

Abstract
Using fMRI, we compared the patterns of fusiform activity produced by viewing English and Chinese for readers who were either English speakers learning Chinese or Chinese‐English bilinguals. The pattern of fusiform activity depended on both the writing system and the reader's native language. Native Chinese speakers fluent in English recruited bilateral fusiform areas when viewing both Chinese and English. English speakers learning Chinese, however, used heavily left‐lateralized fusiform regions when viewing English, but recruited an additional right fusiform region for viewing Chinese. Thus, English learners of Chinese show an accommodation pattern, in which the reading network accommodates the new writing system by adding neural resources that support its specific graphic requirements. Chinese speakers show an assimilation pattern, in which the reading network established for L1 includes procedures sufficient for the graphic demands of L2 without major change. Hum Brain Mapp, 2009. © 2008 Wiley‐Liss, Inc.
Keywords: Chinese word identification, fusiform, language learning, reading, writing systems
INTRODUCTION
Every writing system links print to spoken language, but there are differences in how writing systems make these links. Because of this, reading in two highly contrastive writing systems such as Chinese and English is likely to involve both overlapping and distinct processes. We can exploit differences between the writing systems to ask questions about which aspects of reading might be universal across even highly dissimilar languages, and which are language‐specific, and we can use functional imaging to discover similarities and differences in the neural circuits involved in reading in the two languages [Perfetti, 2003; Tan et al., 2001, 2005]. Our specific questions go beyond this mapping of differences and similarities across languages to questions of learning: Is the neural support for learning to read in a second writing system affected by the structure of this second system? Is it affected by the native language and its writing system?
Reading must begin with the visual analysis of text, so the visual and orthographic properties of a writing system should influence the early stages of word recognition. As an alphabetic system, English word forms are ordered strings of letters that correspond to ordered strings of meaningless sounds (phonemes), and reading words implies a process that uses knowledge of phonemes. In fact, success in alphabetic reading is highly correlated with phonological skills [Wagner and Torgesen, 1987; Wagner et al., 1997], while dyslexia is associated with a deficit in such skills [Rack et al., 1992]. Chinese, in contrast, is a morphosyllabic writing system, with most characters representing a meaning‐bearing syllable with no graphs that correspond to phonemes. Characters include component radicals, and radicals further comprise a set of strokes. In many instances the component radicals can stand alone as characters, and are thus associated with a meaning and pronunciation that is often related to the meaning or pronunciation of the whole character. Chen et al. [1996] argue that the basic functional orthographic units in Chinese word recognition are stroke patterns (lexical radicals and nonradical components) that recur across characters and cannot be further reduced into independently occurring combinations of strokes. Identification of a Chinese character involves recognizing these stroke patterns as well as their positional hierarchical organization within the resulting square character. Although phonological skill has also been linked to success in Chinese reading [So and Siegel, 1997], and phonology is activated rapidly in character identification [Lam et al., 1991; Perfetti and Zhang, 1991; Spinks et al., 2000; Tan and Perfetti, 1998;Tan et al., 1996], the mapping of orthography to phonology occurs at the level of the syllable rather than the phoneme. The theoretical and empirical consequences of this difference are detailed in Perfetti et al. [2005].
Given such differences, we might expect to see corresponding differences in the neural circuits supporting reading in the two writing systems, but the literature addressing this issue presents conflicting findings. While Tan et al. [2005] report both overlapping English/Chinese reading regions and more language‐specific regions in their meta‐analysis, others have found fairly complete overlap between regions used for reading English and Chinese [Chee et al., 1999a, c, 2000, 2001]. There are two main differences between these reports: (1) Tan et al. look specifically at phonological tasks, while Chee et al. use semantic tasks, and (2) Tan and his colleagues compare activity in native Chinese readers to activity in native English readers, while Chee and his colleagues report activity for both languages within proficient Chinese‐English bilinguals. The differences between these studies may afford a way to reconcile their seemingly discrepant results. It is possible that English and Chinese use similar regions for accessing meaning, but different areas for accessing phonology. Alternatively, it is possible that Chinese‐English bilinguals use overlapping regions for reading in the two languages, but that the regions may not be the same ones recruited in monolingual speakers of either language because the reading network may be influenced by experience with a native language.
One specific area of disagreement is whether the left middle frontal gyrus (BA 9) is an area of overlap between Chinese and English. Tan et al. [2001, 2005] reported the left middle frontal region to be significantly more active in reading Chinese than reading English, and Kuo et al. [2001] found it as a key area involved in the silent reading of Chinese that has “not been identified as critically linked to reading in previous studies on alphabetic languages.” In contrast, Chee et al. [1999a, 2000, 2001] also find BA 9 activation, but they find it for both English and Chinese. Either BA 9 could be active when Chinese‐English bilinguals read English, but not when English monolinguals read English, or an alternative would be that BA 9 is used in English semantic, but not phonological, tasks. A study by Chee et al. [1999b] identified regions involved in semantic processing in native English readers by having participants make abstract/concrete classifications. In the visual/reading version of this task, semantic judgments were compared to case judgments as a control. Neither the semantic judgment vs. fixation nor the semantic judgment vs. case judgment revealed involvement of BA 9. This leads us to believe that Chinese‐English bilinguals recruit different brain regions for reading English than native English readers, and that these regions overlap with regions used for reading Chinese. Further support for this comes from a study that directly compared activity in Chinese‐English bilinguals and monolingual English speakers in an English rhyme decision task; left middle frontal activity was observed only for the Chinese‐English bilinguals [Tan et al., 2003].
Since one of the major differences between Chinese and English is the orthographic mapping principal used, we can also look to activity in visual word recognition areas for possible processing differences. One area that shows consistent activation across many languages and word reading tasks is the middle fusiform region of the left ventral occipitotemporal cortex [Bolger et al., 2005]. This region is considered to be vital to the visual recognition of words due to its location along the visual processing stream, its sensitivity to orthographic legality, and its invariant response to differing visual features such as font [McCandliss et al., 2003]. The region has been found across studies of Chinese, Japanese Kana and Kanji, English, and other Western/alphabetic writing systems, and it is generally thought to be left‐lateralized [Bolger et al., 2005]. In adult readers of English, activity in the right hemisphere fusiform is weaker and more variable than the left hemisphere fusiform region [Dehaene et al., 2002], and sensitive in only some individuals to alphabetic stimuli [Cohen et al., 2002]. Part of the reason for the variability in right hemisphere activity may be due to baselines that specifically activate the right hemisphere fusiform to varying degrees, making a subtraction falsely inflate left‐lateralization [Ben‐Shachar et al., 2006]. However, left‐lateralization is also seen in reading when fixation is used as the baseline. Ben‐Shachar et al. [2006] argue that this may be due to feedback from language areas inflating left‐hemisphere activity during reading tasks when compared to the within‐word visual search task they used to find bilateral fusiform activity. Regardless of whether left hemisphere dominance is caused by feed‐forward activation or feedback, English reading tasks seem to activate the left hemisphere fusiform more consistently and robustly than the right.
In Chinese reading, however, activity in the right fusiform gyrus is consistent and highly convergent, as reported in two meta‐analyses of Chinese character reading [Bolger et al., 2005; Tan et al., 2005]. Kuo et al. [2001] also found that bilateral fusiform activation withstands the use of a stricter visuomotor control condition that eliminates many other right hemisphere regions thought to be involved in Chinese reading. We focus on this bilateral ventral occipitotemporal region in our study because of its important function in the early visual processing of words and its stability across reading tasks. The early visual processing is associated with a difference in laterality between Chinese, which shows bilateral activation, and English, which is mostly localized to a left hemisphere region, with more variable and generally weaker right‐hemisphere activity.
Given that there are different processing demands for reading Chinese and English, and that these differences are reflected in the neural activity seen in the ventral visual processing stream [Liu and Perfetti, 2003] and the left middle frontal region, how does one adapt to learning a new writing system once native reading networks are in place? We sought to distinguish between two broad possibilities about the outcome of learning a new writing system, which we refer to as the System Accommodation Hypothesis and the System Assimilation Hypothesis. The System Accommodation Hypothesis predicts that the brain must accommodate the new language by recruiting neural structures that can support the specific processing demands of the new writing system [Perfetti and Liu, 2005]. A high degree of accommodation would involve recruitment of entirely new areas for processing the new writing system, but accommodation could also simply amplify the activity in an area less important for reading in the native language. In general, this can be thought of as a qualitative change in the pattern of activity. Alternatively, the System Assimilation Hypothesis predicts that the brain will assimilate the new writing system, using neural circuits that are already in place for the native language to process the new writing system. A high degree of assimilation would result in no structural or functional differences between processing the two languages, but a pattern of assimilation could also describe overlapping brain structures being used for slightly different functional processes in the two languages.
Previous studies of Chinese‐English bilinguals suggest that when Chinese and English are learned together, overlapping brain structures are used to process the two languages. This network of regions is more similar to that of native Chinese speakers than native English speakers as it includes the left middle frontal gyrus (BA 9) [Chee et al., 1999a, 2000, 2001, 2003; Tan et al., 2003] and bilateral fusiform regions [Xue et al., 2004]. Additionally, the overlap for Chinese speakers in processing of Chinese and English occurs regardless of age of acquisition [Chee et al., 1999c] and proficiency [Chee et al., 2001], including low proficiency students [Xue et al., 2004]. These results have been used to make generalizations about second language learning and bilingualism, but none of these studies address whether the pattern of assimilation holds true for native speakers of English with Chinese as a second language. Given that native speakers of English show a different pattern of activity when reading English than Chinese/English bilinguals, it is an open question as to whether learners of Chinese will utilize the L1 (English) systems for Chinese, or whether they will use the more native Chinese‐like pattern of regions, including the right fusiform and left middle frontal gyrus.
Using a laboratory learning task in which native English speakers learned to read a set of Chinese characters, Liu et al. [2006] showed evidence of accommodation. Native English speakers, even after only a relatively short learning period, utilized the right hemisphere fusiform region and bilateral middle frontal regions. Since the pattern of activity was similar to that of native Chinese speakers reading Chinese, the study raises the question of whether the features of some writing systems may determine the necessary network required to read a language, regardless of native language.
To explore this question, we studied both native Chinese speakers who were also proficient in English (Chinese bilinguals) and native English speakers who had completed a year of college‐level Chinese instruction. With this design, we were able to look for evidence of a high degree of accommodation in either direction (with English or Chinese as the second language), and to extend the laboratory learning results to a more proficient population who had learned their second language outside of the lab. We mapped participants' brain activity while they passively viewed Chinese and English stimuli. We focused on activity in the ventral visual processing stream, primarily in bilateral middle fusiform regions, as well as the left middle frontal gyrus, to look for evidence of assimilation or accommodation of second language (L2) writing systems. If the brain areas for a reader's L2 overlap highly with those for that reader's native language (L1), there is evidence that his brain's existing reading network is assimilating the new writing system. If the brain areas for the reader's L2 instead overlap with those seen for a native speaker of L2, there is evidence that his brain's reading network has changed to accommodate the new writing system.
METHODS
Subjects
Twenty‐five healthy volunteers were recruited to participate for payment. Seventeen were native speakers of Chinese who were proficient in English as a second language. These Chinese participants were recruited through the University of Pittsburgh Chinese Student Association. All had TOEFL scores of at least 550, as required for acceptance to the University. Eight volunteers were native speakers of English who had completed 1 year of Chinese instruction at the University of Pittsburgh. These participants were recruited through the second semester Chinese classes. Data from four participants were eliminated due to excessive head motion, one participant withdrew from the study, and three participants were eliminated due to technical errors with the data acquisition. The remaining groups used in analysis consisted of 11 native Chinese speakers proficient in reading English (6 females, 5 males) and six native English speakers who had completed 1 year of Chinese language instruction (3 females, 3 males).
Apparatus
A 1.5 Tesla GE Signa whole‐body scanner and standard radio frequency coil were used for data collection. Structural images were obtained prior to stimulus presentation using a standard T1‐weighted pulse sequence in 36 contiguous slices (3.8 mm thick) parallel to the AC/PC line. In‐plane resolution was 3.75 mm × 3.75 mm. A one‐shot spiral pulse sequence was used during stimulus presentation to obtain oblique axial functional images matching the location of the middle 26 slices of the structural images [TR = 2,000 ms, TE = 35 ms, FOV = 24 cm, flip angle = 70°]. “E‐prime” software [Schneider et al., 2002] was used for stimulus presentation.
Materials
Seven types of stimuli were used: English words, English pseudowords, consonant strings, Chinese characters, Chinese pseudocharacters, Chinese noncharacters, and pinyin (an alphabetic representation of pronunciations of Chinese characters used in teaching Chinese). English words were 4–6 letters long (mean = 4.4) with a Kucera‐Francis written frequency greater than 100 (mean = 271.4) [Kucera and Francis, 1967]. Familiarity ratings [Coltheart, 1981] were greater than 550 (mean = 594.9) on a scale from 100 to 700. English pseudowords were in CVC (consonant‐vowel‐consonant) form, but were not words in Chinese or English when pronounced. The psuedowords were designed to be similar to pinyin for planned comparisons between the two stimulus types. Any differences between pinyin and pseudowords should be due to pin‐yin being associated with a meaning and tonal pronunciation. Consonant strings were matched to words for length and percent occurrence of individual consonants. Chinese characters consisted of two radicals, and were known to have been used frequently in the first year curriculum for the University of Pittsburgh Chinese classes from which we recruited our participants. Thus the characters should have been familiar to all participants. Pseudocharacters consisted of two real radicals, but the radical occurring on the left of each pseudocharacter never occurs on the left in any real Chinese character. Noncharacters were composed of two false radicals formed from real strokes. Pinyin ranged from 2 to 5 letters (mean = 3.13) and was marked for tone. All participants should have been familiar with pinyin as it was taught in the Chinese courses in which participants were enrolled and in the standard reading curriculum in China.
Procedure
Each scanning session consisted of four 7‐min runs. Within each run, the seven types of stimuli were presented in 30‐s blocks alternating with seven 30‐s blocks of fixation. Thirty stimuli (or 30 fixation crosses) were presented in white on a black background in each block alternating with 30 black screens. Each stimulus or fixation cross and each black screen was presented for 500 ms. Participants were asked to passively view the stimuli.
Analysis
Data were analyzed using the NeuroImaging Software package (NIS 3.5), developed at the University of Pittsburgh and Princeton University, and Functional Imaging Software Widgets (fiswidgets), a graphical computing environment [Fissell et al., 2003]. Images were reconstructed and corrected for subject motion using Automated Image Registration [Woods et al., 1993]. Runs in which motion exceeded 3 mm or 3 degrees in any direction were not used in analysis, which excluded four participants but no additional runs. The images were detrended to correct for scanner drift within runs using a voxelwise linear detrend. The structural images acquired for each brain were stripped to remove the skull and coregistered to a common reference brain, chosen from among the participants [Woods et al., 1993]. Functional images were transformed into the same common space, normalized by a mean scaling of each image to match global mean image intensities across subjects, and smoothed using a three‐dimensional Gaussian filter (8 mm FWHM) to account for anatomical differences between subjects. The AFNI software program [Cox, 1996] was used to visualize the data and to warp the data into Talairach space [Talairach and Tournoux, 1988].
The first three images (corresponding to the first 6 s) of each stimulus block, and the first four images (corresponding to the first 8 s) of each fixation block were removed from analysis to account for the slow rise of the BOLD response at the beginning of the stimulus blocks and its slow return to baseline in the fixation blocks. Statistical comparisons, then, reflected differences between peak activity levels and baseline levels of activity.
A whole‐brain voxel‐wise ANOVA with subjects as a random factor was performed which simply compared the viewing of a stimulus to the viewing of fixation across all stimulus types and all participants (including both native English and native Chinese readers). Regions of interest (ROIs) were determined by identifying regions containing at least eight contiguous voxels, with P < 0.000005. This corresponds to a corrected P‐value of < 0.005. Although a resulting left fusiform region connected superiorly to more primary visual areas, an automated peak‐finding algorithm verified that the two regions (fusiform and primary visual) had separable peaks [Wheeler et al., 2006]. Therefore, we made the criterion more stringent with 0.000001 < P < 0.000005 so that the two regions separated. This analysis revealed 10 regions that responded with a difference between stimulus blocks and fixation blocks (see Table I). Of those regions, only the fusiform gyri, lingual gyri, and left middle frontal regions showed more activity for stimulus blocks than fixation blocks. We focused our investigation on the fusiform gyri and two left middle frontal regions. For each participant, a mean percent signal change was computed for each region for each condition. This was computed as (stimulus – following fixation)/following fixation × 100.
Table I.
Regions of least six contiguous voxels showing a difference between stimulus blocks and fixation blocks with P < 0.000005
| Region | Voxels | x | y | z |
|---|---|---|---|---|
| Stimulus > Fixation | ||||
| L fusiform | 13 | −37 | −54 | −20 |
| R fusiform | 50 | 40 | −38 | −17 |
| L lingual gyrus | 69 | −23 | −79 | −7 |
| R lingual gyrus | 57 | 32 | −76 | −4 |
| L middle frontal | ||||
| Posterior (BA9/BA6) | 53 | −48 | 4 | 35 |
| Anterior | 8 | −44 | 23 | 27 |
| Fixation > Stimulus | ||||
| R cingulate gyrus | 16 | 10 | −19 | 38 |
| L posterior cingulate | 12 | −18 | −52 | 15 |
| R insula | 10 | 41 | −11 | 11 |
| L insula | 71 | −41 | −3 | −4 |
RESULTS
Fusiform Gyri
A left fusiform region was identified in the coordinate space of the Talairach and Tournoux atlas [Talairach and Tournoux, 1988] with the peak voxel at x = −37, y = −54, z = −20. A right fusiform region was identified with the peak voxel at x = 40, y = −38, z = −17 (See Fig. 1). Average changes in activation for all stimulus types and for both native languages were stimulus > baseline. Therefore, all F‐tests reported are differences in positive activation.
Figure 1.
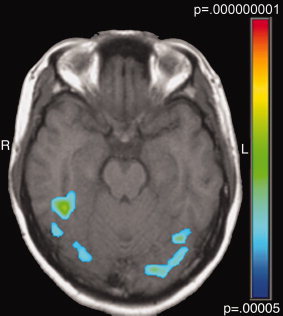
Left and right fusiform regions were identified in a voxel‐wise ANOVA, comparing stimulus blocks to fixation blocks across all participants and stimulus types. ROI's were required to contain six contiguous voxels in which P < 0.000005. Peak voxels for the left‐hemisphere region are T.C. = −37, −54, −20 and for the right‐hemisphere region are T.C. = 40, −38, −17.
To consider the overall pattern of activity in these fusiform regions, including how hemisphere might play a role, we included data from both the left and right hemisphere ROIs in an ANOVA of percent signal change with hemisphere (left vs. right), stimulus language (English vs. Chinese) and three levels of lexicality (words and characters, pseudowords and pseudocharacters, or nonwords and noncharacters) as within‐subjects factors, and participant native language (English or Chinese) as a between‐subjects factor. As necessary to understand complex interactions, we also conducted the appropriate two‐way ANOVAs and t‐tests within participant groups, the results of which are also reported when needed for clarity.
No four‐way interaction was found. However we did find two reliable three‐way interactions. The first was an interaction of participant native language × stimulus language × lexicality, F(2, 30) = 6.37, P < 0.01. While English speakers did not show an effect of lexicality in the fusiform region, Chinese speakers showed a lexicality effect for English stimuli only, F(2, 20) = 8.5, P < 0.01. T‐tests revealed that the native Chinese speakers showed the expected pattern: the nonwords activated the fusiform region less than either pseudowords or words, t(10) = 4.3, P = 001, t(10) = 3.0, P < 0.05, respectively. Our observation of fusiform sensitivity to lexicality only for Chinese participants reading English is consistent with literature showing that lexicality effects in this region are dependent on stimulus duration as well as proficiency; these findings include the observation that a long stimulus duration, such as the 500 ms exposure duration used in this study, is not optimal for probing fine‐grained stimulus differences in the fusiform gyrus [Cohen et al., 2003; Phillips et al., Varying stimulus duration reveals functional differences in left occipitotemporal cortex, submitted].
The second three‐way interaction was among participant native language, stimulus language, and hemisphere, F(1, 15) = 18.33, P = 0.001. Figures 2 and 3 illustrate that the three‐way interaction stems from the fact that the native Chinese speakers show a similar bilateral locus and amount of activity when viewing stimuli in the two languages, whereas the native English speakers show left‐lateralized activity when viewing English stimuli but bilateral activity when viewing Chinese stimuli. This is confirmed by the lack of a significant hemisphere × stimulus language interaction in the native Chinese speakers, F(1, 10) <1, contrasting with a highly reliable interaction between hemisphere and stimulus language in the native English speakers, F(1, 5) = 29.1, P < 0.01, as seen in Figure 2.
Figure 2.
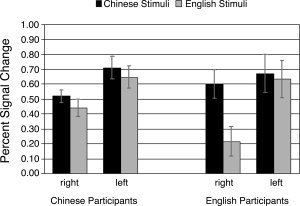
Activity in the left and right fusiform areas is characterized by a three‐way interaction between hemisphere, participant native language, and stimulus language. While native Chinese speakers activate the regions equally for Chinese and English, native English speakers show more activity for Chinese stimuli than for English stimuli in the right hemisphere, t(5) = 4.1, P = 0.01 and more activity in the left hemisphere than the right hemisphere when viewing English stimuli, t(5) = 3.3, P < 0.05.
Figure 3.
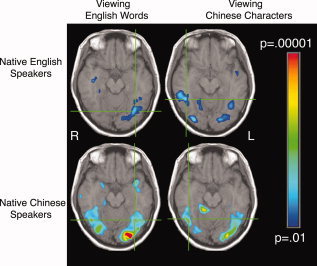
Native English speakers and native Chinese speakers viewing English words and Chinese characters compared to fixation. Peak voxels shown in the crosshairs are: for the left hemisphere ROI seen in native English speakers viewing English words, T.C. = −35, −65, −12; for the right hemisphere ROI seen in native English speakers viewing Chinese, T.C. = 43, −44, −9; for the left hemisphere ROI seen in native Chinese speakers viewing English words, T.C. = −33, −56, −12; for the right hemisphere ROI seen in native Chinese speakers viewing Chinese characters, T.C. = 43, −54, −12.
The fact that the English participants activated the right hemisphere much more for Chinese than for English stimuli also accounts for a main effect of stimulus language (Chinese > English, F(1, 15) = 7.6, P < 0.05) and an overall hemisphere × stimulus language interaction, F(1, 15) = 21.9, P < 0.001, since Chinese stimuli activated the right hemisphere, but not the left, more than English stimuli. All of the native English speakers showed at least twice the increase in activation for Chinese than for English in the right hemisphere compared to the left, and while all six showed greater activity for Chinese in the right hemisphere, only four showed greater activity for Chinese in the left.
Overall, native speakers of English who had completed a year of Chinese instruction showed left‐lateralized fusiform activity when viewing English words compared to fixation. When viewing Chinese characters, however, the native English speakers showed bilateral fusiform activity. Native Chinese speakers who are bilingual in Chinese and English showed bilateral fusiform activity when viewing Chinese characters compared to fixation. Very similar bilateral fusiform activity was seen in the Chinese participants when viewing English words compared with fixation. These results support the accommodation hypothesis for the native English speakers, who recruit right hemisphere fusiform cortex for reading Chinese, but the assimilation hypothesis for the native Chinese speakers, who activate highly overlapping bilateral fusiform regions when viewing both English and their native Chinese. Figure 3 compares all cases.
Left Middle Frontal
Two left middle frontal ROIs were identified in our comparison of activity during stimulus blocks to activity during fixation blocks. An anterior middle frontal ROI had a peak coordinate at x = −44, y = 23, z = 27, and a more posterior ROI, located near the junction of the precentral and inferior frontal sulci (BA 9/44), had a peak coordinate of x = −48, y = 4, z = 35. These regions are shown in Figure 4. Again, average changes in activation for all stimulus types and for both groups of readers were stimulus > baseline. Therefore, all F‐tests reported are differences in positive activation.
Figure 4.
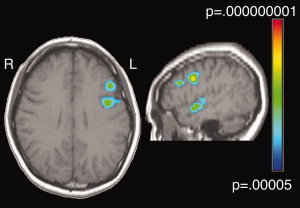
Two left middle frontal ROIs were identified in our comparison of activity during stimulus blocks to activity during fixation blocks. One was more anterior with its peak coordinate at T.C. = −44, 23, 27, and one was more posterior with its peak coordinate at T.C. = −48, 4, 35.
Since only left hemisphere middle frontal ROIs were found, hemisphere was not included in the ANOVA, which was otherwise the same as the ANOVA conducted for the fusiform ROIs: a 2 × 3 × 2 ANOVA of percent signal change with stimulus language (English vs. Chinese) and three levels of lexicality (words and characters, pseudowords and pseudocharacters, or nonwords and noncharacters) as within subjects factors, and participant native language (English or Chinese) as a between‐subjects factor. In the anterior left middle frontal region there was a main effect of stimulus language such that Chinese activated the region more than English, F(1, 15) = 5.37, P < 0.05. There were no other main effects or interactions.
In addition, we report the results of planned comparisons between pinyin (Chinese written using the Roman alphabet, marked for tone) and CVC pseudowords, which were constructed to look similar to pinyin without carrying any meaning or a Chinese pronunciation. If tonal pronunciation and/or meaning are important in modulating activity in the left middle frontal regions, we would expect to see greater activity for pinyin than for pseudowords. This ANOVA was constructed with stimulus type (pseudowords or pinyin) as the within‐subjects factor and participant native language as the between‐subjects factor. There was a trend toward pinyin activating the region more than pseudowords, F(1,15) = 3.53, P = 0.08, but no effect of participant native language and no interaction, Ps > 0.1. Thus, this anterior left middle frontal ROI appears to be driven by the analysis of Chinese characters for both sets of participants. Once again, the data suggest that the native English speakers show a pattern of accommodation to the demands of the second language. In contrast, the native Chinese speakers do not demonstrate a pattern of assimilation in this region: the region is used primarily for analysis of Chinese, but not for English.
We then performed the same analyses on the data from the posterior left middle frontal region. Figure 5 shows the pattern of activity for all stimulus types in this region.
Figure 5.
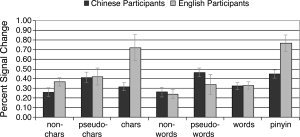
Activity in the posterior middle frontal region, T.C. = −48, 4, 35, showing a sensitivity to Chinese characters and pinyin in native English speakers.
The 2 (stimulus language) × 3 (lexicality) × 2 (participant native language) ANOVA showed no three‐way interactions. However, we did find two significant two‐way interactions. The first was between lexicality and participant native language, F(2, 30) = 8.28, P < 0.01. Chinese participants showed a significant effect of lexicality, F(2, 20) = 13.3, P < 0.001, with t‐tests revealing that the Chinese participants activated the region most for pseudowords and pseudocharacters, which produced a greater response than nonwords and noncharacters, t(10) = 4.90, P= 0.001, or words and characters, t(10) = 4.28, P < 0.01. As expected based on the interaction between lexicality and native language, the specific pattern of lexicality effects differed between the participant groups. Unlike the native Chinese speakers, the native English speakers activated the region most for words and characters, which produced a greater response than either the nonwords and noncharacters, t(15) = 3.08, P < 0.05, or the pseudowords and pseudocharacters, t(15) = 2.78, P < 0.05. The fact that nonwords and noncharacters produced weak activation for both participant groups accounts for the overall main effect of lexicality, F(2, 30) = 11.21, P < 0.001.
There was also an interaction between stimulus language and participant native language, F(1, 15) = 10.65, P < 0.01. To explore the nature of this interaction, separate t‐tests were performed comparing the Chinese and English stimuli within each participant group. The Chinese participants showed no significant effect of stimulus language, t(10) = 0.67, P > 0.5. The English participants, however, activated the region reliably more for Chinese than for English, t(5) = 2.64, P < 0.05.
These results suggest that the native English speakers are recruiting the posterior left middle frontal ROI for reading Chinese only, evidence again of accommodation even beyond the visual areas. The native Chinese speakers, however, activate the posterior left middle frontal ROI equally when viewing stimuli in both languages, evidence again of a pattern of assimilation in which Chinese readers use their native language systems for reading in English. Though the native Chinese speakers activate the ROI most for pseudowords and pseudocharacters while native English speakers show a preference for real words and characters, this may simply reflect a timing or proficiency issue. For the native Chinese readers, pseudowords and pseudocharacters may be eligible for the lexical analysis that (by hypothesis) is the function of the LMFG. This is because the radicals used in the pseudocharacters are associated with real words. For English learners, lexicality in Chinese may be noncompositional [Wang et al., 2004], so that these readers do not show a large response in this area for pseudocharacters. More important is that the region does seem to be differentially sensitive to stimuli with differing lexical status.
If this posterior LMF region is sensitive to lexical properties in Chinese, we would expect to see differences in the level of activity for viewing pinyin and viewing pseudowords, because pinyin is associated with a Chinese tonal pronunciation and meaning. This evidence came from a second ANOVA, with participants native language as a between‐subjects factor, comparing English pseudowords and pinyin, with stimulus type (pseudowords or pinyin) as the within‐subjects factor. There was a main effect of stimulus type, with pinyin activating the region more than pseudowords, F(1, 15) = 17.79, P = 0.001, and there was also a significant interaction between participant native language and stimulus language. The Chinese participants activated the region equally for both stimulus types, t(10) = 0.38, P = 0.71. The English participants activated the region more for pinyin than for pseudowords, t(5) = 4.13, P < 0.01. Thus, native English speakers utilize the left middle frontal region more for pinyin, which has a Chinese tonal pronunciation and is associated with a meaning, than for the pseudowords, which were designed to be similar in form to the pinyin. It is unclear why the native Chinese speakers do not show such a difference, but similar effects in the fusiform suggest that the ability to observe differences can be affected by expertise and exposure duration. As an alternative, pinyin may be more familiar (more recent) to the Chinese learners than to the Chinese native speakers, who see very little pinyin as adults. It is possible that because of this, characters and pinyin are more functionally equivalent for the English speakers than for the Chinese speakers.
DISCUSSION
The results support the general hypothesis that the neural basis for reading in a second language is affected by the reader's first language and its writing system. For Chinese native speakers bilingual in English, brain images acquired during passive viewing showed highly overlapping neural systems for English and Chinese, replicating previous studies showing that native Chinese speakers use a “Chinese” network to read English [Tan et al., 2003] and pinyin [Chen et al., 2002] and to perform a verb generation task in English [Pu et al., 2001]. In our study, this network included bilateral fusiform regions for reading both English and Chinese, in contrast to the left‐lateralized activity reported in native English speakers reading English. This result supports the System Assimilation Hypothesis that brain structures already functional for the writing system of the first language will be applied to a new writing system to the extent possible.
In contrast, native English speakers who were learning Chinese showed an accommodation pattern. Instead of the left‐lateralized regions that support their reading of English, these readers, when they viewed Chinese, showed bilateral fusiform activity similar to that observed for native Chinese speakers. This reversal‐accommodation for English learners of Chinese and assimilation of Chinese bilinguals‐requires an explanation that takes into account the nature of reading in the two writing systems. Before taking up the implication of this claim, we consider another possibility.
A key factor could be the level of skill in the second language/second writing system. English learners of Chinese could show a more bilateral activation because their proficiency in Chinese is low; activity in a similar right hemisphere occipitotemporal region has been negatively correlated with reading skill in English [Shaywitz et al., 2002]. However, a proficiency explanation might also predict a decrease in left hemisphere activity for Chinese stimuli, since Shaywitz et al. [2002] also found a positive correlation between left hemisphere activity and reading skill for children; we do not see this pattern in the data. A crucial test of the accommodation hypothesis thus requires a demonstration of the accommodation pattern with native speakers of English who are highly skilled in Chinese. The accommodation hypothesis predicts that the bilateral pattern will be observed regardless of skill, even if there is some decline in the level of right hemisphere activity with increased skill. We do see that the pattern holds for varying degrees of early learning when we compare our results to the Liu et al. [2006] laboratory‐learning study. In both studies, the native English speakers showed Chinese‐like brain activity when reading Chinese. While the Liu et al. study showed this very early in learning, we found that the pattern was also evident after a year of classroom learning. Until a highly skilled sample is tested, we can appeal to the important fact that the bilateral pattern observed for English native speakers mimics the pattern observed for native Chinese readers. Thus, one might expect that increasing skill in Chinese would move more, not less, toward the Chinese pattern.
Another consideration is that the native English speakers also showed activation in an anterior left middle frontal (LMF) region near an area found to be functional for skilled Chinese readers [Tan et al., 2005]. The function of the LMF region in reading Chinese remains to be specified in detail, although both semantic access [Liu et al., 2006; Tan et al., 2000] and addressed phonology [Liu et al., 2006; Tan et al., 2005] have been proposed, with the possibility that more anterior portions are used in accessing semantics, while more posterior portions are used for phonological processes, even in tasks using English text [Booth et al., 2006; Xu et al., 2001]. Though our silent reading task was not one that taxed phonological processing, the conditions that would have been most phonologically difficult for the English readers would likely be the pinyin condition (since readers must recall the Chinese pronunciations of letters in addition to tone) and the Chinese character condition, where correct pronunciations must be retrieved. Since these are the conditions showing the most activation in posterior frontal regions, a phonological processing hypothesis may account for the results. To the extent the LMF region supports spatial working memory [Courtney et al., 1998], we might expect it to function for the “intensive visualspatial analysis” required for reading Chinese [Tan et al., 2001]. A general function of the LMF region proposed by Perfetti et al. [2006], is that it supports a brief memory for the orthographic form of the character during the retrieval of its associated phonological and semantic constituents. The more anterior middle frontal area we found in our learners was virtually identical (within 3 mm, or 1 voxel, in all dimensions) to the LMF location identified by the meta‐analysis of Tan et al. for skilled native readers. It is also nearly identical to an area found by Liu et al. [2006] in their laboratory study of Chinese learning. Our posterior LMF location is very close to the location found in the Liu et al. [2006] study for learners who were exposed only to Chinese orthography and phonology, but not meaning. If what we are observing are merely the effects of low skill, they are effects that involve both posterior and frontal areas proximal to areas of well‐established importance in skilled Chinese reading. Of course, even if the effects are not due to proficiency differences, regions of activity in learners could reflect different underlying processes than overlapping regions of activity in native readers. The conclusion is not that learners' processes are identical to those used by skilled native speakers, but rather that the processes of the learner are adaptive to features of the new system.
To return to the conclusion concerning accommodation vs. assimilation, the findings of accommodation in one direction (English to Chinese) and assimilation in the other (Chinese to English) suggest that the adaptability of first language reading procedures—their compatibility with the demands of the learned system—are important. Whether a neural system assimilates or accommodates a new writing system depends on the reading procedures that have been acquired in the first system and their compatibility with what is required by the new system. Procedures that develop in reading Chinese may include both local, analytic graphic processes and more global, holistic processes [Perfetti et al., 2006]. The local processes extract high spatial frequency information from stroke sequences to build radicals. The global processes, using lower spatial frequencies, configure character shape based on radicals, which can be arranged in left‐right, up‐down, and inside‐outside manner. Left hemisphere processes may be functional for the local high spatial frequency processes and right hemisphere areas may be functional for the more global shape processes [Delis et al., 1986; Fink et al., 1999; Kitterle et al., 1990; Sergent, 1982].
These bilateral procedures can be applied to linear alphabetic writing as well, although the right hemisphere might seem less critical, given the mainly high spatial information needed to discriminate among English words. If so, then we expect to see evidence that native Chinese speakers read English in ways that suggest holistic as well as local processing. In fact, we do see such evidence. Native Chinese speakers naming English words are more affected than native English speakers by alternating case, a visual manipulation that preserves the ordered letter information required for naming, indicating a greater attention to the word form as a whole [Akamatsu, 1999]. Additionally, ESL students with Chinese as their native language show behavioral patterns consistent with greater attention to visual word form and less assembling of phonology: a large deficit in spelling nonwords compared to real words [Wang and Geva, 2003] and more confusion from orthographic similarity along with less homophone confusion in a semantic category judgment task [Wang et al., 2003].
English speakers, on the other hand, will fail to read Chinese unless they adopt procedures that process global character forms, including the hierarchical organization of high spatial frequency stroke patterns arranged in a lower spatial frequency relational pattern within a character. Thus, the linear alphabetic procedures sufficient for reading English cannot assimilate Chinese writing, and instead accommodation occurs by the addition of neural resources for the visual analysis of Chinese characters.
The collection of evidence supporting neural accommodation in English speakers learning Chinese, including activity in right hemisphere posterior visual areas and activity in the left middle frontal region, gives us confidence that a new neural system is recruited in these learners for reading Chinese. The stark contrast in the pattern for native Chinese speakers who are proficient in English, which was one of no differential activation between the two writing systems in any primary reading area, leads us to suggest that native Chinese speakers are able to assimilate English writing into their existing procedures for reading Chinese and thus into their established reading network.
Contributor Information
Jessica R. Nelson, Email: jen33@pitt.edu.
Charles A. Perfetti, Email: perfetti@pitt.edu.
REFERENCES
- Akamatsu N ( 1999): The effects of first language orthographic features on word recognition processing in English as a second language. Reading Writing 11: 381–403. [Google Scholar]
- Ben‐Shachar M,Dougherty RF,Deutsch GK,Wandell BA ( 2006): Differential sensitivity to words and shapes in ventral occipito‐temporal cortex. Cereb Cortex 17: 1604–1611. [DOI] [PubMed] [Google Scholar]
- Bolger DJ,Perfetti CA,Schneider W ( 2005): Cross‐cultural effect on the brain revisited: Universal structures plus writing system variation. Hum Brain Mapp 25: 92–104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Booth JR,Lu D,Burman DD,Chou T‐L,Jin Z,Peng D‐L,Zhang L,Ding G‐S,Deng Y,Liu L ( 2006): Specialization of phonological and semantic processing in Chinese word reading. Brain Res 1071: 197–207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chee MWL,Caplan D,Soon CS,Sriram N,Tan EWL,Thiel T,Weekes B ( 1999a) Processing of visually presented sentences in Mandarin and English studied with fMRI. Neuron 23: 127–137. [DOI] [PubMed] [Google Scholar]
- Chee MWL, O'Craven, KM ,Begida R,Rosen BR,Savoy RL ( 1999b) Auditory and visual word processing studied with fMRI. Hum Brain Mapp 7: 15–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chee MWL,Tan EWL,Thiel T ( 1999c) Mandarin and English single word processing studied with functional magnetic resonance imaging. J Neurosci 19: 3050–3056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chee MWL,Weekes B,Lee KM,Soon CS,Schreiber A,Hoon JJ,Chee M ( 2000): Overlap and dissociation of semantic processing of Chinese characters, English words, and pictures: Evidence from fMRI. NeuroImage 12: 392–403. [DOI] [PubMed] [Google Scholar]
- Chee MWL,Hon N,Lee HL,Soon CS ( 2001): Relative language proficiency modulates BOLD signal change when bilinguals perform semantic judgments. NeuroImage 13: 1155–1163. [DOI] [PubMed] [Google Scholar]
- Chee MWL,Soon CS,Lee HL ( 2003): Common and segregated neuronal networks for different languages revealed using functional magnetic resonance adaptation. J Cogn Neurosci 15: 85–97. [DOI] [PubMed] [Google Scholar]
- Chen Y,Allport D,Marshall J ( 1996): What are the functional orthographic units in Chinese word recognition: The stroke or the stroke pattern? Q J Exp Psychol A 49: 1024–1043. [Google Scholar]
- Chen Y,Fu S,Iversen SD,Smith SM,Matthews PM ( 2002): Testing for dual brain processing routes in reading: A direct contrast of Chinese character and Pinyin Reading using fMRI. J Cogn Neurosci 14: 1088–1098. [DOI] [PubMed] [Google Scholar]
- Cohen L,Lehericy S,Chochon F,Lemer C,Rivaud S,Dehaene S ( 2002): Language‐specific tuning of visual cortex? Functional properties of the Visual Word Form Area. Brain 125: 1054–1069. [DOI] [PubMed] [Google Scholar]
- Cohen L,Martinaud O,Lemer C,Lehericy S,Samson Y,Obadia M,Slachevsky A,Dehaene S ( 2003): Visual word recognition in the left and right hemispheres: Anatomical and functional correlates of peripheral alexias. Cereb Cortex 13: 1313–1333. [DOI] [PubMed] [Google Scholar]
- Coltheart M ( 1981): The MRC psycholinguistic database. Q J Exp Psychol A 33: 497–505. [Google Scholar]
- Courtney SM,Petit L,Maisog JM,Ungerleider LG,Haxby JV ( 1998): An area specialized for spatial working memory in human frontal cortex. Science 279: 1347–1351. [DOI] [PubMed] [Google Scholar]
- Cox RW ( 1996): AFNI: Software for analysis and visualization of functional magnetic resonance neuroimages. Comput Biomed Res 29: 162–173. [DOI] [PubMed] [Google Scholar]
- Dehaene S,Le Clec'H G,Poline JB,Le Bihan D,Cohen L ( 2002): The visual word form area: A prelexical representation of visual words in the fusiform gyrus. Neuroreport 13: 321–325. [DOI] [PubMed] [Google Scholar]
- Delis DC,Robertson LC,Efron R ( 1986): Hemispheric specialization of memory for visual hierarchical stimuli. Neuropsychologia 24: 205–214. [DOI] [PubMed] [Google Scholar]
- Fink G,Marshall J,Halligan P,Dolan R ( 1999): Hemispheric asymmetries in global/local processing are modulated by perceptual salience. Neuropsychologia 37: 31–40. [DOI] [PubMed] [Google Scholar]
- Fissell K,Tseytlin E,Cunningham D,Iyer K,Carter CS,Schneider W,Cohen JD ( 2003): Fiswidgets: A graphical computing environment for neuroimaging analysis. Neuroinformatics 1: 111–125. [DOI] [PubMed] [Google Scholar]
- Kitterle FL,Christman S,Hellige JB ( 1990): Hemispheric differences are found in the identification, but not the detection, of low versus high spatial frequencies. Percept Psycophys 48: 297–306. [DOI] [PubMed] [Google Scholar]
- Kucera H,Francis WN ( 1967): Computational Analysis of Present‐Day American English. Providence, RI: Brown University Press. [Google Scholar]
- Kuo W‐J,Yeh T‐C,Duann J‐R,Wu Y‐T,Ho L‐T,Hung D,Tzeng OJL,Hsieh J‐C ( 2001): A left‐lateralized network for reading Chinese words: a 3T‐fMRI study. Neuroreport 12: 3997–4001. [DOI] [PubMed] [Google Scholar]
- Lam AS,Perfetti CA,Bell L ( 1991): Automatic phonetic transfer in bidialectal reading. Appl Psycholinguist 12: 299–311. [Google Scholar]
- Liu Y,Perfetti CA ( 2003): The time course of brain activity in reading English and Chinese: An ERP study of Chinese bilinguals. Hum Brain Mapp 18: 167–175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y,Dunlap S,Fiez JA,Perfetti CA ( 2007): Evidence for neural accommodation to a writing system following learning. Hum Brain Mapp 28: 1223–1234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCandliss BD,Cohen L,Dehaene S ( 2003): The visual word form area: Expertise for reading in the fusiform gyrus. Trends Cogn Sci 7: 293–299. [DOI] [PubMed] [Google Scholar]
- Perfetti CA ( 2003): The universal grammar of reading. Sci Stud Read 7: 3–24. [Google Scholar]
- Perfetti CA,Zhang S ( 1991): Phonological processes in reading Chinese characters. J Exp Psychol Learn Mem Cogn 17: 633–643. [Google Scholar]
- Perfetti CA,Liu Y ( 2005): Orthography to phonology and meaning: Comparisons across and within writing systems. Reading Writing 18: 193–210. [Google Scholar]
- Perfetti CA,Liu Y,Tan LH ( 2005): The lexical constituency model: Some implications of research on Chinese for general theories of reading. Psychol Rev 112: 43–59. [DOI] [PubMed] [Google Scholar]
- Perfetti CA,Liu Y,Fiez J,Tan LH ( 2008): The neural bases of reading: The accommodation of the brain's reading network to writing systems In: Cornelissen P,Kringelbach M,Hansen P, editors. The Neural Basis of Reading, Oxford University Press. [Google Scholar]
- Pu Y,Liu HL,Spinks JA,Mahankali S,Xiong J,Feng CM,Tan LH,Fox PT,Gao JH ( 2001): Cerebral hemodynamic response in Chinese (first) and English (second) language processing revealed by event‐related functional MRI. Magn Reson Imaging 19: 643–647. [DOI] [PubMed] [Google Scholar]
- Rack JP,Snowling MJ,Olson RK ( 1992): The nonword reading deficit in developmental dyslexia: A review. Read Res Q 27: 28–53. [Google Scholar]
- Schneider W,Eschman A,Zuccolotto A. 2002. E‐Prime User's Guide. Pittsburgh, PA: Psychology Software Tools. [Google Scholar]
- Sergent J ( 1982): The cerebral balance of power: Confrontation or cooperation? J Exp Psychol Hum Percept Perform 8: 253–272. [DOI] [PubMed] [Google Scholar]
- Shaywitz BA,Shaywitz SE,Pugh KR,Mencl WE,Fulbright RK,Skudlarski P,Constable RT,Marchione KE,Fletcher JM,Lyon GR,Gore JC ( 2002): Disruption of posterior brain systems for reading in children with developmental dyslexia. Biol Psychiatry 52: 101–110. [DOI] [PubMed] [Google Scholar]
- So D,Siegel LS ( 1997): Learning to read Chinese: Semantic, syntactic, phonological and working memory skills in normally achieving and poor Chinese readers. Reading Writing 9: 1–21. [Google Scholar]
- Spinks JA,Liu Y,Perfetti CA,Tan LH ( 2000): Reading Chinese characters for meaning: The role of phonological information. Cognition 76: B1–B11. [DOI] [PubMed] [Google Scholar]
- Talairach J,Tournoux P. 1988. Co‐Planar Stereotactic Atlas of the Human Brain. New York: Thieme. [Google Scholar]
- Tan LH,Perfetti CA ( 1998): Phonological codes as early sources of constraint in Chinese word identification: A review of current discoveries and theoretical accounts. Reading Writing 10: 165–200. [Google Scholar]
- Tan LH,Hoosain R,Siok WW ( 1996): Activation of phonological codes before access to character meaning in written Chinese. J Exp Psychol Learn Mem Cogn 22: 865–882. [Google Scholar]
- Tan LH,Spinks JA,Gao JH,Liu HL,Perfetti CA,Xiong J,Stofer KA,Pu Y,Liu Y,Fox PT ( 2000): Brain activation in the processing of Chinese characters and words: A functional MRI study. Hum Brain Mapp 10: 16–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH,Liu HL,Perfetti CA,Spinks JA,Fox PT,Gao JH ( 2001): The neural system underlying Chinese logograph reading. Neuroimage 13: 836–846. [DOI] [PubMed] [Google Scholar]
- Tan LH,Spinks JA,Feng CM,Siok WT,Perfetti CA,Xiong J,Fox PT,Gao JH ( 2003): Neural systems of second language reading are shaped by native language. Hum Brain Mapp 18: 158–166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan LH,Laird A,Li K,Fox PT ( 2005): Neuroanatomical correlates of phonological processing of Chinese characters and alphabetic words: A meta‐analysis. Hum Brain Mapp 25: 83–91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagner RK,Torgesen JK ( 1987): The nature of phonological processing and its causal role in the acquisition of reading skills. Psychol Bull 101: 192–212. [Google Scholar]
- Wagner RK,Torgesen JK,Rashotte CA,Hecht SA,Barker TA,Burgess SR,Donahue J,Garon T ( 1997): Changing relations between phonological processing abilities and word‐level reading as children develop from beginning to skilled readers: A 5‐year longitudinal study. Dev Psychol 33: 468–479. [DOI] [PubMed] [Google Scholar]
- Wang M,Geva E ( 2003): Spelling performance of Chinese children using English as a second language: Lexical and visual‐orthographic processes. Appl Psycholinguist 24: 1–25. [Google Scholar]
- Wang M,Koda K,Perfetti CA ( 2003): Alphabetic and nonalphabetic L1 effects in English word identification: A comparison of Korean and Chinese English L2 learners. Cognition 87: 129–149. [DOI] [PubMed] [Google Scholar]
- Wang M,Liu Y,Perfetti CA ( 2004): The implicit and explicit learning of Chinese orthographic structure and function by alphabetic readers. Sci Stud of Read 8: 357–379. [Google Scholar]
- Wheeler ME,Shulman GL,Buckner RL,Miezin FM,Velanova K,Petersen SE ( 2006): Evidence for separate perceptual reactivation and search processes during remembering. Cereb Cortex 16: 949–959. [DOI] [PubMed] [Google Scholar]
- Woods RP,Mazziotta JC,Cherry SR ( 1993): MRI‐PET registration with automated algorithm. J Comput Assist Tomogr 17: 536–546. [DOI] [PubMed] [Google Scholar]
- Xu B,Grafman J,Gaillard WD,Ishii K,Vega‐Bermudez F,Pietrini P,Reeves‐Tyer P,DiCamillo P,Theodore W ( 2001): Conjoint and extended neural networks for the computation of speech codes: The neural basis of selective impairment in reading words and pseudowords. Cereb Cortex 11: 267–277. [DOI] [PubMed] [Google Scholar]
- Xue G,Dong Q,Jin Z,Zhang L,Wang Y ( 2004): An fMRI study with semantic access in low proficiency second language learners. NeuroReport 15: 791–796. [DOI] [PubMed] [Google Scholar]