google Scholar]
- Tang et al. (2006).Tang W, Eggertsen G, Chiang JY, Norlin M. Estrogen-mediated regulation of CYP7B1: a possible role for controlling DHEA levels in human tissues. Journal of Steroid Biochemistry and Molecular Biology. 2006;100:42–51. doi: 10.1016/j.jsbmb.2006.02.005. [DOI] [PubMed] [Google Scholar]
- Veazey & Golding (2011).Veazey KJ, Golding MC. Selection of stable reference genes for quantitative rt-PCR comparisons of mouse embryonic and extra-embryonic stem cells. PLOS ONE. 2011;6:e27592. doi: 10.1371/journal.pone.0027592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu et al. (1999).Wu Z, Martin KO, Javitt NB, Chiang JY. Structure and functions of human oxysterol 7alpha-hydroxylase cDNAs and gene CYP7B1. Journal of Lipid Research. 1999;40:2195–2203. [PubMed] [Google Scholar]
- Xie et al. (2012).Xie F, Xiao P, Chen D, Xu L, Zhang B. miRDeepFinder: a miRNA analysis tool for deep sequencing of plant small RNAs. Plant Molecular Biology. 2012;80:75. doi: 10.1007/s11103-012-9885-2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Normalized gene expression for CYP7B1 (A), Wnt10b (B) and Xist (C) using RpL37 and Sdha as combined reference genes. Normalized gene expression for Wnt10b (D), Xist (E) and CYP7B1 using Pgk1 and Sdha as combined reference genes.
Name, accession number and oligonucleotide primer sequences used in this study.
Amplification efficiency for each oligonucleotide primer set.
Mean Ct values and corresponding standard deviation (SD) of each reference gene with respect to developmental stage/
Calculated NormFinder stability values, shaded values are top genes with lowest stability value.
Shaded values are top two genes with lowest std error values.
GeNorm values for each reference gene candidate.Mean M value shaded top two ranked genes.
Average standard deviation (of delta Ct values) for reference genes, sex and developmental stages.
Summary of rankings for comparing gene expression between male and female samples at each embryonic stage.
Data Availability Statement
The following information was supplied regarding data availability: