

Abstract
The present study examined the impacts of major depressive disorder (MDD) on visual and prefrontal cortical activity as well as their connectivity during visual working memory updating and related them to the core clinical features of the disorder. Impairment in working memory updating is typically associated with the retention of irrelevant negative information which can lead to persistent depressive mood and abnormal affect. However, performance deficits have been observed in MDD on tasks involving little or no demand on emotion processing, suggesting dysfunctions may also occur at the more basic level of information processing. Yet, it is unclear how various regions in the visual working memory circuit contribute to behavioral changes in MDD. We acquired functional magnetic resonance imaging data from 18 unmedicated participants with MDD and 21 age-matched healthy controls (CTL) while they performed a visual delayed recognition task with neutral faces and scenes as task stimuli. Selective working memory updating was manipulated by inserting a cue in the delay period to indicate which one or both of the two memorized stimuli (a face and a scene) would remain relevant for the recognition test. Our results revealed several key findings. Relative to the CTL group, the MDD group showed weaker postcue activations in visual association areas during selective maintenance of face and scene working memory. Across the MDD subjects, greater rumination and depressive symptoms were associated with more persistent activation and connectivity related to no-longer-relevant task information. Classification of postcue spatial activation patterns of the scene-related areas was also less consistent in the MDD subjects compared to the healthy controls. Such abnormalities appeared to result from a lack of updating effects in postcue functional connectivity between prefrontal and scene-related areas in the MDD group. In sum, disrupted working memory updating in MDD was revealed by alterations in activity patterns of the visual association areas, their connectivity with the prefrontal cortex, and their relationship with core clinical characteristics. These results highlight the role of information updating deficits in the cognitive control and symptomatology of depression.
Keywords: Depression, Working memory updating, Prefrontal cortex, Visual cortex, Rumination, fMRI, Functional connectivity
Highlights
-
•
Unmedicated individuals with major depressive disorder showed several forms of deficits during visual working memory updating.
-
•
Impaired visual working memory updating performance.
-
•
Diminished category-specific response patterns in visual association areas, particularly those involved in scene processing.
-
•
Loss of frontal functional connectivity with the scene-selective visual region during updating of scene information.
-
•
Rumination is associated with heightened activity and functional connectivity for obsolete information in working memory.
1. Introduction
Cognitive models of major depressive disorder (MDD) propose that cognitive inflexibility including repetitive focus on negative thoughts, perseveration of non-optimal problem-solving strategies, and failure to switch to new relevant information may underline the development and maintenance of the disorder (Gotlib and Joormann, 2010, Nolen-Hoeksema et al., 2008). In behavioral research, cognitive inflexibility in MDD is often demonstrated by poor performance on working memory tasks with an updating component (Gohier et al., 2009, Joormann et al., 2011, Meiran et al., 2011). More specifically, working memory updating deficits have been shown to involve reduced ability to process and maintain task-relevant information (Gruber et al., 2011, Pelosi et al., 2000), discard obsolete material (Berman et al., 2011, Cooney et al., 2010), and inhibit distractors (Davis and Nolen-Hoeksema, 2000, Desseilles et al., 2009). Several studies have associated these deficits with depressive symptom severity (Demeyer et al., 2012, Harvey et al., 2004) and rumination (Demeyer et al., 2012, Watkins and Brown, 2002, Whitmer and Banich, 2007), suggesting a potentially close relationship between information updating and core clinical aspects of depression. Such updating difficulties in MDD are typically interpreted as the result of stimulus-processing biases driven by the emotional content of task material. Yet, impairments have been reported in selective attention, maintenance of task-relevant information, and filtering of distractors in tasks involving simple visual stimuli with minimal emotional influence (e.g., letters, colors, shapes) (Desseilles et al., 2009, Garrett et al., 2011, MacQueen et al., 2000, Silton et al., 2011). Thus, working memory updating dysfunctions can occur at the more basic visual information processing level, potentially underscoring the cognitive inflexibility and emotional dysregulations observed in MDD. Despite such evidence, the extent to which working memory updating deficits impact information processing and contribute to the clinical features of MDD is ambiguous due to inconsistent behavioral findings and limited neuroimaging studies of visual cortical functions.
Previous studies have found working memory updating impairments in MDD and yet evidence was mixed as to whether these impairments are part of a general working memory deficit or a specific dysfunction in depression. Studies which used both updating (n-back) and maintenance (forward digit and visuospatial span) tasks showed depressed subjects performed significantly worse than healthy controls only when updating was required (Harvey et al., 2004, Landro et al., 2001). These findings echo other reports showing intact performance in maintenance (e.g., forward digit span) (Channon et al., 1993) but deficits in manipulation (e.g., backward digit span) (Channon et al., 1993) or updating of working memory content (Joormann and Gotlib, 2008, Levens and Gotlib, 2010, Yoon et al., 2014). Nevertheless, the exact nature of working memory deficits in depression is inconclusive as some studies also reported relatively unaffected n-back task performance in MDD (Barch et al., 2003, Fitzgerald et al., 2008, Harvey et al., 2005). A number of factors may have hindered the attempt to differentiate updating from maintenance impairment. First, most inconsistent findings came from investigations using the n-back task which simultaneously engages multiple complex processes, thus making it challenging to isolate the updating component of working memory. Several studies employing designs more specific to updating (e.g., modified Sternberg task) (Joormann and Gotlib, 2008, Yoon et al., 2014) indeed found deficits in depressed individuals. Second, as emotion interacts with cognitive processes extensively (Gotlib et al., 2004, van Tol et al., 2012), stimulus valence might have obscured basic processing dysfunctions involved in working memory updating in past work. Thus, it is imperative to dissociate the impacts of MDD on differentiable working memory processes (i.e., updating vs. maintenance) while minimizing the influence of emotional biases.
Previous neuroimaging work investigating working memory deficits in MDD has primarily emphasized prefrontal abnormalities due to its putative role in executive functions. While some reported hyperactivity with higher working memory demand (Fitzgerald et al., 2008, Harvey et al., 2005, Walsh et al., 2007, Walter et al., 2007), others also found hypoactivity or no change in prefrontal activation (Korgaonkar et al., 2013, Pu et al., 2011, Schöning et al., 2009). These inconsistent findings raise the question whether working memory impairment can be accounted for by prefrontal dysfunctions alone. Studies of healthy human adults and non-human primates have consistently demonstrated that the visual working memory circuit involves not only prefrontal regions (Cavada and Goldman-Rakic, 1989, Schall et al., 1995) but also visual associations cortices (Gazzaley et al., 2007, Zanto et al., 2011). Indeed, visual association areas preferentially respond, both in terms of activation and connectivity with prefrontal regions, during the selective maintenance of task-relevant visual information in comparison to irrelevant information (Gazzaley et al., 2007, Gazzaley et al., 2005b, Oh and Leung, 2010, Peters et al., 2012). In MDD research, neural aberrations have been reported in multiple visual regions during the selective processing of both emotional (Furey et al., 2013) and non-emotional task-relevant visual stimuli (Desseilles et al., 2009). There has been increasing interest in the impact of depression on visual cortical areas as their dysfunctions may significantly affect an individual's visual experience and internal state (Barrett and Bar, 2009). However, current understanding of visual cortical involvement in visual working memory updating in MDD, both at the regional and inter-regional levels, is sparse.
To investigate the alterations in brain functions during visual working memory updating and their relationship with cognitive performance as well as clinical characteristics of MDD, we utilized a working memory updating paradigm with a retrocue and visual stimuli of little emotional content. Neutral face and scene images were used in a delayed recognition task that contained an informative cue inserted during the delay period after stimulus presentation. We examined fMRI signal associated with stimulus category (face vs. scene) and task relevance (to be remembered vs. to be ignored) during working memory updating. A non-updating condition was included as a control for maintenance and memory load effects. Neural aberration associated with visual working memory updating in MDD was evaluated by examining regional activity, spatial activation patterns, and functional connectivity between prefrontal and visual association cortices. Multivariate pattern analysis was also conducted to examine potential changes in the integrity of visual cortical spatial activation patterns that may not be expressed in amplitude or connectivity. Changes in regional activity and functional connectivity were further assessed in relation to individual differences in task performance and clinical measures (e.g., rumination).
2. Methods and materials
2.1. Participants
Forty-two young adults were recruited to participate in the study: 20 currently unmedicated subjects with MDD (12 females; mean [SD] age = 22.0 [3.1] years) and 22 healthy control (CTL) subjects (12 females; mean [SD] age = 22.2 [3.4] years). All subjects underwent clinical screening assessments, including the Structured Clinical Interview for DSM-IV (SCID) (First et al., 2007). All depressed subjects met DSM-IV criteria for MDD and were in a current major depressive episode. A total of 9 MDD participants (50.0%) had comorbid diagnoses (35.3% social phobia, 29.4% specific phobia, 5.9% post-traumatic stress disorder, and 11.8% generalized anxiety). Six subjects were medication naïve. Individuals who had taken psychotropic medications within the past 8 weeks before fMRI were excluded from participation. Healthy individuals were without history of psychiatric or neurological illnesses based on their SCID and self-reports. The two groups were matched on gender, age, and years of education. All subjects completed the self-report version of the Inventory of Depressive Symptomatology (IDS) (Rush et al., 1996) and the Ruminative Response Scale (RRS) (Nolen-Hoeksema and Morrow, 1991) to assess severity of depressive symptoms and ruminative tendency, respectively. Rumination scores were further analyzed according to the three subscales including Reflection, Brooding, and Depression (Treynor et al., 2003). See Table 1 for demographic and clinical characteristics of participants included in the final analysis. All subjects gave written consent prior to participation. The study was approved by the local Institutional Review Board.
Table 1.
Demographics and clinical characteristics. Abbreviations: MDD, major depressive disorder; CTL, healthy control; IDS, inventory of depressive symptomatology; RRS, ruminative response scale; SD, standard deviation, * significantly different between the two subject groups (p's < 0.001).
| Variable | Group |
|
|---|---|---|
| MDD | CTL | |
| N (N female) | 18 (12) | 21 (12) |
| Age (years): mean (SD) | 22.0 (3.09) | 22.19 (3.38) |
| Education (years): mean (SD) | 12.4 (4.67) | 13.93 (6.48) |
| Ethnicity: Asian, Caucasian, Hispanic | 41%, 47%, 12% | 19%, 57%, 24% |
| Depression duration (years): mean (SD) | 3.87 (2.61) | N/A |
| RRS score: mean (SD)* | 63.52 (9.39) | 32.6 (6.96) |
| IDS score: mean (SD)* | 40.79 (8.70) | 5.57 (5.03) |
2.2. General experiment procedure
The fMRI portion was conducted within a week of the clinical interview for each participant. Prior to fMRI, participants completed the questionnaires, practiced the behavioral tasks, and were acclimated to the imaging procedures in a mock scanner.
2.3. Visual stimuli
Ninety pictures of faces (half were females) with neutral expressions were used as stimuli (Lundqvist et al., 1998, Minear and Park, 2004, Russell et al., 2007) and 90 pictures of scenes (urban houses and buildings) collected from the internet were used as stimuli in the working memory task. All images were scaled to the same size (154 × 186 pixels), converted to black and white, and equalized for overall brightness. Post-experiment stimulus valence ratings (from − 5 to 5) were obtained to determine whether the participants with MDD judged the faces more negatively or positively than the healthy controls. Both groups rated the face and scene images close to 0 or neutral in valence (M [SD] for faces: MDD = − 0.46 [0.92]; CTL = − 0.06 [0.75]; scenes: MDD = 0.79 [1.16]; CTL = 1.37 [1.03]), with no significant group differences (face: t(37) = 1.40, p = 0.17; scene: t(37) = 1.53, p = 0.14). Both subject groups rated scene stimuli to be more positive than face stimuli (p's < 0.002). For the localizer task, another set of face, scene, and common objects (20 images per category) was selected.
2.4. Behavioral tasks
2.4.1. Working memory task
We used a delayed recognition task with a cue inserted during the delay period similar to our previous study of healthy young adults (Oh and Leung, 2010). The task included two updating conditions (Remember Face, Remember Scene) and a control condition (Remember Both). See Fig. 1A for the task paradigm. At the beginning of each trial, a fixation cross was presented for 4 s, which turned green for 200 ms as a warning 500 ms before stimulus presentation. Two images (a face and a scene) were then presented sequentially, each for 800 ms with a 200-ms inter-stimulus interval, followed by a checkerboard mask displayed for 800 ms. The order of stimulus presentation was pseudorandomized and counterbalanced such that face and scene images appeared first in an equal number of trials. After a delay of 2.2 s, a cue word (“Face”, “Scene”, or “Both”) appeared at the center of the screen for 1 s, indicating the stimulus category/categories to be remembered for the recognition test. After a 9-s postcue delay, either the target or a new image of the cued category was presented as a probe for recognition. A face probe would be presented on trials with the “Face” cue, a scene probe for the “Scene” cue, and either a face or a scene probe for the “Both” cue (50/50 chance). Thus, all cues were fully informative; the “Face” and “Scene” cues required remembering only the relevant item whereas the “Both” cue required remembering both items for probe recognition. Chance of a matching and nonmatching probe was equal (50/50) for each condition. Participants responded with a button press to indicate whether the probe was the remembered item. There were 4 trials per condition per run for 6 runs, giving a total of 24 trials per condition.
Fig. 1.
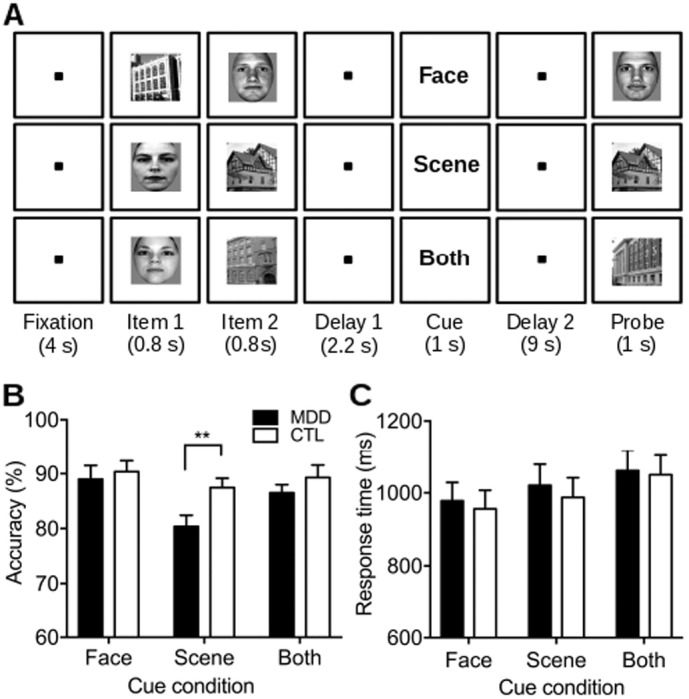
Task design and behavioral results. (A) Schematic of the working memory updating task. The three task conditions (Remember Face, Remember Scene, and Remember Both) were indicated by the cue word displayed after the presentation of the two picture stimuli, instructing the participants to remember only the face, scene, or both pictures, respectively. For simplicity, the visual mask and ITI (8–12 s) were not shown in the figure. (B) Mean accuracy (± standard error of the mean [SEM]) for each group by cue type. The MDD participants performed significantly worse on the Remember Scene condition compared to healthy controls (corrected for multiple comparisons). (C) Mean response time (± SEM) for each group by cue type. ** p ≤ 0.03.
2.4.2. Localizer task
We used 1-back task with three visual categories (face, scene, and object) to detect brain regions that exhibit preferential responses to faces and scenes. Each stimulus was presented for 800 ms with a 1200-ms ISI. The task consisted of 12 stimulus blocks (4 per category). Each stimulus block lasted 16 s and was separated by 16 s of fixation. Participants responded with a button press to indicate whether the currently displayed stimulus matched the immediately preceding stimulus.
2.5. Image data acquisition, preprocessing and analysis
Whole-brain images were acquired using the Siemens Trio 3 T System (Siemens, Erlangen Germany) at the Stony Brook University SCAN Center. High-resolution anatomical images were acquired (MPRAGE: TR = 1900 ms, TE = 2.53 ms, flip angle = 9 degrees, Matrix = 256 × 256, FOV = 220 × 220 mm, 176 slices, slice thickness = 1 mm). In-plane anatomical images of 33 axial-oblique slices, parallel to the anterior commissure–posterior commissure (AC–PC), were also collected (TR = 300 ms, TE = 5 ms, flip angle = 60 degrees, matrix = 256 × 256, FOV 220 × 220 mm2, slice thickness = 3.5 mm with 0.5 mm gap). During working memory task performance, functional images were acquired in the same orientation and slice thickness/gap as the inplane images using a single-shot T2* weighted EPI sequence (TR = 2000 ms, TE = 30 ms, flip angle = 80 degree, matrix = 72 × 72, FOV = 220 × 220 mm2, effective voxel size = 3.06 × 3.06 × 4 mm3, 180 volumes per run). Identical imaging parameters were used for collecting 200 volumes in a single run of the localizer task which was performed after the working memory task.
All images were preprocessed using SPM8 (http://www.fil.ion.ucl.ac.uk/spm/software/spm8/). Images were first screened by visual inspection for obvious artifacts and motion. Standard preprocessing steps were applied to each dataset including slice timing correction, volume alignment for motion correction, and coregistration of anatomical to the mean EPI image. A unified segmentation algorithm was applied to the high-resolution structural images to separate the gray matter, white matter, and CSF. The functional and anatomical images were then spatially normalized and transformed into a common MNI space, using affine nonlinear transformation, and then spatially smoothed with a 4-mm full-width at half-maximum Gaussian kernel.
The final analysis included 39 subjects: 18 MDD and 21 CTL. Data from two MDD and one CTL participants were excluded due to excessive head movements. Runs with significant motion (> 3-mm translation peak-to-peak movement and/or 1.5-degree rotation) were removed. Outlier volumes were identified when frame-to-frame displacement exceeded 0.5 mm and/or rotation > 1.5°, using the Artifact Detection Tools (ART, www.nitrc.org/projects/artifact_detect/). Outlier volumes ranged from 2 to 26% of all volumes across subjects (on average: MDD, 13.1%; CTL: 9.8%), and did not differ significantly between the two subject groups (p > 0.3).
First-level analysis was conducted using the General Linear Model (GLM). For each individual, a design matrix was constructed for the four events (stimulus presentation, cue, postcue delay, and probe) for each cue condition in the working memory task. The events were convolved with a canonical hemodynamic response function. Outlier volumes and the head motion parameters were accounted for in the GLM. The estimated parameters of the regressors (beta weights) were then calculated for each voxel. For the second-level analysis, treating subjects as a random effect, two-samples t-tests were conducted to evaluate the effects of interest using the corresponding contrast image from each individual of each group. We focused on the postcue delay to examine brain activations during working memory updating in MDD relative to CTL participants.
To control for familywise type I error, all group maps were thresholded at voxelwise p < 0.001 and corrected for multiple comparisons at false discovery rate, FDR < 0.05.
2.6. Regions-of-interest analysis
Category-selective areas in the ventral pathway were examined using regions-of-interest (ROIs) analysis. We used a group probabilistic map approach (Wang et al., 2016) to define face- (Fox et al., 2009) and scene-related (Epstein et al., 2007, Walther et al., 2009) regions using the localizer task data. First, face and scene activation maps were generated for each individual using their face versus scene and scene versus face contrasts, respectively. The resulting activation maps were thresholded (Z > 2.3, p < 0.001, uncorrected) and binarized. The binarized images for each visual category (face and scene) were then averaged across subjects, resulting in a probabilistic map for each category. Each voxel within each probabilistic map thus had a value representing the proportion of subjects with suprathreshold activation for the corresponding visual category. Thus, these probabilistic maps showed the interindividual variability of face- and scene-related activations. To create the final functional ROI masks, only voxels from the probabilistic maps with a value ≥ 0.2 were kept (i.e., at least 20% of the subjects showed suprathreshold activation in these voxels). The ROIs were further constrained by eliminating voxels with a high probability of being outside the gray matter (using the group averaged anatomical image, with 30% gray matter probability) (Fig. 2). This method of selecting ROIs is consistent with previous studies investigating visually selective regions and has been shown to be reliable in identifying activations sensitive to visual demand (Kawabata Duncan and Devlin, 2011, Wang et al., 2016, Zhen et al., 2015). The face network contained regions commonly associated with face processing, including the fusiform face area (FFA), occipital face area (OFA), and superior temporal sulcus (STS). Similarly, the scene network contained regions commonly associated with scene processing, including the parahippocampal place area (PPA), transverse occipital sulcus (TOS), and retrosplenial cortex (RSC). These ROIs were used for the reported univariate and multivariate analyses. Due to the increased concern of partial volume effect by including more heterogeneous voxels, for the psychophysiological interaction analysis, the right FFA and PPA were individually defined. We focused on the right hemisphere ROIs due to their stronger visual selectivity as shown in previous studies (Epstein et al., 2001, Jonas et al., 2016, Kanwisher et al., 1997). Peak voxels were identified at p < 0.001 and were required to have a cluster of at least three contiguous voxels. ROIs were spheres of 5-mm radius centered at the peak coordinates. (Note: these subject-specific ROIs produced similar results for the univariate and multivariate analysis as the group probabilistic ROIs.) For all face and scene network ROIs, the beta weights were extracted for the postcue delay period activity and connectivity of each cue condition for each individual. Two-way ANOVA's were conducted to determine main effects of group and cue condition and their interactions.
Fig. 2.
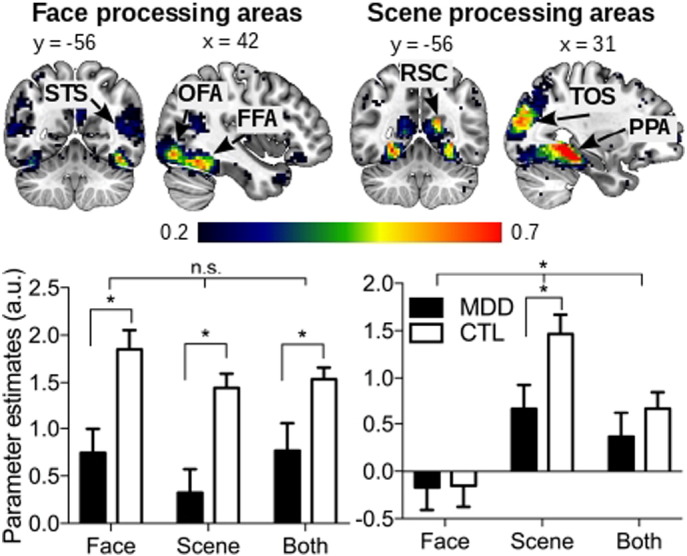
Top row: Group probabilistic maps of face network (left) and scene network (right) activation using data from the localizer task. Color bar indicates the proportion of participants who showed suprathreshold visual category-selective activation. Bottom row: Bar charts showing postcue delay activity in the face (left) and scene (right) networks during the three working memory updating conditions. Significant Group × Condition interaction was found in the scene network. Relative to the CTL group, the MDD group showed significantly lower activation of the face network and scene network when maintaining the task-relevant face and scene item was required, respectively. * corrected p ≤ 0.05. Abbreviations: FFA, fusiform face area; OFA, occipital face area; STS, superior temporal sulcus; PPA, parahippocampal place area; TOS, transverse occipital sulcus; RSC, retrosplenial cortex.
To confirm the visual category (face vs scene) selectivity of brain activation in the face and scene networks using the localizer task data, we used a signal detection theory measure (Green et al., 1969):
where μ and σ were the mean and standard deviation of the ROI activity during the specific task event.
2.7. Functional connectivity analysis
The frontal functional connectivity with FFA and PPA was estimated using the psychophysiological interactions (PPI) (Friston et al., 1997) using the gPPI toolbox (McLaren et al., 2012) (http://www.nitrc.org/projects/gppi). A PPI model was created for each subject with three main components: the physiological term which represents the time series from the seed region, the psychological term which represents the task conditions (e.g., Remember Face and Remember Scene), and the psychophysiological interaction term. The PPI was computed as the element-by-element product of the deconvolved time series of the seed region and a task condition vector (Garraux, 2005, Stephan et al., 2003). All 3 conditions (Remember Face, Remember Scene, and Remember Both) for each task event (stimulus presentation, cue and postcue delay, and probe) were included in the model. PPI of category-specific working memory updating was calculated for each subject by contrasting the postcue delay event of the two updating conditions. As the interaction between the left middle frontal gyrus (MFG) and visual association areas has been previously implicated in visual working memory updating in healthy adults (Chadick and Gazzaley, 2011, Gazzaley et al., 2007), we independently defined the left MFG in each subject using the load effect (Remember Both vs Remember Face/Scene) and used this ROI as the seed. Beta weights of the psychophysiological interaction term between left MFG-right FFA/PPA coupling and the experimental condition during the postcue delay period were extracted and used in two-way ANOVA tests.
2.8. Multivariate pattern analysis (MVPA)
We used linear support vector machines (SVM) to conduct multivoxel pattern classification of category-specific activity patterns in the face and scene network ROIs (see Regions-of-interest analysis) during the working memory task. Methodological details of the analysis can be found in our previous study (Han et al., 2013). Briefly, classifiers were trained in the localizer task data to discriminate face vs scene related activation patterns in the working memory task data using the LibLinear SVM package (http://www.csie.ntu.edu.tw/~cjlin/liblinear/) with L2-regularization, L2-loss function, and bias = 1. In building these binary classification models, the regularization vs. loss tradeoff parameter C was determined (from the set [0.001, 0.1, 1, 10, 1000]) for each cross-validation fold by using a subset of the training samples of each fold for the nested cross validation (see below). Classification accuracy for each individual in discriminating Remember Face vs Remember Scene from the working memory task was determined using the leave-one-out cross-validation approach.
More specifically, for each subject, in each fold of cross validation, the classifier was trained on data from the localizer task and tested on each trial of the working memory task. This procedure was repeated until all trials were tested. Training samples were the averaged activation patterns of two scans during the localizer task block (5th and 6th scans) and testing samples were the average of two scans during early probe period (the 12th and 13th scans).
Since the ROIs are functionally specialized in response to faces or scenes, the reported classification accuracy of category-specific activation patterns for each ROI was calculated based on the classifier's performance on the trial type congruent with the ROI's category selectivity (i.e., percentage of Remember Face and Remember Scene trials that were correctly classified using the activation patterns of the face network and scene network ROI, respectively). Average classification accuracy and standard error of the mean were computed across the participants in each group.
MVPA was done in each participant's native space without smoothing. To ensure spatial consistency across analyses, ROIs in MNI space used in the univariate analysis were transformed back into the participant's native space, using the inverse normalization matrix (output from segmentation-based normalization of SPM8).
3. Results
3.1. Behavioral results
While both groups performed well on the working memory task, the MDD participants performed slightly worse across all task conditions relative to the CTL participants (Fig. 1B & C). Two-way ANOVA's with the three cue conditions and two subject groups as the within- and between-subject factors revealed a significant main effect of Condition (accuracy: F(2, 74) = 8.77, p < 0.001; RT: F(2, 74) = 26.81, p < 0.001), while the Group × Condition interaction did not reach the significance threshold (accuracy: F(2, 74) = 2.26, p = 0.11). This weak group by condition interaction was driven by the MDD group's lower accuracy on the Remember Scene condition compared to the CTL group (t(37) = 2.68, corrected p = 0.03). All other effects were not significant (p's > 0.12). Both groups showed faster RT on the updating conditions, indicating cue-related performance facilitation. Furthermore, the two groups did not differ in their performance on the more demanding non-updating condition (Remember Both) (accuracy and RT: p's > 0.3). Taken together, the MDD group's impairment on the working memory task was especially pronounced in the updating condition in which removing the irrelevant face and maintaining the relevant scene item were required.
3.2. fMRI results
3.2.1. Scene- and face-related regions in the ventral pathway and visual category selectivity
We first determined whether the stimulus-related responses in the visual association areas were comparable between the MDD and CTL groups. Face- and scene-related ROIs were similarly identified for both groups using the localizer task data (Fig. 2) (For further information on group maps of face- and scene-related activation, see Table 2). Using the signal detection theory equation (see Methods), we found no group differences in visual category selectivity in the localizer task data for either the face or scene network (corrected p's > 0.18).
Table 2.
Face- and scene-related brain activations for the MDD and CTL groups. Suprathreshold activation clusters are shown for the Face > Scene and Scene > Face contrasts, using data from the localizer task. Peak MNI coordinates in mm, t values and Z scores are listed for each region. Abbreviations: FFA, fusiform face area; OFA, occipital face area; STS, superior temporal sulcus; PPA, parahippocampal place area; TOS, transverse occipital sulcus; RSC, retrosplenial cortex. All clusters were significant at threshold corrected for multiple comparisons at false discovery rate, FDR, p < 0.05 unless otherwise noted.
| MNI coordinates |
|||||||
|---|---|---|---|---|---|---|---|
| Region | Cluster size | x | y | z | t | Z | p (FDR) |
| MDD group | |||||||
| Face > Scene | |||||||
| Right OFA | 30 | 42 | − 79 | − 14 | 6.5 | 4.5 | 0.005 |
| Precuneus | 74 | 3 | − 55 | 37 | 5.4 | 4.1 | 0.001 |
| Right amygdala | 30 | 21 | − 7 | − 14 | 6.5 | 4.5 | 0.005 |
| Left amygdala | 40 | − 21 | − 4 | − 17 | 7.2 | 4.8 | 0.002 |
| Scene > Face | |||||||
| Right PPA | 299 | 30 | − 46 | − 11 | 11.7 | 6.1 | 0.001 |
| Left PPA | 302 | − 21 | − 46 | − 14 | 10.1 | 5.7 | 0.001 |
| Right TOSa | 205 | 39 | − 79 | 28 | 8.9 | 5.4 | 0.001 |
| Left TOS | 418 | − 33 | − 85 | 13 | 9.4 | 5.5 | 0.001 |
| Right RSCa | 45 | 18 | − 52 | 16 | 7.9 | 5 | 0.001 |
| Left RSC | 55 | − 15 | − 58 | 16 | 6.5 | 4.5 | 0.001 |
| CTL group | |||||||
| Face > Scene | |||||||
| Right FFA | 152 | 45 | − 49 | − 23 | 7.1 | 5 | 0.001 |
| Left FFA | 28 | − 42 | − 46 | − 20 | 6.7 | 4.8 | 0.01 |
| Right OFAa | 28 | 39 | − 82 | − 11 | 6.7 | 4.8 | 0.001 |
| Left OFA | 33 | − 42 | − 82 | − 8 | 6.1 | 4.6 | 0.005 |
| Right STS | 102 | 42 | − 43 | 16 | 5.4 | 4.2 | 0.001 |
| Left STS | 18 | − 54 | − 55 | 4 | 4.5 | 3.7 | 0.035 |
| Precuneus | 35 | 3 | − 55 | 25 | 6.8 | 4.8 | 0.005 |
| Right amygdala | 73 | 21 | − 1 | − 17 | 6.7 | 4.8 | 0.001 |
| Left amygdala | 63 | − 21 | − 7 | − 14 | 8.4 | 5.5 | 0.001 |
| Scene > Face | |||||||
| Right PPA | 379 | 33 | − 37 | − 11 | 12.4 | 6.5 | 0.001 |
| Left PPA | 365 | − 27 | − 46 | − 11 | 21.1 | Inf | 0.001 |
| Right TOSa | 394 | 33 | − 82 | 13 | 10.9 | 6.2 | 0.001 |
| Left TOSa | 285 | − 30 | − 82 | 28 | 12.5 | 6.5 | 0.001 |
| Right RSCa | 90 | 15 | − 52 | 10 | 10.5 | 6.1 | 0.001 |
| Left RSCa | 48 | − 15 | − 55 | 10 | 7.4 | 5.1 | 0.001 |
Cluster separated at p < 0.0001.
3.2.2. Postcue activity during working memory updating
Our main focus was to examine whether the face and scene network activity during selective working memory maintenance was altered in the MDD group. For both groups, postcue activity (measured by beta weights) showed category selectivity by cue condition (i.e., greater responses to the visually preferred and task-relevant category; see Fig. 2, bottom graphs). For the face network, the two-way ANOVA's (Group [MDD, CTL] × Condition [Remember Face, Remember Scene, Remember Both]) revealed a significant main effect of Condition (F(2, 74) = 3.93, p = 0.024) and Group (F(1, 37) = 81.70, p < 0.001) but not the Group × Condition interaction (F < 1). Post hoc tests revealed significantly lower beta weights for all three conditions in the MDD group relative to the CTL group (corrected p's < 0.05). The scene network showed a significant main effect of Condition (F(2, 74) = 46.28, p < 0.001), and significant Group × Condition interaction (F(2, 74) = 4.61, p = 0.013), but not the main effect of Group (F(1, 37) = 1.76, p = 0.19). Post hoc analyses indicated that the interaction effect was driven by the significantly lower activation for the Remember Scene condition in the MDD group compared to CTL group (t(1, 37) = 2.46, corrected p = 0.05). Thus, both face and scene networks in MDD demonstrated reduced activation when the face and scene items were respectively cued for selective maintenance.
Exploratory whole-brain analyses showed postcue delay activation in all scene-related regions (i.e., PPA, TOS, and RSC) in both groups during Remember Scene compared to Remember Face, although the MDD group showed significantly weaker scene-related activation in bilateral TOS/middle occipital gyrus and left posterior parietal (p < 0.05 FDR corrected; Table 3). Similar to previous studies (Gazzaley et al., 2005a, Oh and Leung, 2010), neither groups showed suprathreshold activations in the Remember Face versus Remember Scene contrast as face-related activation was more variable across subjects. Further, consistent with our previous study (Oh and Leung, 2010) using a similar paradigm, the healthy adults showed a memory load effect (i.e., Remember Both vs. Remember Face/Scene) in several regions including bilateral MFG, left superior parietal lobule, and precuneus. In contrast, load-related activations were much weaker in the MDD group, only found at a very lenient threshold (p < 0.05, uncorrected).
Table 3.
Brain activation in each group and group differences during selective scene working memory. Suprathreshold activation clusters are shown for the Remember Scene > Remember Face contrast during postcue delay of the working memory task. Abbreviations: MOG, middle occipital gyrus. See Table 2 for other annotations and abbreviations. All clusters were significant at threshold corrected for multiple comparisons at false discovery rate, FDR, p < 0.05 unless otherwise noted.
| MNI coordinates |
|||||||
|---|---|---|---|---|---|---|---|
| Region | Cluster size | x | y | z | t | Z | p (FDR) |
| MDD group | |||||||
| Right PPA | 250 | 27 | − 43 | − 8 | 11.6 | 6 | 0.001 |
| Left PPA | 218 | − 24 | − 43 | − 8 | 11.3 | 6 | 0.001 |
| Right TOS | 55 | 39 | − 82 | 22 | 6.2 | 4.3 | 0.001 |
| Left TOS | 30 | − 39 | − 85 | 19 | 5.2 | 4 | 0.005 |
| CTL group | |||||||
| Right PPAa | 304 | 30 | − 31 | − 17 | 14.7 | 7 | 0.001 |
| Left PPA | 294 | − 27 | − 46 | − 11 | 12.5 | 6.5 | 0.001 |
| Right TOSa | 213 | 30 | − 85 | 19 | 10.8 | 6.2 | 0.001 |
| Left TOS | 390 | − 30 | − 88 | 10 | 10.7 | 6.1 | 0.001 |
| Left posterior parietalb | − 33 | − 88 | 25 | 8.3 | 5.4 | 0.001 | |
| − 33 | − 82 | 31 | 8 | 5.3 | 0.001 | ||
| Right RSC | 105 | 21 | − 55 | 13 | 6.7 | 4.8 | 0.001 |
| Left RSC | 108 | − 15 | − 61 | 19 | 6.2 | 4.6 | 0.001 |
| CTL > MDD | |||||||
| Right TOS/MOG | 26 | 30 | − 85 | 16 | 4.4 | 3.9 | 0.043 |
| Left TOS/MOG | 35 | − 33 | − 88 | 4 | 5.2 | 4.5 | 0.021 |
| Left posterior parietal | 37 | − 24 | − 82 | 37 | 4.7 | 4.1 | 0.021 |
Cluster separated at p < 0.0001.
Part of the left TOS cluster.
One particular goal of the current study was to determine whether core clinical features of MDD were related to the visual cortical abnormalities during working memory updating. Thus, we examined the relationship between individual differences in rumination and depressive symptoms and postcue delay activation associated with the processing of no-longer-relevant stimuli. Across individuals with MDD, higher Brooding subcomponent of rumination correlated with heightened postcue delay face network activity during Remember Scene (ignore face) (r = 0.72, p = 0.001) (Fig. 3A) but not with the scene network activity during Remember Face (ignore scene) (r = 0.31, p = 0.21). Conversely, pronounced depressive symptoms significantly correlated with greater postcue delay scene network activity during Remember Face (ignore scene) (r = 0.59, p = 0.01) (Fig. 3B) but not with postcue delay face network activity during Remember Scene (ignore face) (r = 0.43, p = 0.08). No significant correlations were found in the CTL group (r2's < 0.14, p's > 0.09). Taken together, the higher the rumination and depressive symptoms reported by the MDD subjects, the greater the postcue activity in visual association regions associated with the processing of no-longer relevant information.
Fig. 3.
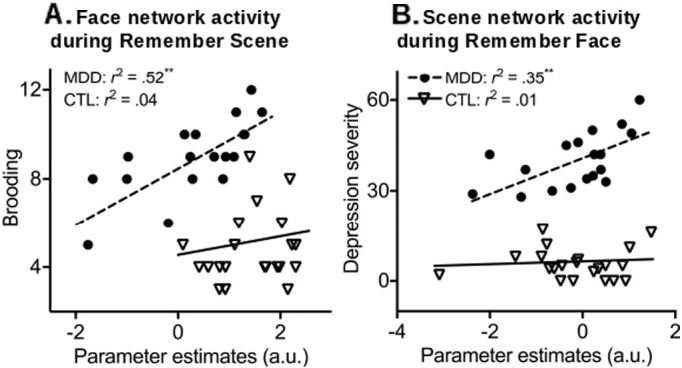
Relationship between clinical measures of MDD and maintenance of no-longer-relevant information. The MDD group showed that (A) higher rumination (Brooding component) was associated with heightened face network activity when the face item should be ignored, and (B) more pronounced depressive symptoms were associated with heightened scene network activity when the scene item should be ignored. No significant correlations were found for the CTL group. ** p ≤ 0.01.
3.2.3. Classification of activation patterns during probe recognition
We applied multivoxel pattern analysis to examine spatial patterns of activation in the face and scene networks during probe recognition of the two updating conditions. In both groups, the classifier trained on the activation patterns of the localizer task was able to discriminate activation patterns during the working memory task probe recognition in correspondence to the probed visual category. For both groups, classification accuracy was significantly above chance for both the face and scene networks (p's < 0.017, one-sample t-tests). However, the classification accuracy for the scene network was significantly lower for the MDD group compared to the CTL group (t(37) = 2.25, p = 0.03; see Fig. 4). This further showed that not only the amplitude but also the spatial pattern of scene-related activation during working memory was altered in the MDD group.
Fig. 4.
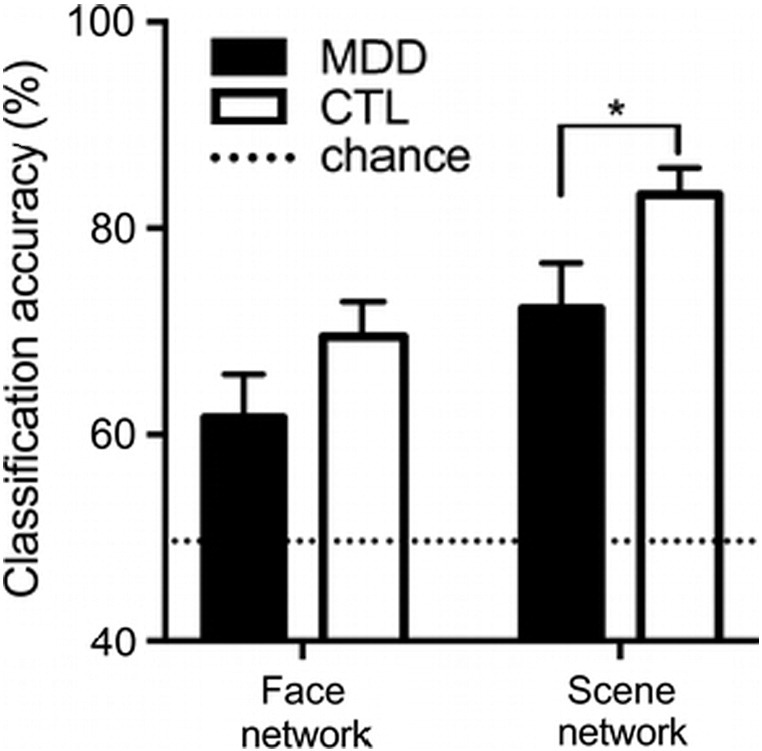
Significantly weaker category-selective spatial patterns of activation during selective scene working memory in MDD. Bar graph shows above-chance (> 50%) classification accuracy of face and scene trials using spatial activation patterns of the face and scene network, respectively. * p < 0.05.
3.2.4. Postcue functional connectivity during working memory updating
We used the left MFG as the seed in examining functional connectivity with the right FFA and PPA since combining heterogeneous time series across visual regions can be problematic for the PPI analysis. Individual subjects' beta weights for this functional connectivity were extracted and entered into two-way ANOVA's to characterize potential impacts of MDD (Fig. 5). The left MFG-right FFA connectivity showed a significant main effect of Condition (F(2, 74) = 8.95, p < 0.001) but not the main effect of Group (F < 1) or Group × Condition interaction (F(2, 74) = 1.09, p = 0.34). The right PPA-left MFG showed a significant main effect of Condition (F(2, 74) = 3.79, p < 0.03) and significant Group × Condition interaction (F(2,74) = 4.0, p < 0.03) but not significant Group main effect (F < 1). Post hoc analyses showed a significant effect of Condition only in the CTL group (F(2, 40) = 8.27, p < 0.001) but not the MDD group (F < 1), indicating a lack of differentiating modulation effects by updating cues on functional connectivity in the depressed individuals.
Fig. 5.
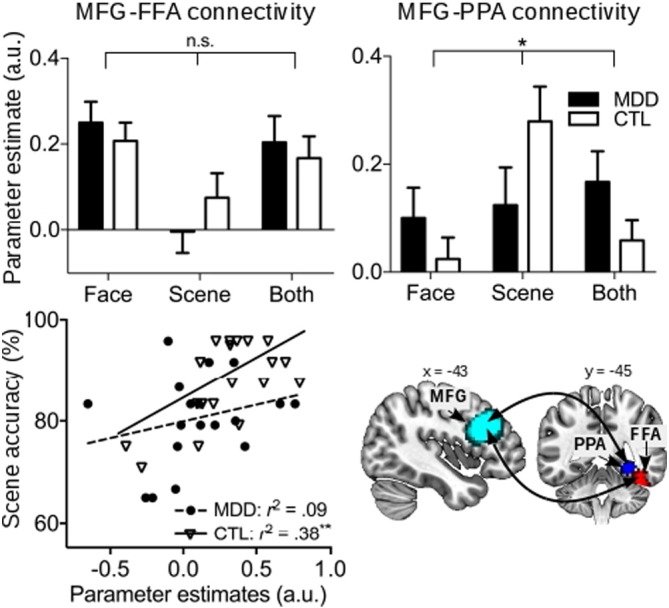
Top row: left MFG connectivity with the right FFA (left) and right PPA (right) during working memory updating. MFG-PPA connectivity showed significant Group × Condition interaction. Bottom row: relationship between MFG-PPA connectivity strength and performance on the Remember Scene condition (left), and ROI locations of the left MFG (cyan), FFA (red), and PPA (blue) in all subjects (right). * p < 0.05, ** p < 0.01.
We further examined the relationship between functional connectivity and behavioral performance. Across all subjects, the beta weights for left MFG-right FFA connectivity during Remember Face significantly correlated with performance accuracy on the Remember Face condition (r = 0.48, p = 0.003) after controlling for age, IDS, and RRS (data not shown). Similarly, across all subjects, the beta weights for left MFG-right PPA connectivity during Remember Scene significantly correlated with performance accuracy on the Remember Scene condition (r = 0.49, p = 0.002) (Fig. 5) after controlling for age, IDS, and RRS. Thus, stronger prefrontal connectivity with the visual association areas was associated with better updating performance across subjects. However, when examining the MFG-PPA connectivity correlation separately by group, only the CTL group showed a significant correlation with behavioral performance (r = 0.62, p = 0.01) but not the MDD group (p = 0.36). Similar effect was not observed for the MFG-FFA connectivity. The Brooding subscale of rumination also significantly correlated with the left MFG-right FFA connectivity during the Remember Scene (ignore face) condition in the MDD (r = 0.51, p = 0.03) but not CTL group (r = 0.28, p = 0.21). In sum, the MDD group showed a lack of preferential responses in terms of prefrontal-PPA functional connectivity for the task-relevant scene item during updating while heightened prefrontal-FFA coupling was associated with increased in ruminative brooding tendency across subjects. In general, the strength of prefrontal-visual association connectivity predicted performance on both working memory updating conditions across all subjects.
4. Discussion
Using a design that manipulates the task relevance of working memory items of two distinct visual category, we showed that visual association areas in individuals with MDD exhibited altered regional activation patterns and functional connectivity with prefrontal areas during working memory updating, particularly in the Remember Scene condition. We also demonstrated that enhanced functional connectivity between the prefrontal and visual association areas predicted better performance across subjects, and that rumination and depressive symptoms were associated with the maintenance of obsolete visual information in working memory. In contrast, little or no significant deficit was evident for the non-updating (Remember Both) condition. These findings extend previous work by showing visual working memory updating dysfunctions in MDD involves neural aberrations not only at the prefrontal but also at the visual cortical level.
4.1. Altered regional activity and spatial activation patterns in visual association areas during working memory updating
Compared to the CTL group, alterations in activation during working memory updating were found across both face and scene-processing networks in MDD. The scene network in MDD also showed weaker spatial patterns of activation associated with the task-relevant scene information. These findings add to the growing body of research showing that the neural correlates of impaired visual working memory in MDD extend beyond the structures typically associated with working memory (e.g., prefrontal cortex) or emotion modulation (e.g., amygdala) to include visual cortical areas. Several studies have reported alterations along the ventral visual pathway including middle occipital as well as more downstream areas (e.g., fusiform gyrus) during working memory (Furey et al., 2013), attention (Desseilles et al., 2011, Desseilles et al., 2009), and visual categorization (Fu et al., 2008, Suslow et al., 2010) tasks. In particular, a recent pharmacological fMRI study (Furey et al., 2013) showed that anti-muscarinic scopolamine both enhanced visual working memory performance and partially recovered middle occipital activation which also correlated with the degree of clinical response to the drug treatment in depressed patients. Spatial activation patterns in visual regions have also been used to successfully distinguish between healthy and depressed individuals (Costafreda et al., 2009, Marquand et al., 2008, Zeng et al., 2012). As spatial patterns in the ventral visual cortex are thought to carry information about the task-relevant visual information in working memory (Christophel and Haynes, 2014, Han et al., 2013, Harrison and Tong, 2009, Xing et al., 2013), our findings not only demonstrate the impact of depression on category-selective activation but also potential loss of visual information.
Our findings of altered visual cortical functions in MDD offer several cognitive and clinical implications. First, as the visual association regions play a crucial role in perception (Allison et al., 1994), attention (Kastner and Ungerleider, 2000), and working memory (Christophel et al., 2012, Harrison and Tong, 2009), their dysfunction can contribute to impaired cognitive control of information processing and ultimately depressive symptoms. Specifically, perseverative focus on previous adverse experiences can lead to the formation of negative schemas (i.e., automatic thoughts and negative self-referential beliefs) in depressed individuals (Disner et al., 2011). Failure to update using subsequent inputs can prevent adaptive interpretations and appraisals, leading to sustained negative affect and emotion dysregulation that are central to depression (Gotlib and Joormann, 2010). Second, our findings showed that visual cortical dysfunctions can contribute to cognitive inflexibility in situations with little or no negative affect. While most studies have associated working memory impairments in MDD with “negativity bias” (i.e., enhanced attention/memory for negative emotional material), emerging evidence indicates that alterations in visual cortical processing occur without emotional influence from visual stimuli (Desseilles et al., 2009, Garrett et al., 2011). Nevertheless, the current study does not suggest an independent role of visual cortical aberration in the etiology of depression. Rather, dysfunctional visual regions during working memory likely contribute to cognitive control deficits through interacting with higher-order areas to exacerbate the sensitivity to and persistent processing of task-irrelevant information.
Several factors may have led to the disrupted activity and spatial patterns of activation in the visual association areas. One possibility is that intrusive no-longer-relevant items interfered with the maintenance of the task-relevant items in working memory. This is supported by the correlation between the pronounced clinical measures (i.e., rumination and depressive symptoms) and the increased activation and connectivity associated with the irrelevant visual information in MDD. Such relationship, along with the impaired behavioral performance, suggests that depressive mood state and ruminative processing style may have reduced the ability to inhibit obsolete material which then interfered with the ability to focus on the relevant material (see below). This interpretation is consistent with the effects of stimulus competition and interference on working memory integrity demonstrated in previous behavioral studies (Bays et al., 2011, Gorgoraptis et al., 2011, Zokaei et al., 2011). Another possibility is the disruption of task-related signal from the prefrontal cortex. Unlike the CTL group, the MDD subjects did not show significant load-related activation during working memory maintenance. Together with weakened prefrontal-visual association area connectivity, these results suggest that prefrontal alterations may contribute to updating deficits at the visual cortical level in MDD. Finally, abnormal general visual processing which could manifest as impairment at the encoding stage of working memory in MDD could also lead to similar effects. This is less likely as visual category selectivity of visual cortical activation for faces and scenes during the stimulus presentation period was comparable between the two subject groups. Furthermore, the MDD subjects were not particularly impaired on the more demanding non-updating condition. While visual association areas have been shown to play a role in both perception and working memory, these results suggest differentiable visual cortical alterations in selective maintenance of task-relevant visual information during working memory updating.
4.2. Altered functional connectivity between prefrontal and visual areas during working memory updating
As the ability to selectively respond to task demands is not considered an intrinsic property of visual association regions (Gazzaley et al., 2007), their aberrant activity during working memory updating is likely related to altered modulatory signals from higher-order structures. Indeed, we found deficient coupling between the right PPA and left MFG during selective scene processing while the strength of MFG-PPA and MFG-FFA connectivity predicted across-subject task performance, suggesting the involvement of multiple regions in working memory dysfunctions in MDD. Our results are consistent with findings from a recent study (Desseilles et al., 2011) which related abnormal effective connectivity between the intraparietal sulcus and visual region V4 to the deficient attentional filtering of distracting information in depressed individuals. The interaction between prefrontal, parietal, and visual areas has been investigated extensively in healthy human and non-human primates. During the short-term retention of visual information, the putative top-down signal from the prefrontal cortex may serve to enhance or dampen neural processing in visual areas in correspondence to task goals (for a review, see Gilbert and Li, 2013). The loss of this signal can severely impair visual processing and task performance (Tomita et al., 1999). In further support of this notion, we also observed that when separated by group, the strength of the MFG-PPA connectivity significantly correlated with performance accuracy only in the CTL group but not the MDD group. This suggests a potential disconnect between prefrontal processes and task-relevant behaviors in MDD. In conjunction with the MDD group's poorer Remember Scene performance, these results pointed to the reduced modulatory role of the prefrontal cortex in the fronto-visual cortical circuit during visual working memory and its potential negative impact on behavioral outcome. To our knowledge, our study is the first to show alterations in prefrontal-visual association connectivity in MDD during working memory updating and directly relate this connectivity to behavioral outcome.
It is plausible that the diminished updating cue effects in connectivity in our data was related to prefrontal abnormalities. In previous studies, MDD-related alterations in prefrontal anatomy have been documented including reduction in glucose metabolism (Baxter et al., 1989, Kennedy et al., 2001), fractional anisotropy (Liao et al., 2013), and regional cerebral blood flow (Bench et al., 1992, Oda et al., 2003). While the investigation of anatomical integrity is beyond the scope of this study, we found that the MDD group did not show similar prefrontal responses to working memory load as in the CTL group. This lack of differential prefrontal responses between the updating and non-updating task conditions indicates impaired updating such that depressed subjects may have maintained both items to a certain extent even after being cued to update. This explanation is in line with the theoretical model proposing that the prefrontal network serves as a gating mechanism (O'Reilly and Frank, 2004). In sum, the abnormal prefrontal signal may potentially lead to lower functional connectivity with the visual regions, which in turn affects the selective processing of task relevant information and inhibition of task irrelevant information in visual regions, ultimately incurring behavioral consequences in MDD.
4.3. Disrupted working memory updating and rumination in depression
In examining the relationship between rumination and the neural substrates of working memory updating deficits, we found that brooding, a subcomponent of rumination, significantly correlated with both increased activation in the face network and prefrontal connectivity with the right FFA during the Remember Scene condition. As brooding represents moody pondering, our results suggest that maladaptive mood-related thinking patterns were associated with the neural correlates of irrelevant face processing while remembering scene was task relevant. Such association helps shed light on a number of points. First, it provides a neural basis for the reliable observation in neuropsychological studies which showed that rumination in MDD is related to cognitive inflexibility, including ineffectual negative material updating (Joormann and Gotlib, 2008, Yoon et al., 2014), reduced internal shifting capacity (De Lissnyder et al., 2011), and impaired suppression of distraction (MacQueen et al., 2000). Second, it reveals potentially differentiable impacts of negative mood state on the selective processing of face relative to scene information. While the depressed subjects did not rate the face or scene stimuli to be negative compared to the healthy control in our post task evaluation, only the correlation between brooding and the neural correlates of irrelevant face representation was significant.
Previous research has emphasized bias for interpersonal stimuli such as faces in the maintenance of depressive symptoms (Hames et al., 2013). A recent study has also shown that the STS, which is implicated in processing faces, exhibits preferential responses to social input (Deen et al., 2015). As the STS was part of our face network, it is plausible that depressive mood in our MDD sample may have triggered the biased processing of faces as well as the social characteristic of faces, leading to the ruminative and persistent retention of the irrelevant face in working memory. Interestingly, the neural correlates of irrelevant scene processing significantly correlated with symptom severity but not rumination, which suggests that depression affects the updating of non-interpersonal information but not through a ruminative behavioral style. These different relationships between face and scene neural processing and distinct clinical features may have contributed to the asymmetrical findings of impairments in the face and scene network activity and connectivity in our MDD group. It is noteworthy that while the subjects' stimulus valence ratings were close to neutral, both groups perceived faces to be slightly more negative than scenes. This phenomenon has been previous reported in depressed individuals who are more likely to judge emotionally neutral or ambiguous to be negative than healthy controls (Lawson and MacLeod, 1999, Leppänen et al., 2004). Thus, the combined tendency for bias toward the social nature of faces and negativity may have facilitated and/or prioritized the processing of the face stimulus at the expense of scene-related processing. Further research is needed to dissociate the impacts of symptom severity and rumination on different stimulus types in depression.
5. Limitations
The current study has a relatively small MDD sample size primarily due to the difficulty generally encountered in recruiting unmedicated depressed individuals. However, given the converging findings for impairments observed across behavioral, clinical, and neural measures, our results provide novel findings in specifying working memory dysfunctions in unmedicated individuals with MDD. Given the relatively young age of our participants, our results may potentially reflect deficits patterns specific to this age group. Thus, generalization to other clinical populations may be limited and future studies should test on a wider age range.
6. Conclusions
Findings from the current study provide novel evidence for the relationship between impaired visual working memory updating, altered regional neural activity, spatial activation patterns, and abnormal functional connectivity between visual and prefrontal areas in MDD. These findings suggest that clinical characteristics of MDD and cognitive deficits may be closely connected through a common mechanism involving alterations in visual and prefrontal processing during the control of visual information selection and maintenance. Such alterations are potentially related to the reduced ability to enhance task relevant representation and inhibit obsolete information, contributing to both cognitive difficulties and prolonged depressive states in MDD. This may be a useful model for further investigation of this multifaceted disorder.
Role of funding source
This work was supported by funding from the National Institute of Mental Health (RO1 MH069942, Klein) and Stony Brook Research Foundation (Leung).
Acknowledgments
The authors wish to thank Vickie Bhatia, Allison Danzig, Lauren Delaparte, Dawna Shimabukuro, and Megan Tudor for their help with screening participants. We thank Alyssa Barbas, Catherine Barnes, Gabrielle Baylon, Willa Dean, and Jacob Fogel for assistance with data management. We thank Xufeng Han for help with multivariate pattern analysis. We also thank Kim Burke and Elisabeth Caperalli of the SCAN center for their technical support.
References
- Allison T., McCarthy G., Nobre A.C., Puce A., Belger A. Human extrastriate visual cortex and the perception of faces, words, numbers, and colors. Cereb. Cortex. 1994;4:544–554. doi: 10.1093/cercor/4.5.544. [DOI] [PubMed] [Google Scholar]
- Barch D.M., Sheline Y.I., Csernansky J.G., Snyder A.Z. Working memory and prefrontal cortex dysfunction: specificity to schizophrenia compared with major depression. Biol. Psychiatry. 2003;53:376–384. doi: 10.1016/s0006-3223(02)01674-8. [DOI] [PubMed] [Google Scholar]
- Barrett L.F., Bar M. See it with feeling: affective predictions during object perception. Philos. Trans. R. Soc. B. 2009:1325–1334. doi: 10.1098/rstb.2008.0312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baxter L.R., Jr., Schwartz J.M., Phelps M.E., Mazziotta J.C., Guze B.H., Selin C.E., Gerner R.H., Sumida R.M. Reduction of prefrontal cortex glucose metabolism common to three types of depression. Arch. Gen. Psychiatry. 1989;46:243–250. doi: 10.1001/archpsyc.1989.01810030049007. [DOI] [PubMed] [Google Scholar]
- Bays P.M., Marshall L., Husain M. Temporal dynamics of encoding, storage, and reallocation of visual working memory. J. Vis. 2011;11:1–15. doi: 10.1167/11.10.6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bench C.J., Friston K.J., Brown R.G., Scott L.C., Frackowiak R.S.J., Dolan R.J. The anatomy of melancholia–focal abnormalities of cerebral blood flow in major depression. Psychol. Med. 1992;22:607–615. doi: 10.1017/s003329170003806x. [DOI] [PubMed] [Google Scholar]
- Berman M.G., Nee D.E., Casement M., Kim H.S., Deldin P., Kross E., Gonzalez R., Demiralp E., Gotlib I.H., Hamilton P., Joormann J., Waugh C., Jonides J. Neural and behavioral effects of interference resolution in depression and rumination. Cogn. Affect. Behav. Neurosci. 2011;11:85–96. doi: 10.3758/s13415-010-0014-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cavada C., Goldman-Rakic P.S. Posterior parietal cortex in rhesus monkey: II. Evidence for segregated corticocortical networks linking sensory and limbic areas with the frontal lobe. J. Comp. Neurol. 1989;287:422–445. doi: 10.1002/cne.902870403. [DOI] [PubMed] [Google Scholar]
- Chadick J.Z., Gazzaley A. Differential coupling of visual cortex with default or frontal-parietal network based on goals. Nat. Neurosci. 2011;14:830–832. doi: 10.1038/nn.2823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Channon S., Baker J.E., Robertson M.M. Working memory in clinical depression: an experimental study. Psychol. Med. 1993;23:87–91. doi: 10.1017/s0033291700038873. [DOI] [PubMed] [Google Scholar]
- Christophel T.B., Haynes J.D. Decoding complex flow-field patterns in visual working memory. NeuroImage. 2014;91:43–51. doi: 10.1016/j.neuroimage.2014.01.025. [DOI] [PubMed] [Google Scholar]
- Christophel T.B., Hebart M.N., Haynes J.-D. Decoding the contents of visual short-term memory from human visual and parietal cortex. J. Neurosci. 2012;32:12983–12989. doi: 10.1523/JNEUROSCI.0184-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooney R.E., Joormann J., Eugène F., Dennis E.L., Gotlib I.H. Neural correlates of rumination in depression. Cogn. Affect. Behav. Neurosci. 2010;10:470–478. doi: 10.3758/CABN.10.4.470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Costafreda S.G., Khanna A., Mourao-Miranda J., Fu C.H.Y. Neural correlates of sad faces predict clinical remission to cognitive behavioural therapy in depression. Neuroreport. 2009;20:637–641. doi: 10.1097/WNR.0b013e3283294159. [DOI] [PubMed] [Google Scholar]
- Davis R.N., Nolen-Hoeksema S. Cognitive inflexibility among ruminators and nonruminators. Cogn. Ther. Res. 2000;24:699–711. [Google Scholar]
- De Lissnyder E., Koster E.H.W., Raedt R. Emotional interference in working memory is related to rumination. Cogn. Ther. Res. 2011;36:348–357. [Google Scholar]
- Deen B., Koldewyn K., Kanwisher N., Saxe R. Functional organization of social perception and cognition in the superior temporal sulcus. Cereb. Cortex. 2015;25:4596–4609. doi: 10.1093/cercor/bhv111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demeyer I., De Lissnyder E., Koster E.H.W., De Raedt R. Rumination mediates the relationship between impaired cognitive control for emotional information and depressive symptoms: a prospective study in remitted depressed adults. Behav. Res. Ther. 2012;50:292–297. doi: 10.1016/j.brat.2012.02.012. [DOI] [PubMed] [Google Scholar]
- Desseilles M., Balteau E., Sterpenich V., Dang-Vu T.T., Darsaud A., Vandewalle G., Albouy G., Salmon E., Peters F., Schmidt C., Schabus M., Gais S., Degueldre C., Phillips C., Luxen A., Ansseau M., Maquet P., Schwartz S. Abnormal neural filtering of irrelevant visual information in depression. J. Neurosci. 2009;29:1395–1403. doi: 10.1523/JNEUROSCI.3341-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desseilles M., Schwartz S., Dang-Vu T.T., Sterpenich V., Ansseau M., Maquet P., Phillips C. Depression alters “top-down” visual attention: a dynamic causal modeling comparison between depressed and healthy subjects. NeuroImage. 2011;54:1662–1668. doi: 10.1016/j.neuroimage.2010.08.061. [DOI] [PubMed] [Google Scholar]
- Disner S.G., Beevers C.G., Haigh E.a.P., Beck A.T. Neural mechanisms of the cognitive model of depression. Nat. Rev. Neurosci. 2011;12:467–477. doi: 10.1038/nrn3027. [DOI] [PubMed] [Google Scholar]
- Epstein R., DeYoe E.a., Press D.Z., Rosen A.C., Kanwisher N. Neuropsychological evidence for a topographical learning mechanism in parahippocampal cortex. Cogn. Neuropsychol. 2001;18:481–508. doi: 10.1080/02643290125929. [DOI] [PubMed] [Google Scholar]
- Epstein R.a., Higgins J.S., Jablonski K., Feiler A.M. Visual scene processing in familiar and unfamiliar environments. J. Neurophysiol. 2007;97:3670–3683. doi: 10.1152/jn.00003.2007. [DOI] [PubMed] [Google Scholar]
- First M., Spitzer R., Gibbon M., Williams J. 2007. SCID-I/P. [Google Scholar]
- Fitzgerald P.B., Srithiran A., Benitez J., Daskalakis Z.Z., Oxley T.J., Kulkarni J., Egan G.F. An fMRI study of prefrontal brain activation during multiple tasks in patients with major depressive disorder. Hum. Brain Mapp. 2008;29:490–501. doi: 10.1002/hbm.20414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox C.J., Iaria G., Barton J.J.S. Defining the face processing network: optimization of the functional localizer in fMRI. Hum. Brain Mapp. 2009;30:1637–1651. doi: 10.1002/hbm.20630. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friston K.J., Buechel C., Fink G.R., Morris J., Rolls E., Dolan R.J. Psychophysiological and modulatory interactions in neuroimaging. NeuroImage. 1997;6:218–229. doi: 10.1006/nimg.1997.0291. [DOI] [PubMed] [Google Scholar]
- Fu C.H.Y., Williams S.C.R., Cleare A.J., Scott J., Mitterschiffthaler M.T., Walsh N.D., Donaldson C., Suckling J., Andrew C., Steiner H., Murray R.M. Neural responses to sad facial expressions in major depression following cognitive behavioral therapy. Biol. Psychiatry. 2008;64:505–512. doi: 10.1016/j.biopsych.2008.04.033. [DOI] [PubMed] [Google Scholar]
- Furey M.L., Drevets W.C., Hoffman E.M., Frankel E., Speer A.M., Zarate C.a. Potential of pretreatment neural activity in the visual cortex during emotional processing to predict treatment response to scopolamine in major depressive disorder. JAMA Psychiat. 2013;70:280–290. doi: 10.1001/2013.jamapsychiatry.60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garraux G. Shared brain areas but not functional connections controlling movement timing and order. J. Neurosci. 2005;25:5290–5297. doi: 10.1523/JNEUROSCI.0340-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrett A., Kelly R., Gomez R., Keller J., Schatzberg A.F., Reiss A.L. Aberrant brain activation during a working memory task in psychotic major depression. Am. J. Psychiatry. 2011;168:173–182. doi: 10.1176/appi.ajp.2010.09121718. [DOI] [PubMed] [Google Scholar]
- Gazzaley A., Cooney J.W., McEvoy K., Knight R.T., D'Esposito M. Top-down enhancement and suppression of the magnitude and speed of neural activity. J. Cogn. Neurosci. 2005;17:507–517. doi: 10.1162/0898929053279522. [DOI] [PubMed] [Google Scholar]
- Gazzaley A., Cooney J.W., Rissman J., D'Esposito M. Top-down suppression deficit underlies working memory impairment in normal aging. Nat. Neurosci. 2005;8:1298–1300. doi: 10.1038/nn1543. [DOI] [PubMed] [Google Scholar]
- Gazzaley A., Rissman J., Cooney J., Rutman A., Seibert T., Clapp W., D'Esposito M. Functional interactions between prefrontal and visual association cortex contribute to top-down modulation of visual processing. Cereb. Cortex. 2007;17(Suppl. 1):i125–i135. doi: 10.1093/cercor/bhm113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert C.D., Li W. Top-down influences on visual processing. Nat. Rev. Neurosci. 2013;14:350–363. doi: 10.1038/nrn3476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gohier B., Ferracci L., Surguladze S.a., Lawrence E., El Hage W., Kefi M.Z., Allain P., Garre J.-B., Le Gall D. Cognitive inhibition and working memory in unipolar depression. J. Affect. Disord. 2009;116:100–105. doi: 10.1016/j.jad.2008.10.028. [DOI] [PubMed] [Google Scholar]
- Gorgoraptis N., Catalao R.F.G., Bays P.M., Husain M. Dynamic updating of working memory resources for visual objects. J. Neurosci. 2011;31:8502–8511. doi: 10.1523/JNEUROSCI.0208-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gotlib I.H., Joormann J. Cognition and depression: current status and future directions. Annu. Rev. Clin. Psychol. 2010;6:285–312. doi: 10.1146/annurev.clinpsy.121208.131305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gotlib I.H., Krasnoperova E., Yue D.N., Joormann J. Attentional biases for negative interpersonal stimuli in clinical depression. J. Abnorm. Psychol. 2004;113:121–135. doi: 10.1037/0021-843X.113.1.121. [DOI] [PubMed] [Google Scholar]
- Green D.M., Swets J.a., Nevin J.a. Signal detection theory and operant behavior. J. Exp. Anal. Behav. 1969;3:475–480. [Google Scholar]
- Gruber O., Zilles D., Kennel J., Gruber E., Falkai P. A systematic experimental neuropsychological investigation of the functional integrity of working memory circuits in major depression. Eur. Arch. Psychiatry Clin. Neurosci. 2011;261:179–184. doi: 10.1007/s00406-010-0165-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hames J.L., Hagan C.R., Joiner T.E. Interpersonal processes in depression. Annu. Rev. Clin. Psychol. 2013;9:355–377. doi: 10.1146/annurev-clinpsy-050212-185553. [DOI] [PubMed] [Google Scholar]
- Han X., Berg A.C., Oh H., Samaras D., Leung H.-C. Multi-voxel pattern analysis of selective representation of visual working memory in ventral temporal and occipital regions. NeuroImage. 2013;73:8–15. doi: 10.1016/j.neuroimage.2013.01.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrison S.a., Tong F. Decoding reveals the contents of visual working memory in early visual areas. Nature. 2009;458:632–635. doi: 10.1038/nature07832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harvey P.-O., Fossati P., Pochon J.-B., Levy R., Lebastard G., Lehéricy S., Allilaire J.-F., Dubois B. Cognitive control and brain resources in major depression: an fMRI study using the n-back task. NeuroImage. 2005;26:860–869. doi: 10.1016/j.neuroimage.2005.02.048. [DOI] [PubMed] [Google Scholar]
- Harvey P.O., Le Bastard G., Pochon J.B., Levy R., Allilaire J.F., Dubois B., Fossati P. Executive functions and updating of the contents of working memory in unipolar depression. J. Psychiatr. Res. 2004;38:567–576. doi: 10.1016/j.jpsychires.2004.03.003. [DOI] [PubMed] [Google Scholar]
- Jonas J., Jacques C., Liu-shuang J., Brissart H., Colnat-coulbois S., Maillard L., Rossion B. A face-selective ventral occipito-temporal map of the human brain with intracerebral potentials. PNAS. 2016 doi: 10.1073/pnas.1522033113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Joormann J., Gotlib I.H. Updating the contents of working memory in depression: interference from irrelevant negative material. J. Abnorm. Psychol. 2008;117:182–192. doi: 10.1037/0021-843X.117.1.182. [DOI] [PubMed] [Google Scholar]
- Joormann J., Levens S.M., Gotlib I.H. Sticky thoughts: depression and rumination are associated with difficulties manipulating emotional material in working memory. Psychol. Sci. 2011;22:979–983. doi: 10.1177/0956797611415539. [DOI] [PubMed] [Google Scholar]
- Kanwisher N., McDermott J., Chun M.M. The fusiform face area: a module in human extrastriate cortex specialized for face perception. J. Neurosci. 1997;17:4302–4311. doi: 10.1523/JNEUROSCI.17-11-04302.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kastner S., Ungerleider L.G. Mechanisms of visual attention in the human cortex. Annu. Rev. Neurosci. 2000;23:315–341. doi: 10.1146/annurev.neuro.23.1.315. [DOI] [PubMed] [Google Scholar]
- Kawabata Duncan K.J., Devlin J.T. Improving the reliability of functional localizers. NeuroImage. 2011;57:1022–1030. doi: 10.1016/j.neuroimage.2011.05.009. [DOI] [PubMed] [Google Scholar]
- Kennedy S.H., Evans K.R., Krüger S., Mayberg H.S., Meyer J.H., McCann S., Arifuzzman a.I., Houle S., Vaccarino F.J. Changes in regional brain glucose metabolism measured with positron emission tomography after paroxetine treatment of major depression. Am. J. Psychiatry. 2001;158:899–905. doi: 10.1176/appi.ajp.158.6.899. [DOI] [PubMed] [Google Scholar]
- Korgaonkar M.S., Grieve S.M., Etkin A., Koslow S.H., Williams L.M. Using standardized fMRI protocols to identify patterns of prefrontal circuit dysregulation that are common and specific to cognitive and emotional tasks in major depressive disorder: first wave results from the iSPOT-D study. Neuropsychopharmacology. 2013;38:863–871. doi: 10.1038/npp.2012.252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landro N.I., Stiles T.C., Sletvold H. Neuropsychological function in nonpsychotic unipolar major depression. Neuropsychiatry Neuropsychol. Behav. Neurol. 2001;14:233–240. [PubMed] [Google Scholar]
- Lawson C., MacLeod C. Depression and the interpretation of ambiguity. Behav. Res. Ther. 1999;37:463–474. doi: 10.1016/s0005-7967(98)00131-4. [DOI] [PubMed] [Google Scholar]
- Leppänen J.M., Milders M., Bell J.S., Terriere E., Hietanen J.K. Depression biases the recognition of emotionally neutral faces. Psychiatry Res. 2004;128:123–133. doi: 10.1016/j.psychres.2004.05.020. [DOI] [PubMed] [Google Scholar]
- Levens S.M., Gotlib I.H. Updating positive and negative stimuli in working memory in depression. J. Exp. Psychol. Gen. 2010;139:654–664. doi: 10.1037/a0020283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao Y., Huang X., Wu Q., Yang C., Kuang W., Du M., Lui S., Yue Q., Chan R.C.K., Kemp G.J., Gong Q. Is depression a disconnection syndrome? Meta-analysis of diffusion tensor imaging studies in patients with MDD. J. Psychiatry Neurosci. 2013;38:49–56. doi: 10.1503/jpn.110180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundqvist D., Flykt A., Öhman A. CD ROM from Dep. Clin. Neurosci. Psychol. Sect. Karolinska Institutet; 1998. The Karolinska directed emotional faces (KDEF) [Google Scholar]
- MacQueen G.M., Tipper S.P., Young L.T., Joffe R.T., Levitt A.J. Impaired distractor inhibition on a selective attention task in unmedicated, depressed subjects. Psychol. Med. 2000;30:557–564. doi: 10.1017/s0033291799001981. [DOI] [PubMed] [Google Scholar]
- Marquand A.F., Mourão-Miranda J., Brammer M.J., Cleare A.J., Fu C.H.Y. Neuroanatomy of verbal working memory as a diagnostic biomarker for depression. Neuroreport. 2008;19:1507–1511. doi: 10.1097/WNR.0b013e328310425e. [DOI] [PubMed] [Google Scholar]
- McLaren D.G., Ries M.L., Xu G., Johnson S.C. A generalized form of context-dependent psychophysiological interactions (gPPI): a comparison to standard approaches. NeuroImage. 2012;61:1277–1286. doi: 10.1016/j.neuroimage.2012.03.068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meiran N., Diamond G.M., Toder D., Nemets B. Cognitive rigidity in unipolar depression and obsessive compulsive disorder: examination of task switching, Stroop, working memory updating and post-conflict adaptation. Psychiatry Res. 2011;185:149–156. doi: 10.1016/j.psychres.2010.04.044. [DOI] [PubMed] [Google Scholar]
- Minear M., Park D.C. A lifespan database of adult facial stimuli. Behav. Res. Methods Instrum. Comput. 2004;36:630–633. doi: 10.3758/bf03206543. [DOI] [PubMed] [Google Scholar]
- Nolen-Hoeksema S., Morrow J. A prospective study of depression and posttraumatic stress symptoms after a natural disaster: the 1989 Loma Prieta earthquake. J. Pers. Soc. Psychol. 1991;61:115–121. doi: 10.1037//0022-3514.61.1.115. [DOI] [PubMed] [Google Scholar]
- Nolen-Hoeksema S., Wisco B.E., Lyubomirsky S. Rethinking rumination. Perspect. Psychol. Sci. 2008;3:400–424. doi: 10.1111/j.1745-6924.2008.00088.x. [DOI] [PubMed] [Google Scholar]
- Oda K., Okubo Y., Ishida R., Murata Y., Ohta K., Matsuda T., Matsushima E., Ichimiya T., Suhara T., Shibuya H., Nishikawa T. Regional cerebral blood flow in depressed patients with white matter magnetic resonance hyperintensity. Biol. Psychiatry. 2003;53:150–156. doi: 10.1016/s0006-3223(02)01548-2. [DOI] [PubMed] [Google Scholar]
- Oh H., Leung H.-C. Specific and nonspecific neural activity during selective processing of visual representations in working memory. J. Cogn. Neurosci. 2010;22:292–306. doi: 10.1162/jocn.2009.21250. [DOI] [PubMed] [Google Scholar]
- O'Reilly R.C., Frank M.J. Making working memory work: a computational model of learning in the prefrontal cortex and basal ganglia. Neural Comput. 2004;18:283–328. doi: 10.1162/089976606775093909. [DOI] [PubMed] [Google Scholar]
- Pelosi L., Slade T., Blumhardt L.D., Sharma V.K. Working memory dysfunction in major depression: an event-related potential study. Clin. Neurophysiol. 2000;111:1531–1543. doi: 10.1016/s1388-2457(00)00354-0. [DOI] [PubMed] [Google Scholar]
- Peters J.C., Roelfsema P.R., Goebel R. Task-relevant and accessory items in working memory have opposite effects on activity in extrastriate cortex. J. Neurosci. 2012;32:17003–17011. doi: 10.1523/JNEUROSCI.0591-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pu S., Yamada T., Yokoyama K., Matsumura H., Kobayashi H., Sasaki N., Mitani H., Adachi A., Kaneko K., Nakagome K. A multi-channel near-infrared spectroscopy study of prefrontal cortex activation during working memory task in major depressive disorder. Neurosci. Res. 2011;70:91–97. doi: 10.1016/j.neures.2011.01.001. [DOI] [PubMed] [Google Scholar]
- Rush A.J., Gullion C.M., Basco M.R., Jarrett R.B., Trivedi M.H. The Inventory of Depressive Symptomatology (IDS): psychometric properties. Psychol. Med. 1996;26:477–486. doi: 10.1017/s0033291700035558. [DOI] [PubMed] [Google Scholar]
- Russell B.C., Torralba A., Murphy K.P., Freeman W.T. LabelMe: a database and web-based tool for image annotation. Int. J. Comput. Vis. 2007;77:157–173. [Google Scholar]
- Schall J.D., Morel a., King D.J., Bullier J. Topography of visual cortex connections with frontal eye field in macaque: convergence and segregation of processing streams. J. Neurosci. 1995;15:4464–4487. doi: 10.1523/JNEUROSCI.15-06-04464.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schöning S., Zwitserlood P., Engelien A., Behnken A., Kugel H., Schiffbauer H., Lipina K., Pachur C., Kersting A., Dannlowski U., Baune B.T., Zwanzger P., Reker T., Heindel W., Arolt V., Konrad C. Working-memory fMRI reveals cingulate hyperactivation in euthymic major depression. Hum. Brain Mapp. 2009;30:2746–2756. doi: 10.1002/hbm.20702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silton R.L., Heller W., Engels A.S., Towers D.N., Spielberg J.M., Edgar J.C., Sass S.M., Stewart J.L., Sutton B.P., Banich M.T., Miller G.A. Depression and anxious apprehension distinguish frontocingulate cortical activity during top-down attentional control. J. Abnorm. Psychol. 2011;120:272–285. doi: 10.1037/a0023204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stephan K.E., Marshall J.C., Friston K.J., Rowe J.B., Ritzl A., Zilles K., Fink G.R. Lateralized cognitive processes and lateralized task control in the human brain. Science. 2003;301:384–386. doi: 10.1126/science.1086025. [DOI] [PubMed] [Google Scholar]
- Suslow T., Konrad C., Kugel H., Rumstadt D., Zwitserlood P., Schöning S., Ohrmann P., Bauer J., Pyka M., Kersting A., Arolt V., Heindel W., Dannlowski U. Automatic mood-congruent amygdala responses to masked facial expressions in major depression. Biol. Psychiatry. 2010;67:155–160. doi: 10.1016/j.biopsych.2009.07.023. [DOI] [PubMed] [Google Scholar]
- Tomita H., Ohbayashi M., Nakahara K. Top-down signal from prefrontal cortex in executive control. Nature. 1999;401:699–703. doi: 10.1038/44372. [DOI] [PubMed] [Google Scholar]
- Treynor W., Gonzalez R., Nolen-Hoeksema S. Rumination reconsidered : a psychometric analysis. Cogn. Ther. Res. 2003;27:247–259. [Google Scholar]
- van Tol M.-J., Demenescu L.R., van der Wee N.J.a., Kortekaas R., Marjan M A N., Boer J.a.D., Renken R.J., van Buchem M.a., Zitman F.G., Aleman A., Veltman D.J. Functional magnetic resonance imaging correlates of emotional word encoding and recognition in depression and anxiety disorders. Biol. Psychiatry. 2012;71:593–602. doi: 10.1016/j.biopsych.2011.11.016. [DOI] [PubMed] [Google Scholar]
- Walsh N.D., Williams S.C.R., Brammer M.J., Bullmore E.T., Kim J., Suckling J., Mitterschiffthaler M.T., Cleare A.J., Pich E.M., Mehta M.a., Fu C.H.Y. A longitudinal functional magnetic resonance imaging study of verbal working memory in depression after antidepressant therapy. Biol. Psychiatry. 2007;62:1236–1243. doi: 10.1016/j.biopsych.2006.12.022. [DOI] [PubMed] [Google Scholar]
- Walter H., Wolf R.C., Spitzer M., Vasic N. Increased left prefrontal activation in patients with unipolar depression: an event-related, parametric, performance-controlled fMRI study. J. Affect. Disord. 2007;101:175–185. doi: 10.1016/j.jad.2006.11.017. [DOI] [PubMed] [Google Scholar]
- Walther D.B., Caddigan E., Fei-Fei L., Beck D.M. Natural scene categories revealed in distributed patterns of activity in the human brain. J. Neurosci. 2009;29:10573–10581. doi: 10.1523/JNEUROSCI.0559-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X., Zhen Z., Song Y., Huang L., Kong X., Liu J. The hierarchical structure of the face network revealed by its functional connectivity pattern. J. Neurosci. 2016;36:890–900. doi: 10.1523/JNEUROSCI.2789-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watkins E., Brown R.G. Rumination and executive function in depression: an experimental study. J. Neurol. Neurosurg. Psychiatry. 2002;72:400–402. doi: 10.1136/jnnp.72.3.400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitmer A.J., Banich M.T. Inhibition versus switching deficits in different forms of rumination. Psychol. Sci. 2007;18:546–553. doi: 10.1111/j.1467-9280.2007.01936.x. [DOI] [PubMed] [Google Scholar]
- Xing Y., Ledgeway T., McGraw P.V., Schluppeck D. Decoding working memory of stimulus contrast in early visual cortex. J. Neurosci. 2013;33:10301–10311. doi: 10.1523/JNEUROSCI.3754-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoon K.L., LeMoult J., Joormann J. Updating emotional content in working memory: a depression-specific deficit? J. Behav. Ther. Exp. Psychiatry. 2014;45:368–374. doi: 10.1016/j.jbtep.2014.03.004. [DOI] [PubMed] [Google Scholar]
- Zanto T.P., Rubens M.T., Thangavel A., Gazzaley A. Causal role of the prefrontal cortex in top-down modulation of visual processing and working memory. Nat. Neurosci. 2011;14:656–661. doi: 10.1038/nn.2773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng L.-L., Shen H., Liu L., Wang L., Li B., Fang P., Zhou Z., Li Y., Hu D. Identifying major depression using whole-brain functional connectivity: a multivariate pattern analysis. Brain. 2012;135:1498–1507. doi: 10.1093/brain/aws059. [DOI] [PubMed] [Google Scholar]
- Zhen Z., Yang Z., Huang L., Kong X.z., Wang X., Dang X., Huang Y., Song Y., Liu J. Quantifying interindividual variability and asymmetry of face-selective regions: a probabilistic functional atlas. NeuroImage. 2015;113:13–25. doi: 10.1016/j.neuroimage.2015.03.010. [DOI] [PubMed] [Google Scholar]
- Zokaei N., Gorgoraptis N., Bahrami B., Bays P.M., Husain M. Precision of working memory for visual motion sequences and transparent motion surfaces. J. Vis. 2011;11:1–18. doi: 10.1167/11.14.2. [DOI] [PMC free article] [PubMed] [Google Scholar]