

Abstract
RNA sequencing studies have identified hundreds of non‐coding RNAs in bacteria, including regulatory small RNA (sRNA). However, our understanding of sRNA function has lagged behind their identification due to a lack of tools for the high‐throughput analysis of RNA–RNA interactions in bacteria. Here we demonstrate that in vivo sRNA–mRNA duplexes can be recovered using UV‐crosslinking, ligation and sequencing of hybrids (CLASH). Many sRNAs recruit the endoribonuclease, RNase E, to facilitate processing of mRNAs. We were able to recover base‐paired sRNA–mRNA duplexes in association with RNase E, allowing proximity‐dependent ligation and sequencing of cognate sRNA–mRNA pairs as chimeric reads. We verified that this approach captures bona fide sRNA–mRNA interactions. Clustering analyses identified novel sRNA seed regions and sets of potentially co‐regulated target mRNAs. We identified multiple mRNA targets for the pathotype‐specific sRNA Esr41, which was shown to regulate colicin sensitivity and iron transport in E. coli. Numerous sRNA interactions were also identified with non‐coding RNAs, including sRNAs and tRNAs, demonstrating the high complexity of the sRNA interactome.
Keywords: CLIP‐Seq, CRAC, EHEC, enterohaemorrhagic E. coli, non‐coding RNA
Subject Categories: Methods & Resources; Microbiology, Virology & Host Pathogen Interaction; RNA Biology
Introduction
Advances in RNA sequencing technologies and associated applications have driven a revolution in our understanding of the complexity of the transcriptome. For diverse bacterial species, a single RNA‐Seq experiment can reveal hundreds of novel non‐coding RNAs. Bacterial small RNA (sRNA) species regulate translation of mRNAs involved in a diverse range of physiological processes including carbon, amino acid and metal ion utilization (Papenfort & Vogel, 2014), horizontal transfer of DNA (Papenfort et al, 2015), biofilm formation (Holmqvist et al, 2010) and virulence gene expression (Chao & Vogel, 2010). Canonically, sRNAs repress mRNA translation by base pairing that covers the ribosome‐binding site and/or directing the transcript for cleavage and degradation. It is now apparent that there are many variations on this canonical theme including activation of translation (Soper et al, 2010), repression by cleavage alone (Pfeiffer et al, 2009), cleavage inhibition (Papenfort et al, 2013), transcriptional attenuation (Bossi et al, 2012) and sRNA sponging (Figueroa‐Bossi et al, 2009; Tree et al, 2014; Miyakoshi et al, 2015). The majority of sRNAs in E. coli require the RNA chaperone Hfq to anneal with target mRNAs (Gottesman & Storz 2011). Hfq can present sRNAs for interaction with the pool of mRNA targets, increasing the local concentration of interaction partners and providing a positively charged lateral surface to aid annealing (Panja et al, 2013).
In principal, targets for sRNA interactions can be predicted using sequence‐based analysis; however, few sequence or structural features are conserved between the many different sRNA targets, making false positives a major problem (Backofen et al, 2014; Künne et al 2014). To overcome this, target prediction programmes have used the presence of a tract of 6 or more consecutive base pairs (the seed sequence) and the predicted accessibility of the seed region (Peer & Margalit, 2011). Phylogenetic conservation of seed sequences also improves the likelihood of identifying functionally significant interactions but is not applicable to transcripts encoded within variable regions of the genome, such as pathogenicity islands. In consequence, determining the targets for sRNAs and their regulatory function has generally required the investigation of individual RNAs, often by using transcriptomics to indirectly identify mRNAs with altered stability following sRNA expression or depletion.
A number of recent studies have implemented in vitro and in vivo techniques to directly identify interactions between non‐coding RNAs and their RNA targets. These have included approaches using individual microRNAs or bacterial sRNAs as baits, with or without chemical modifications to improve capture of interacting RNAs. High‐throughput sequencing allows identification of target RNAs interacting with the bait RNA (Imig et al, 2015). This approach unexpectedly identified a spacer region from the tRNA‐Leu precursor as a target for RyhB (Lalaouna et al, 2015). An approach to experimentally profile transcriptome‐wide RNA–RNA interactions in eukaryotic cells has been described that uses proximity‐dependent ligation of duplexed RNAs to capture RNA interactions in vivo and has been termed CLASH (UV‐crosslinking, ligation and sequencing of hybrids) (Helwak et al, 2013) (Fig 1A). RNA–RNA duplexes are UV‐crosslinked to a protein “bait” allowing selective capture of RNAs and stringent purification of the RNA–protein complex. A small fraction of RNAs covalently bound to the protein remain duplexed during purification and these can be ligated into a single contiguous RNA molecule with T4 RNA ligase (Helwak et al, 2013) or by endogenous RNA ligases (Grosswendt et al, 2014). An alternative methodology uses a joining linker to ligate the constrained duplex ends of the RNAs (Sugimoto et al, 2015). In each case, a proportion of sequencing reads recovered (typically ~1–2%) consist of read segments that non‐contiguously map to the transcriptome. These hybrid reads can be identified in silico and indicate sites of intra‐ or intermolecular RNA–RNA interactions occurring on the bait protein.
Figure 1. UV‐crosslinking of RNase E reveals binding sites transcriptome‐wide.

- Schematic of CLASH protocol for purification of RNA–RNA interactions. RNAs were UV‐crosslinked to RNase E‐HTF in vivo and purified using M2 anti‐FLAG resin. RNAs were trimmed using RNase A/T1 and further purified under denaturing conditions. RNA linkers were ligated to the immobilized RNA–RNase E complexes. Duplexed RNAs may be ligated into a single contiguous molecule (left, CLASH) that gives information on RNA–RNA interaction occurring on RNase E. The remaining single RNAs reveal the site of RNase E binding within the transcriptome. Linker‐ligated RNA–RNase E complexes were size‐selected by SDS–PAGE and RNAs recovered for library preparation and sequencing. The schematic on the right represents the key steps in preparing UV‐crosslinked RNA–protein complexes to map RNA–protein interactions sites (∼99% of reads recovered), and RNA–RNA interaction sites (∼1% of reads recovered). Colours correspond to key words in the flow diagram.
- The 5′ UTR of rne is bound by RNase E and non‐genomically encoded oligo(A) tails are maximally recovered −9nt from the rne start codon. Known stem loop structures (HP1–3) and the ribosomal binding site (RBS) are shaded grey.
- RNase E binding and oligoadenylation of the pldB‐yigL dicistronic transcript. The reported RNase E cleavage site (red dashed line) and SgrS binding site (grey shading) are indicated.
- Length of non‐genomically encoded oligo(A) tails recovered from RNase E‐bound reads.
- Position of Hfq binding and oligoadenylation relative to RNase E binding peaks. The cumulative position of Hfq and oligoadenylation peaks was determined relative to RNase E binding peaks for 672 RNase E binding sites that were within 1 kb of an Hfq binding peak. A detailed description of the data processing is presented in the Appendix Supplementary Methods.
RNase E is an endonuclease that plays key roles in both the catalytic activity and assembly of the RNA degradosome, a complex responsible for the majority of RNA processing and bulk RNA turnover (Mackie, 2013). The C‐terminal domain of RNase E interacts with RhlB (helicase), PNPase (polynucleotide polymerase and 3′ to 5′ exoribonuclease activities) and PAPI (poly(A) polymerase). Both PAPI and PNPase can add oligonucleotide tails (oligo(A) or A‐rich, respectively) to the 3′ ends of RNAs following RNase E cleavage. This creates a single‐stranded “landing pad” that promotes subsequent degradation by 3′‐exonucleases (Khemici & Carpousis, 2004). In CLASH analyses, the 3′ ends of sequence reads will not generally correspond to in vivo cleavage sites because the RNA fragments are treated with RNase during library preparation. However, the presence of a non‐encoded oligo(A) tract at the 3′ end of sequence reads is a clear indication that this represents a site that was cleaved and then oligoadenylated in vivo.
We previously reported that UV‐crosslinking and high‐throughput sequencing (CRAC) can be used to identify the binding sites for the RNA chaperone, Hfq, at base pair resolution in the model prokaryote E. coli and the related human pathogen, enterohaemorrhagic E. coli (EHEC) (Tree et al, 2014). These studies revealed that for many sRNA–mRNA interactions, the Hfq binding site is closely associated with the mRNA seed sequence. Formation of the sRNA–mRNA duplex at the Hfq binding site is predicted to induce dissociation from the single‐stranded RNA binding site on the chaperone, providing directionality to the reaction (Tree et al, 2014). The endonuclease activity of RNase E is strongly stimulated by the presence of a free 5′ monophosphate on the substrate and a 5′ triphosphate therefore stabilizes newly synthesized mRNAs (Mackie, 1998). Recent work has demonstrated that sRNA–mRNA duplexes can guide RNase E cleavage of the mRNA by providing a free 5′ monophosphate to stimulate cleavage (Bandyra et al, 2012).
Together, these results indicated that formation of an sRNA–mRNA duplex may cause dissociation from Hfq and then direct RNase E cleavage of the mRNA. To test this model, we have identified targets of sRNA‐mediated degradation transcriptome‐wide and in vivo by applying CLASH to RNase E.
Results
UV‐crosslinking identifies in vivo binding sites for RNase E
We reasoned that duplexed sRNA–mRNA pairs might be transiently associated with RNase E prior to mRNA degradation, allowing tagged RNase E to act as a bait in the capture of in vivo interactions by UV‐crosslinking (CLASH) (Fig 1A). To facilitate affinity purification of RNA–RNase E complexes, the chromosomal copy of RNase E (rne) was C‐terminally tagged with a tandem affinity His6‐TEV cleavage site‐FLAG tag (HTF). RNase E is essential for cell viability and was previously shown to retain function when C‐terminally FLAG‐tagged at the same site (Morita et al, 2005; Worrall et al, 2008). The strain expressing only RNase E‐HTF was viable and showed normal processing of 9S rRNA precursor into mature 5S rRNA (Ghora & Apirion, 1978), indicating that the fusion protein is functional (Fig EV1A). Following UV‐crosslinking in actively growing cells, RNA–RNase E‐HTF complexes were affinity‐purified under denaturing conditions and crosslinked RNAs were trimmed using mild RNase A/T1 digestion. T4 RNA ligase was added to join RNase E‐associated RNA duplexes into hybrid sequences, and to ligate Illumina sequencing compatible linkers to the ends of RNA fragments. Silver staining of eluates revealed co‐precipitated proteins, with a clearly separated protein at the expected molecular weight of 118 kDa (Fig EV1B). We confirmed that this band was RNase E using LC‐MS/MS. RNA–RNase E complexes were transferred to nitrocellulose, excised from the appropriate fragment of the membrane and recovered by protease digestion. Sequencing libraries were prepared by RT–PCR. Duplicate UV‐crosslinking experiments showed a strong correlation in the number of reads mapping to individual transcripts (Spearman correlation = 0.97), and 79% of RNase E binding sites in dataset #2 (lower read depth) were also recovered in dataset #1. Sequence reads were mapped to the genome and represent sites of RNase E–RNA interaction (read statistics presented in Table EV1). Read clusters with > 10 reads were identified in 75% of annotated mRNAs, likely representing the repertoire of mRNAs expressed under our experimental conditions (Fig EV2), as RNase E is reported to be the primary factor responsible for initiating bulk mRNA turnover. In addition, close to 1% of reads were mapped to non‐contiguous sites in the genome and represented RNA–RNA hybrid reads (see below).
Figure EV1. C‐terminally tagged RNase E is functional and facilitates purification of labelled RNA–RNase E complexes.

- RNase E is required for processing of 9S rRNA into mature 5S rRNA, and we assessed 5S rRNA processing in our His‐FLAG‐tagged RNase E strain. Enterohaemorrhagic E. coli (EHEC) O157:H7 str. Sakai and the isogenic rne‐HTF insertion mutant were grown to an OD600 of 0.6 in LB broth at 37°C and total RNA was harvested. To compare wild‐type and impaired 5S rRNA processing, we additionally cultured E. coli str. K12 (N3433) and the isogenic temperature‐sensitive rne‐3071 mutant (N3431) to OD600 0.6. The rne‐3071 mutant was temperature‐shifted to 43°C for 30 min and total RNA harvested. 1 μg of total RNA was separated on an 8% TBE–urea polyacrylamide gel and blotted for 9S rRNA. Mature 5S rRNA is shown in the SYBR‐stained loading control below.
- RNA–protein complexes were eluted from Ni‐NTA resin (see Materials and Methods) and precipitated and 50% of the eluate separated on a NuPAGE Bis‐Tris 4–12% gel and silver‐stained. E. coli O157:H7 str. Sakai (untagged; negative control) and the isogenic rne‐HTF mutant were analysed for stringency of the dual‐affinity purification.
- Replicate [32P]‐labelled RNA–RNase E complexes were separated on NuPAGE Bis‐Tris 4–12% gradient gels and transferred to nitrocellulose membranes. The labelled complexes were imaged by autoradiography and complexes with a molecular mass equivalent to RNase E–RNA fragments (dashed boxes) were excised from the membrane (see Materials and Methods).
Source data are available online for this figure.
Figure EV2. Circos plot of RNase E binding sites (grey), non‐genomically encoded oligoadenylation sites (red), and Hfq binding sites (blue).
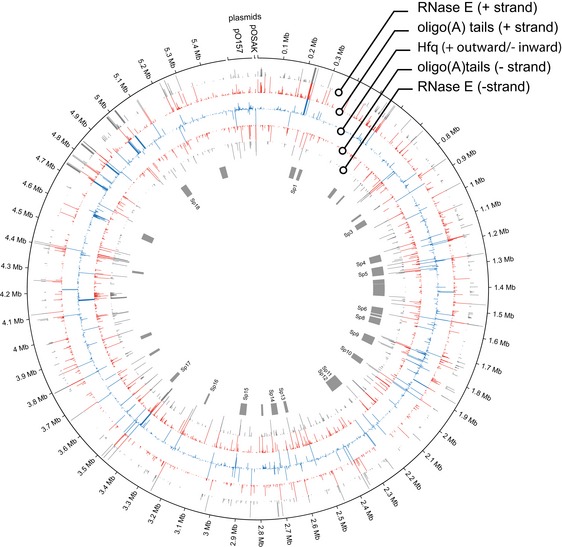
Positive strand binding sites are shown in the outer rings and negative strand binding in the inner rings. Pathogenicity islands (S‐loops [Sp]) are shown in grey (centre).
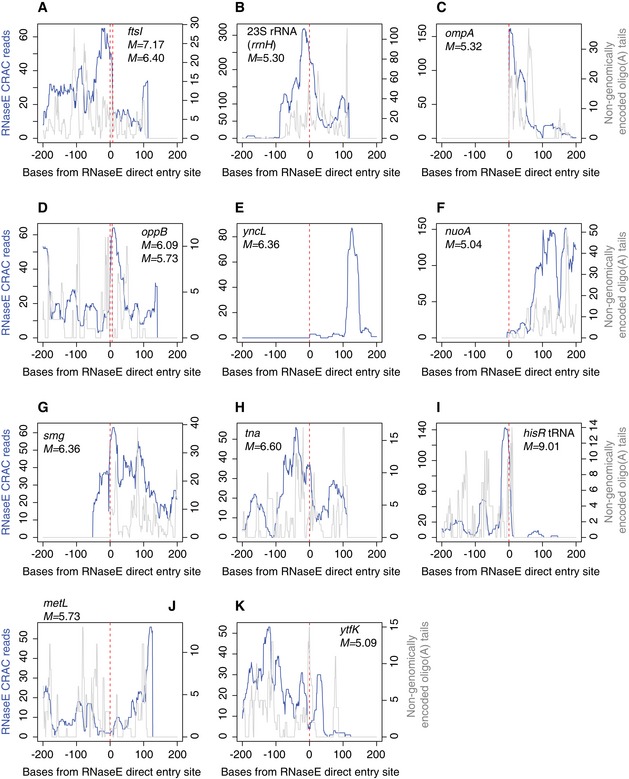
As an initial step to verify our approach, we tested whether UV‐crosslinking of RNA–RNase E complexes in vivo recovered known RNase E binding sites. Photocrosslinking experiments have demonstrated that RNase E autoregulates the stability of its own transcript (rne) by binding the hairpin structures HP1–HP3 within the 5′ UTR (Diwa et al, 2000; Schuck et al, 2009). We found that RNase E indeed binds to all three HP structures in vivo. Oligoadenylated reads, which are strongly indicative of endogenous 3′ ends (Khemici & Carpousis, 2004), peaked at −9 nts relative to the rne start codon, indicating that RNase E cleaves the rne transcript near the ribosomal binding site (Fig 1B). The small RNA SgrS binds pldB at +935−955 nt and stabilizes the yigL transcript by occluding an RNase E cleavage site at +948−955 nt within the dicistronic pldB‐yigL mRNA (Papenfort et al, 2013). In agreement with this study, we find that RNase E binds 5′ of this cleavage site and overlaps the SgrS interaction site (Fig 1C). RNase E cleavage sites were recently mapped transcriptome‐wide, identifying sites of 5′ monophosphate‐independent (“direct entry”) RNA cleavage (Clarke et al, 2014). We assessed RNase E binding at reported RNase E direct entry sites. Thirteen sites had > 50 reads within 200 nt of the direct entry cleavage site and ten showed a clear peak in RNase E binding or oligoadenylation at the direct entry site (Fig EV3). We conclude that our in vivo RNase E binding sites agree with published interactions and represent bona fide targets.
Figure EV3. RNase E binding and oligoadenylation at RNase E direct entry sites.
-
A–KDirect entry sites identified by Clarke et al (2014) were screened for peaks > 50 reads ± 200 nt from the direct entry site in our EHEC RNase E crosslinking dataset #1. RNase E binding (blue) and oligoadenylation (grey) are shown. Gene names, enrichment scores (M) and coordinates for direct entry sites were taken from Clarke et al (2014). For panels (A and D), two closely associated direct entry sites are indicated on the same plot.
Relationship between RNase E, Hfq and oligoadenylation sites
We previously reported that non‐genomically encoded oligo(A) tails of 2–6 nt were present in 5% of Hfq‐bound sequences (Tree et al, 2014). This indicates that Hfq binding sites are associated with endogenous 3′ ends that are oligoadenylated by PAPI. Oligo(A) tails were found in 0.7% of RNase E‐bound reads and were predominately (76%) between 2 and 6 nt in length (Fig 1D). Hfq interacts with RNase E (Morita et al, 2005; Worrall et al, 2008), and sRNA interactions with an mRNA can facilitate RNase E recruitment and cleavage (Ikeda et al, 2011; Prévost et al, 2011; Bandyra et al, 2012). To gain insights into the arrangement of binding and cleavage sites, we compared the distribution of oligoadenylated sequences and Hfq crosslinking relative to RNase E binding sites. Maximal Hfq binding was cumulatively found five base pairs 5′ of the RNase E binding maximum (Fig 1E) although we note a significant overlap in these binding sites. In contrast, reads with oligo(A) tails, reflecting in vivo cleavage sites, were maximally recovered 13 base pairs 3′ of the peak in RNase E binding (Fig 1E).
These results support a model in which RNase E is frequently recruited to Hfq binding sites with a five base pair 3′‐offset leading to RNA cleavage 13 nt downstream of the RNase E binding site and addition of a 2‐ to 6‐nt oligo(A) tail. Recovery of more distant RNase E cleavage and oligoadenylation sites is limited by the length of the sequencing read. However, we note that our observations are consistent with in vitro characterization of the MicC–ompD interaction that directs RNase E cleavage 6 base pairs downstream of the sRNA–mRNA duplex (Bandyra et al, 2012).
RNA–RNA interactions are recovered by RNase E‐CLASH
In CLASH analyses, RNA duplexes that are bound by RNase E can be ligated together and recovered as cDNA sequencing reads that map non‐contiguously to distinct sites in the transcriptome. These were identified and mapped using the Hyb software package (Travis et al, 2014). From 21.9 M mapped reads, we recovered 176,874 RNA–RNA interactions (0.8%, Tables EV1 and EV2) including 1,733 sRNA–mRNA interactions (Table EV3). There was substantial overlap between hybrids recovered in the two replicate datasets, and 41% of interactions identified in replicate #2 were also recovered in the larger replicate #1 dataset. We used the approach of Sharma et al (2016) to assess the theoretical false discovery rate expected from random ligation of RNAs in solution, and find that 58.8% of RNA–RNA interactions have an FDR < 0.05 (Table EV2 and Appendix Supplementary Methods).
To verify that RNase E‐CLASH recovered bona fide sRNA–mRNA interactions, we looked for 125 experimentally verified sRNA–mRNA pairs within our datasets (Table EV4). Small RNA interactions were taken from sRNATarBase 3.0 (Wang et al, 2015), inspected for concordance with published sites and corrected where necessary (corrections to sRNATarBase 3.0 are presented in Table EV4). RNase E‐CLASH analysis identified a statistically significant number of known sRNA–mRNA pairs (14/125, P < 6.6 × 10−4; Table EV5 and Appendix Supplementary Methods) including the sRNA–mRNA pair MicA–ompA (Fig 2A and B) (Rasmussen et al, 2005; Udekwu et al, 2005). We performed RNA‐Seq on total RNA from EHEC and found that the recovery of hybrid reads was only weakly correlated with RNA abundance (Spearman correlation = 0.15; Fig EV4A), but was moderately correlated with RNase E crosslinking to single RNAs (Spearman correlation = 0.44; Fig EV4B). Similar results were found for the 125 known sRNA–mRNA interactions where hybrid recovery correlates more significantly with RNase E crosslinking (Spearman correlation = 0.15 for mRNA binding; Fig EV4C–F). Hybrid recovery is likely a function of both sRNA and mRNA association with RNase E, and we find a general trend towards higher numbers of hybrid reads for known sRNA–mRNA interactions where both single RNAs were strongly crosslinked to RNase E (Fig EV4H). These results are consistent with hybrid reads being derived from RNA interactions on RNase E rather than from total cellular RNA. Small RNAs interact with mRNAs through base pairing, and hybrid reads generated from duplexed RNAs are predicted to have a lower‐than‐random free energy of interaction (∆G) (i.e. greater stability). We compared the distribution of free energies for all RNA–RNA interactions identified (Fig 2C) and for sRNA–mRNA pairs (Fig 2D) with randomly paired hybrid read halves. The distribution of free energies from RNase E‐CLASH RNA–RNA interactions was significantly lower than for random pairs. These results are consistent with the hybrid sequences being derived from duplexed RNAs associated with RNase E.
Figure 2. Relative position, RNA class, and base pairing strength of small RNA hybrids recovered by RNase E‐CLASH .

- In silico folding of hybrid reads using the UNAfold suite of tools was used to predict base pairing and interaction strength between hybrid read halves. Reads mapping to the interaction between MicA and ompA mRNA are shown (ompA start codon in bold).
- Hfq binding (blue), RNase E binding (grey) and oligoadenylation (red) of the MicA binding site in the ompA mRNA. The MicA seed sequence is shaded blue.
- Interaction strength (Gibbs free energy) of all RNA–RNA interactions recovered by RNase E‐CLASH (solid line) and for randomly paired hybrid read halves (dashed line).
- Interaction strength of sRNA–mRNA hybrid read halves (solid line) and randomly paired sRNA and mRNA read halves (dashed line).
- Position of sRNA–mRNA interactions relative to the mRNA start codon.
- Distribution of sRNA interactions recovered with each class of RNA.
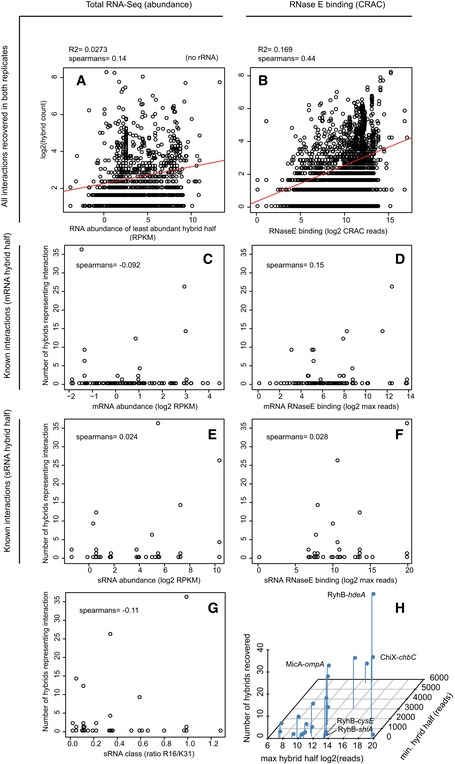
Figure EV4. Correlations between recovery of hybrids, total RNA abundance and RNase E crosslinking.
-
A, BFor all interactions recovered in both replicates, the number of hybrid reads mapping to an interaction was correlated with RNA abundance (of the least abundant hybrid half; A), and RNase E crosslinking (also for the lowest hybrid half; B).
-
C–FThe number of hybrids recovered for 125 experimentally verified sRNA–mRNA interactions was also correlated with RNA abundance at the target (C) and sRNA (E), and RNase E crosslinking at the mRNA (D) and sRNA (F).
-
GFor each known sRNA–mRNA interaction, the number of hybrids recovered was also plotted against the ratio of HfqR16/HfqK31 IP (previously determined by Schu et al 2015). The HfqR16/HfqK31 ratio is indicative of sRNA class where Class I < 1 < Class II sRNAs. For each plot, the Spearman correlation between the variables is shown.
-
HRNase E crosslinking and hybrid recovery for known sRNA–mRNA interactions including those described in Fig 5. The number of hybrids representing an interaction (z‐axis) was plotted against RNase E crosslinking for the lower (y‐axis) and higher (x‐axis) crosslinked halves.
Interactions between sRNAs and mRNAs that impair 30S ribosome binding and translation are generally positioned within a window extending from 50 nt upstream to 15 nt (five codons) downstream of the start codon (Bouvier et al, 2008). Binding sites for sRNAs identified by RNase E‐CLASH were enriched within this window on mRNAs (Fig 2E), in agreement with 30S occlusion as a major pathway for sRNA function.
RNase E acts as a scaffold for the RNA degradosome and plays important roles in the degradation and processing of all RNA classes in E. coli (Mackie, 2013). We therefore determined the proportion of unique sRNA interactions that were contributed by each RNA class (Fig 2F). Messenger RNA coding regions and 5′ UTRs are characterized substrates for sRNA interactions and constituted 43.1 and 2% of interactions, respectively (reads that included sequences from both the 5′ UTR and CDS were categorized as CDS). The free sRNA pool can be “buffered” by sRNA–tRNA interactions (Lalaouna et al, 2015), which represented 3.5% of interactions in our dataset. In addition, sRNA interactions were recovered with rRNAs (35%) and other ncRNAs (6S, tmRNA, RnpB RNA, CsrB; 0.9%). Hybrids between different sRNA species were recovered, for both sRNAs encoded in the “core” genome (1.8%, 87 interactions) and pathogenicity islands (0.6%, 29 interactions), indicating an extensive sRNA–sRNA interaction network. These included the previously identified interaction between the bacteriophage‐encoded anti‐sRNA, AgvB, and the conserved core sRNA GcvB (82 unique hybrids) (Tree et al, 2014). Small RNAs can also be generated from the 3′ UTRs of mRNAs (Guo et al, 2014; Miyakoshi et al, 2015). 0.9% of hybrids with sRNAs mapped within 50 nt downstream of mRNA translation termination sites, potentially reflecting interactions involving 3′ UTRs or 3′ UTR‐derived sRNAs. For all RNA classes presented in Fig 2F, the distribution of free energies of interacting RNAs was significantly lower than randomly paired hybrid halves (P < 1 × 10−9).
Our results indicate that sRNA–mRNA interactions recovered by RNase E‐CLASH have significantly lower free energy than randomly paired RNA sequences and are predominately found close to the start codon, consistent with these hybrid sequences originating from in vivo sRNA–mRNA interactions. Numerous sRNA interactions were recovered with diverse ncRNA classes, including sRNA, rRNA, tRNA and other ncRNAs, revealing a complex network of sRNA interactions.
Filtering functionally relevant RNA–RNA interactions
Proximity‐dependent ligation protocols can potentially yield false‐positive data through spurious ligation events, mapping artefacts or errors introduced during reverse transcription and PCR (Ramani et al, 2015). Since highly recovered interactions have a higher percentage of true positives (Ramani et al, 2015), ligation events can be weighted on the number of unique sequencing reads corresponding to individual interactions. We additionally used known and predicted attributes of sRNA–mRNA interactions to prioritize interactions for further analysis. This was based on (i) the number of unique sequence reads corresponding to the interaction; (ii) detection of the interaction in replicate datasets; (iii) recovery of the hybrid sequences in both RNA1–RNA2 and RNA2–RNA1 orientations, indicating ligation at opposite ends of the duplex; (iv) inclusion of a non‐genomically encoded oligo(A) tail at the 3′ end of the target RNA sequence, which is indicative of sRNA‐directed cleavage and subsequent tailing; and (v) overlap of both hybrid regions with Hfq binding sites determined by UV‐crosslinking and indicating Hfq dependence (see Appendix Supplementary Methods). We confirmed that experimentally verified sRNA–mRNA interactions had a higher distribution of scores compared to total sRNA–mRNA interactions recovered when applying these criteria (Fig EV5 and Appendix Supplementary Methods).
Figure EV5. To determine whether the scoring criteria provided a useful ranking of functional sRNA–mRNA interactions, we looked at the distribution of scores assigned to 14 experimentally verified sRNA–mRNA interactions (see Table EV5).

-
A–DCumulative distribution functions for the total (unverified sRNA–mRNA) interactions (black) and for verified sRNA–mRNA interactions. Verified sRNA–mRNA interactions were found to have a significantly lower distribution of interaction strength (free energy of interaction, ∆G; panel A). For our scoring criteria, we looked the effectiveness of ranking interactions by the number of unique reads recovered (B), by applying scoring criteria 1–4 (i.e. excluding an overlap with a Hfq binding site) (C), or for scoring interactions on all five criteria (D). We applied a two‐sample Kolmogorov–Smirnov test to each CDF (p indicated in plot) and found that scoring interactions on all five criteria gave the most significant separation of verified sRNA–mRNA interactions.
Strikingly, sRNA interactions that satisfied all five criteria, and were represented by multiple unique hybrid reads, were recovered for all RNA classes examined: mRNA, tRNA, rRNA, ncRNA, sRNA (both core and pathogen specific [EcOnc]) and mRNA antisense transcripts (Fig 3). The sRNA interactions with the most hybrid reads representing an interaction were with tRNA species and these interactions were also coincident with Hfq binding sites, indicating that tRNA is a major target for a subset of sRNAs.
Figure 3. RNase E‐CLASH recovers RNA–RNA interactions between diverse RNA classes.

RNA classes are labelled on the outer ring (rRNA has been omitted to clarify interactions between other RNA classes). RNA–RNA interactions with more than two unique hybrid sequences are presented. The thickness of the link represents the number of unique hybrid sequences recovered (up to a maximum of 50 sequences). RNA–RNA interactions where both hybrid halves overlap an Hfq binding site are coloured blue and non‐overlapping interactions are coloured green.
Several characterized sRNAs target functionally related sets of mRNAs, allowing coordinated adaption of the transcriptome in response to specific challenges. Functionally related clusters of mRNA targets within an sRNA interactome may therefore constitute a further indication of reliability, as well as providing insights into the biological roles of the sRNAs involved. We therefore clustered functionally related sRNA interactions with a score of ≥ 1.1 using BiNGO (Maere et al, 2005) (Appendix Supplementary Methods). Consistent with previous reports (Sharma et al, 2011), targets for the core sRNA GcvB were enriched for mRNAs involved in branched‐chain amino acid metabolism. The targets of seven other sRNAs showed significant enrichment of specific ontology classes (Table EV6). In particular, the EHEC‐specific sRNA Esr41 (EcOnc14 in our earlier analysis) was significantly enriched for targets annotated as “signal transduction”. Esr41 bound three mRNAs with products involved in iron uptake: CirA (receptor for the iron‐binding, catecholate siderophore), ChuA (haem receptor) and Bfr (bacterioferritin), which were analysed in more detail (see below). These results indicate that functionally related sRNA targets can be defined using gene ontology and are a further indicator of reliability.
sRNA–RNA interactions define seed motifs
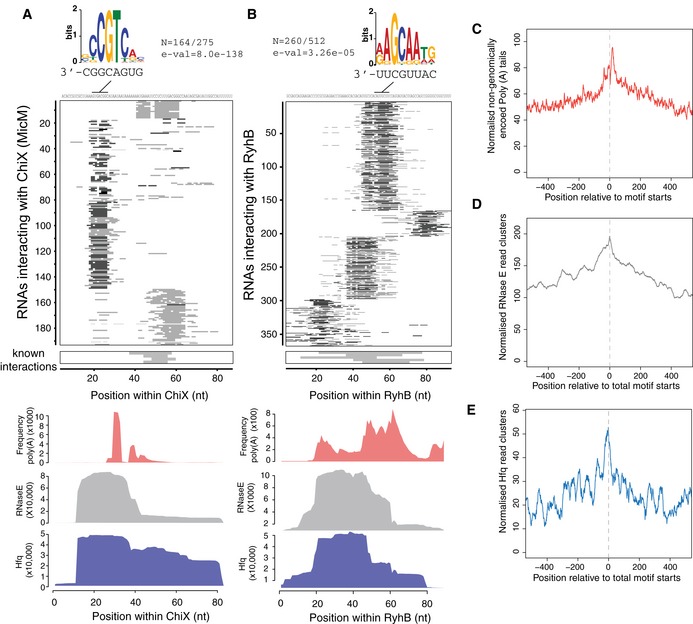
Within characterized sRNAs, a single “seed sequence” can initiate binding to multiple, distinct RNA targets. However, between sRNAs the seeds are heterogeneous in location and sequence, making them difficult to predict using only bioinformatic approaches (Peer & Margalit, 2011; Backofen et al, 2014). To identify putative, novel sRNA seed regions, we analysed sRNA–target RNA interactions. The base‐paired nucleotides between each sRNA and target RNA were predicted by folding the hybrid read in silico using the UNAfold suite of tools. The base‐paired nucleotides within the sRNA were plotted for each interaction (Fig 4 and Appendix Fig S1). Conserved sites of target base pairing were considered to be a seed region. Multiple seed regions were apparent in the sRNAs ChiX, RyhB, ArcZ, GadY, MgrR and Spot42. The motif discovery tool MEME (Bailey & Elkan, 1994) was then applied to identify conserved sequence motifs within target mRNAs that might be recognized by each sRNA seed. Highly enriched motifs were identified (e‐value < 10−4) within target RNAs for 12 sRNAs. GcvB was reported to recognize the consensus motif CACAaCAY in mRNAs through interactions with the GU‐rich R1 seed region located at bases 66–89 (Sharma et al, 2011). We found that GcvB–target interactions were positioned within this R1 seed region (Appendix Fig S1D) and MEME identified the consensus motif ACAATAWC within GcvB‐targeted RNAs that has complementary to bases 69–76 of the GcvB R1 seed region (Appendix Fig S1D). The consensus motif suggests that base G72 of GcvB frequently participates in G‐U wobble interactions. For the 12 sRNAs with statistically significant target motifs, a complementary sequence was identified within the sRNA and likely represents a seed sequence (Fig 4A and B and Appendix Fig S1).
Figure 4. sRNA–RNA interactions identify sRNA seed sequences.
-
A, B(Top) Sequences interacting with ChiX (A) or RyhB (B) were analysed for conserved motifs using MEME. Motifs that were enriched in the target RNAs are shown above the heatmap with proportion of target RNAs carrying the motif (N) and the expected value for the motif (e‐val). The complementary sequence motifs within ChiX or RyhB are shown below the logo. (Middle) Heatmaps showing the position of predicted base pairing for each interaction within ChiX or RyhB. Grey bars boxed below indicate the position of base pairing with experimentally verified mRNAs (known interactions). (Bottom) Hfq (blue) and RNase E (grey) binding sites within ChiX and RyhB. Hfq‐ and RNase E‐bound sequence, and non‐genomically encoded oligo(A) tails (red) within ChiX and RyhB are shown as line plots where the x‐axis position correlates with heatmaps above.
-
C–ECumulative plots of oligoadenylation (C), Hfq (D) and RNase E (E) binding at predicted seed motifs.
The seed sequence of the sRNA–mRNA pair MicC–ompD guides RNase E cleavage 6 nt downstream of the duplex (Bandyra et al, 2012). To determine whether this is a general phenomenon, we cumulatively analysed RNase E binding, oligoadenylation and Hfq binding relative to statistically significant seed motifs identified in target RNAs (Fig 4C–E). Oligo(A) tails were found to be maximally recovered 10 nt from the 3′ end of the seed motif (8‐nt motif length) consistent with seed‐directed RNase E cleavage. Hfq‐bound reads were maximally recovered in the 10 nt 5′ to the seed motif, indicating that Hfq binding sites are often closely associated with the identified seed motifs.
Our results experimentally define seed motifs for sRNAs with multiple interactions and demonstrate that many sRNAs use more than one site for target RNA interactions. The newly identified sRNA seed motifs appear to direct RNase E cleavage and oligoadenylation of target RNAs at sites 3′ of the seed interaction.
Functional testing of sRNA–mRNA interactions
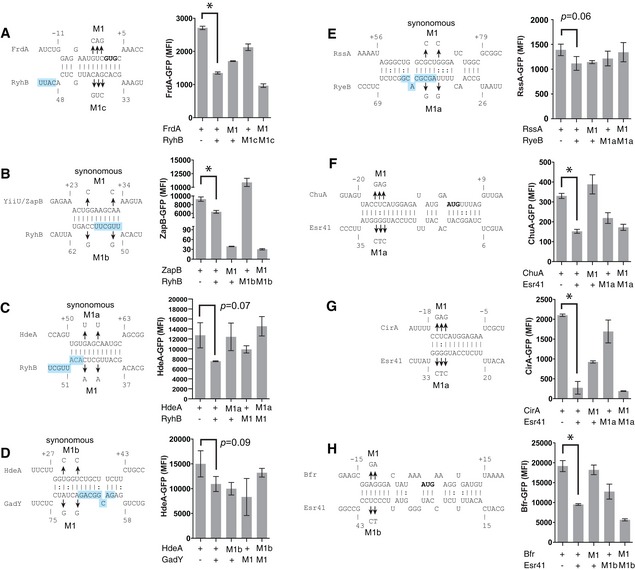
To assess whether sRNA–mRNA interactions defined by RNase E‐CLASH function in regulating gene expression, we used a two‐plasmid system for monitoring translation of superfolder GFP fusions (Corcoran et al, 2012). Translational fusions were constructed for sRNA–mRNAs interactions with high scores, as defined above: hdeA‐RyhB (score = 8.9), zapB‐RyhB (7.6), rssA‐RyeB (7.2), frdA‐RyhB (6.7), hdeA‐GadY (5.8) (Fig 5A–E), and for interactions with lower scores that were supported by the ontological analysis chuA‐Esr41 (4.1), cirA‐Esr41 (3.1) and bfr‐Esr41 (4.2) (Fig 5F–H). Expression levels were reduced for all 8 of the fusions when co‐expressed with the cognate sRNA. Mutations introduced into the mRNAs and sRNAs de‐repressed the frdA‐RyhB and hdeA‐RyhB interactions, and all three Esr41 interactions. Point mutations in RyhB similarly relieved repression of zapB; however, synonymous mutations within the mRNA abolished expression and destabilized the transcript as assessed by qPCR (data not shown). A rare leucine codon was introduced into zapB by the M1 synonymous mutation, potentially explaining the poor translation of this mRNA. Introduction of compensatory mutations restored RyhB control of frdA, and Esr41 control of chuA, cirA and bfr verifying direct sRNA–mRNA interactions for these pairs and confirming that functional sRNA–mRNA interactions are recovered by the RNase E‐CLASH method.
Figure 5. Confirmation of high‐scoring sRNA–mRNA interactions and interactions with the EHEC‐specific sRNA, Esr41.
-
A–H(Left) Base pairing patterns for the sRNA–mRNA pairs identified by CLASH were calculated using IntaRNA software (Busch et al, 2008) and are shown for frdA‐RyhB (A), zapB‐RyhB (B), hdeA‐RyhB (C), hdeA‐GadY (D), rssA‐RyeB (E), chuA‐Esr41 (F), cirA‐Esr41 (G) and bfr‐Esr41 (H). Point mutations and predicted sRNA seed sequences (blue shading) are indicated. (Right) Median fluorescence intensity (MFI) was assessed by FACS for mRNA‐sfGFP fusions with compensatory base changes. In each case, introduction of a point mutation into an sRNA or mRNA construct (M1) is expected to reduce sRNA repression, which should be restored by combining complementary point mutants (last bar, M1‐M1). A two‐tailed t‐test was used to calculate significance from biological triplicate cultures. Error bars represent SEM. *P < 0.05.
The EHEC‐specific sRNA Esr41 controls iron transport and storage
Our previous analysis of Hfq binding sites using UV‐crosslinking identified numerous novel sRNAs within the pathogenicity islands of enterohaemorrhagic E. coli, referred to as EcOnc RNAs, but their RNA targets remained largely unknown (Sudo et al, 2014; Tree et al, 2014). The RNase E‐CLASH dataset contained 810 unique hybrids with pathogenicity island‐encoded EcOnc sRNAs identifying many target transcripts (Fig 3 and Table EV7). The EHEC‐specific sRNA, Esr41 (EcOnc14 in our earlier analysis), was previously shown to affect the abundance of the fliC transcript and cell motility (Sudo et al, 2014). Here we have demonstrated that Esr41 regulates expression of the iron transport and storage proteins CirA, ChuA and Bfr (Fig 5F–H). The mRNA interactome of Esr41 is similar to the “core” genome‐encoded sRNA, RyhB (Massé et al, 2005). We therefore additionally analysed translation of the chuA, cirA and bfr fusions in the presence of constitutively expressed RyhB (Fig 6A). Esr41 and RyhB repressed bfr to comparable levels, but Esr41 had a greater repressive effect on chuA translation, consistent with it base pairing closer to the chuA RBS. In contrast, Esr41 repressed cirA translation by 7.6‐fold, whereas RyhB positively regulated cirA translation.
Figure 6. The EHEC‐specific small RNA Esr41 regulates iron uptake, storage and colicin resistance.

-
A–CFACS analysis of constitutively expressed sfGFP fusions to chuA (A), cirA (B) or bfr (C) in the presence of Esr41 (orange), RyhB (blue) or a scrambled RNA control (red). The histogram shows GFP fluorescence for each sRNA–mRNA fusion.
-
DEsr41 confers resistance to colicins 1A and 1B in the sensitive background, E. coli DH5α. Top agar lawns of DH5α expressing a control scrambled RNA (pJV300), Esr41 (pZE12::esr41) or RyhB (pZE12::ryhB) were spotted with colicins indicated (top). Zones of clearing indicate sensitivity to the tested colicin.
-
EEsr41 confers a competitive disadvantage on EHEC under iron‐limiting conditions. Wild‐type EHEC and isogenic ∆esr41 strain (black), or EHEC ∆esr41 and the chromosomally repaired strain EHEC ∆esr41::esr41 were inoculated at equal densities and cultured in MEM‐HEPES media for 3 days. The proportion of each strain was determined at each time point (days) and is expressed as a ratio relative to the starting inoculum where 1 is an equal fitness (see Appendix Supplementary Methods).
Esr41 is encoded on the pathogenicity island SpLE1 that also encodes the tellurite, phage and colicin resistance gene cluster ter (Whelan et al, 1997), and the enterobactin receptor Iha. Colicin 1A is a pore‐forming toxin that uses the siderophore receptor CirA to enter the cell and cause bacterial cell death. RyhB confers sensitivity to colicin 1A through de‐repression of CirA (Salvail et al, 2013), and we investigated the effect of Esr41 on colicin sensitivity. Constitutive expression of Esr41 conferred complete resistance to colicin 1A in the sensitive E. coli background, DH5α, but did not affect resistance in the EHEC background that is already colicin resistant (Fig 6 and data not shown). Deletion of esr41 in EHEC strain ZAP198 conferred a fitness advantage in iron‐limited medium (MEM‐HEPES supplemented with 250 nM Fe(NO3)3 and 0.1% glucose) consistent with repression of iron transporters by Esr41 (Fig 6E). Complementation of the esr41 mutant by chromosomal knock‐in of esr41 restored the growth disadvantage to the esr41 mutant.
These results demonstrate that, consistent with mRNA interactions identified by RNase E‐CLASH, Esr41 regulates iron uptake and homeostasis in EHEC and can confer resistance to colicin 1A and colicin 1B in a sensitive background.
Discussion
We demonstrate that interaction networks for bacterial sRNAs can be determined experimentally by UV‐crosslinking sRNA–target RNA duplexes to RNase E. Our results revealed sRNA interactions with diverse RNAs including stable RNA species: rRNA and tRNA, other non‐coding RNAs, and many different mRNAs. Here we have focused on the association of RNase E with sRNA–mRNA duplexes. The CLASH analyses of RNase E‐associated RNA duplexes recovered around 0.8% hybrids. This frequency is similar to that seen in previous analyses of human miRNAs associated with Argonaute 1 (Ago1) (Helwak et al, 2013) and double‐stranded RNAs bound to Staufen (Sugimoto et al, 2015). In contrast, analysis of our previous Hfq UV‐crosslinking data identified far fewer hybrids (~0.001% of mapped reads). Consistent with this finding, we previously found that many Hfq binding motifs overlap the mRNA seed sequence, suggesting that for these sRNA–mRNA interactions, duplex formation would likely dissociate the RNAs from Hfq (Tree et al, 2014). We therefore postulated that duplexes formed on Hfq are rapidly transferred to RNase E.
For a subset of sRNAs, we were able to define seed sequences within the sRNA and identify enriched motifs within target RNAs. Our analyses indicate that sRNAs commonly utilize multiple seed regions for target RNA base pairing. Target RNA seed sequences were closely associated with Hfq binding sites. This is consistent with our earlier model that duplex formation will render many Hfq binding motifs double stranded, promoting release of the base‐paired RNAs and preventing re‐binding to Hfq (Fig 4E). Oligoadenylation peaked 10 nt 3′ of the seed motif, indicating that many seed interactions direct cleavage of the mRNA and terminal nucleotide addition by poly(A) polymerase or PNPase (Fig 4C). This is consistent with in vitro results demonstrating RNase E cleavage of target RNAs is guided to 5–6 nt 3′ of a duplexed 13‐mer or sRNA (Bandyra et al, 2012).
The mechanism of sRNA‐directed, RNase E cleavage has features in common with miRNA‐directed cleavage by human Argonaute 2 (hAgo2). RNA targets that are fully complementary to the miRNA displace the PAZ domain of hAgo2 and induce a conformational change that results in cleavage of the miRNA–target duplex (Ameres et al, 2007; Wang et al, 2009). Thus, productive base pairing of the miRNA and target is sensed by competition between hAgo2 and the target RNA resulting in dissociation of the miRNA 3′ end. For the Hfq‐RNase E complex, we suggest that sRNA–mRNA duplex formation at the Hfq binding motif dissociates the sRNA–mRNA pair from Hfq allowing interaction with RNase E and sRNA‐directed cleavage of the target RNA 3′ of the seed motif.
A striking result from our RNase E‐CLASH analysis was the range of RNA classes identified in RNA–RNA hybrids. The transcriptomes of both E. coli and Salmonella encode small RNAs embedded within mRNAs (Guo et al, 2014; Miyakoshi et al, 2015) lending weight to the idea of a genomic palimpsest even in prokaryotes (Tuck & Tollervey, 2011) and potentially obscuring clear annotation of transcript classes. However, it is notable that all classes of RNA analysed were found in sRNA–RNA duplexes. We and others have identified small RNA species that act as sRNA sponges and this appears to be widespread. We recovered 152 unique sRNA–sRNA interactions in our CLASH data. These included our previously characterized interaction between the pathogenicity‐associated sRNA AgvB and core sRNA GcvB (Tree et al, 2014). These results indicate that an extensive network of sponging interactions occur between sRNAs. Recent work demonstrated that sRNA interactions with tRNA spacer regions play important roles in “buffering” sRNA interactions to enhance specificity (Lalaouna et al, 2015). We identified 320 unique sRNA–tRNA interactions, including the previously reported RyhB–tRNA–Leu interaction (Lalaouna et al, 2015). We note that six sRNA–tRNA interactions contain > 10 nt of pre‐tRNA sequence, indicating that minimally, these interactions occur before tRNA 5′ and 3′ maturation. Hfq has previously been shown to interact with tRNAs (Zhang et al, 2003; Lee & Feig, 2008; Tree et al, 2014), suggesting a role in facilitating sRNA–tRNA interactions. Extensive interactions of miRNAs with tRNA and rRNA have also been identified (Helwak et al, 2013) and it seems that these stable RNA species may act universally to buffer non‐coding RNA interactions. These may stabilize sRNAs or miRNAs that are temporarily in excess over cognate targets and help prevent their inappropriate binding elsewhere.
The EHEC‐specific sRNA Esr41/EcOnc14 was independently identified by Sudo et al (2014) and in our previous analysis of Hfq binding sites. We initially investigated the role of Esr41 in promoting colicin resistance through repression of CirA, and we were able to confirm that Esr41 confers complete colicin 1A and colicin 1B resistance when provided in trans in the colicin‐sensitive background, DH5α. Colicin 1B is used by Salmonella Typhimurium to clear commensal Escherichia coli species (part of the normal flora) during gastrointestinal colonization (Nedialkova et al, 2014). Our results demonstrate that resistance to colicin 1B can be conferred by expression of a single, pathogen‐specific small RNA. In contrast, the core genome‐encoded sRNA RyhB promotes colicin 1A sensitivity through translational activation of CirA (Salvail et al, 2013).
Esr41 is encoded within a large pathogenicity island (SpLE1 or O‐island 43/48) that confers colicin, tellurite and bacteriophage resistance, and also encodes the iron transporter/adhesin Iha. We were not able to test for decreased colicin 1A sensitivity in an EHEC ∆esr41 strain due to the presence of the adjacent colicin resistance ter gene cluster. However, Esr41 targets identified by CLASH and confirmed by mutations included mRNAs encoding the iron transport and storage proteins ChuA, CirA and Bfr. A role in iron homeostasis is corroborated by competitive index experiments, demonstrating that deletion of esr41 confers a fitness advantage to EHEC under relatively iron‐limited conditions (250 nM Fe), indicating that Esr41 limits iron transport by repression of select iron receptors. The Iha gene is located upstream of Esr41 and encodes a receptor for the ferric iron‐binding siderophore, enterobactin. We speculate that Esr41 is co‐selected with Iha as Esr41‐mediated repression of CirA (catecholate siderophore receptor), ChuA (haem receptor) and Bfr (bacterioferritin) would redirect iron transport through a pathway involving enterobactin and Iha, favouring maintenance of the O‐island.
While this work was in revision, a related technique for sequencing sRNA–RNA interactions termed RIL‐Seq was described (Melamed et al, 2016). This is conceptually similar to RNase E‐CLASH, excepting that Hfq is used as a scaffold to capture sRNA–RNA duplexes and the purification is performed under native conditions as opposed to CLASH that uses a stringent purification protocol. Stringency is introduced into RIL‐Seq analysis in silico where hybrid reads are filtered for statistical enrichment. We find a comparable number of statically significant sRNA–mRNA interactions are recovered by both techniques in log phase cells (633 using RIL‐Seq and 782 using RNase E‐CLASH) and similar sRNA seed regions and motifs are recovered for abundant sRNAs (e.g. ArcZ, MgrR, GcvB and CyaR), suggesting that both techniques capture bona fide sRNA–RNA interactions. Notably, the pools of RNA–RNA interactions recovered in association with Hfq and RNase E are expected to be different. RNase E processes a broad range of RNA species and is expected to associate with a subset of all sRNA–mRNA interactions that specifically result in target degradation.
We conclude that CLASH recovers functional RNA–RNA interactions when applied to RNase E in E. coli, allowing high‐throughput identification of functional RNA targets for many sRNA species. A key advantage of this high‐throughput approach is the ability to identify interactions that would not be predicted by extrapolating our current understanding of sRNA biology. We anticipate that profiling RNA interactions using CLASH will reveal diverse roles for both coding and non‐coding RNAs in cell physiology.
Materials and Methods
Bacterial strains, plasmids and culture conditions
For CLASH analysis, Escherichia coli O157:H7 str. Sakai (GenBank Acc# NC_002695.1) was used to construct a dual‐affinity‐tagged HTF strain. Bacterial strains, plasmids and primers are presented in Table EV8. Strains were routinely grown on LB agar plates and broth supplemented with antibiotics where appropriate. For crosslinking and phenotypic experiments, E. coli O157:H7 was grown under virulence‐inducing conditions in MEM‐HEPES media (Sigma M7278) supplemented with 250 nM Fe(NO3)3 and 0.1% glucose.
Preparation of CLASH sequencing libraries
Cells grown to OD 0.8 in MEM‐HEPES (M7278) supplemented with 250 nM Fe(NO3)3 and 0.1% glucose were crosslinked with 1,800 mJ of UV‐C. Cells were harvested by centrifugation at 4,000 g for 10 min, weighed and resuspended in 50 ml of ice‐cold PBS. The cells were divided into 1 g pellets and snap‐frozen in a dry ice/ethanol bath. One volume (1 ml/g) of lysis buffer [50 mM Tris–HCl (pH 7.8), 1.5 mM MgCl2, 150 mM NaCl, 0.1% Nonidet P‐40, 5 mM β‐mercaptoethanol and 1 tablet “cOmplete” EDTA‐free protease inhibitor (Roche)/50 ml] and 3 V of 0.1‐mm zirconia beads were added to a cell pellet and vortexed 5 × 1 min with 1‐min intervals on ice. Cell lysates were cleared by centrifugation (4,000 g for 20 min) and the supernatant was transferred to 1.5‐ml microcentrifuge tubes and cleared at 16,000 g for a further 20 min. Supernatants were added to 200 μl of pre‐washed M2 anti‐FLAG resin (Sigma‐Aldrich) and incubated overnight. The resin was washed twice with 10 ml of TNM1000 (50 mM Tris–HCl pH 7.8, 1 M NaCl, 0.1% NP‐40, 5 mM β‐mercaptoethanol) and twice in 10 ml TMN150 (50 mM Tris–HCl pH 7.8, 150 mM NaCl, 0.1% NP‐40, 5 mM β‐mercaptoethanol), resuspended in 500 μl of TMN150 and incubated with 20–30 U of TEV protease for 2 h at 18°C. The slurry was centrifuged through a Bio‐Rad Bio‐spin column and the eluate collected. Approximately 500 μl of eluate was incubated with 0.15 U of RNace‐IT (Agilent) at 20°C for 7 min. The digestion was stopped by the addition of 0.4 g of guanidine–HCl, 300 mM NaCl and 10 mM imidazole (pH 8.0). 100 μl of Ni‐NTA slurry was pre‐washed twice in 750 μl of wash buffer I (6 M guanidine–HCl, 50 mM Tris–HCl pH 7.8, 300 mM NaCl, 0.1% NP‐40 and 5 mM β‐mercaptoethanol). Eluates were added to the washed resin and incubated overnight at 4°C. The resin was washed twice with 750 μl of ice‐cold wash buffer I and twice with 750 μl of 1× PNK buffer (50 mM Tris–HCl pH 7.8, 10 mM MgCl2, 0.5% NP‐40 and 5 mM β‐mercaptoethanol). The eluates were transferred into a spin column (Pierce, Thermo Fisher, 69705). The subsequent reactions were performed in 80 μl reaction volumes on‐column. 3′ ends were dephosphorylated by incubating for 45 min at 20°C with thermosensitive alkaline phosphatase (TSAP, Promega) and RNasin (Promega) in PNK reaction buffer (50 mM Tris–HCl pH 7.8, 10 mM MgCl2 and 10 mM β‐mercaptoethanol). The resin was washed once with 400 μl of wash buffer I and three times with 400 μl of 1× PNK buffer. The resin was incubated with tobacco acid pyrophosphatase (Epicentre) in 1× TAP buffer (Epicentre) and incubated at 20°C for 2 h, washed once with 400 μl of wash buffer I and then three times with 400 μl of 1× PNK buffer. The 5′ ends of bound RNAs were radiolabelled by phosphorylation with T4 PNK (4 μl, Sigma) and 32P‐γATP (4 μl, PerkinElmer BLU502Z) in PNK reaction buffer for 100 min at 20°C, after which 100 nM of cold ATP was added and incubated for a further 50 min to complete 5′ end phosphorylation. The resin was washed once with 400 μl of wash buffer I and three times with 400 μl of 1× PNK buffer. To add 3′ linkers, the resin was incubated with 4 μl of T4 RNA ligase I (NEB) and 8 μl of miRCat‐33 3′ linker (IDT) in PNK reaction buffer with 2 μl of RNasin (Promega) at 16°C for 16 h and then washed once with 400 μl of wash buffer I and three times with 1× PNK buffer. To add 5′ linkers, the resin was incubated with 4 μl of T4 RNA ligase I (NEB) and 1 μl of 100 μM 5′ linker (IDT; Table EV8) in PNK reaction buffer with 2 μl of RNasin (Promega) and 1 mM ATP at 16°C for 16 h. The resin was washed three times with wash buffer II (50 mM Tris–HCl pH 7.8, 50 mM NaCl, 10 mM imidazole, 0.1% NP‐40, 5 mM β‐mercaptoethanol). 200 μl of elution buffer (wash buffer II supplemented with 150 mM imidazole) was added to the resin and incubated at RT for 5 min. RNase E–RNA complexes were eluted into a clean microcentrifuge tube, and the elution was repeated. Complexes were precipitated with 100 μl of TCA and 40 μg of glycogen by incubating on ice for 30–60 min and centrifugation at 4°C for 20 min (16,000 g). Supernatants were removed and pellets washed with 800 μl of ice‐cold acetone. Precipitate was centrifuged again at 16,000 g, supernatants were removed, and pellets were air‐dried. The pellet was resuspended in 30 μl of 1× NuPAGE loading buffer. The sample was loaded onto a NuPAGE 4–12% Bis‐Tris PAGE gel (Invitrogen) and run in MOPS SDS running buffer (Invitrogen). 32P‐labelled RNase E complexes were transferred to a nitrocellulose membrane (Amersham Hybond ECL) by wet transfer using a Bio‐Rad mini‐Trans blot module in NuPAGE transfer buffer (Invitrogen). Complexes were visualized by autoradiography using Kodak BioMax MS film and developed films realigned to the membrane. The high molecular weight complex (> 115 kDa) was excised from the membrane (see Fig EV1C). The labelled RNA was recovered by incubating the membrane fragment in 400 μl of wash buffer II supplemented with 1% SDS, 5 mM EDTA and 100 μg of proteinase K, for 2 h at 55°C. The supernatant containing labelled RNA fragments was transferred to a clean microcentrifuge tube. To precipitate the RNA fragments, 50 μl of 3 M NaOAc pH 5.2 and 500 μl of phenol:chloroform:isoamylalcohol was added, vortexed and centrifuged for 5 min at RT. The aqueous phase was transferred to a clean microcentrifuge tube and 1 ml of ice‐cold EtOH and 20 μg of glycogen added. The precipitation was incubated at −80°C for 30 min and centrifuged at 16,000 g for 20 min, followed by a wash with 500 μl of ice‐cold 70% EtOH and air‐drying. The RNA pellet was resuspended in 13 μl of RT buffer I (miRCat RT oligo and 5 mM dNTPs) and reverse‐transcribed using Superscript III as per the manufacturer's instructions. cDNA was amplified using Takara LA Taq, P5 and PE_miRCat PCR primers (Table EV8), and 2 μl of cDNA. cDNAs were amplified for 20–24 cycles to minimize bias in amplicons. 3–10 PCRs were pooled and ethanol‐precipitated. PCR products were separated on a 3% metaphor agarose gel and smeared amplicons above primer dimers indicated in control samples were gel‐extracted using a MinElute gel extraction Kit (Qiagen). Libraries were pooled and submitted for single‐end 100‐bp HiSeq2500 sequencing at GenePool (University of Edinburgh). Sequence data has been deposited at NCBI GEO (series GSE77463).
Analysis of CLASH hybrids
Sequencing reads generated by RNase E‐CLASH were analysed using the hyb package (Travis et al, 2014). Details of the in silico analysis are presented in Appendix Supplementary Methods.
Confirmation of sRNA–mRNA interactions and phenotypic characterization of Esr41
We employed the two‐plasmid system described by Corcoran et al (2012) to monitor translation efficiency of mRNA‐sfGFP fusions. Plasmids containing small RNAs were cloned as described in Urban and Vogel (2007) excepting Esr41 was inserted into pZE12 using inverse PCR. Briefly, the mutagenic primers Esr41.ZE12.F and ZE12.5P.R were used to amplify a fragment of pZE12::luc that was DpnI‐treated, gel‐extracted and subsequently recircularized with T4 DNA ligase and transformed into DH5α. Clones containing an Esr41 insert were confirmed by sequencing. For mRNA fusions, clones were generated essentially as described in Corcoran et al (2012). Briefly, transcript start sites were identified using RegulonDB and the corresponding site in E. coli O157:H7 str. Sakai identified using BLAST. Primers were designed to amplify from the transcription start site to within the CDS encompassing the predicted region of sRNA–mRNA interaction (Table EV8). PCR products were cloned using NsiI and NheI (Fast digest enzymes, Thermo) and positive clones confirmed by sequencing. Point mutations were introduced using mutagenic primers listed in Table EV8 and confirmed by Sanger sequencing. Detailed methods for FACS and qPCR analysis of superfolder GFP fusions are presented in Appendix Supplementary Methods.
Competitive index experiments
Indicated strains were grown overnight in LB at 37°C and the culture OD600 adjusted to provide equal densities. Competing strains were inoculated at 1/1,000 into MEM‐HEPES supplemented with 250 nM Fe(NO3)3 and 0.1% glucose. At 24‐h intervals, the culture was diluted 1/1,000 in fresh media for a total of three subcultures (3‐days growth). Cultures were diluted and plated onto LB plates to obtain well‐separated colonies and 100 colonies were replica plated onto LB agar and LB agar supplemented with nalidixic acid (30 μg/ml) to select for marked strains. Competition experiments were repeated with nalidixic acid resistance and sensitivity in the opposite strain to account for any fitness cost associated with nalidixic acid resistance.
Colicin sensitivity testing
Colicin 1A and B lysates were prepared from E. coli harbouring p3Z/Col1A and p3Z/ColB as described in Brickman and Armstrong (1996). Colicin 1B was prepared from Salmonella Typhimurium SL1344 by inducing with 1 μg/ml of mitomycin C and filtering the supernatant. Colicin V was prepared from E. coli strain NCTC50147 (Public Health England, UK) as described for colicin 1B. To test sensitivity to colicins, a top agar lawn of E. coli DH5α was prepared and 5 μl of colicin lysate spotted onto the lawn. Plates were incubated overnight at 37°C and scanned.
Author contributions
JJT, DLG and DT designed the experiments. SAW, JJT, SPM and KWL performed the experimental work. JJT, GK, NPD, IP and TGA analysed the data. All authors contributed to writing and editing the manuscript.
Conflict of interest
The authors declare that they have no conflict of interest.
Supporting information
Appendix
Expanded View Figures PDF
Table EV1
Table EV2
Table EV3
Table EV4
Table EV5
Table EV6
Table EV7
Table EV8
Source Data for Figure EV1
Review Process File
Acknowledgements
We thank Eric Masse for providing constructs for expressing colicins 1A and B. JJT and SAW were supported by funding from the Australian National Health and Medical Research Council (APP1067241). DLG, DT and SPM were supported by Wellcome Trust funding (WT090231MA) and research at the Roslin Institute is supported by BBSRC Institute grant funding (BB/J004227/1). DT was supported by Wellcome Trust funding (077248). Work in the Wellcome Trust Centre for Cell Biology is supported by Wellcome Trust core funding (092076). GK was supported by Wellcome Trust grant 097383 and by the MRC. MRW acknowledges funding from the Australian Government NCRIS scheme and the New South Wales State Government RAAP scheme.
The EMBO Journal (2017) 36: 374–387
See also: J Hör & J Vogel (February 2017)
Contributor Information
David Tollervey, Email: d.tollervey@ed.ac.uk.
Jai J Tree, Email: j.tree@unsw.edu.au.
References
- Ameres SL, Martinez J, Schroeder R (2007) Molecular basis for target RNA recognition and cleavage by human RISC. Cell 130: 101–112 [DOI] [PubMed] [Google Scholar]
- Backofen R, Amman F, Costa F, Findeiß S, Richter AS, Stadler PF (2014) Bioinformatics of prokaryotic RNAs. RNA Biol 11: 470–483 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bailey TL, Elkan C (1994) Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol 2: 28–36 [PubMed] [Google Scholar]
- Bandyra KJ, Said N, Pfeiffer V, Górna MW, Vogel J, Luisi BF (2012) The seed region of a small RNA drives the controlled destruction of the target mRNA by the endoribonuclease RNase E. Mol Cell 47: 943–953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bossi L, Schwartz A, Guillemardet B, Boudvillain M, Figueroa‐Bossi N (2012) A role for Rho‐dependent polarity in gene regulation by a noncoding small RNA. Genes Dev 26: 1864–1873 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouvier M, Sharma CM, Mika F, Nierhaus KH, Vogel J (2008) Small RNA binding to 5′ mRNA coding region inhibits translational initiation. Mol Cell 32: 827–837 [DOI] [PubMed] [Google Scholar]
- Brickman TJ, Armstrong SK (1996) Colicins B and Ia as novel counterselective agents in interspecies conjugal DNA transfers from colicin‐sensitive Escherichia coli donors to other Gram‐negative recipient species. Gene 178: 39–42 [DOI] [PubMed] [Google Scholar]
- Busch A, Richter AS, Backofen R (2008) IntaRNA: efficient prediction of bacterial sRNA targets incorporating target site accessibility and seed regions. Bioinformatics 24: 2849–2856 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chao Y, Vogel J (2010) The role of Hfq in bacterial pathogens. Curr Opin Microbiol 13: 24–33 [DOI] [PubMed] [Google Scholar]
- Clarke JE, Kime L, Romero AD, McDowall KJ (2014) Direct entry by RNase E is a major pathway for the degradation and processing of RNA in Escherichia coli . Nucleic Acids Res 42: 11733–11751 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corcoran CP, Podkaminski D, Papenfort K, Urban JH, Hinton JCD, Vogel J (2012) Superfolder GFP reporters validate diverse new mRNA targets of the classic porin regulator, MicF RNA. Mol Microbiol 84: 428–445 [DOI] [PubMed] [Google Scholar]
- Diwa A, Bricker AL, Jain C, Belasco JG (2000) An evolutionarily conserved RNA stem–loop functions as a sensor that directs feedback regulation of RNase E gene expression. Genes Dev 14: 1249–1260 [PMC free article] [PubMed] [Google Scholar]
- Figueroa‐Bossi N, Valentini M, Malleret L, Bossi L (2009) Caught at its own game: regulatory small RNA inactivated by an inducible transcript mimicking its target. Genes Dev 23: 2004–2015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghora BK, Apirion D (1978) Structural analysis and in vitro processing to p5 rRNA of a 9S RNA molecule isolated from an rne mutant of E. coli . Cell 15: 1055–1066 [DOI] [PubMed] [Google Scholar]
- Gottesman S, Storz G (2011) Bacterial small RNA regulators: versatile roles and rapidly evolving variations. Cold Spring Harb Perspect Biol 3: a003798 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grosswendt S, Filipchyk A, Manzano M, Klironomos F, Schilling M, Herzog M, Gottwein E, Rajewsky N (2014) Unambiguous Identification of miRNA: target site interactions by different types of ligation reactions. Mol Cell 54: 1042–1054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo MS, Updegrove TB, Gogol EB, Shabalina SA, Gross CA, Storz G (2014) MicL, a new σE‐dependent sRNA, combats envelope stress by repressing synthesis of Lpp, the major outer membrane lipoprotein. Genes Dev 28: 1620–1634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helwak A, Kudla G, Dudnakova T, Tollervey D (2013) Mapping the human miRNA interactome by CLASH reveals frequent noncanonical binding. Cell 153: 654–665 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmqvist E, Reimegård J, Sterk M, Grantcharova N, Römling U, Wagner EGH (2010) Two antisense RNAs target the transcriptional regulator CsgD to inhibit curli synthesis. EMBO J 29: 1840–1850 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikeda Y, Yagi M, Morita T, Aiba H (2011) Hfq binding at RhlB‐recognition region of RNase E is crucial for the rapid degradation of target mRNAs mediated by sRNAs in Escherichia coli . Mol Microbiol 79: 419–432 [DOI] [PubMed] [Google Scholar]
- Imig J, Brunschweiger A, Mmer ABU, Guennewig B, Mittal N, Kishore S, Tsikrika P, Gerber AEP, Zavolan M, Hall J (2015) miR‐CLIP capture of a miRNA targetome uncovers a lincRNA H19‐miR‐106a interaction. Nat Chem Biol 11: 107–114 [DOI] [PubMed] [Google Scholar]
- Khemici V, Carpousis AJ (2004) The RNA degradosome and poly(A) polymerase of Escherichia coli are required in vivo for the degradation of small mRNA decay intermediates containing REP‐stabilizers. Mol Microbiol 51: 777–790 [DOI] [PubMed] [Google Scholar]
- Künne T, Swarts DC, Brouns SJJ (2014) Planting the seed: target recognition of short guide RNAs. Trends Microbiol 22: 74–83 [DOI] [PubMed] [Google Scholar]
- Lalaouna D, Carrier M‐C, Semsey S, Brouard J‐S, Wang J, Wade JT, Massé E (2015) A 3′ external transcribed spacer in a tRNA transcript acts as a sponge for small RNAs to prevent transcriptional noise. Mol Cell 58: 393–405 [DOI] [PubMed] [Google Scholar]
- Lee T, Feig AL (2008) The RNA binding protein Hfq interacts specifically with tRNAs. RNA 14: 514–523 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackie GA (1998) Ribonuclease E is a 5′‐end‐dependent endonuclease. Nature 395: 720–723 [DOI] [PubMed] [Google Scholar]
- Mackie GA (2013) RNase E: at the interface of bacterial RNA processing and decay. Nat Rev Microbiol 11: 45–57 [DOI] [PubMed] [Google Scholar]
- Maere S, Heymans K, Kuiper M (2005) BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 21: 3448–3449 [DOI] [PubMed] [Google Scholar]
- Massé E, Vanderpool CK, Gottesman S (2005) Effect of RyhB small RNA on global iron use in Escherichia coli . J Bacteriol 187: 6962–6971 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melamed S, Peer A, Faigenbaum‐Romm R, Gatt YE, Reiss N, Bar A, Altuvia Y, Argaman L, Margalit H (2016) Global mapping of small RNA‐target interactions in bacteria. Mol Cell 63: 884–897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyakoshi M, Chao Y, Vogel J (2015) Cross talk between ABC transporter mRNAs via a target mRNA‐derived sponge of the GcvB small RNA. EMBO J 34: 1478–1492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morita T, Maki K, Aiba H (2005) RNase E‐based ribonucleoprotein complexes: mechanical basis of mRNA destabilization mediated by bacterial noncoding RNAs. Genes Dev 19: 2176–2186 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nedialkova LP, Denzler R, Koeppel MB, Diehl M, Ring D, Wille T, Gerlach RG, Stecher B (2014) Inflammation fuels colicin Ib‐dependent competition of Salmonella serovar Typhimurium and E. coli in enterobacterial blooms. PLoS Pathog 10: e1003844 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panja S, Schu DJ, Woodson SA (2013) Conserved arginines on the rim of Hfq catalyze base pair formation and exchange. Nucleic Acids Res 41: 7536–7546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papenfort K, Sun Y, Miyakoshi M, Vanderpool CK, Vogel J (2013) Small RNA‐mediated activation of sugar phosphatase mRNA regulates glucose homeostasis. Cell 153: 426–437 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papenfort K, Vogel J (2014) Small RNA functions in carbon metabolism and virulence of enteric pathogens. Front Cell Infect Microbiol 4: 91 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papenfort K, Espinosa E, Casadesús J, Vogel J (2015) Small RNA‐based feedforward loop with AND‐gate logic regulates extrachromosomal DNA transfer in Salmonella . Proc Natl Acad Sci USA 25: E4772–E4781 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peer A, Margalit H (2011) Accessibility and evolutionary conservation mark bacterial small‐RNA target‐binding regions. J Bacteriol 193: 1690–1701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pfeiffer V, Papenfort K, Lucchini S, Hinton JCD, Vogel J (2009) Coding sequence targeting by MicC RNA reveals bacterial mRNA silencing downstream of translational initiation. Nat Struct Mol Biol 16: 840–846 [DOI] [PubMed] [Google Scholar]
- Prévost K, Desnoyers G, Jacques JF, Lavoie F, Massé E (2011) Small RNA‐induced mRNA degradation achieved through both translation block and activated cleavage. Genes Dev 25: 385–396 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramani V, Qiu R, Shendure J (2015) High‐throughput determination of RNA structure by proximity ligation. Nat Biotechnol 33: 980–984 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rasmussen AA, Eriksen M, Gilany K, Udesen C, Franch T, Petersen C, Valentin‐Hansen P (2005) Regulation of ompA mRNA stability: the role of a small regulatory RNA in growth phase‐dependent control. Mol Microbiol 58: 1421–1429 [DOI] [PubMed] [Google Scholar]
- Salvail H, Caron M‐P, Bélanger J, Massé E (2013) Antagonistic functions between the RNA chaperone Hfq and an sRNA regulate sensitivity to the antibiotic colicin. EMBO J 32: 2764–2778 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schu DJ, Zhang A, Gottesman S, Storz G (2015) Alternative Hfq‐sRNA interaction modes dictate alternative mRNA recognition. EMBO J 34: 2557–2573 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuck A, Diwa A, Belasco JG (2009) RNase E autoregulates its synthesis in Escherichia coli by binding directly to a stem‐loop in the rne 5′ untranslated region. Mol Microbiol 72: 470–478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma CM, Papenfort K, Pernitzsch SR, Mollenkopf H‐J, Hinton JCD, Vogel J (2011) Pervasive post‐transcriptional control of genes involved in amino acid metabolism by the Hfq‐dependent GcvB small RNA. Mol Microbiol 81: 1144–1165 [DOI] [PubMed] [Google Scholar]
- Sharma E, Sterne‐Weiler T, O'Hanlon D, Blencowe BJ (2016) Global mapping of human RNA‐RNA interactions. Mol Cell 62: 618–626 [DOI] [PubMed] [Google Scholar]
- Soper T, Mandin P, Majdalani N, Gottesman S, Woodson SA (2010) Positive regulation by small RNAs and the role of Hfq. Proc Natl Acad Sci USA 107: 9602–9607 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sudo N, Soma A, Muto A, Iyoda S, Suh M, Kurihara N, Abe H, Tobe T, Ogura Y, Hayashi T, Kurokawa K, Ohnishi M, Sekine Y (2014) A novel small regulatory RNA enhances cell motility in enterohemorrhagic Escherichia coli . J Gen Appl Microbiol 60: 44–50 [DOI] [PubMed] [Google Scholar]
- Sugimoto Y, Vigilante A, Darbo E, Zirra A, Militti C, D'Ambrogio A, Luscombe NM, Ule J (2015) hiCLIP reveals the in vivo atlas of mRNA secondary structures recognized by Staufen 1. Nature 519: 491–494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Travis AJ, Moody J, Helwak A, Tollervey D, Kudla G (2014) Hyb: a bioinformatics pipeline for the analysis of CLASH (crosslinking, ligation and sequencing of hybrids) data. Methods 65: 263–273 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tree JJ, Granneman S, McAteer SP, Tollervey D, Gally DL (2014) Identification of bacteriophage‐encoded anti‐sRNAs in pathogenic Escherichia coli . Mol Cell 55: 199–213 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuck AC, Tollervey D (2011) RNA in pieces. Trends Genet 27: 422–432 [DOI] [PubMed] [Google Scholar]
- Udekwu KI, Darfeuille F, Vogel J, Reimegård J, Holmqvist E, Wagner EGH (2005) Hfq‐dependent regulation of OmpA synthesis is mediated by an antisense RNA. Genes Dev 19: 2355–2366 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Urban JH, Vogel J (2007) Translational control and target recognition by Escherichia coli small RNAs in vivo. Nucleic Acids Res 35: 1018–1037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Y, Juranek S, Li H, Sheng G, Wardle GS, Tuschl T, Patel DJ (2009) Nucleation, propagation and cleavage of target RNAs in Ago silencing complexes. Nature 461: 754–761 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J, Liu T, Zhao B, Lu Q, Wang Z, Cao Y, Li W (2015) sRNATarBase 3.0: an updated database for sRNA‐target interactions in bacteria. Nucleic Acids Res 44: D248–D253 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whelan KF, Sherburne RK, Taylor DE (1997) Characterization of a region of the IncHI2 plasmid R478 which protects Escherichia coli from toxic effects specified by components of the tellurite, phage, and colicin resistance cluster. J Bacteriol 179: 63–71 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worrall JA, Górna M, Crump NT, Phillips LG, Tuck AC, Price AJ, Bavro VN, Luisi BF (2008) Reconstitution and analysis of the multienzyme Escherichia coli RNA degradosome. J Mol Biol 382: 870–883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang A, Wassarman KM, Rosenow C, Tjaden BC, Storz G, Gottesman S (2003) Global analysis of small RNA and mRNA targets of Hfq. Mol Microbiol 50: 1111–1124 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix
Expanded View Figures PDF
Table EV1
Table EV2
Table EV3
Table EV4
Table EV5
Table EV6
Table EV7
Table EV8
Source Data for Figure EV1
Review Process File