

ABSTRACT
Wounding is the first event that occurs in plant regeneration. However, wound signaling in plant regeneration is barely understood. Using a simple system of de novo root organogenesis from Arabidopsis thaliana leaf explants, we analyzed the genes downstream of wound signaling. Leaf explants may produce at least two kinds of wound signals to trigger short-term and long-term wound signaling. Short-term wound signaling is primarily involved in controlling auxin behavior and the fate transition of regeneration-competent cells, while long-term wound signaling mainly modulates the cellular environment at the wound site and maintains the auxin level in regeneration-competent cells. YUCCA (YUC) genes, which are involved in auxin biogenesis, are targets of short-term wound signaling in mesophyll cells and of long-term wound signaling in regeneration-competent cells. The expression patterns of YUCs provide important information about the molecular basis of wound signaling in plant regeneration.
KEYWORDS: Arabidopsis thaliana, De novo root organogenesis, plant regeneration, wound signals, wound signaling
Plant cells are highly plastic and have powerful regenerative abilities.1-4 Wounding is the first event in plant regeneration.1, 5 Studies on wounding have suggested several candidates for wound signals, such as electrical pulses, hydraulic pressure, Ca2+, reactive oxygen species, the oligopeptide systemin, oligosaccharides, jasmonic acid, salicylic acid, ethylene, abscisic acid, and changes in various metabolic processes.6,7 Many studies have tried to clarify the effects of wounding on regeneration by analysis of the genes downstream of wound signaling.8-27 However, our knowledge about whether and how the above physical and chemical signals serve as wound signal(s) in plant regeneration is still limited.
To study the role of wound signaling in de novo root organogenesis, we cultured leaf explants on B5 medium to regenerate adventitious roots.28 Wounding has complex biological effects, and may have multiple roles in de novo root organogenesis from leaf explants. Our previous studies revealed that at least two pathways are triggered by wounding of leaf explants: short-term and long-term wound-signaling pathways.
Short-term wound signaling is required for auxin production and cell fate transition (indicated by the red flow path in Fig. 1A).10,29 This pathway comprises at least three stages of signal delivery (Fig. 1A). In stage I, wounding first triggers short-term wound signaling that lasts from seconds to hours. This wound signal spreads very rapidly from the wound site to mesophyll cells and activates YUCCA1 (YUC1) and YUC4 expression in mesophyll cells within 4 hours. Stage II involves auxin behavior. Auxin begins to be produced by YUCs in mesophyll cells within 4 hours and then polar-transported to regeneration-competent cells in the vasculature near the wound site at around 12 hours after wounding. In stage III, the accumulation of auxin in regeneration-competent cells activates expression of WUSCHEL RELATED HOMEOBOX11 (WOX11) and WOX12, which start the cell fate transition to form root founder cells at around 1 to 2 days d after wounding.
Figure 1.
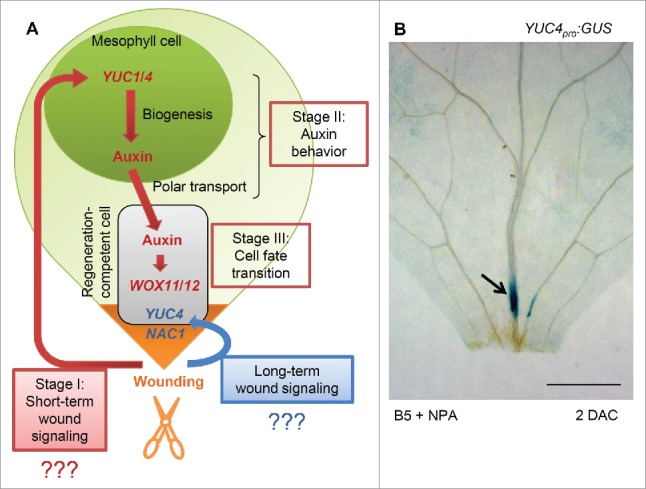
Wound signaling in de novo root organogenesis. (A) Model of short-term (red flow path) and long-term (blue flow path) wound signaling-mediated pathways in de novo root organogenesis. Orange region indicates wound site. (B) YUC4 expression in leaf explant cultured on B5 medium containing 1 μM NPA. Arrow shows GUS signal in regeneration-competent cells at 2 DAC. Leaf explant was cultured in dark conditions,28 and GUS staining was performed as described previously.29,30 Scale bar, 500 μm in (B).
The long-term wound-signaling pathway functions for a relatively long time, for around 2 days after wounding (indicated by the blue flow path in Fig. 1A).11 One event downstream of long-term wound signaling is the activation of a group of NAC (NAM, ATAF1,2, and CUC2) transcription factor genes including NAC1 (Fig. 1A).11 NAC1 is expressed in many cells at the wound site around 1 to 2 days after detachment of the leaf explant. However, NAC1 does not affect the fate transition of regeneration-competent cells because WOX11 and WOX12 expression are normally activated when NAC1 function is blocked. The role of NAC1 might be to control the cellular environment, including cell wall metabolism. On the other hand, the long-term wound-signaling pathway also activates YUC4 expression in regeneration-competent cells near the wound site at around 2 days after wounding (Fig. 1A),10 and this might contribute to maintaining a high auxin level in regeneration-competent cells. Here, we analyzed YUC4pro:GUS10 in leaf explants cultured on B5 medium containing naphthylphthalamic acid (NPA, a polar auxin transport inhibitor) to test whether YUC4 is a direct target of long-term wound signaling or is activated by auxin accumulation in regeneration-competent cells. Treatment with NPA can prevent auxin accumulation in regeneration-competent cells in leaf explants.29 We observed strong β-glucuronidase (GUS) signals in regeneration-competent cells near the wound in the leaf explant cultured on medium containing NPA at 2 days after culture (DAC) (Fig. 1B), suggesting that YUC4 might be a direct target of long-term wound signaling in regeneration-competent cells. Currently, it is unclear whether the activation of NAC1 and YUC4 is controlled dependently or independently by long-term wound signaling.
The short-term and long-term wound-signaling pathways act differently in leaf explants. First, the short-term wound signal is produced at the wound site and quickly moves to the mesophyll cells to activate YUC1 and YUC4 expression.10 Therefore, this signal might be a moving signal that can spread from cell to cell. Long-term wound signaling functions at the wound site,11 suggesting that it is a local signal. Second, short-term wound signaling lasts for seconds to hours,10 while long-term signaling lasts for at least 2 days.11 This suggests that the short-term wound signal has a short lifetime, while the long-term signal has a longer lifetime or is continuously produced in response to wounding. Third, short-term wound signaling seems to function in both the detached explant and in the wounded leaf residue on the source plant, because YUC1 and YUC4 expression are quickly activated in leaf explants as well as at the wound site on the source plant.10 However, long-term wound signaling activates NAC1 expression primarily at the wound site on the detached explant, and not in the leaf residue on the source plant.11 This indicates that long-term wound signaling has a role in distinguishing different types of wounding. Overall, it seems that wounding produces complex molecules that serve as different types of wound signals in de novo root organogenesis from leaf explants.
Although many of the downstream effects of wounding have been identified, our knowledge about wound signals and wound signaling at the molecular level is still very limited. Recent studies have suggested that jasmonic acid, ethylene, and electrical pulses have effects on tissue repair,9,15,17,25 raising the possibility that they may be candidates for wound signals to trigger regeneration. In our regeneration system of de novo root organogenesis, YUC1 and YUC4 serve as targets of short-term wound signaling in mesophyll cells, and NAC1 and YUC4 are targets of long-term wound signaling at the wound site. It will be interesting to test the upstream molecular events of YUCs and NAC1 to study the molecular basis of wounding during de novo root organogenesis.18
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding
This work was supported by grants from National Basic Research Program of China (973 Program, 2014CB943500/2012CB910503), the National Natural Science Foundation of China (91419302/31422005) and Youth Innovation Promotion Association CAS (2014241).
References
- 1.Xu L, Huang H. Genetic and epigenetic controls of plant regeneration. Curr Top Dev Biol 2014; 108:1-33; PMID:24512704; http://dx.doi.org/ 10.1016/B978-0-12-391498-9.00009-7 [DOI] [PubMed] [Google Scholar]
- 2.Ikeuchi M, Ogawa Y, Iwase A, Sugimoto K. Plant regeneration: cellular origins and molecular mechanisms. Development 2016; 143:1442-51; PMID:27143753; http://dx.doi.org/ 10.1242/dev.134668 [DOI] [PubMed] [Google Scholar]
- 3.Pulianmackal AJ, Kareem AV, Durgaprasad K, Trivedi ZB, Prasad K. Competence and regulatory interactions during regeneration in plants. Front Plant Sci 2014; 5:142; PMID:24782880; http://dx.doi.org/ 10.3389/fpls.2014.00142 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Su YH, Zhang XS. The hormonal control of regeneration in plants. Curr Top Dev Biol 2014; 108:35-69; PMID:24512705; http://dx.doi.org/ 10.1016/B978-0-12-391498-9.00010-3 [DOI] [PubMed] [Google Scholar]
- 5.Lup SD, Tian X, Xu J, Perez-Perez JM. Wound signaling of regenerative cell reprogramming. Plant Sci 2016; 250:178-87; PMID:27457994; http://dx.doi.org/ 10.1016/j.plantsci.2016.06.012 [DOI] [PubMed] [Google Scholar]
- 6.Leon J, Rojo E, Sanchez-Serrano JJ. Wound signalling in plants. J Exp Bot 2001; 52:1-9; PMID:11181708; http://dx.doi.org/ 10.1093/jexbot/52.354.1 [DOI] [PubMed] [Google Scholar]
- 7.Maffei ME, Mithofer A, Boland W. Before gene expression: early events in plant-insect interaction. Trends Plant Sci 2007; 12:310-6; PMID:17596996; http://dx.doi.org/ 10.1016/j.tplants.2007.06.001 [DOI] [PubMed] [Google Scholar]
- 8.Iwase A, Mitsuda N, Koyama T, Hiratsu K, Kojima M, Arai T, Inoue Y, Seki M, Sakakibara H, Sugimoto K, et al.. The AP2/ERF transcription factor WIND1 controls cell dedifferentiation in Arabidopsis. Curr Biol 2011; 21:508-14; PMID:21396822; http://dx.doi.org/http://dx.doi.org/ 10.1016/j.cub.2011.02.020 [DOI] [PubMed] [Google Scholar]
- 9.Melnyk CW, Schuster C, Leyser O, Meyerowitz EM. A Developmental Framework for Graft Formation and Vascular Reconnection in Arabidopsis thaliana. Curr Biol 2015; 25:1306-18; PMID:25891401; http://dx.doi.org/ 10.1016/j.cub.2015.03.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Chen L, Tong J, Xiao L, Ruan Y, Liu J, Zeng M, Huang H, Wang JW, Xu L.. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis. J Exp Bot 2016; 67:4273-84; PMID:27255928; http://dx.doi.org/ 10.1093/jxb/erw213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chen X, Cheng J, Chen L, Zhang G, Huang H, Zhang Y, Xu L.. Auxin-independent NAC pathway acts in response to explant-specific wounding and promotes root tip emergence during de novo root organogenesis in Arabidopsis. Plant Physiol 2016; 170:2136-45; PMID:26850273; http://dx.doi.org/27212234 10.1104/pp.15.01733 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Efroni I, Mello A, Nawy T, Ip PL, Rahni R, DelRose N, Powers A, Satija R, Birnbaum KD. Root Regeneration Triggers an Embryo-like Sequence Guided by Hormonal Interactions. Cell 2016; 165:1721-33; PMID:27212234; http://dx.doi.org/ 10.1016/j.cell.2016.04.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Xu J, Hofhuis H, Heidstra R, Sauer M, Friml J, Scheres B. A molecular framework for plant regeneration. Science 2006; 311:385-8; PMID:16424342; http://dx.doi.org/ 10.1126/science.1121790 [DOI] [PubMed] [Google Scholar]
- 14.Sena G, Wang X, Liu HY, Hofhuis H, Birnbaum KD. Organ regeneration does not require a functional stem cell niche in plants. Nature 2009; 457:1150-3; PMID:19182776; http://dx.doi.org/ 10.1038/nature07597 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Asahina M, Azuma K, Pitaksaringkarn W, Yamazaki T, Mitsuda N, Ohme-Takagi M, Yamaguchi S, Kamiya Y, Okada K, Nishimura T, et al.. Spatially selective hormonal control of RAP2.6L and ANAC071 transcription factors involved in tissue reunion in Arabidopsis. Proc Natl Acad Sci U S A 2011; 108:16128-32; PMID:21911380; http://dx.doi.org/ 10.1073/pnas.1110443108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ahkami AH, Lischewski S, Haensch KT, Porfirova S, Hofmann J, Rolletschek H, Melzer M, Franken P, Hause B, Druege U, et al.. Molecular physiology of adventitious root formation in Petunia hybrida cuttings: involvement of wound response and primary metabolism. New Phytol 2009; 181:613-25; PMID:19076299; http://dx.doi.org/ 10.1111/j.1469-8137.2008.02704.x [DOI] [PubMed] [Google Scholar]
- 17.Kral N, Ougolnikova AH, Sena G. Externally imposed electric field enhances plant root tip regeneration. Regeneration 2016; 3(3):156-67; PMID:27606066; http://dx.doi.org/ 10.1002/reg2.59 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sang YL, Cheng ZJ, Zhang XS. Endogenous auxin biosynthesis and de novo root organogenesis. J Exp Bot 2016; 67:4011-3; PMID:27402616; http://dx.doi.org/ 10.1093/jxb/erw250 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Delessert C, Wilson IW, Van Der Straeten D, Dennis ES, Dolferus R. Spatial and temporal analysis of the local response to wounding in Arabidopsis leaves. Plant Mol Biol 2004; 55:165-81; PMID:15604673; http://dx.doi.org/ 10.1007/s11103-004-0112-7 [DOI] [PubMed] [Google Scholar]
- 20.Sukumar P, Maloney GS, Muday GK. Localized induction of the ATP-binding cassette B19 auxin transporter enhances adventitious root formation in Arabidopsis. Plant Physiol 2013; 162:1392-405; PMID:23677937; http://dx.doi.org/ 10.1104/pp.113.217174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chupeau MC, Granier F, Pichon O, Renou JP, Gaudin V, Chupeau Y. Characterization of the early events leading to totipotency in an Arabidopsis protoplast liquid culture by temporal transcript profiling. Plant Cell 2013; 25:2444-63; PMID:23903317; http://dx.doi.org/ 10.1105/tpc.113.109538 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Pitaksaringkarn W, Ishiguro S, Asahina M, Satoh S. ARF6 and ARF8 contribute to tissue reunion in incised Arabidopsis inflorescence stems. Plant Biotechnol J 2014; 34:49-53; http://dx.doi.org/ 10.5511/plantbiotechnology.13.1028b [DOI] [Google Scholar]
- 23.Asahina M, Satoh S. Molecular and physiological mechanisms regulating tissue reunion in incised plant tissues. J Plant Res 2015; 128:381-8; PMID:25736731; http://dx.doi.org/ 10.1007/s10265-015-0705-z [DOI] [PubMed] [Google Scholar]
- 24.Grafi G, Barak S. Stress induces cell dedifferentiation in plants. Biochim Biophys Acta 2015; 1849:378-84; PMID:25086338; http://dx.doi.org/ 10.1016/j.bbagrm.2014.07.015 [DOI] [PubMed] [Google Scholar]
- 25.Reid JB, Ross JJ. Regulation of tissue repair in plants. Proc Natl Acad Sci U S A 2011; 108:17241-2; PMID:21960442; http://dx.doi.org/ 10.1073/pnas.1114432108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Heyman J, Cools T, Vandenbussche F, Heyndrickx KS, Van Leene J, Vercauteren I, Vanderauwera S, Vandepoele K, De Jaeger G, Van Der Straeten D, et al.. ERF115 controls root quiescent center cell division and stem cell replenishment. Science 2013; 342:860-3; PMID:24158907; http://dx.doi.org/ 10.1126/science.1240667 [DOI] [PubMed] [Google Scholar]
- 27.Ge Y, Liu J, Zeng M, He J, Qin P, Huang H, Xu L.. Identification of WOX family genes in Selaginella kraussiana for studies on stem cells and regeneration in lycophytes. Front Plant Sci 2016; 7:93; PMID:26904063; http://dx.doi.org/ 10.3389/fpls.2016.00093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chen X, Qu Y, Sheng L, Liu J, Huang H, Xu L. A simple method suitable to study de novo root organogenesis. Front Plant Sci 2014; 5:208; PMID:24860589; http://dx.doi.org/ 10.3389/fpls.2014.00208 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Liu J, Sheng L, Xu Y, Li J, Yang Z, Huang H, Xu L. WOX11 and 12 are involved in the first-step cell fate transition during de novo root organogenesis in Arabidopsis. Plant Cell 2014; 26:1081-93; PMID:24642937; http://dx.doi.org/ 10.1105/tpc.114.122887 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.He C, Chen X, Huang H, Xu L. Reprogramming of H3K27me3 is critical for acquisition of pluripotency from cultured Arabidopsis tissues. PLoS Genet 2012; 8:e1002911; PMID:22927830; http://dx.doi.org/ 10.1371/journal.pgen.1002911 [DOI] [PMC free article] [PubMed] [Google Scholar]