

Abstract
There is currently no treatment for effectively slowing the progression of Alzheimer's disease, so early prevention is very important. Numerous studies have shown that flavonoids can improve memory impairment. The present study investigated the effects of myricetin, a member of the flavonoids, on intracerebroventricular streptozotocin induced neuronal loss and memory impairment in rat models of Alzheimer's disease. Myricetin at 5 or 10 mg/kg was intraperitoneally injected into rats over 21 days. Control rats were treated with 10 mL/kg saline. Behavioral test (the shuttle box test) was performed on day 22 to examine learning and memory in rats. Immediately after that, hematoxylin-eosin staining was performed to observe the morphological change in hippocampal CA3 pyramidal neurons. Myricetin greatly increased the number of hippocampal CA3 pyramidal neurons and improved learning and memory impairments in rats with Alzheimer's disease. These findings suggest that myricetin is beneficial for treatment of Alzheimer's disease.
Keywords: nerve regeneration, myricetin, Alzheimer's disease, streptozotocin, hippocampus, pyramidal neurons, CA3 region, behavioral test, neural regeneration
Introduction
Alzheimer's disease is one of the most prevalent types of dementia. It usually occurs in people aged 65 years or older (Newman et al., 2007). Both environmental and genetic factors are relevant in the pathogenesis of AD. The unusual deposition of amyloid-beta (Aβ) plaques and the abnormal aggregation of tau proteins are neuropathological properties of AD (Vauzour, 2014). Neuronal degeneration and cognitive impairment are the most typical features of this disorder (Song et al., 2014), which can directly affect the patient's ability to recall or recognize new information processed in the hippocampus (Machado et al., 2009). The hippocampus is a part of the brain consisting of CA1, CA2, and CA3 fields and the subiculum. Information originating from dentate gyrus is transferred to CA3 and then integrated in CA1 (Mu et al., 2011).
Strong evidence has recently come to light to support the positive influences of flavonoids on the brain (Spencer, 2007; Macready et al., 2009; Rendeiro et al., 2009; Kumar et al., 2013; Vauzour, 2014; Rendeiro et al., 2015; Wang et al., 2016). Flavonoids are rich in compounds that reduce neurodegenerative efficacy (Vauzour et al., 2008). In addition, they strongly promote neural networks by stimulating neurogenesis (Vauzour et al., 2008; Oberbauer et al., 2013). They have also been reported to enhance cognitive functions (Macready et al., 2009). Flavonoids exist in many vegetables and fruits and greatly influence different brain functions, such as cerebrovascular blood flow and synaptic plasticity, which are related to learning and memory (Rendeiro et al., 2015). Particularly, they are able to control and conserve neuronal survival, differentiation, and long-term potentiation in memory (Krishnaveni, 2012). The first and most important characteristics of flavonoids are their antioxidant activity and their ability to suppress reactive oxygen species formation (Kumar et al., 2013).
Numerous studies have confirmed neurogenesis in hippocampal neurons (Eriksson et al., 1998; Dong et al., 2003; Bruel-Jungerman et al., 2007; Liu et al., 2011; Mu et al., 2011). A large body of evidence has shown that flavonoids play a key role in neuronal improvement because of their high proportion of antioxidant features (Jung et al., 2010; Kim et al., 2010; Choi et al., 2012; Huang et al., 2012; Rendeiro et al., 2015; Wang et al., 2016). Some studies have also shown that flavonoids prevent and restrict neuronal damage in the hippocampal pyramidal layer (Rendeiro et al., 2009; Tongjaroenbuangam et al., 2011; Lee et al., 2012; Ashrafpour et al., 2015; Wang et al., 2016).
Myricetin (3,3′,4′5,5′,7-hexahydroxylflavone) is a common natural flavonoid found in many fruits, vegetables, and herbs. It is well-known for its antioxidant activity and anti-apoptotic effects (Kang et al., 2010). Myricetin plays a substantial role in the treatment and prevention of some diseases due to its potent iron-chelating capability, antioxidant and free-radical scavenging activities (Yao et al., 2014). Myricetin, as a well-known flavonoid with anti-oxidative, anti-inflammatory, anti-apoptotic, and neuroprotective properties, reportedly plays an important role in the hippocampus of stressed mice (Ma et al., 2015).
According to these studies, we assumed that myricetin as a potential flavonoid is beneficial for learning and memory in experimental models of Alzheimer's disease. In addition, it can improve neurons in hippocampal CA3 pyramidal layers in rats with Alzheimer's disease. The present study examined the possible effects of myricetin on learning and memory impairments and hippocampal CA3 pyramidal neurons in Wistar rat models of Alzheimer's disease induced by intracerebroventricular (i.c.v.) streptozotocin (STZ).
Materials and Methods
Animals
Fifty adult male Wistar rats, weighing 220–250 g, were provided by Pasteur Institute, Tehran, Iran. The rats were housed two per cage in an animal house, kept at 22 ± 2°C and a 12 hour dark/light cycle, and were given free access to food and water.
Rats were anesthetized by intraperitoneal (i.p.) injection of 10% ketamine (50 mg/kg) and 2% xylazine (5 mg/kg) (Struck et al., 2011) and all efforts were made to minimize the pain and distress of experimental animals. All procedures for the treatment of animals were reviewed and approved by the Research and Ethics Committee of the Biology School, University of Arak, Iran and performed according to the National Institute of Health Guidelines for the Care and Use of Laboratory Animals.
Treatment
All rats were equally and randomly divided into five groups. In the control group, rats daily received i.c.v. (10 μL for each lateral ventricle) and i.p. (10 mL/kg) injections of normal saline. In the STZ group, rats received injection of STZ (3 mg/kg, i.c.v., 10 μL on each side, on first and third days) under the stereotaxic surgery and normal saline (10 mL/kg, i.p.). In the STZ + 5 mg/kg myricetin and STZ + 10 mg/kg myricetin groups, rats received injection of STZ (3 mg/kg, i.c.v.) and myricetin (5 and 10 mg/kg respectively, i.p.). In the saline + myricetin group, rats received injection of normal saline (10 μL for each lateral ventricle, i.c.v.) and myricetin (10 mg/kg, i.p.). Myricetin/saline i.p. administration started 1 day before stereotactic surgery and was performed in the morning between 9:00 and 12:00 over 21 days. After 21 days, the passive avoidance test (the shuttle box test) was performed on the 22nd day. Immediately after that, rat brains were removed and stained with hematoxylin-eosin to examine the effects of myricetin on hippocampal CA3 pyramidal neurons.
Drugs
The drugs used in the study were STZ (Sigma-Aldrich, St. Louis, MO, USA) and myricetin (Sigma-Aldrich). They were dissolved in sterile 0.9% saline just before the experiment. STZ was administered into the lateral ventricle, and myricetin or saline was i.p. injected.
Stereotaxic surgery
After anesthesia induction, the animal's head was shaved and then placed into a stereotaxic instrument (Stoelting, Chicago, IL, USA). 10 μL of STZ (ICV) was injected into each lateral ventricle via a Hamilton syringe. The stereotaxic coordinates were chosen based on the rat brain atlas of Paxinos and Watson (1998): anteriorposterior = –0.8 mm; mediolateral = ± 1.4 mm; dorso-ventral = –3.5 mm from bregma.
Behavioral test
At the end of the treatment, the passive avoidance test (the shuttle box test) was performed. It consisted of two equally-sized compartments, a bright room (white compartment, 20 cm × 20 cm × 30 cm), and a dark one (black compartment, 20 cm × 20 cm × 30 cm). The chambers were separated by a guillotine-type door.
Prior to the experiment, all animals were placed into the apparatus for approximately 30 minutes (habituation stage). Afterward, each rat was individually placed in the bright chamber and the guillotine door was opened. As soon as the rat entered the dark chamber, the door was closed, and an electric shock was delivered by an isolated stimulator (1Hz, 3 seconds, 1/5 MA) through the grid floor. Twenty-four hours later, each rat was placed in the bright chamber for the retention test. The step-through latency (STL) and the total time spent in the dark chamber (TDC) were measured.
Tissue preparation and hematoxylin-eosin staining
Rats were deeply anesthetized with 3.5% chloral hydrate (35 mg/100 g, i.p.), and vessels were perfusion fixed by intracardiac infusion with phosphate buffer solution (0.1 M, pH 7.4) followed by 200 mL of 4% paraformaldehyde fixative in phosphate buffer solution (0.1 M, pH 7.4) for 15 minutes. Rat brains were removed, isolated, post fixed in 4% para formaldehyde (24 hours), and embedded in paraffin. Coronal sections, 10 μm in thickness, were taken from the dorsal hippocampus and stained with hematoxylin and eosin. Finally, the numbers of damaged and intact neurons in the hippocampal CA3 pyramidal layer were counted (Koike et al., 2008) using a light microscope (BX40, Olympus, New York, USA) connected to a camera (Olympus, DP12), and quantitatively analyzed by ImageJ software (Neuroscience Center of Shahid Beheshti University, Iran).
Statistical analysis
All results are expressed as the mean ± SEM. For the behavioral test, STL and TDC were analyzed by one-way analysis of variance (ANOVA) and Tukey's post-hoc test. Statistical results were analyzed using SPSS 16.0 software (SPSS, Chicago, IL, USA). Differences between groups in the average number of intact and damaged neurons in the hippocampal CA3 pyramidal layer were analyzed using GraphPad Prism Software (version 5.00) (GraphPad Software Inc., San Diego, CA, USA). A value of P < 0.05 was considered statistically significant.
Results
Effect of myricetin on cognitive function of rats with Alzheimer's disease
Passive avoidance test results showed that STL was decreased and TDC was increased in the STZ group, indicating memory impairment, than in the control group (P < 0.05). STL was significantly increased and TDC was significantly decreased in the STZ + 10 mg/kg myricetin group than in the STZ group (P < 0.01). There were no significant differences in STL and TDC between STZ + 5 mg/kg myricetin and STZ groups (P > 0.05). STL was increased and TDC was decreased in the saline + 10 mg/kg myricetin group than in the STZ group (Figure 1).
Figure 1.
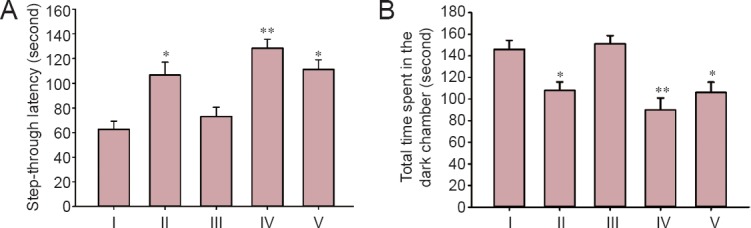
Effect of myricetin (5 and 10 mg/kg) on learning and memory of Alzheimer's disease rats measured by passive avoidance test (the shuttle box test).
(A, B) Step-through latency (A) and total time spent in the dark chamber (B) in passive avoidance test were measured after intracerebroventricular injection of STZ followed by 21-day treatment of myricetin. Values are expressed as the mean ± SEM with 10 rats in each group. *P < 0.05, **P < 0.01, vs. STZ group (one-way analysis of variance followed by Tukey's post-hoc test). I–V: STZ (intracerebroventricular injection of STZ + intraperitoneal injection of normal saline), control (intracerebroventricular and intraperitoneal injections of normal saline), STZ + 5 mg/kg myricetin (intracerebroventricular injection of STZ + intraperitoneal injection of 5 mg/kg myricetin), STZ +10 mg/kg myricetin (intracerebroventricular injection of STZ + intraperitoneal injection of 10 mg/kg myricetin), saline + 10 mg/kg myricetin (intracerebroventricular injection of normal saline + intraperitoneal injection of 10 mg/kg myricetin) groups, respectively. STZ: Streptozotocin.
Effect of myricetin on hippocampal CA3 pyramidal neuron density in rats with Alzheimer's disease
Histological studies revealed striking changes in neuron morphology in each group. The number of intact neurons in the hippocampal CA3 pyramidal layer was markedly decreased in the STZ group, and there were some irregularities in the layer; however, the number of intact neurons was significantly increased in the control group and saline + myricetin group. Despite the fact that STZ greatly damaged neurons, myricetin (10 mg/kg/d, i.p.) significantly increased the number of intact neurons containing large, round, and regular neurons (Figure 2A–E).
Figure 2.
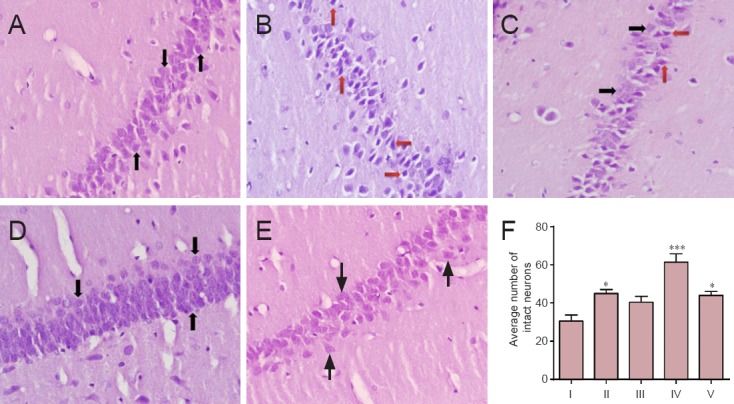
Effect of myricetin (5 and 10 mg/kg) on morphology of hippocampal CA3 pyramidal neurons of rats with Alzheimer's disease (hematoxylin-eosin staining, original magnification ×40).
(A) Control group (natural and intact neurons with arranged CA3 layer); (B) STZ group (significant neural loss and an irregular layer); (C) STZ + 5 mg/kg myricetin (most of the cells were damaged; however, there were some intact neurons); (D) STZ + 10 mg/kg myricetin (neurons were neatly arranged with a lot of intact and round cells); (E) saline +10 mg/kg myricetin (regular layer and significant increases in intact neuron). Black arrows indicate intact neurons and red arrows represent damaged neurons (F) The average number of intact neurons in hippocampal CA3 pyramidal layer. All data are represented as the mean ± SEM. *P < 0.05, ***P < 0.001, vs. STZ group (one-way analysis of variance followed by Tukey's post-hoc test). I–V: STZ (intracerebroventricular injection of STZ + intraperitoneal injection of normal saline), control (intracerebroventricular and intraperitoneal injections of normal saline), STZ + 5 mg/kg myricetin (intracerebroventricular injection of STZ + intraperitoneal injection of 5 mg/kg myricetin), STZ +10 mg/kg myricetin (intracerebroventricular injection of STZ + intraperitoneal injection of 10 mg/kg myricetin), saline + 10 mg/kg myricetin (intracerebroventricular injection of normal saline + intraperitoneal injection of 10 mg/kg myricetin) groups, respectively. STZ: Streptozotocin.
Although i.c.v. injection of STZ resulted in severe hippocampal CA3 pyramidal neuronal damage, saline did not lead to any severe neuronal damage in this area in control rats; the number of damaged neurons in the STZ group was significantly increased than that in the control group (P < 0.05). However, the number of damaged neurons was significantly decreased in the STZ + 10 mg/kg myricetin group than in the STZ group (P < 0.01) (Figure 2F). These results suggest that myricetin (10 mg/kg/d) exhibited significant protective effects on STZ-induced neuronal damage.
Discussion
The hippocampus plays a vital role in learning and memory. Moreover, hippocampal CA3 pyramidal neurons are significantly vulnerable to the negative effects of STZ. Therefore, hippocampal CA3 neurons located near the DG were the target of the current study. Hematoxylin-eosin staining showed that the number of intact neurons was greatly decreased in the STZ group; however, myricetin (10 mg/kg) alleviated neuronal damage in hippocampal CA3 region of rats receiving i.p. injection of STZ. This result indicates that myricetin at 10 mg/kg/d can promote neuronal recovery and at the same time improve neurogenesis in the hippocampus. i.c.v. injection of STZ in rats lead to learning, memory and cognitive impairments and worsened inflammatory response in the central nervous system (Baluchnejadmojarad., 2009; Khalili et al., 2010). The underlying mechanism remains unclear; however, it is likely attributed to the peripheral performance in the body (Grunblatt et al., 2007).
STZ in rat brain promoted inflammatory response after 3 weeks (Chu et al., 2005; Lester-Coll et al., 2006). Following 3 weeks of i.c.v. injection of STZ, great changes were observed in the rat brain (Agrawal et al., 2009). Free radicals induce neurotoxicity. Moreover, they lead to the symptoms of Alzheimer's disease (Khalili et al., 2010). There is evidence that STZ injections into the brain induce insulin resistance and lead to an increase in amyloid precursor protein processing to produce beta-amyloid and tau pathology (Kamat, 2015). Tau protein and Aβ are the most important pathological properties of AD. They are responsible for upcoming pathological events that result in a lack of cognitive performance, neuropsychiatric alterations, and eventually lead to neuronal death (Kamat, 2015).
Flavonoids can reportedly affect different kinds of synaptic plasticity, regulate receptor activation, signaling pathways and gene expression, and promote long-term potentiation (Rendeiro et al., 2012). In spite of many studies reporting flavonoid-mediated neuroprotection, there is no enough information regarding the interaction of flavonoids or their circulating metabolites with the blood-brain barrier (Youdim et al., 2004). Yao et al. (2014) demonstrated that myricetin is a lipophilic compound with low water solubility but high solubility in organic solvents or with solubilizers. It can pass through biological membranes such as the blood-brain barrier. Moreover, they show strong neuroprotective effects under in vivo conditions (Das et al., 2008; Hamaguchi et al., 2009), and their metabolites have been found in the brain tissue of rodents after oral administration (Paulke et al., 2006). These results display their ability to cross the blood-brain barrier (Paulke et al., 2006; Franco et al., 2010). In some age-related diseases, oxidative stress may contribute to the breakdown of blood-brain barrier, so flavonoids can enter the brain more easily than under the healthy state (Youdim et al., 2004).
It is strongly believed that Alzheimer's disease and other neurodegenerative disorders are triggered by multifactorial processes, including neuroinflammation, glutamatergic excitotoxicity, increases in iron, and/or the depletion of endogenous antioxidants (Jellinger, 2001). Flavonoids can inhibit apoptosis and other damage in neurons through neurotoxic species, but not directly. They modulate a number of protein kinase and lipid kinase signaling cascades, like PI3 kinase (PI3K)/Akt, tyrosine kinase, protein kinase C (PKC), and MAPK signaling pathways (Spencer, 2009). Spencer (2009) reported that flavonoids induce angiogenesis and new nerve cell growth in the hippocampus. Under the destructive effects of STZ, microglia and astrocytes produce cytokines (interleukin-1β, tumor necrosis factor-α) and nitric oxide (NO•) that lead to glial-induced neuronal death. Flavonoids play a truly significant role in the inhibition of NO•, interleukin-1β, and TNF-α in activated microglial cells (Spencer, 2009). Flavones have been shown to significantly inhibit 6-hydroxydopamine-induced JNK activation and neuronal cell death, and they may suppress JNK activity and apoptosis induced by hydrogen peroxide, 4-hydroxy-2-nonenal, and tumor necrosis factor-α (Kobuchi et al., 1999).
The present study clearly demonstrated that myricetin (10 mg/kg/d; i.p.) significantly inhibited cognitive impairments caused by i.c.v. injection of STZ. It can also increase the survival of hippocampal CA3 pyramidal neurons. The present findings add evidence to treatment of Alzheimer's disease using myricetin as a flavonoid.
Footnotes
Funding: Funding: The study was funded by the Department of Biology, Faculty of Science, Arak University, Iran, No. 38156-8-8349.
Conflicts of interest: None declared.
Plagiarism check: This paper was screened twice using CrossCheck to verify originality before publication.
Peer review: This paper was double-blinded and stringently reviewed by international expert reviewers.
Copyedited by Li CH, Song LP, Zhao M
References
- Agrawal R, Tyagi E, Shukla R, Nath C. A study of brain insulin receptors, AChE activity and oxidative stress in rat model of ICV STZ induced dementia. Neuropharmacology. 2009;56:779–787. doi: 10.1016/j.neuropharm.2009.01.005. [DOI] [PubMed] [Google Scholar]
- Ashrafpour M, Parsaei S, Sepehri H. Quercetin improved spatial memory dysfunctions in rat model of intracerebroventricular streptozotocin-induced sporadic Alzheimer's disease. Natl J Physiol. 2015;5:411–415. [Google Scholar]
- Baluchnejadmojarad T. The effect of genistein on intracerebroventricular streptozotocin-induced cognitive deficits in male rat. Basic Clin Neurosci. 2009;1:17–21. [Google Scholar]
- Bruel-Jungerman E, Rampon C, Laroche S. Adult hippocampal neurogenesis, synaptic plasticity and memory: facts and hypotheses. Rev Neurosci. 2007;18:93–114. doi: 10.1515/revneuro.2007.18.2.93. [DOI] [PubMed] [Google Scholar]
- Choi GN, Kim JH, Kwak JH, Jeong CH, Jeong HR, Lee U, Heo HJ. Effect of quercetin on learning and memory performance in ICR mice under neurotoxic trimethyltin exposure. Food Chem. 2012;132:1019–1024. [Google Scholar]
- Chu WZ, Qian CY. Expressions of Abeta1-40, Abeta1-42, tau202, tau396 and tau404 after intracerebroventricular injection of streptozotocin in rats. Di Yi Jun Yi Da Xue Xue Bao. 2005;25:168–170. [PubMed] [Google Scholar]
- Das S, Mandal AK, Ghosh A, Panda S, Das N, Sarkar S. Nanoparticulated quercetin in combating age related cerebral oxidative injury. Curr Aging Sci. 2008;1:169–174. doi: 10.2174/1874609810801030169. [DOI] [PubMed] [Google Scholar]
- Dong H, Csernansky CA, Goico B, Csernansky JG. Hippocampal neurogenesis follows kainic acid-induced apoptosis in neonatal rats. J Neurosci. 2003;23:1742–1749. doi: 10.1523/JNEUROSCI.23-05-01742.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eriksson PS, Perfilieva E, Björk-Eriksson T, Alborn AM, Nordborg C, Peterson DA, Gage FH. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4:1313–1317. doi: 10.1038/3305. [DOI] [PubMed] [Google Scholar]
- Francoa JL, Posser T, Missau F, Pizzolatti MG, Santos A, Souza DO, Aschner M, Rocha JBT, Dafre AL, Farina M. Structure-activity relationship of flavonoids derived from medicinal plants in preventing methylmercury-induced mitochondrial dysfunction. Environ Toxicol Pharmacol. 2010;30:272–278. doi: 10.1016/j.etap.2010.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grunblatt E, Salkovic-Petrisic M, Osmanovic J, Riederer P, Hoyer S. Brain insulin system dysfunction in streptozotocin intracerebroventricularly treated rats generates hyperphosphorylated tau protein. J Neurochem. 2007;101:757–770. doi: 10.1111/j.1471-4159.2006.04368.x. [DOI] [PubMed] [Google Scholar]
- Hamaguchi T, Ono K, Murase A, Yamada M. Phenolic compounds prevent Alzheimer's pathology through different effects on the amyloid-beta aggregation pathway. Am J Pathol. 2009;175:2557–2565. doi: 10.2353/ajpath.2009.090417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang JJ, Liu X, Wang XQ, Yang LH, Qi DS, Yao RQ. Effects of quercetin on the learning and memory ability of neonatal rats with hypoxic-ischemic brain damage. Zhongguo Dang Dai Er Ke Za Zhi. 2012;14:454–457. [PubMed] [Google Scholar]
- Jellinger KA. Cell death mechanisms in neurodegeneration. J Cell Mol Med. 2001;5:1–17. doi: 10.1111/j.1582-4934.2001.tb00134.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jung WY, Park SJ, Park DH, Kim JM, Kim DH, Ryu JH. Quercetin impairs learning and memory in normal mice via suppression of hippocampal phosphorylated cyclic AMP response element-binding protein expression. Toxicol Lett. 2010;197:97–105. doi: 10.1016/j.toxlet.2010.05.001. [DOI] [PubMed] [Google Scholar]
- Kamat PK. Streptozotocin induced Alzheimer's disease like changes and the underlying neural degeneration and regeneration mechanism. Neural Regen Res. 2015;10:1050–1052. doi: 10.4103/1673-5374.160076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang KA, Wang ZH, Zhang R, Piao MJ, Kim KC, Kang SS, Kim YW, Lee J, Park D, Hyun JW. Myricetin protects cells against oxidative stress-induced apoptosis via regulation of PI3K/Akt and MAPK signaling pathways. Int J Mol Sci. 2010;11:4348–4360. doi: 10.3390/ijms11114348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khalili M, Hamzeh F. Effects of active constituents of Crocus sativus L., crocin on streptozocin-induced model of sporadic Alzheimer's disease in male rats. Iran Biomed J. 2010;14:59–65. [PMC free article] [PubMed] [Google Scholar]
- Kim JK, Choi SJ, Cho HY, Hwang HJ, Kim YJ, Lim ST, Kim CJ, Kim HK, Peterson S, Shin DH. Protective effects of kaempferol (3,4’,5,7-tetrahydroxyflavone) against amyloid beta peptide (Aβ)-induced neurotoxicity in ICR mice. Biosci Biotechnol Biochem. 2010;74:397–401. doi: 10.1271/bbb.90585. [DOI] [PubMed] [Google Scholar]
- Kobuchi H, Roy S, Sen CK, Nguyen HG, Packer L. Quercetin inhibits inducible ICAM-1 expression in human endothelial cells through the JNK pathway. Am J Physiol. 1999;277:403–411. doi: 10.1152/ajpcell.1999.277.3.C403. [DOI] [PubMed] [Google Scholar]
- Koike M, Shibata M, Tadakoshi M, Gotoh K, Komatsu M, Waguri S, Kawahara N, Kuida K, Nagata S, Kominami E, Tanaka K, Uchiyama Y. Inhibition of autophagy prevents hippocampal pyramidal neuron death after hypoxic-ischemic injury. Am J Pathol. 2008;172:454–469. doi: 10.2353/ajpath.2008.070876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krishnaveni M. Flavonoid in enhancing memory function. J Pharm Res. 2012;5:3870–3874. [Google Scholar]
- Kumar S, Pandey AK. Chemistry and biological activities of flavonoids: an overview. ScientificWorldJournal 2013. 2013:162750. doi: 10.1155/2013/162750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee JH, Lee SR. The effect of Baicalein on hippocampal neuronal damage and metalloproteinase activity following transient global cerebral ischaemia. Phytother Res. 2012;26:1614–1619. doi: 10.1002/ptr.4644. [DOI] [PubMed] [Google Scholar]
- Lester-Coll N, Rivera EJ, Soscia SJ, Doiron K, Wands JR, de la Monte SM. Intracerebroventricular streptozotocin model of type 3 diabetes relevance to sporadic Alzheimer's disease. J Alzheimers Dis. 2006;9:13–33. doi: 10.3233/jad-2006-9102. [DOI] [PubMed] [Google Scholar]
- Liu JX, Pinnock SB, Herbert J. Novel control by the CA3 region of the hippocampus on neurogenesis in the dentate gyrus of the adult rat. PLoS One. 2011;6:1–8. doi: 10.1371/journal.pone.0017562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma Z, Wang G, Cui L, Wang Q. Myricetin attenuates depressant-like behavior in mice subjected to repeated restraint stress. Int J Mol Sci. 2015;16:28377–28385. doi: 10.3390/ijms161226102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Machado S, Cunha M, Minc D, Portella CE, Velasques B, Basile LF, Cagy M, Piedade R, Ribeiro P. Alzheimer's disease and implicit memory. Arq Neuropsiquiatr. 2009;67(2-A):334–342. doi: 10.1590/s0004-282x2009000200034. [DOI] [PubMed] [Google Scholar]
- Macready AL, Kennedy OB, Ellis JA, Williams CM, Spencer JP, Butler LT. Flavonoids and cognitive function: a review of human randomized controlled trial studies and recommendations for future studies. Genes Nutr. 2009;4:227–242. doi: 10.1007/s12263-009-0135-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mu Y, Gage FH. Adult hippocampal neurogenesis and its role in Alzheimer's disease. Mol Neurodegener. 2011;6:85. doi: 10.1186/1750-1326-6-85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newman M, Musgrave IF, Lardelli M. Alzheimer disease: amyloidogenesis, the presenilins and animal models. Biochim Biophys Acta. 2007;1772:285–297. doi: 10.1016/j.bbadis.2006.12.001. [DOI] [PubMed] [Google Scholar]
- Oberbauer E, Urmann C, Steffenhagen C, Bieler L, Brunner D, Furtner T, Humpel C, Bäumer B, Bandtlow C, Couillard-Despres S, Rivera FJ, Riepl H, Aigner L. Chroman-like cyclic prenylflavonoids promote neuronal differentiation and neurite outgrowth and are neuroprotective. J Nutr Biochem. 2013;24:1953–1962. doi: 10.1016/j.jnutbio.2013.06.005. [DOI] [PubMed] [Google Scholar]
- Paulke A, Schubert-Zsilavecz M, Wurglics M. Determination of St. John's wort flavonoid-metabolites in rat brain through high performance liquid chromatography coupled with fluorescence detection. J Chromatogr B Analyt Technol Biomed Life Sci. 2006;832:109–113. doi: 10.1016/j.jchromb.2005.12.043. [DOI] [PubMed] [Google Scholar]
- Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 4th edition. San Diego: Academic Press; 1998. [Google Scholar]
- Rendeiro C, Rhodes JS, Spencer JP. The mechanisms of action of flavonoids in the brain: Direct versus indirect effects. Neurochem Int. 2015;89:126–139. doi: 10.1016/j.neuint.2015.08.002. [DOI] [PubMed] [Google Scholar]
- Rendeiro C, Guerreiro JD, Williams CM, Spencer JP. Flavonoids as modulators of memory and learning: molecular interactions resulting in behavioural effects. Proc Nutr Soc. 2012;71:246–262. doi: 10.1017/S0029665112000146. [DOI] [PubMed] [Google Scholar]
- Rendeiro C, Spencer JP, Vauzour D, Butler LT, Ellis JA, Williams CM. The impact of flavonoids on spatial memory in rodents: from behaviour to underlying hippocampal mechanisms. Genes Nutr. 2009;4:251–270. doi: 10.1007/s12263-009-0137-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Song J, Hur BE, Bokara KK, Yang W, Cho HJ, Park KA, Lee WT, Lee KM, Lee JE. Agmatine improves cognitive dysfunction and prevents cell death in a streptozotocin-induced Alzheimer rat model. Yonsei Med J. 2014;55:689–699. doi: 10.3349/ymj.2014.55.3.689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spencer JP. Flavonoids and brain health: multiple effects underpinned by common mechanisms. Genes Nutr. 2009;4:243–250. doi: 10.1007/s12263-009-0136-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spencer JP. The interactions of flavonoids within neuronal signalling pathways. Genes Nutr. 2007;2:257–273. doi: 10.1007/s12263-007-0056-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Struck MB, Andrutis KA, Ramirez HE, Battles AH. Effect of a short-term fast on ketamine-xylazine anesthesia in rats. J Am Assoc Lab Anim Sci. 2011;50:344–348. [PMC free article] [PubMed] [Google Scholar]
- Tongjaroenbuangam W, Ruksee N, Chantiratikul P, Pakdeenarong N, Kongbuntad W, Govitrapong P. Neuroprotective effects of quercetin, rutin and okra (Abelmoschus esculentus Linn.) in dexamethasone-treated mice. Neurochem Int. 2011;59:677–685. doi: 10.1016/j.neuint.2011.06.014. [DOI] [PubMed] [Google Scholar]
- Vauzour D. Effect of flavonoids on learning, memory and neurocognitive performance: relevance and potential implications for Alzheimer's disease pathophysiology. J Sci Food Agric. 2014;94:1042–1056. doi: 10.1002/jsfa.6473. [DOI] [PubMed] [Google Scholar]
- Vauzour D, Vafeiadou K, Rodriguez-Mateos A, Rendeiro C, Spencer JP. The neuroprotective potential of flavonoids: a multiplicity of effects. Genes Nutr. 2008;3:115–126. doi: 10.1007/s12263-008-0091-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang H, Wang H, Cheng H, Che Z. Ameliorating effect of luteolin on memory impairment in an Alzheimer's disease model. Mol Med Rep. 2016;13:4215–4220. doi: 10.3892/mmr.2016.5052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yao Y, Lin G, Xie Y, Ma P, Li G, Meng Q, Wu T. Preformulation studies of myricetin: a natural antioxidant flavonoid. Pharmazie. 2014;69:19–26. [PubMed] [Google Scholar]
- Youdim KA, Shukitt-Hale B, Joseph JA. Flavonoids and the brain: interactions at the blood-brain barrier and their physiological effects on the central nervous system. Free Radic Biol Med. 2004;37:1683–1693. doi: 10.1016/j.freeradbiomed.2004.08.002. [DOI] [PubMed] [Google Scholar]