

Abstract
Brucella abortus is a Gram-negative, facultative intracellular bacterium that causes brucellosis, a worldwide zoonotic disease leading to undulant fever in humans and abortion in cattle. The immune response against this bacterium relies on the recognition of microbial pathogen-associated molecular patterns, such as lipoproteins, lipopolysaccharides, and DNA; however, the immunostimulatory potential of B. abortus RNA remains to be elucidated. Here, we show that dendritic cells (DCs) produce significant amounts of IL-12, IL-6, and IP-10/CXCL10, when stimulated with purified B. abortus RNA. IL-12 secretion by DCs stimulated with RNA depends on TLR7 while IL-6 depends on TLR7 and partially on TLR3. Further, only TLR7 plays a role in IL-12 production induced by B. abortus infection. Moreover, cytokine production in DCs infected with B. abortus or stimulated with bacterial RNA was reduced upon pretreatment with MAPK/NF-κB inhibitors. By confocal microscopy, we demonstrated that TLR7 is colocalized with B. abortus in LAMP-1+ Brucella-containing vacuoles. Additionally, type I IFN expression and IP-10/CXCL10 secretion in DCs stimulated with bacterial RNA were dependent on TLR3 and TLR7. Our results suggest that TLR3 and TLR7 are not required to control Brucella infection in vivo, but they play an important role on sensing B. abortus RNA in vitro.
Keywords: B. abortus, innate immunity, toll-like receptors, RNA, cytokines, type I IFN
Introduction
The innate immunity is important to the initial recognition of pathogens, the development of a neutralizing response, and mobilization of the adaptive immunity (1). This defense relies on a limited set of evolutionary conserved pattern recognition receptors (PRRs) that recognize specific pathogen-associated molecular patterns (PAMPs) commonly present in microbes but not in mammals. Upon detection of PAMPs, some PRRs trigger an inflammatory response leading to the efficient destruction of the invading pathogens (2). Among these PRRs, the toll-like receptors (TLRs) family has gained more attention (1, 3).
Toll-like receptors are a well-conserved family of proteins, characterized by an extracellular leucine-rich repeat domain and an intracellular toll/IL-1 receptor-like domain (4). In mammals, the TLR family comprises more than 12 members (1). Although TLR1–TLR9 are conserved between humans and mice, TLR10 is not functional in mice because of a retrovirus insertion, and TLR11, TLR12, and TLR13 are lost in the human genome (5). Surface TLRs such as TLR1, TLR2, TLR4, TLR5, TLR6, and TLR11 mainly recognize microbial membrane components including lipids, lipoproteins, and flagella. TLR3, TLR7, TLR8, and TLR9 are preferentially expressed in intracellular vesicles of the endoplasmic reticulum (ER), endosomes, and lysosomes and recognize both microbial and viral nucleic acid motifs [double-stranded RNA, single-stranded RNA (ssRNA), and CpG-DNA are recognized by TLR3, TLR7/8, and TLR9, respectively] (6). TLRs responsible for sensing nucleic acids are expressed in various cell types, including dendritic cells (DCs), macrophages, and B cells (7). Microbial nucleic acids are an important class of PAMPs and self/non-self recognition is based on different parameters, such as their sequence, structure, molecular modifications, and localization (mislocalized self nucleic acids such as extranuclear DNA or extracellular RNA are recognized as damage-associated molecular patterns) (8). There are recent reports on the properties of nucleic acid recognition for viral RNA and bacterial DNA (9, 10); however, little is known about the immunogenicity of prokaryotic RNA. Moreover, the role of TLRs acting as bacterial RNA sensors is controversial. Initially, it was reported that bacteria from group B Streptococcus (GBS) potently induced type I IFN in conventional DCs (cDCs) by a mechanism that required TLR7, MyD88, and the transcription factor IRF-1. These molecules colocalized with bacterial products in degradative vacuoles bearing lysosomal markers, linking lysosomal recognition of bacterial RNA with a robust IFN response (11). Further, Eberle et al. demonstrated that bacterial RNA is a potent trigger for type I IFN secretion in human peripheral blood mononuclear cells. The same study reported that murine plasmacytoid DCs from TLR7-deficient mice were unable to trigger an immune response against bacterial RNA (12). In addition, Love et al. demonstrated that the induction of IFN-α and IFN-λ1 (a type III IFN) by Borrelia burgdorferi RNA or live spirochetes requires TLR7-dependent signaling and an enhanced IRF7 expression (13). The TLR-mediated sensing to bacterial RNA depends on the cellular type studied and the chosen model of infection and some studies reported cases in which TLRs are not essential. For instance, Deshmukh et al. reported that the recognition of ssRNA from GBS and other Gram-positive bacteria by macrophages and monocytes depends on the adaptors MyD88 and UNC93B, but not TLRs (14). It was also demonstrated by Gratz et al. that in cDCs and macrophages stimulated with Streptococcus pyogenes RNA, type I IFN was induced in the absence of TLR3, TLR7, and TLR9 (15).
Brucella abortus is a facultative, intracellular Gram-negative bacterium that causes brucellosis, an important zoonotic infection that causes reproductive disease in domestic animals and chronic debilitating disease in humans (16, 17). Brucella infects and multiplies in various cell types, including macrophages, DCs, and non-phagocytic cells (18). The first line of defense against brucellosis includes phagocytosis by professional phagocytes (neutrophils, macrophages, and DCs), and natural killer (NK) cells, recognition of PAMPs by PRRs (e.g., TLRs), secretion of cytokines and chemokines, and activation of the complement system (19, 20). Signaling B. abortus infection via TLRs has been investigated by several groups including ours. These studies have highlighted the involvement of TLR2, TLR4, TLR6, and TLR9, as well as a dependence of MyD88 on immune responses against this bacterium (21–26). To evade immune surveillance, Brucella is devoid of conspicuous molecular determinants such as pili, fimbriae, and capsules, and it possess non-canonical surface molecules, such as lipopolysaccharide (LPS), ornithine-containing lipids, lipoproteins, and flagella, structures that lack marked PAMP activities and hence are very weak inducers of innate immunity (27). However, there are still few reports in the literature related to the host innate immune response against Brucella-derived nucleic acids (24, 28–30). Moreover, the role of Brucella RNA during bacterial infections remains undeciphered. In this study, we focused on the stimulatory activity of B. abortus RNA, as well as the role of TLR3 and TLR7 in cell signaling pathways and host protection against infection.
Materials and Methods
Bacteria
Brucella abortus smooth virulent strain S2308 was obtained from our laboratory collection. All work with Brucella, including animal experiments, was conducted in the biosafety cabinet or a primary containment device within a dedicated laboratory. Appropriate laboratory coats, gloves, and protective eyewear were provided to ensure the safety of personnel. All personnel working in the biosafety laboratory were trained and approved for entry by an individual knowledgeable in the biosafety practices. Frozen stocks were prepared with bacteria previously grown in Brucella broth liquid medium (BB) (Becton Dickinson, Franklin Lakes, NJ, USA) at 37°C at 180 rpm. The bacterial culture was centrifuged after 72 h of growth and the pellet was resuspended in saline solution plus 25% glycerol. Aliquots of these cultures were serially diluted, plated on BB agar 1.5%, and after incubation for 72 h at 37°C, bacterial numbers were determined by counting CFU.
Mice
TLR3, TLR7, or MyD88 KO mice were kindly provided by Dr. Shizuo Akira, Osaka University, Japan. Wild-type (WT) strain C57BL/6 mice were obtained from the Federal University of Minas Gerais (UFMG, Belo Horizonte, Brazil). Genetically deficient and control mice were maintained at our facilities and used at 6–8 weeks of age. All animal experiments were preapproved by the Institutional Animal Care and Use Committee of the UFMG (CETEA no. 104/2011). Mice were housed in filter-top cages and provided with sterile water and food ad libitum.
Generation of Bone Marrow-Derived DCs
Bone marrow cells were obtained from femur and tibiae of KO and WT mice, and they were differentiated into DCs as previously described (26, 31, 32). Briefly, bone marrow cells were cultured in RPMI medium 1640 (Thermo Fisher Scientific, Waltham, MA, USA) containing 10% fetal bovine serum (FBS), 100 U/mL penicillin, 100 µg/mL streptomycin, and 20 ng/mL murine recombinant granulocyte-macrophage colony-stimulating factor (GM-CSF). Petri dishes containing 1 × 107 cells were incubated at 37°C in a 5% CO2 atmosphere. On day 3 of incubation, a further 5 mL of fresh complete medium containing GM-CSF was added, and on days 5 and 7, the medium was replaced with fresh supplemented medium containing GM-CSF. On day 10, non-adherent cells (immature DCs) were harvested, seeded in 24-well plates (5 × 105 cells/well), and then incubated at 37°C in a 5% CO2 atmosphere until use.
Isolation of B. abortus Total RNA from Bacterial Cell Culture
Bacteria were cultured for 3 days at 37°C in 10 mL BB liquid medium, and the bacterial suspension was pelleted into 10 eppendorfs. The supernatants were removed and each pellet was resuspended in 1 mL TRIzol® (Thermo Fisher Scientific, Waltham, MA, USA). The procedure of RNA isolation was performed following instructions of the manufacturer. Briefly, each TRIzol-bacterial suspension was vortexed, then 0.2 mL chloroform was added and the mixture was centrifuged at 11,000 × g during 10 min at 4°C. The aqueous phase was transferred to another eppendorf and 0.5 mL ice-cold isopropanol was added. The mixture was incubated at room temperature for 10 min and then centrifuged at 11,000 × g during 15 min at 4°C. The supernatant was discarded and the precipitate was washed with 1 mL ice-cold ethanol 75%, through centrifugation at 7,500 × g during 10 min at 4°C. The pellet was air-dried and resuspended using 50 µL nuclease-free water. Total RNA was quantified using the Nanodrop 2000 spectrophotometer (Nanodrop 2000; Thermo Scientific), treated with DNase I and/or RNases A/T1 (Thermo Scientific) and stored at −80°C until use.
In Vitro Stimulation of DCs
Stimulation of the DCs was performed by adding RPMI plus 10% FBS (0.5 mL/well) containing B. abortus [multiplicity of infection (MOI) 100:1], Escherichia coli LPS (1 µg/mL; Sigma-Aldrich), or B. abortus total RNA (2 µg/mL) complexed with DOTAP Liposomal Transfection Reagent (Sigma-Aldrich, St. Louis, MO, USA). Briefly, DOTAP was mixed with bacterial RNA (5:1 ratio) in 100 µL/well serum-free RPMI and incubated for 20 min at room temperature. Then, complexes were added to the cells in the presence of 12.5% FBS and cell cultures were incubated for 24 h at 37°C in a 5% CO2 atmosphere. Culture supernatants were collected after 24 h of stimulation and kept at −80°C until use. To the remaining cells, it was added 0.350 mL/well of TRIzol® to further RNA isolation.
RNA Isolation from DCs and Quantitative Real-time PCR
Reverse transcription of 1 µg from total RNA was performed using Illustra™ Ready-To-Go RT-PCR Beads (GE Healthcare, Little Chalfont, UK), following instructions of the manufacturer. Quantitative real-time PCR was conducted in a final volume of 10 µL containing the following: SYBR® Green PCR Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), oligo-dT cDNA as the PCR template, and 5 µM of primers. The PCR reaction was performed with StepOne Plus Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA), using the following cycling parameters: 60°C for 10 min, 95°C for 10 min, 40 cycles of 95°C for 15 s and 60°C for 1 min, and a dissociation stage of 95°C for 15 s, 60°C for 1 min, 95°C for 15 s, and 60°C for 15 s. Primers were used to amplify a specific 100–120 bp fragment corresponding to specific gene targets as follows: IFN-β, forward (5′-AGCTCCAAGAAAGGACGAACAT-3′); IFN-β, reverse (5′-GCCCTGTAGGTGAGGTTGATCT-3′); β-actin, forward (5′-AGGTGTGCACCTTTTATTGGTCTCAA-3′); β-actin, reverse (5′-TGTATGAAGGTTTGGTCTCCCT-3′). All data are presented as relative expression units after normalization to the β-actin gene. PCR measurements were conducted in triplicate.
Inhibition of MAPK/NF-κB Signaling Pathways in DCs
DCs were pretreated with vehicle (0.1% DMSO) or selective inhibitors for ERK1/2 (U0126; 10, 50, and 100 µM), JNK (SP600125; 1, 5, and 10 µM), p38 (SB203580; 5, 25, and 50 µM), and NF-κB (BAY1170-82; 10, 100, and 500 µM) (all from Cell Signaling Technology, Danvers, MA, USA) for 45 min. Subsequently, cells were stimulated with B. abortus S2308 strain (MOI 100:1) or with bacterial RNA (2 µg/mL) for 24 h, and harvested supernatants were used to measure cytokine production by ELISA.
Measurement of IL-12, IL-6, IP-10/CXCL10, and TNF-α by ELISA
Collected supernatants of stimulated DCs were frozen until they were assayed for determination of cytokine concentrations using Mouse DuoSet ELISA (R&D Systems), according to the manufacturer’s specifications.
Confocal Microscopy
Dendritic cells were differentiated on 12 mm glass coverslips in 24-well plates, as described before. Infection with the B. abortus was performed at a MOI of 100:1. After 24 h of infection, cells were washed twice with PBS and fixed in 4% paraformaldehyde, pH 7.4, at 37°C for 15 min. Once fixed, cells were incubated with the primary antibody rat anti-lysosomal-associated membrane protein 1 (LAMP-1) diluted 1:200 (Abcam, Cambridge, UK) and rabbit anti-TLR7 diluted 1:100 (Abcam, Cambridge, UK) in PBS 0.1% saponin overnight. Then, cells were washed three times with PBS 0.1% saponin and were incubated for 1.5 h at room temperature with an anti-rabbit secondary antibody conjugated to Alexa 546 (Invitrogen, Carlsbad, CA, USA) and with an anti-rat secondary antibody conjugated to Alexa 488 (Abcam, Cambridge, UK), both diluted 1:1,000. The cells were washed twice with PBS 0.1% saponin and once with PBS then mounted with the mounting medium ProLong Gold with DAPI (Invitrogen, Carlsbad, CA, USA). Stained cells were examined by confocal microscopy (Nikon C2 confocal microscope) and image analysis was performed using ImageJ software version 1.47n.
Enumeration of B. abortus from Spleens
To count residual B. abortus CFU in the spleens of C57BL/6 WT, TLR3 KO, TLR7 KO, and MyD88 KO mice, five mice from each group were examined at 1 and 3 weeks postinfection. Spleens were homogenized with 10 mL of saline solution. To enumerate viable bacteria, homogenized spleen samples were serially diluted 10-fold with saline solution and plated on BB agar (Becton Dickinson, Franklin Lakes, NJ, USA). CFUs were counted after 3 days of incubation at 37°C.
Statistical Analysis
All experiments were repeated at least three times with similar results and figures show data from one representative experiment. Graphs and data analysis were done using GraphPad Prism 5 (GraphPad Software), using one-way ANOVA, two-way ANOVA (Bonferroni post hoc test), or Student’s t-test (Tukey’s post hoc test).
Results
Lack of TLR7 Affects Proinflammatory Cytokine Production by DCs Activated with B. abortus or Its RNA
To determine whether TLR3 and TLR7 are required to induce proinflammatory cytokine production in DCs stimulated with B. abortus, cells from C57BL/6, TLR3 KO, TLR7 KO, and MyD88 KO were cultured with B. abortus (MOI 100:1) and after 24 h the supernatants were tested for production of IL-12, IL-6, and TNF-α by ELISA. IL-12 was partially reduced in TLR7 KO cells but not in TLR3 KO DCs (Figure 1A). Additionally, secretion of IL-6 and TNF-α was unaffected by the absence of TLR3 and TLR7 (Figures 1C,E). Furthermore, IL-12, IL-6, and TNF-α production were totally abrogated in MyD88 KO cells.
Figure 1.

Secretion of proinflammatory cytokines in dendritic cells (DCs) after stimulation with Brucella abortus or bacterial RNA. DCs were stimulated with B. abortus strain S2308 (MOI 100:1) or lipopolysaccharide (LPS) (1 µg/mL) (A,C,E), or bacterial RNA (2 μg/mL) (B,D). Supernatants were harvested after 24 h after stimulation and cytokine secretion was determined by ELISA. Significant differences in relation to wild-type mice are denoted (for ***p < 0.001, two-way ANOVA test).
Next, we assessed the potential of B. abortus RNA as TLR3 and/or TLR7 agonist to induce proinflammatory cytokines. DCs from C57BL/6, TLR3 KO, TLR7 KO, and MyD88 KO were stimulated with purified B. abortus RNA and after 24 h the supernatants were tested for production of IL-12, IL-6, and TNF-α. B. abortus RNA was able to induce the production of IL-12 and IL-6 in WT cells (Figures 1B,D) but not TNF-α (data not shown). Further, IL-12 and IL-6 secretion were completely abrogated in TLR7 and MyD88 KO cells when compared to the WT DCs. However, IL-6 secretion was only partially dependent on TLR3 KO cells (Figure 1D). These results suggest that, upon B. abortus infection, only the lack of TLR7 partially affected IL-12 production in DCs. On the other hand, TLR3 and TLR7 are required to induce a cytokine response in DCs stimulated with Brucella-purified RNA. Additionally, our results also indicated that bacterial RNA treatment with RNases A/T1 abolished IL-12 and IL-6 in DCs, indicating that no other possible contaminant would be responsible for the cytokine production observed (Figures 1B,D).
ERK1/2, JNK, and NF-κB but Not p38 Signaling Are Involved in Cytokine Production in DCs Stimulated with B. abortus or Bacterial RNA
Since B. abortus or its purified RNA induce the production of proinflammatory cytokines in DCs, we evaluated the role of MAPK components and the NF-κB transcription factor during cell activation. DCs from WT mice were treated with selective inhibitors against ERK1/2 (U0126), p38 (SB203580), JNK (SP600125), and NF-κB (BAY1170-82) before stimulation with B. abortus or bacterial RNA. Harvested supernatants were used to assess IL-12, IL-6, and TNF-α production (Figure 2). Treatment of DCs with the ERK1/2 inhibitor before infection with B. abortus reduced the production of all three cytokines, whereas the cytokine production was not significantly altered by inhibition of p38. Moreover, inhibition of JNK in DCs stimulated with B. abortus resulted in a decrease in IL-12 and IL-6 but not TNF-α. It was also observed that IL-12, IL-6, and TNF-α production are reduced in infected cells treated with a selective inhibitor of NF-κB (Figures 2A,C,E).
Figure 2.

MAPK/NF-κB signaling pathways involved on cytokine production in dendritic cells (DCs) after stimulation with Brucella abortus or bacterial RNA. DCs from C57BL/6 mice were pretreated for 45 min with increasing concentrations (in micromolars) of U0126, SB203580, SP600125, and BAY11-7082 (ERK1/2, p38, JNK, and NF-κB inhibitors, respectively) or vehicle, as indicated, and then stimulated with B. abortus (MOI 100:1) (A,C,E) or bacterial RNA (2 µg/mL) (B,D). Supernatants were harvested after 24 h after stimulation and cytokine secretion was determined by ELISA. Abbreviations: NI, non-infected; C, Control, DOTAP only;, RNA, DOTAP + RNA. Significant differences in relation to vehicle treatment are denoted (for *p < 0.05, **p < 0.01, and ***p < 0.001, two-way ANOVA test).
We also assessed whether the pretreatment with MAPK/NF-κB inhibitors would affect cytokine production in DCs stimulated with B. abortus RNA. Both ERK1/2 and JNK inhibitors, as well as the NF-κB inhibitor, were able to reduce IL-6 production in DCs stimulated with bacterial RNA (Figure 2D). As observed for live bacteria, p38 inhibitor did not reduce the production of any studied cytokine in the supernatant of these cells. Besides, IL-12 production was diminished only in supernatants from DCs pretreated with JNK and NF-κB inhibitors (Figure 2B), which suggests that DCs infected with live bacteria may be exposed to other bacterial agonists that are recognized via ERK besides to JNK and NF-κB. Importantly, the reduction in cytokine production mediated by U0126, SP600125, and BAY1170-82 is not a consequence of toxicity caused by the inhibitors since treatment induces slight levels of cell death and only in high concentrations (Figure S1 in Supplementary Material). Once again, TNF-α production was not detected in DCs stimulated with B. abortus RNA, pretreated or not with the inhibitors previously mentioned (data not shown). All together, these results indicate that MAPK and NF-κB activation participates in proinflammatory cytokine production in DCs stimulated with B. abortus or its RNA.
TLR7 and LAMP-1 Are Colocalized in B. abortus-Infected DCs
Results shown in Figure 1 suggest that IL-12 production in DCs stimulated with B. abortus or its purified RNA depends on the signaling via TLR7. Upon entry into mammalian cells, B. abortus resides within a membrane-bound, ER-derived compartment, the Brucella-containing vacuole (BCV). Before reaching the ER, the bacteria ensure its intracellular survival by inhibiting fusion of the intermediate BCV with late endosomes and lysosomes, in which we observe the presence of LAMP-1 (33). Therefore, to gain insight into the role of TLR7 in the response to B. abortus infection, we performed confocal microscopy in infected DCs, labeling cells with anti-TLR7 and anti-LAMP-1 antibody, and visualizing by confocal microscopy. Our results suggest that upon infection of WT DCs with B. abortus, TLR7 traffics to LAMP-1+ endosomes, spreading throughout the cells (Figure 3). Indeed, colocalization of TLR7 and Brucella in LAMP-1+ compartments could facilitate sensing of RNA released from bacteria and subsequent cytokine production. As expected, there was no TLR7 staining in TLR7 KO DCs. Further, TLR7 KO DCs showed phenotypically distinct clustered, membrane-bound LAMP-1+ BCV-like structures, when compared to WT DCs. All together, these results suggest that TLR7 is involved in a B. abortus-induced signaling pathway, leading to a proinflammatory cytokine production.
Figure 3.
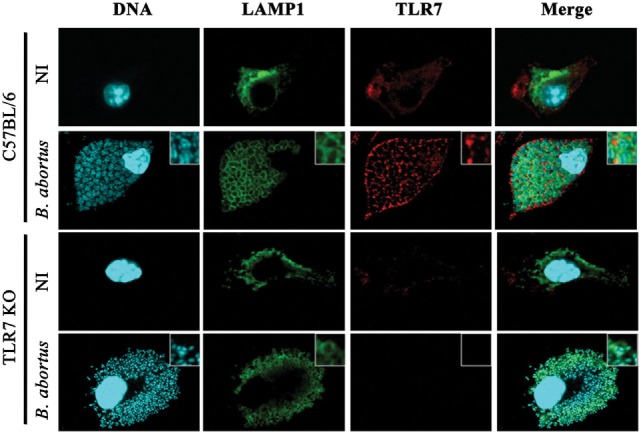
Dendritic cells (DCs) activated by Brucella abortus promote TLR7 colocalization with lysosomal-associated membrane protein 1 (LAMP-1)-positive compartments. DCs were stimulated with B. abortus (MOI 100:1) for 24 h. Subsequently, macrophages were fixed and stained to detect host cell and bacterial DNA (blue), TLR7 (red), and LAMP-1 (green) by confocal microscopy. The colocalization between TLR7 and LAMP-1 is highlighted in inserts. NI, non-infected.
TLR3 and TLR7 Are Required for Type I IFN Expression and IP-10/CXCL10 Secretion in DCs Stimulated with B. abortus RNA
It is known that upon TLR3 and TLR7 recognition of nucleic acids released into endosomes after virus or bacteria internalization or lysis results in type I IFN production, particularly IFN-β (34, 35). Therefore, we decided to investigate the levels of IFN-β expression and IP-10/CXCL10 production, a surrogate marker of type I IFN (36, 37), in TLR3 and TLR7 KO DCs activated with B. abortus or its purified RNA. We have observed no significant reduction on IFN-β expression and IP-10/CXCL10 production in TLR3 and TLR7 KO cells infected with B. abortus when compared to the WT (Figures 4A,C). Bacterial RNA was able to induce the expression of IFN-β and IP-10/CXCL10 production in WT DCs; however, the levels of these cytokines were diminished in TLR3 KO and TLR7 KO cells (Figures 4B,D). IFN-β expression and IP-10/CXCL10 production were only partially reduced in MyD88 KO cells because the IFN-responsive gene expression can also be activated via TLR3–TRIF pathway (38, 39). However, when MyD88 KO cells were activated with bacterial RNA a dramatic reduction on cytokine levels were observed. The previous treatment of RNA with RNases abolished the differences between KO and WT cells.
Figure 4.

Type I IFN expression and IP-10/CXCL10 secretion in dendritic cells (DCs) stimulated with Brucella abortus or bacterial RNA. DCs were stimulated with B. abortus strain S2308 (MOI 100:1) (A,C) or bacterial RNA (2 µg/mL) (B,D) for 24 h. Total RNA was isolated from these cells, first-strand cDNA synthesis was performed as indicated in Section “Materials and Methods” and subjected to qPCR analysis. Supernatants were analyzed by ELISA. Significant differences related to wild-type mice are denoted (***p < 0.001, two-way ANOVA test).
TLR3 and TLR7 Are Not Required to Control B. abortus Infection In Vivo
Results obtained from in vitro studies indicate that TLR3 and TLR7 play a role in cytokine production and type I IFN expression upon stimulation with B. abortus RNA but are dispensable for the induction of an immune response against live bacteria. Therefore, to test the role of these receptors in host defense against Brucella infection in vivo, C57BL/6 WT, TLR3 KO, TLR7 KO, and MyD88 KO mice to test the role of these receptors in host defense against Brucella infection in vivo. Mice were infected intraperitoneally with 106 CFU B. abortus and the number of bacteria in the spleen was quantified. TLR3 and TLR7 KO mice were able to effectively control Brucella infection as the WT animals at 1 and 3 weeks postinfection (Figure 5). MyD88 KO were used as a control group and showed the highest CFU numbers in the spleen compared to other strains tested here, indicating their susceptibility to infection.
Figure 5.

The control of Brucella abortus infection in vivo in mice deficient for TLR3, TLR7, and for the adaptor molecule MyD88. Residual B. abortus CFU in the spleen of wild-type (WT) and KO mice were determined after 1 (A) and 3 (B) weeks postinfection. Significant differences in relation to WT mice are denoted (***p < 0.001, Student’s t-test).
Discussion
Here, we provided evidence that B. abortus RNA function as an immunostimulatory PAMP that induces the production of proinflammatory cytokines in bone marrow-derived DCs. DCs stimulated with bacterial RNA produced significant amounts of IL-12 and IL-6 (Figures 1B,D), and this production is decreased when cells are pretreated with MAPK/NF-κB inhibitors (Figures 2B,D). Moreover, B. abortus RNA induces IFN-β expression (Figure 4B) and IP-10/CXCL10 production (Figure 4D). On the other hand, although we observed a robust secretion of TNF-α in DCs stimulated with B. abortus (Figure 1E), there was no detectable TNF-α production in RNA-stimulated cells (data not shown). Results reported by Eberle et al. indicated that TNF-α secretion in murine DCs induced by bacterial RNA is independent of endosomal maturation and consequently independent of endosomal nucleic acid-sensing TLRs (12). Furthermore, the transfection of DCs with RNA complexed to DOTAP, a cationic lipid that mimics the chemical and physical attributes of biological lipids (40), allows direct delivery of RNA to endosomes, avoiding a possible nucleic acid sensing by cytosolic sensors, for example, members of retinoic acid-inducible gene I (RIG-I)-like receptors family: RIG-I, melanoma differentiation factor 5 (MDA5), and laboratory of genetics and physiology 2 (LGP2) (38).
Our data reported here also reveal that B. abortus RNA is recognized in a TLR3- and TLR7-dependent manner by DCs. However, the involvement of these two receptors is only noticed upon stimulation with purified RNA complexed to DOTAP and not upon infection with live bacteria (Figures 1 and 4). Alternatively, the immune responses against B. abortus infection could be triggered by major agonists such as DNA, lipoproteins, or LPS, and probably such stimuli are more efficient in activating proinflammatory cytokine secretion and type I IFN expression than bacterial RNA. A recent review published by our group describes B. abortus DNA as a major agonist to activate innate responses against this bacterium, and this activation is mediated by the endosomal TLR9 as well as cytosolic molecules such as AIM2 and STING (30). Moreover, endosomal TLRs may interact with each other, resulting in a unidirectional and hierarchical inhibition. In a work performed by Wang and colleagues in 2006, it was demonstrated a cooperative interaction among TLR7, TLR8, and TLR9, in HEK293 cells transfected with human or murine TLRs in a paired combination (39). In this study, the results indicated that TLR8 inhibits TLR7 and TLR9, and TLR9 inhibits TLR7 but TLR7 does not inhibit TLR8 neither TLR9. Furthermore, in mice, it is possible that the inflammatory response triggered upon TLR7 activation may be modulated by TLR8 in all DC subsets. We cannot rule out the possibility that B. abortus RNA is sensed by other PRRs in a MyD88-dependent pathway, since both IFN-β expression and IP-10/CXCL10 secretion are abrogated in MyD88 KO DCs (Figures 4B,D). TLR13, a PRR involved in sensing a specific sequence from bacterial 23S rRNA, is present in mice but not in humans (41, 42). It is possible that TLR13 may play a role in B. abortus RNA sensing, but this hypothesis needs further investigation. The assumptions aforementioned could also explain the results obtained in Figure 5, in which we compared bacterial clearance in vivo in WT, TLR3 KO, and TLR7 KO mice and we observed no significant differences among these studied groups. Similar findings were previously described by Oliveira and coworkers, where the macrophages derived from NOD1, NOD2, and RIP2 KO mice showed an altered IL-12 and TNF-α secretion but no differences were observed among these groups in vivo (43).
Brucella abortus has developed sophisticated molecular strategies to ensure its survival inside host cells by forming BCVs, which traffics from the endocytic vesicle to the ER, where the bacterium proliferates (44). BCV maturation is evidenced by the exclusion of LAMP-1, possibly by recycling LAMP-1+ membranes while interacting with the ER (45). Results shown in the Figure 3 demonstrate that, after 24 h of infection, B. abortus colocalized with TLR7 in BCVs that retain LAMP-1 labeling. However, it is important to highlight that literature on BCV maturation is based on studies using macrophages, and BCVs from infected DCs could present alterations on their membrane constitution. In addition, it is possible that in infected DCs ER-localized Brucella could hold membrane rearrangements dependent upon a subset of autophagy-associated proteins, producing vacuolar LAMP-1+ compartments that are phenotypically distinct from ER-derived BCVs, so called autophagic Brucella-containing vacuoles (44).
In summary, our findings shown here suggest that B. abortus RNA is an important immunostimulatory nucleic acid, and bacterial RNA-initiated TLR3 and TLR7 signaling contributes to cytokine response and type I IFN expression in murine DCs. To the best of our knowledge, this study reports for the first time the involvement of RNA sensing TLRs on innate immune responses against Brucella infection and highlights the importance of nucleic acid sensing during bacterial diseases.
Author Contributions
PC, MG, and SO designed the project and experiments. PC, MG, EG, and GG carried out most of the experiments. PC and SO wrote the manuscript. PC carried out statistical analysis and prepared figures. SO submitted this paper. All the authors reviewed the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank Dr. Gustavo B. Menezes for the use of his Nikon confocal microscope.
Funding
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Fundação de Amparo à Pesquisa do estado de Minas Gerais (FAPEMIG), CAPES/PVE, CAPES/PNPD, CNPq/CT-Biotec, CNPq/CBAB, and National Institutes of Health R01 AI116453.
Supplementary Material
The Supplementary Material for this article can be found online at http://journal.frontiersin.org/article/10.3389/fimmu.2017.00028/full#supplementary-material.
References
- 1.Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell (2006) 124:783–801. 10.1016/j.cell.2006.02.015 [DOI] [PubMed] [Google Scholar]
- 2.Medzhitov R, Janeway C., Jr Innate immunity. N Engl J Med (2000) 343:338–44. 10.1056/NEJM200008033430506 [DOI] [PubMed] [Google Scholar]
- 3.Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev (2009) 22:240–73. 10.1128/CMR.00046-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Medzhitov R, Preston-Hurlburt P, Janeway CA, Jr. A human homologue of the Drosophila toll protein signals activation of adaptive immunity. Nature (1997) 388:394–7. 10.1038/41131 [DOI] [PubMed] [Google Scholar]
- 5.Kawai T, Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int Immunol (2009) 21:317–37. 10.1093/intimm/dxp017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mifsud E, Tan A, Jackson D. TLR agonists as modulators of the innate immune response and their potential as agents against infectious disease. Front Immunol (2014) 5:79. 10.3389/fimmu.2014.00079 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Iwasaki A, Medzhitov R. Toll-like receptor control of the adaptive immune responses. Nat Immunol (2004) 5:987–95. 10.1038/ni1112 [DOI] [PubMed] [Google Scholar]
- 8.Desmet CJ, Ishii KJ. Nucleic acid sensing at the interface between innate and adaptive immunity in vaccination. Nat Rev Immunol (2012) 12:479–91. 10.1038/nri3247 [DOI] [PubMed] [Google Scholar]
- 9.Barbalat R, Ewald SE, Mouchess ML, Barton GM. Nucleic acid recognition by the innate immune system. Annu Rev Immunol (2011) 29:185–214. 10.1146/annurev-immunol-031210-101340 [DOI] [PubMed] [Google Scholar]
- 10.Wu J, Chen ZJ. Innate immune sensing and signaling of cytosolic nucleic acids. Annu Rev Immunol (2014) 32:461–88. 10.1146/annurev-immunol-032713-120156 [DOI] [PubMed] [Google Scholar]
- 11.Mancuso G, Gambuzza M, Midiri A, Biondo C, Papasergi S, Akira S, et al. Bacterial recognition by TLR7 in the lysosomes of conventional dendritic cells. Nat Immunol (2009) 10:587–94. 10.1038/ni.1733 [DOI] [PubMed] [Google Scholar]
- 12.Eberle F, Sirin M, Binder M, Dalpke AH. Bacterial RNA is recognized by different sets of immunoreceptors. Eur J Immunol (2009) 39:2537–47. 10.1002/eji.200838978 [DOI] [PubMed] [Google Scholar]
- 13.Love AC, Schwartz I, Petzke MM. Borrelia burgdorferi RNA induces type I and III interferons via toll-like receptor 7 and contributes to production of NF-kappaB-dependent cytokines. Infect Immun (2014) 82:2405–16. 10.1128/IAI.01617-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Deshmukh SD, Kremer B, Freudenberg M, Bauer S, Golenbock DT, Henneke P. Macrophages recognize streptococci through bacterial single-stranded RNA. EMBO Rep (2011) 12:71–6. 10.1038/embor.2010.189 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gratz N, Hartweger H, Matt U, Kratochvill F, Janos M, Sigel S, et al. Type I interferon production induced by Streptococcus pyogenes-derived nucleic acids is required for host protection. PLoS Pathog (2011) 7:e1001345. 10.1371/journal.ppat.1001345 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Corbel MJ. Brucellosis: an overview. Emerg Infect Dis (1997) 3:213–21. 10.3201/eid0302.970219 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Byndloss MX, Tsolis RM. Brucella spp. virulence factors and immunity. Annu Rev Anim Biosci (2016) 4:111–27. 10.1146/annurev-animal-021815-111326 [DOI] [PubMed] [Google Scholar]
- 18.Archambaud C, Salcedo SP, Lelouard H, Devilard E, de Bovis B, Van Rooijen N, et al. Contrasting roles of macrophages and dendritic cells in controlling initial pulmonary Brucella infection. Eur J Immunol (2010) 40:3458–71. 10.1002/eji.201040497 [DOI] [PubMed] [Google Scholar]
- 19.Diacovich L, Gorvel JP. Bacterial manipulation of innate immunity to promote infection. Nat Rev Microbiol (2010) 8:117–28. 10.1038/nrmicro2295 [DOI] [PubMed] [Google Scholar]
- 20.Oliveira SC, de Oliveira FS, Macedo GC, de Almeida LA, Carvalho NB. The role of innate immune receptors in the control of Brucella abortus infection: toll-like receptors and beyond. Microbes Infect (2008) 10:1005–9. 10.1016/j.micinf.2008.07.005 [DOI] [PubMed] [Google Scholar]
- 21.Delpino MV, Barrionuevo P, Macedo GC, Oliveira SC, Genaro SD, Scian R, et al. Macrophage-elicited osteoclastogenesis in response to Brucella abortus infection requires TLR2/MyD88-dependent TNF-alpha production. J Leukoc Biol (2012) 91:285–98. 10.1189/jlb.04111185 [DOI] [PubMed] [Google Scholar]
- 22.Duenas AI, Orduna A, Crespo MS, Garcia-Rodriguez C. Interaction of endotoxins with toll-like receptor 4 correlates with their endotoxic potential and may explain the proinflammatory effect of Brucella spp. LPS. Int Immunol (2004) 16:1467–75. 10.1093/intimm/dxh148 [DOI] [PubMed] [Google Scholar]
- 23.de Almeida LA, Macedo GC, Marinho FA, Gomes MT, Corsetti PP, Silva AM, et al. Toll-like receptor 6 plays an important role in host innate resistance to Brucella abortus infection in mice. Infect Immun (2013) 81:1654–62. 10.1128/IAI.01356-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Huang LY, Ishii KJ, Akira S, Aliberti J, Golding B. Th1-like cytokine induction by heat-killed Brucella abortus is dependent on triggering of TLR9. J Immunol (2005) 175:3964–70. 10.4049/jimmunol.175.6.3964 [DOI] [PubMed] [Google Scholar]
- 25.Oliveira SC, de Almeida LA, Carvalho NB, Oliveira FS, Lacerda TL. Update on the role of innate immune receptors during Brucella abortus infection. Vet Immunol Immunopathol (2012) 148:129–35. 10.1016/j.vetimm.2011.05.036 [DOI] [PubMed] [Google Scholar]
- 26.Macedo GC, Magnani DM, Carvalho NB, Bruna-Romero O, Gazzinelli RT, Oliveira SC. Central role of MyD88-dependent dendritic cell maturation and proinflammatory cytokine production to control Brucella abortus infection. J Immunol (2008) 180:1080–7. 10.4049/jimmunol.180.2.1080 [DOI] [PubMed] [Google Scholar]
- 27.Martirosyan A, Gorvel JP. Brucella evasion of adaptive immunity. Future Microbiol (2013) 8:147–54. 10.2217/fmb.12.140 [DOI] [PubMed] [Google Scholar]
- 28.Gomes MT, Campos PC, Oliveira FS, Corsetti PP, Bortoluci KR, Cunha LD, et al. Critical role of ASC inflammasomes and bacterial type IV secretion system in caspase-1 activation and host innate resistance to Brucella abortus infection. J Immunol (2013) 190:3629–38. 10.4049/jimmunol.1202817 [DOI] [PubMed] [Google Scholar]
- 29.Gomes MT, Campos PC, Pereira GS, Bartholomeu DC, Splitter G, Oliveira SC. TLR9 is required for MAPK/NF-kappaB activation but does not cooperate with TLR2 or TLR6 to induce host resistance to Brucella abortus. J Leukoc Biol (2016). 10.1189/jlb.4A0815-346R [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Campos PC, Gomes MT, Guimaraes G, Costa Franco MM, Marim FM, Oliveira SC. Brucella abortus DNA is a major bacterial agonist to activate the host innate immune system. Microbes Infect (2014) 16:979–84. 10.1016/j.micinf.2014.08.010 [DOI] [PubMed] [Google Scholar]
- 31.Trant CG, Lacerda TL, Carvalho NB, Azevedo V, Rosinha GM, Salcedo SP, et al. The Brucella abortus phosphoglycerate kinase mutant is highly attenuated and induces protection superior to that of vaccine strain 19 in immunocompromised and immunocompetent mice. Infect Immun (2010) 78:2283–91. 10.1128/IAI.01433-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chow A, Toomre D, Garrett W, Mellman I. Dendritic cell maturation triggers retrograde MHC class II transport from lysosomes to the plasma membrane. Nature (2002) 418:988–94. 10.1038/nature01006 [DOI] [PubMed] [Google Scholar]
- 33.Starr T, Ng TW, Wehrly TD, Knodler LA, Celli J. Brucella intracellular replication requires trafficking through the late endosomal/lysosomal compartment. Traffic (2008) 9:678–94. 10.1111/j.1600-0854.2008.00718.x [DOI] [PubMed] [Google Scholar]
- 34.Kawai T, Akira S. TLR signaling. Cell Death Differ (2006) 13:816–25. 10.1038/sj.cdd.4401850 [DOI] [PubMed] [Google Scholar]
- 35.Perry AK, Chen G, Zheng D, Tang H, Cheng G. The host type I interferon response to viral and bacterial infections. Cell Res (2005) 15:407–22. 10.1038/sj.cr.7290309 [DOI] [PubMed] [Google Scholar]
- 36.Qian C, An H, Yu Y, Liu S, Cao X. TLR agonists induce regulatory dendritic cells to recruit Th1 cells via preferential IP-10 secretion and inhibit Th1 proliferation. Blood (2007) 109:3308–15. 10.1182/blood-2006-08-040337 [DOI] [PubMed] [Google Scholar]
- 37.Gautier G, Humbert M, Deauvieau F, Scuiller M, Hiscott J, Bates EE, et al. A type I interferon autocrine-paracrine loop is involved in toll-like receptor-induced interleukin-12p70 secretion by dendritic cells. J Exp Med (2005) 201:1435–46. 10.1084/jem.20041964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Vabret N, Blander JM. Sensing microbial RNA in the cytosol. Front Immunol (2013) 4:468. 10.3389/fimmu.2013.00468 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Wang J, Shao Y, Bennett TA, Shankar RA, Wightman PD, Reddy LG. The functional effects of physical interactions among toll-like receptors 7, 8, and 9. J Biol Chem (2006) 281:37427–34. 10.1074/jbc.M605311200 [DOI] [PubMed] [Google Scholar]
- 40.Leventis R, Silvius JR. Interactions of mammalian cells with lipid dispersions containing novel metabolizable cationic amphiphiles. Biochim Biophys Acta (1990) 1023:124–32. 10.1016/0005-2736(90)90017-I [DOI] [PubMed] [Google Scholar]
- 41.Hidmark A, von Saint Paul A, Dalpke AH. Cutting edge: TLR13 is a receptor for bacterial RNA. J Immunol (2012) 189:2717–21. 10.4049/jimmunol.1200898 [DOI] [PubMed] [Google Scholar]
- 42.Oldenburg M, Kruger A, Ferstl R, Kaufmann A, Nees G, Sigmund A, et al. TLR13 recognizes bacterial 23S rRNA devoid of erythromycin resistance-forming modification. Science (2012) 337:1111–5. 10.1126/science.1220363 [DOI] [PubMed] [Google Scholar]
- 43.Oliveira FS, Carvalho NB, Zamboni DS, Oliveira SC. Nucleotide-binding oligomerization domain-1 and -2 play no role in controlling Brucella abortus infection in mice. Clin Dev Immunol (2012) 2012:861426. 10.1155/2012/861426 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Starr T, Child R, Wehrly TD, Hansen B, Hwang S, Lopez-Otin C, et al. Selective subversion of autophagy complexes facilitates completion of the Brucella intracellular cycle. Cell Host Microbe (2012) 11:33–45. 10.1016/j.chom.2011.12.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Celli J, de Chastellier C, Franchini DM, Pizarro-Cerda J, Moreno E, Gorvel JP. Brucella evades macrophage killing via VirB-dependent sustained interactions with the endoplasmic reticulum. J Exp Med (2003) 198:545–56. 10.1084/jem.20030088 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.