

Abstract
The etiologies underlying variation in adult cognitive performance and cognitive aging have enjoyed much attention in the literature, but much of that attention has focused on broad factors, principally general cognitive ability. The current review provides meta-analyses of age trends in heritability of specific cognitive abilities and considers the profile of genetic and environmental factors contributing to cognitive aging to address the ‘missing heritability’ issue. Our findings, based upon evaluating 27 reports in the literature, suggest that verbal ability demonstrated declining heritability after age 60, as did spatial ability and perceptual speed more modestly. Trends for general memory, working memory, and spatial ability generally indicated stability, or small increases in heritability in mid-life. Equivocal results were found for executive function. A second meta-analysis then considered the gap between twin-based versus SNP-based heritability derived from population-based GWAS studies. Specifically, we considered twin correlation ratios to agnostically re-evaluate biometrical models across age and by cognitive domain. Results modestly suggest that nonadditive genetic variance may become increasingly important with age, especially for verbal ability. If so, this would support arguments that lower SNP-based heritability estimates result in part from uncaptured non-additive influences (e.g., dominance, gene-gene interactions), and possibly gene-environment (GE) correlations. Moreover, consistent with longitudinal twin studies of aging, as rearing environment becomes a distal factor, increasing genetic variance may result in part from nonadditive genetic influences or possible GE correlations. Sensitivity to life course dynamics is crucial to understanding etiological contributions to adult cognitive performance and cognitive aging.
Keywords: Lifespan development, cognitive aging, specific cognitive abilities, twins, missing heritability, meta-analysis
Several thorough reviews of behavior genetics of cognitive aging have appeared in the last 5 years (Finkel & Reynolds, 2009; Finkel & Reynolds, 2010; Johnson et al., 2014; Reynolds & Finkel, in press, 2014). Because of the scope of the issues inherent in genetic investigations of cognitive aging, most of these reviews relied on fairly global overviews that allow for identification of major trends and issues. As these reviews exist, we feel at liberty to focus on two very specific issues that serve to showcase the two primary approaches currently in use in investigations of genetic influences on cognitive aging. First, behavior genetic approaches have expanded in the last decade or so to consider a broader array of domains such as executive functioning and working memory. Sufficient data is available to examine age trends not only in general cognitive ability, but also at the level of these specific abilities, providing the necessary data to support and interpret molecular approaches. Second, with the incredible advances in molecular genetics in the last decade, researchers have been combing the genome to identify specific genes associated with cognitive function and cognitive aging. It should be possible, then, to trace the proportion of genetic variance associated with cognitive functioning to a complete set of cognitive genes. However, research on these issues has not produced a straightforward answer. In the current chapter we use meta-analytic approaches to address the worldwide literature on twin- and family-based studies of adult cognition and cognitive aging and consider these results in light of empirical findings of “missing” heritability.
Adult lifespan: decreasing, stable, or increasing heritability?
Meta-analyses of genetic influences on general cognitive ability in childhood and young adulthood have demonstrated that the resulting heritability estimates and patterns of correlations are consistent with the predictions of polygenic theory (T. J. Bouchard & McGue, 1981; Chipuer, Rovine, & Plomin, 1990; Erlenmeyer-Kimling & Jarvik, 1963). In other words, the more genetically related individuals are, the more similar their cognitive ability [see Box 1.1]. However, meta-analyses focusing on age differences in heritability estimates report decreasing twin similarity and increasing heritability from childhood through young adulthood (Bergen et al., 2007; Bouchard, 2013; McCartney et al., 1990). Multiple effects are assumed to account for these age differences: as twins grow up and move out of the house, parents have decreasing control over their environments and twins begin to select their own environments. That selection is not independent of their genetically-influenced personality and cognitive phenotypes (i.e., active gene by environment correlation plays a role).
Box 1.1. Genetic contribution to differential cognitive aging patterns: Heritability.
Twin and family-based methods aim to uncover etiologies of cognitive traits and rely on comparisons of genetic and environmental relatedness to estimate their contributions to individual differences (e.g., see Falconer & Mackay, 1996; Plomin, DeFries, et al., 2013; Sham, 1998; Visscher et al., 2008). A biometrical model of cognitive performance describes etiological factors that contribute to individual differences or variance in cognitive performance (VCOG), including genetic factors that act additively (VA), genetic factors that act non-additively such as dominance (VD), shared rearing or common environmental factors (VC), and nonshared unique environmental experiences plus any measurement error (VE) (c.f. Falconer & Mackay, 1996; Plomin, DeFries, et al., 2013; Sham, 1998):
| [1a] |
Moreover, the relative genetic and environmental contributions can be estimated relative to the total variance in cognitive performance:
| [1b] |
and, restated as:
| [1c] |
Broad-sense heritability (h2B) indexes the proportion of variance in cognitive performance that is due to additive and nonadditive genetic variances while narrow-sense heritability (h2N) indexes the proportion of variance in cognitive performance scores that is simply due to additive genetic variance (Falconer & Mackay, 1996; Plomin, DeFries, et al., 2013; Sham, 1998):
To estimate these components of variances using twins reared together, the covariance (C) or correlation (R) among each twin's cognitive performance score with their cotwin's cognitive performance score are computed for MZ twin pairs (CMZ, RMZ) and for DZ twin pairs (CDZ, RDZ). For MZ twins reared together their cognitive scores covary or correlate because they are genetically identical with respect to segregating genes, and hence for additive and nonadditive genetic influences, in addition to a common, indeed shared, rearing environment (Falconer & Mackay, 1996; Plomin, DeFries, et al., 2013; Sham, 1998):
| [2a] |
| [2b] |
For DZ twins reared together, they covary or correlate with one another for cognitive performance because on average they share half of their segregating genes, ¼ of dominance deviations, plus a common (shared) rearing environment (Falconer & Mackay, 1996; Plomin, DeFries, et al., 2013; Sham, 1998):
| [3a] |
| [3b] |
By comparing the difference in the covariances, or correlations, we may evaluate the contributions of genetic versus environmental influences, under the assumptions of equal environments and random mating. However, we have too few pieces of information in data from twins reared together to estimate both VD, which tends to increase MZ similarity relative to DZ similarity, and VC, which increases both MZ and DZ similarity. Hence researchers must choose which of these to estimate, for example, by comparing the ratio of MZ twins to DZ twin correlations RMZ/ RDZ (Sham, 1998). Under a scenario with simply additive genetic contributions to pair similarity this ratio would be exactly equal to 2.0 since MZ twins are twice as similar than DZ twins with respect to segregating genes, 100% vs 50%. To the extent that dominance contributes to pair similarity the ratio of correlations would exceed 2.0 and VD instead of VC can be estimated. However, if common environmental experiences were the sole contributor to pair similarity, both MZ and DZ, the ratio of correlations would be 1.0; hence for ratios between 1.0 and 2.0 VC can be estimated in place of VD. Sham (1998) notes that ratios below 1.0 or above 4.0 do not conform to traditional biometrical models and in such cases neither ACE nor ADE models should therefore be fitted (cf. equations 2b and 3b above).
Software packages such as OpenMx (Boker, et al., 2011) or MPlus (Muthén & Muthén, 1998-2012; Prescott, 2004) or similar are used to fit biometrical models such as that described in equation 1a to evaluate genetic and environmental contributions to individual differences. The precision of the variance components estimates are typically calculated within software packages; however, approximate se's for heritability, broad and narrow, can be estimated from intraclass correlations and sample sizes depending on the study design (e.g., Sham, 1998).
It should be noted that in longitudinal studies, multivariate extensions of the described biometrical model are applied, and unscaled genetic and environmental variance estimates are emphasized (i.e., Va, VD, VC, Ve) (e.g., McArdle, et al., 1998). Considering unscaled or ‘raw’ variances allows one to evaluate whether genetic and environmental influences are accumulating or declining with age in tandem, or differentially, which may otherwise be obscured if relying on heritability and environmentality estimates. Indeed, unscaled variances may provide clues about potential genetic and environmental (co) actions (e.g., see Reynolds, et al., 2005; Reynolds, et al., 2014; Reynolds et al., 2007).
However, evolutionary pressures and environmental influences continue to change as adulthood progresses, in addition to the variable action of genetic factors. What can we predict, then, about stability or change in heritability of cognitive ability in the second half of the lifespan? The active gene by environmental correlation that develops in the first half of the lifespan could continue into the second, resulting in steadily increasing heritability as MZ twins choose more similar environments than DZ twins. Alternately, as individuals age beyond child-bearing and child-rearing years, the evolutionary imperative to weed out deleterious genetic mutations wanes (Hamilton, 1966; Kirkwood et al., 2011). Therefore, the consequence of mutations may begin to accumulate, resulting in increasing divergence of MZ and DZ twin correlations (as MZs are more likely to share the mutations) and thus increasing heritability with age. In contrast, as twins continue to age and experience their separate lives, they may continue to “grow apart,” as McCartney and colleagues (McCartney, et al., 1990) claimed. The accumulation of unique environmental factors (environmental assaults, individual experiences, disease) including stochastic chance processes (Finch & Kirkwood, 200) may drive down both MZ and DZ twin similarity, resulting in decreased heritability and increased estimates of nonshared environmental variance. Finally, it is possible that all of these influences have achieved a state of equilibrium by midlife, resulting in fairly stable heritability estimates across the second half of the lifespan.
Two recent analyses have attempted to address the issue of age differences and age changes in heritability of general cognitive ability in adulthood. In a fairly comprehensive review of existing cross-sectional and longitudinal twin studies of cognitive aging extant at that time, Finkel and Reynolds (Finkel & Reynolds, 2009) concluded that heritability of general cognitive ability increased to 80% in midlife but then decreased to 60% in late adulthood, with a corresponding increase in nonshared environmental variance. In contrast, Christiansen and McGue (Christiansen & McGue, 2013) recently reported heritability estimates hovering around 60% in cross-sectional twin data including over 2000 pairs and covering most of adulthood (age 46 – 90). Thus, evidence for both stable and decreasing heritability with age has been reported.
It is possible that these contrasting results are a consequence of differing definitions of general cognitive ability. In fact, it may be misguided to focus our efforts to understand the etiology of cognitive aging on general cognitive ability. Beginning with Schaie's classic longitudinal studies of cognitive aging (Schaie, 1996), the psychometric work by Horn and Cattell (Horn & Cattell, 1966, 1967), and the application of modern growth analysis (McArdle et al., 2002; McArdle et al., 2000), it has been clear that cognitive ability is not a unitary construct and that there are different patterns of cognitive aging across cognitive domains. Similarly, twin studies of cognitive aging that focus on individual cognitive domains report different patterns of genetic and environmental variance across domains and ages (Johansson et al., 2004; Lee et al., 2010; Lessov-Schlagger et al., 2007; McArdle et al., 1998; Reynolds et al., 2005). Therefore, in the hope of clarifying the etiology of cognitive aging, we have conducted a meta-analysis of cross-sectional twin and family studies of specific cognitive abilities in adulthood to examine patterns of age differences in heritability estimates across cognitive domains.
Meta-analysis of heritability estimates
A review of the literature generated 27 articles reporting cross-sectional results from 19 different twin and family studies of adulthood and aging that incorporated measures of specific cognitive functioning (see Supplement 1). Although there are several longitudinal twin studies of cognitive aging, only 5 reported information about specific cognitive abilities, a number insufficient to support the meta-analytic techniques used here. Because at most half of the studies reported estimates of environmental variance (shared or unique) and very few estimated dominance, this meta-analysis focused on narrow-sense heritability. Table 1 presents the characteristics of the 178 heritability estimates collected from these studies, clustered in 6 major cognitive domains: verbal (35 estimates), spatial (20), general memory (45), speed (27), executive functioning (24), and working memory (27). Clustering into domains followed the authors' labels or standard interpretations of the individual tests. Digit span tasks were considered measures of working memory. General memory included primarily measures of episodic memory. Three studies reported twin correlations (TC) instead of heritability estimates (H); structural equation modeling was used to calculate heritability estimates from the twin correlations and number of pairs (Boker et al., 2011). Taken together, the 19 studies included 9916 participants (1929 MZ pairs and 1890 DZ pairs) ranging in age from 14 to 98, with a mean age of 55 (median = 65). Fifty-three percent of the participants were women. A full description of all studies and cognitive tasks is available from the authors.
Table 1. Studies included in the meta-analysis.
| Study | Source | Country | Type | N indv | N pair | Agea | Datab | % Female | Cognitive Domain (# tests) |
|---|---|---|---|---|---|---|---|---|---|
| Central Valley of Costa Rica | Greenwood et al., 2011 | Costa Rica | Family | 253 | NAc | 73.3 (12.7) | H | 65 | Executive function (3) General memory (4) Verbal (2) |
| Diabetes Heart Study | Cox et al., 2014 | U.S. | Family | 550 | NA | 67.3 (8.8) | H | 55 | Executive function (3) General memory (1) Speed (1) |
| Genetics of Brain Structure and Function | Glahn et al., 2013 | U.S. | Family | 1129 | NA | 44.2 (14.0) | H | 62 | Executive function (3) General memory (6) Speed (3) Verbal (1) Working memory (7) |
| German Observational Study of Adult Twins | Neubauer et al., 2000 | Germany | Twin | 600 | 169 MZ 131 DZ |
34.3 (13.0) | H | 78 | Executive function (4) Spatial (2) Speed (1) Verbal (3) Working memory (2) |
| Italian Twin Registry | Giubelei et al., 2008 | Italy | Twin | 186 | 35 MZ 58 DZ |
67.6 (4.7) | H | 61 | Executive function (3) General memory (1) Spatial (1) |
| Keio Twin Study | Ando et al., 2001 | Japan | Twin | 472 | 142 MZ 94 DZ |
19.9 (3.5) | TC | 67 | Executive function (2) General memory (2) Spatial (1) Verbal (1) |
| Kinki University Adult Twin Registry | Hayakawa et al., 1992 | Japan | Female twins | 66 | 30 MZ 3 DZ |
50-78 | TC | 100 | Spatial (1) Speed (1) Working memory (1) |
| Male twins | 128 | 46 MZ 18 DZ |
50-78 | TC | 0 | Spatial (1) Speed (1) Working memory (1) |
|||
| Longitudinal Study of Aging Danish Twins | McGue et al., 2001 | Denmark | Twin | 806 | 168 MZ 235 DZ |
80.0 (4.0) | H | 70 | General memory (1) Verbal (1) Working memory (1) |
| Minnesota Study of Twins Reared Apart | Bouchard et al., 1990 | U.S. | Twin | 112 | 56 MZA | 41.0 (12.0) | TC | NRd | Verbal (1) |
| Minnesota Twin Study of Adult Development and Aging | Finkel & McGue, 1993 | U.S. | Twin | 320 | 93 MZ 67 DZ |
67.0 (6.7) | H | 50 | General memory (4) Speed (1) |
| Finkel et al., 1995 | Younger Twin | 140 | 41 MZ 21 DZ |
38.9 (4.4) | H | 75 | Spatial (1) Speed (1) Verbal (1) Working memory (1) |
||
| Middle Aged Twin | 176 | 50 MZ 30 DZ |
60.1 (3.9) | H | 56 | Spatial (1) Speed (1) Verbal (1) Working memory (1) |
|||
| Older Twin | 66 | 41 MZ 40 DZ |
69.3 (5.8) | H | 54 | Spatial (1) Speed (1) Verbal (1) Working memory (1) |
|||
| Finkel & McGue, 1998 | Younger Twin | 294 | 86 MZ 61 DZ |
43.3 (8.3) | H | 68 | General memory (3) Verbal (1) |
||
| Older Twin | 388 | 115 MZ 79 DZ |
69.8 (10.5) | H | 54 | General memory (3) Verbal (1) |
|||
| Finkel & McGue, 2007 | Twin | 738 | 185 MZ 131 DZ |
58.7 (NR) | H | 60 | Speed (2) | ||
| National Heart Lung Blood Institute | Swan et al., 1990 | U.S. | Twin | 524 | 134 MZ 133 DZ |
63 (NR) | H | 0 | Speed (1) |
| Swan et al., 1999 | Twin | 366 | 94 MZ 89 DZ |
71.8 (2.9) | H | 0 | General memory (1) Verbal (3) |
||
| Netherlands Twin Registry | Posthuma et al., 2001 | The Netherlands | Twin-family | 688 | 102 MZ 131 DZ |
37 (NR) | H | 56 | Speed (1) Verbal (1) |
| Posthuma et al., 2002 | Twin-family | 135 | 24 MZ 31 DZ |
30.0 (9.0) | H | 45 | Working memory (1) | ||
| Norwegian Twin Panel | Tambs et al., 1994 | Norway | Twin | 160 | 40 MZ 40 DZ |
41.0 (8.1) | H | 53 | Spatial (4) Speed (1) Verbal (5) Working memory (2) |
| Older Australian Twin Study | Lee et al., 2012 | Australia | Twin | 472 | 117 MZ 98 DZ |
71.0 (5.2) | H | 66 | Executive function (3) Working memory (1) |
| OCTO-Twin | McClearn et al., 1997 | Sweden | Twin | 388 | 90 MZ 104 MZ |
82.3 (NR) | H | 64 | General memory (1) Spatial (1) Speed (1) Verbal (1) |
| Johansson et al., 1999 | Twin | 564 | 125 MZ 157 DZ |
83.3 (3.1) | H | 64 | General memory (4) Working memory (2) |
||
| Russian Twin Study | Malykh et al., 2005 | Russia | Twin | 160 | 40 MZ 40 DZ |
21.7 (3.3) | H | 50 | Verbal (1) |
| Swedish Adoption Twin Study of Aging | Pedersen et al., 1992 | Sweden | Twin | 645 | 113 MZ 189 DZ |
65.6 (8.4) | H | 60 | General memory (3) Spatial (3) Speed (3) Verbal (3) Working memory (2) |
| Swedish Adoption Twin Study of Aging | Finkel et al., 1995 | Younger Twin | 102 | 21 MZ 30 DZ |
41.1 (5.9) | H | 47 | Spatial (1) Speed (1) Verbal (1) Working memory (1) |
|
| Middle-Aged Twin | 146 | 35 MZ 29 DZ |
58.8 (4.3) | H | 49 | Spatial (1) Speed (1) Verbal (1) Working memory (1) |
|||
| Older Twin | 164 | 30 MZ 51 DZ |
71.6 (4.8) | H | 66 | Spatial (1) Speed (1) Verbal (1) Working memory (1) |
|||
| Finkel & Pedersen, 2014 | Twin | 241 | 64 MZ 96 DZ |
66.5 (7.8) | H | 61 | Speed (2) | ||
| TwinsUK | Singer et al., 2006 | U.K. | Twin | 556 | 108 MZ 170 DZ |
56.0 (11.0) | H | 100 | General memory (1) Speed (2) Verbal (1) Working memory (1) |
| Vietnam Era Twin Study of Aging | Kremen et al., 2005 | U.S. | Twin | 690 | 179 MZ 169 DZ |
47.9 (3.3) | H | 0 | Verbal (1) |
| Kremen et al., 2014 | Twin | 1237 | 349 MZ 265 (DZ) |
55.4 (2.5) | H | 0 | General memory (6) |
Mean age (standard deviation) if reported, otherwise age range
Type of data reported: H = heritability estimate, TC = twin correlations
NA indicates “not applicable”
NR indicates “not reported”
Identifying a common metric of dispersion/accuracy represented the greatest challenge to our meta-analysis. A few studies reported confidence intervals (16.9%) and another 15.7% reported standard errors for their estimates. Forty-five percent reported twin correlations, which could be used to generate heritability estimates and corresponding standard errors via calculation (McCartney, et al., 1990; Sham, 1998) or structural equation modeling (Boker, et al., 2011). However, 22.5% reported neither standard errors nor sufficient information to estimate standard errors. In the interest of consistency, we chose to calculate an approximate standard error for each heritability estimate following Koots and Gibson (Koots & Gibson, 1996), who used 2/√N for parent-offspring pairs, where N is the total number of twin pairs in the current case. The average difference between this metric and the reported or calculated standard errors (where available) was .007; the smallest difference by an order of magnitude of any of the standard error metrics we investigated.
Two approaches were used to summarize the data points within each domain. First, the common standard error estimate (2/√N) was used to calculate 95% confidence intervals around each heritability estimate. Estimates were then plotted (with confidence intervals) by the mean age of the sample (left side of Figures 1-6). Where mean age was not reported, the midpoint of the reported age range was used. A reference line is included in the figures to indicate the average heritability across ages. Second, a multi-level random effects regression model was fitted to the data points (SAS Proc Mixed; SAS, Cary, NC), including both linear and quadratic age components (right side of Figures 1-6). Covariates included gender, publication year, and region (U.S., Europe, non-European). Estimates were adjusted for sampling variability by including the estimated standard errors as a level 1 predictor with unit-constrained variance (Hox, 2010), and between-study random effects estimated. Given the small sample of heritability estimates within each domain (ranging from 20 to 45), there was insufficient power to differentiate between linear and quadratic models in 5 of the 6 cognitive domains. Model fit statistics and parameter estimates with standard errors are reported in Supplemental Tables 1 and 2.
Figure 1.
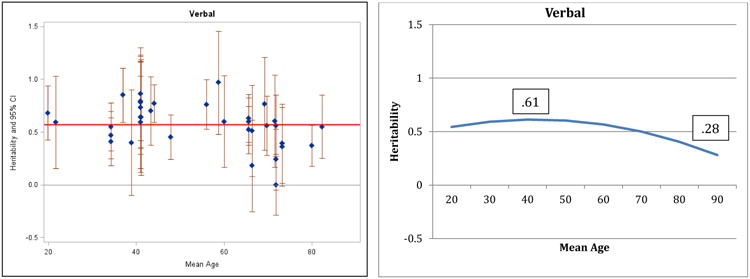
Heritability of Verbal Ability: Estimates and expected curve across age.
Results for verbal ability are presented in Figure 1: both the individual point estimates presented on the left and the fitted curve presented on the right suggest decreasing heritability in the second half of the lifespan and the quadratic regression fit significantly better than the linear model (change in model fit = 6.4, df = 1, p<.05). A reference line in the left panel indicates average heritability across the entire age range: before age 60, 75% of the study heritabilities are greater than the average, whereas after age 60, 73% of the heritabilities are at or below the average. Maximum (.61) and minimum (.28) heritabilities estimated by the regression model are reported in the right panel. The pattern of decreasing heritability indicated here for verbal ability is similar to the pattern reported by Finkel and Reynolds (2009) for general cognitive ability; the same pattern is suggested by the results for spatial ability (Figure 2) and perceptual speed (Figure 4), although the comparisons of regression models did not achieve significance. Note that most measures of general cognitive function rely on four primary domains (verbal, spatial, memory, and speed) and in the current analyses 3 of those 4 domains manifested the same pattern reported for general cognitive ability by Finkel and Reynolds (2009). In contrast, the pattern of results for general memory (Figure 3) and working memory (Figure 5) suggest stability or possible increases in heritability, although the quadratic model did not achieve significance.
Figure 2.
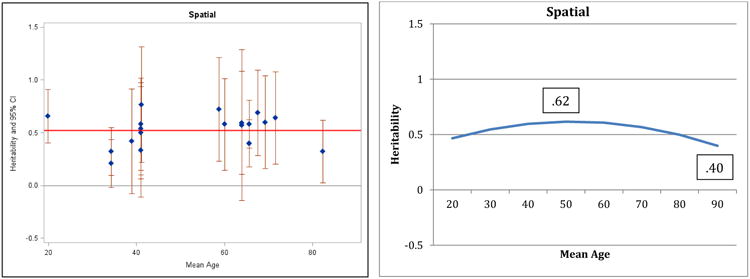
Heritability of Spatial Ability: Estimates and expected curve across age.
Figure 4.
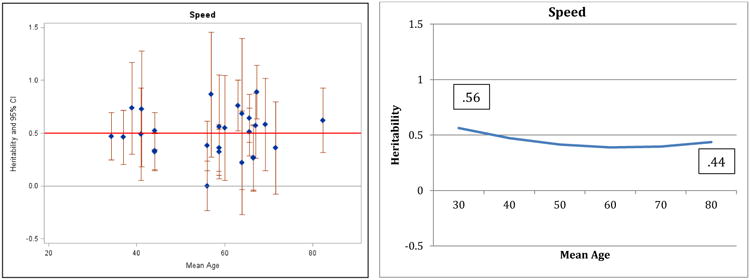
Heritability of Perceptual Speed: Estimates and expected curves across age.
Figure 3.
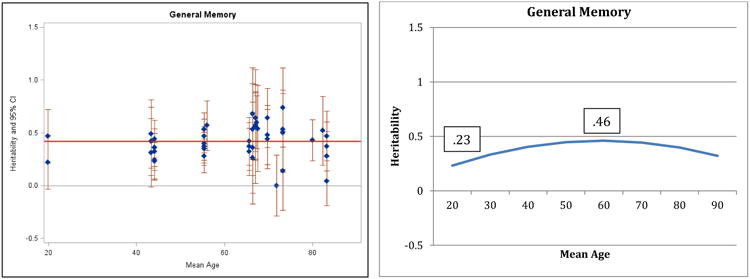
Heritability of General Memory: Estimates and expected curve across age.
Figure 5.
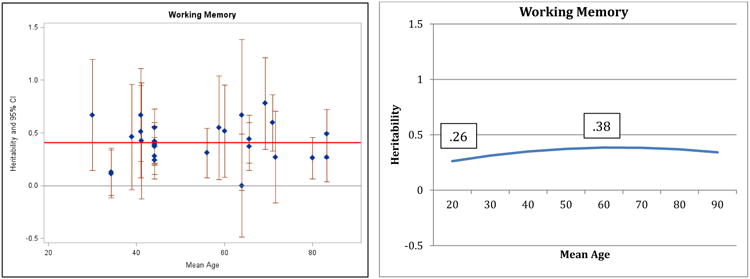
Heritability of Working Memory: Estimates and expected curve across age.
Executive function (Figure 6) presents the sparsest data and the greatest puzzle in these analyses. The individual point estimates and the quadratic regression model for executive functioning suggest an increase in heritability in late adulthood. However, closer examination of individual heritability estimates suggested that the age trends for executive functioning might be more complicated, with multiple inflection points across adulthood. A model including three linear slopes (age 20-50, age 50-65, age 65-90) provided a marginally better fit to the data than the linear model for executive functioning (change in model fit = 4.8, df=2, p<.10); whereas the quadratic model did not improve fit over the linear model (change in model fit = 1.0, df=1, ns). Both the quadratic and three-slope models are presented in right half of Figure 6; the different heritability estimates at age 90 arising from the two models are indicated. As a result of the paucity of data, it is impossible to determine whether heritability for executive function increases to .77 or decreases to .27 in late adulthood. Even examination of the few longitudinal data points available for executive function cannot provide clarification: both modest increases (.52 to .61) and modest decreases (.52 to .43) in heritability estimates for measures of executive function are reported from wave 2 (mean age 72.7) to wave 3 (mean age 76.6) of the National Heart Lung Blood Institute twin study (Lessov-Schlagger et al, 2007).
Figure 6.
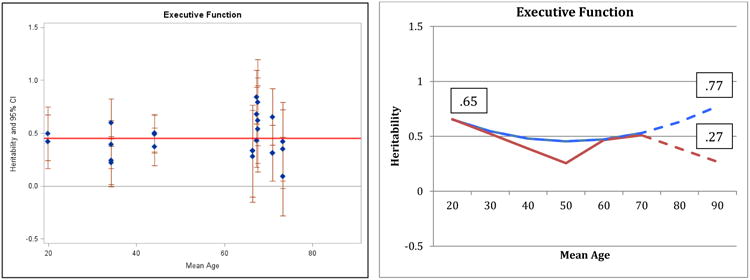
Heritability of Executive Function: Estimates and expected curves across age.
Thus, the meta-analyses revealed some evidence for all three possible patterns of differences in heritability with age (increase, stability, decrease), and the patterns varied distinctly by cognitive domain. Interestingly, three domains of cognitive function that have been associated with structural changes in specific brain regions (Raz, 2000) – general memory, working memory, and executive functioning – show patterns of modestly increasing heritability with age, at some point in the lifespan. It is possible that, as Hamilton (1966) suggested, increasing heritability results from the accumulation with age of deleterious genetic mutations that impact physiological function within the brain. The APOE risk haplotype e4 may have maximal impacts on dementia risk particularly before age 70 (Blacker et al., 1997) but has shown broad impacts to multiple domains of cognitive aging (Davies et al., 2014; Finkel et al., 2011; Reynolds et al., 2006). Other genes evaluated likewise do not appear to be associated particularly with narrow cognitive domains, such as SORL1 (e.g., episodic memory, spatial ability (Reynolds et al., 2013), or BDNF (e.g., hippocampal-dependent memory processes, executive functioning, global cognitive functioning; see (Honea et al., 2013; Kambeitz et al., 2012; Mandelman & Grigorenko, 2012)). Verbal ability, spatial ability, perceptual speed, and possibly executive functioning show patterns of declining heritability, generally after age 60. With respect to these components of cognition, then, it appears that twins continue to “grow apart” in late adulthood as McCartney et al. (1990) suggested: individual life choices and experiences result in decreasing twin similarity and decreasing heritability.
Overstating the case for heritable influences? Missing heritability
Measured gene approaches such as GWAS, which assay millions of single nucleotide polymorphisms scattered across the genome, have brought about alternative methods to twin and family designs. For example, the Genome-wide Complex Trait Analysis (GCTA) estimates the aggregate contribution of SNP variants across the genome to individual differences in a phenotype, summarized as a narrow-sense SNP-based heritability (Davies et al., 2011; Deary et al., 2012). While the GCTA approach is conceptually similar to family-based biometrical approaches, the GCTA estimates the genetic relatedness (i.e., genomic relationship matrix; GRM) among otherwise unrelated individuals. In other words, even genetically unrelated individuals can coincidentally share some gene variants, though the average genetic relatedness in an unrelated sample is zero (Yang et al., 2011).
GCTA estimates of SNP-based heritability for fluid and crystalized abilities have been estimated at .51 and .40, respectively, based upon GWAS data collected on 3511 unrelated persons from the CAGES consortium ranging in age from 44 to 93 years (Davies, et al., 2011). Moreover, 24% of the cognitive change between childhood (11 years) and older adult cognitive performance at ages 65 and older (residual based) is accounted for by common SNPs as measured using GWA SNP data on a subset of 1940 persons from the CAGES consortium (Deary, et al., 2012). Reanalysis of three of the CAGES consortium samples (N=1804) applied GCTA using a genome-scan approach to estimate the contribution of about 500k autosomal SNPs, reporting “population-sense heritabilities (h2ps)” of .36 for crystalized ability, .19 for fluid abilities, and .26 for cognitive change (Rowe et al., 2013). The range of SNP-based heritability estimates across the two studies for fluid ability is attributed to broader age composition of the larger CAGES sample set (Davies, et al., 2011)versus the older ages represented in the reduced sample set for which the genome scan was applied (Rowe, et al., 2013). Nonetheless, these SNP-based heritability estimates for fluid and crystalized ability measures are lower than our meta-analytic heritability estimates, i.e., 56-62% at the peak for verbal, spatial and speed traits (see Figures 1, 2, and 4). This difference is broadly consistent with GCTA estimates of heritability versus twin-based estimates in childhood within the same sample: SNP-based heritability (about .20-.30) versus traditional twin samples (.40 - .60) were substantial though lower (Plomin, Haworth, et al., 2013; Trzaskowski et al., 2013).
Moreover, recent investigations of Alzheimer Disease (AD), for which there has been heavy emphasis on consistency of phenotyping, and for which GWAS have paid off relatively more handsomely than in other domains, have reports of ‘missing heritability’. SNP-based heritability estimation suggests genetic influences explain approximately 33% of the total liability or risk for AD: APOE itself accounts for 6%, while nine extant identified and confirmed genes account for 2% (ABCA7, BIN1, CD33, CD2AP, CLU, CR1, MS4A6A, MS4A4E, PICALM) (Ridge et al., 2013); additional candidates have been identified based on a meta-analysis of GWAS studies with over 74000 individuals (Lambert et al., 2013). Yet, the median twin-based heritability for AD is higher at about 57% (see Gatz et al., 2014; Reynolds & Phillips, under review, 2014). Likewise, a study of the HRS sample suggested similar findings when evaluating cognitive functioning constructed from telephone screening performance based on the MMSE plus an episodic verbal recall memory task (C. Zhang & Pierce, 2014). The SNP-based heritability ranged from 15-33% on overall performance while for longitudinal linear decline, SNP heritability was far lower at about 5%. One can debate the linearity vs. nonlinearity of the trajectory shape as to whether the heritability of the slope was underestimated (c.f. Reynolds, et al., 2005), but these findings coupled with the findings for AD collectively suggest that the SNP heritability fails to approach heritability values from twin studies. Thus, if up to 33% of AD risk or cognitive functioning is explained by common variants, of which only a fraction is due to identified candidates, more work remains to identify additional candidates and to ‘close’ the gap between the twin-based and SNP-based heritability estimates.
There are a number of explanations proposed for this gap between SNP heritability estimates and twin- or family-based heritability estimates. These include the idea that GWAS does not, by design, capture: other structural variants beyond SNPs, rare variants, poorly tagged or multiple independent variants, dominance and epistasis (GxG interaction), epigenetics, GE interplay in the form of gene-environment correlation (e.g., active niche-picking), and GxE1 interaction (e.g., see Bloom et al., 2013; Eichler et al., 2010; Gusev et al., 2013; Hemani et al., 2013; Kaprio, 2012; Koch, 2014; Liu & Leal, 2012; Maher, 2008; Plomin, 2013; G. Zhang et al., 2012; Zuk et al., 2012). Hence, while these factors likely play a role in contributing to human traits such as cognitive abilities they may not be accounted for when applying GCTA and similar methods. We emphasize that layered on top of these discussions must be the recognition that any of these explanations may show age-related dependencies.
Among models proposed to evaluate or uncover missing heritability, we consider the limiting pathways model and the local (vs genome-wide) heritability approach.
Limiting pathways model
The limiting pathway model (LP; Zuk, et al., 2012) suggests that heritability based on twin studies may contribute to ‘phantom heritability’ in cases where gene pathways interact, i.e., where there is epistasis. Indeed, the greater the number of gene pathways that hypothetically contribute, the greater the occurrence of phantom heritability (Zuk, et al., 2012). According to their models and simulations, phantom heritability in principle can occur even when patterns of twin correlations suggest C and may even be higher where both C and D influence a trait (Zuk, et al., 2012). Alternate designs, such as GCTA, offer lower bound narrow heritability estimates. Hence, the authors of the LP model propose that studies focus on individuals from isolate populations which may provide a more reasonable solution using IBD (identity-by-decent) sharing (Zuk, et al., 2012).However, reliance on isolate populations presents another set of issues in terms of trait phenomena and generalizability, among others. Moreover, while emerging work supports the hypothesis that epistasis may be more common than previously realized (Bloom, et al., 2013), findings contrary to that of the LP model are appearing (e.g., Maki-Tanila & Hill, 2014). Indeed, the quantification of epistasis represents a challenge in that even if it is substantial, epistatic effects may contribute largely to additive genetic variance (Maki-Tanila & Hill, 2014).
‘Local’ and genome-wide heritability
Models that aggregate effects across all possible contributing variants at particular loci, i.e., “local” heritability or loci-based heritability, as well as in aggregate across regions or the entire genome collectively, may provide avenues for discerning particular gene or regional contributions to an outcome (Gusev, et al., 2013; Rowe, et al., 2013) and inform the missing heritability debate. Via simulation and application to nine disease traits, Gusev and colleagues (Gusev, et al., 2013) considered the contribution of the most significant GWAS SNP at a locus (h2GWAS), the contribution of all significant SNPs at a locus based on conditional linear modeling (h2GWAS,Joint), the contribution of all SNPs at a locus constructed using the standard variance component approach (h2g; i.e., using GCTA), and the contribution of SNPs at a locus using a variance component adjusted for linkage disequilibrium (LD) (h2g,LD). Based on simulated genome-wide data modeled on the Wellcome Trust Case Control Consortium WTCCC1:CAD cohort, findings supported the conclusion that h2g was approximately unbiased if the causal variants were randomly sampled (i.e., not weighted towards more common variants) but clearly biased in both directions if the causal variants were not randomly sampled and included more common variants (e.g., returning estimates that were 62% to110% of the true heritability value).In comparison, while h2g,LD was routinely downward biased (returning estimates at 94-95% of the true value),it was less perturbed at the sampling of the causal variants. Furthermore, this LD-residualized variance components approach (h2g,LD) was also advocated in genome-wide and local-heritability contexts with the h2g,LD explaining about 29-30% more heritable variance. Indeed, comparisons of the aforementioned heritability estimators at the local level at known GWAS loci for the selected traits suggested the presence of additional common SNPs while at the genome-wide level the additional low frequency (rare) SNPs contribute to the complex disease traits evaluated. With respect to the missing heritability question, this work suggests that even when considering an additive model, additional heritability may be uncovered when adjusting the standard variance component model (GCTA) for LD. Relevant work focused on rare variants suggests, however, that while a portion of rare variant contributions to a trait may be captured in aggregate analyses, underestimation may be routine due to bi-directional effects of causal variants and presence of noncausal variants (Liu & Leal, 2012). Hence, aggregate methods may effectively lose some signal from rare variants, which may otherwise be highly penetrant and individually impactful.
Overall, as GWAS chips have become denser, sample sizes have become larger, and considerations of genetic architecture have advanced, the missing heritability gap has narrowed across a number of traits (Gusev, et al., 2013; van Dongen & Boomsma, 2013). Moreover, with respect to cognition, when considering the same measures of cognitive ability at the same ages (in childhood), methods such as GCTA have explained about two-thirds of the heritability estimated from twin models within the same sample (Plomin, Haworth, et al., 2013). However, across the approaches and models undertaken to evaluate this gap few to none have considered that genetic contributions, and hence heritability, may differ across the lifespan. Recognition of this may provide an insight into possible mechanisms that contribute to the gap in heritability estimates particularly for cognitive ability and aging.
Meta-analysis of MZ/DZ correlation ratios
If the missing heritability gap is due to unaccounted for nonadditivity (dominance and epistasis), and in the absence of shared environment (C=0), this should be evident in ratios of MZ/DZ correlations that exceed 2.0 (see equations 1-3 in Box 1.1). However, we note that MZ-DZ correlation ratios that exceed 4.0, which does not conform to a traditional ADE model, could result from intensive gene-environment interplay, e.g., active gene-environment correlational (rGE) processes such as niche-picking whereby environments are selected that suit genetically influenced traits (Plomin, DeFries, et al., 2013; Scarr & McCartney, 1983). As described above, GE correlation is theorized to explain the increasing heritability for general cognitive ability with age from childhood into adulthood that peaks in young-old age before apparently declining (Bouchard, 2013; Finkel & Reynolds, 2009; Plomin, DeFries, et al., 2013). Hence, for example, individuals may choose (or not) higher educational, occupational and leisure time environments, and spouses, conducive to their partly heritable abilities that may further bolster or moderate their abilities. Such an occurrence would therefore drive increasing MZ similarity over age relative to DZ similarity (c.f. McCartney, et al., 1990). Thus, GE processes could in principle result in increasing MZ/DZ correlation ratios such that they exceed 4.0. A further evaluation of twin correlation ratios, over age, may point to processes that contribute to missing heritability gap. That is, the missing heritability gap may narrow or widen with age.
We conducted a meta-analysis of log transformed MZ/DZ ratios of intraclass correlations based on Sham (1998) for each of the 6 cognitive domains represented in the collected twin data similar to that described above for the meta-analysis of heritability estimates. A total of 103 twin correlations were reported across domains. We dropped two negative ratio values from analysis resulting from small negative correlations among DZ pairs (-.04 to -.11) given that these ratios did not conform to expected values (Sham, 1998). With an analysis sample of 101 correlations across domains and up to 21 ratio values within a domain, we limited covariates to age and age-squared, with age centered at the median age of 65 years. For executive functioning with only 8 data points, we simply adjusted for age. Analyses accounted for sampling variance using the inverse number of twin pairs reported given that the standard error of a ratio of independent correlations is not yet established. Random effects at the study level were nonsignificant (median=4.18%, range 0.00 to 16.75%, all p>.30), but were retained in the model. Sensitivity analyses suggested that study dependency impacted confidence intervals compared to point estimates (not shown). Results suggest that at age 65 (the centering value), ratios tilt towards ADE models especially for verbal (see Table 2, Intercept effect). For general memory, an ACE model was supported based on the fixed effect estimate, although the confidence interval does not rule out ADE. With respect to perceptual speed, the answer is essentially equivocal as to ACE or ADE with a large confidence interval although the point estimate lies just over the ADE boundary. For the remaining traits, particularly working memory, as well as executive functioning, and spatial, the estimates favor an ADE model, although the confidence intervals do not exclude an ACE model. The age covariates were not individually significant (see Table 2).However, together the age covariates accounted for noticeable variance across domains (median=14.70%, range = 1.64% to 19.92%), with spatial ability reporting the smallest effect and executive functioning the largest. Model fit statistics and information criteria are reported in Supplemental Table 3.
Table 2.
Multilevel fixed effect estimates of MZ/DZ ratios by cognitive domain.
| Domain | N Obs | Fixed Effect | Estimate (ln) | se | LCL | UCL | Expected rMZ/rDZ | “Model” |
|---|---|---|---|---|---|---|---|---|
| Verbal | 21 | Intercept | 1.379 | 0.112 | 1.114 | 1.644 | 2.97 | ADE* |
| Age | 0.000 | 0.009 | -0.020 | 0.021 | -- | |||
| Age2 | 0.000 | 0.000 | -0.001 | 0.000 | -- | |||
| Spatial | 14 | Intercept | 1.148 | 0.102 | 0.885 | 1.410 | 2.15 | ADE |
| Age | -0.005 | 0.009 | -0.030 | 0.018 | -- | -- | ||
| Age2 | 0.000 | 0.000 | -0.001 | 0.001 | -- | -- | ||
| Speed | 20 | Intercept | 1.116 | 0.092 | 0.892 | 1.340 | 2.05 | A?E |
| Age | 0.007 | 0.011 | -0.017 | 0.031 | -- | -- | ||
| Age2 | 0.001 | 0.000 | 0.000 | 0.002 | -- | -- | ||
| General Memory | 23 | Intercept | 1.017 | 0.185 | 0.565 | 1.469 | 1.76 | ACE |
| Age | 0.004 | 0.011 | -0.019 | 0.028 | -- | -- | ||
| Age2 | 0.000 | 0.000 | 0.000 | 0.001 | -- | -- | ||
| Working Memory | 15 | Intercept | 1.453 | 0.190 | 1.004 | 1.902 | 3.28 | ADE |
| Age | -0.012 | 0.012 | -0.044 | 0.020 | -- | -- | ||
| Age2 | -0.001 | 0.001 | -0.003 | 0.001 | -- | -- | ||
| Executive Functioning | 8 | Intercept | 1.283 | 0.125 | -0.307 | 2.872 | 2.61 | ADE |
| Age | 0.006 | 0.004 | -0.005 | 0.017 | -- | -- |
Note. Each ratio was transformed accordingly: ln (rMZ/rDZ +1). N Obs = number of data points contributing to each analysis. Analyses were weighted based on pair sample size. The fixed effect estimates and standard errors are presented from multi-level modeling in SAS Proc Mixed (Cary, NC), where LCL=95% Lower Confidence Limit and UCL = 95% Upper confidence limit. The intercept reflects the expected ratio in natural log units, adjusted for age- and age-squared terms centered at 65 years, with the exception of Executive Functioning that entered only the linear age term. If the intercept estimates exceed 1.099 that would indicate an ADE model may be warranted when back-calculated into raw ratio units (i.e., exceed 2.0). Back-calculated expected rMZ/rDZ ratios are provided. ADE* = support of ADE model based on 95% CI of the intercept that excludes values 1.099 and below.
Figure 7 presents the individual ratios, log transformed, with the corresponding fixed effect estimate and its confidence interval from the age-adjusted model described above. The forest plots suggest that among the evident heterogeneity of individual ratios, the ratios tend to increase in magnitude with age, up to about age 65, perhaps most noticeably for verbal. The exception is perceptual speed, wherein the ratios tend to decrease with age and cluster towards an ACE model. We note that some of the individual ratio estimates show values exceeding the expected upper boundary of 4.0, inconsistent with an ADE model. This is predominantly the case in studies with average ages between 60 and 75. This outcome is particularly interesting in light of accelerating declines that may be observed at approximately 65 years (see Finkel et al., 2003; Reynolds, et al., 2005). Moreover, after age 75, there are no studies reporting ratio values that exceed 4.0.Hence, peak GE correlational processes may occur around this age before increasing frailty sets and when older adults are still generally able enough to take advantage of their increased leisure time after retirement to select environments that impact verbal ability; however, later in adulthood, as frailty increases and options become more limited, GxE interaction may become more salient (Reynolds et al., 2014). Moreover, increasing evidence for ADE comes with increasing distance from rearing environments; hence rearing environment becomes a more distal impact while proximal environmental influences (even from mid-adulthood) remain strong.
Figure 7.
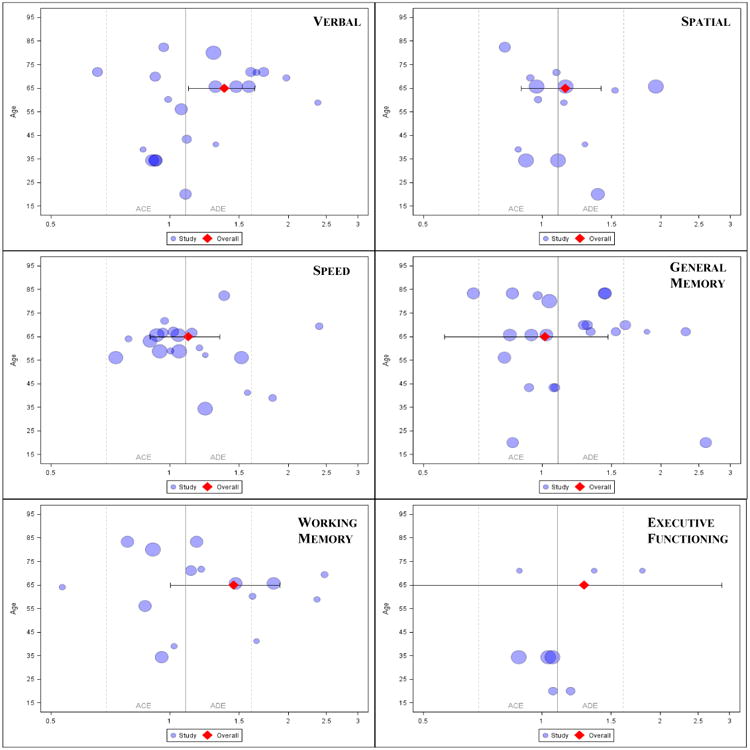
MZ/DZ correlation ratios by study (log transformed) with overall estimate and 95% CI.
Note. Each ratio was transformed accordingly: ln(rMZ/rDZ +1). The blue circles represent study ratio values scaled relative to pair sample size. The red diamond represents the overall fixed effect estimate with a 95% CI. The solid gray vertical line at approximately 1.10 natural log units is equivalent to a raw ratio of 2.0. Dashed gray vertical lines indicate boundaries in natural log units equivalent to raw ratios of 1.0 and 4.0, respectively.
We would be remiss, however, in not pointing out that the limited number of observations for analysis, and hence power, also hampered consideration of other potential moderators (e.g., cohort, country, etc.) beyond these initial and modest examinations of age differences. Yet, it is interesting to consider that even in light of the circumstance such that epistatic effects may contribute mainly to additive genetic variance (Maki-Tanila & Hill, 2014), potential shifts in the relative nonadditive contributions, or GE interplay, may become evident across time. Of course, large longitudinal twin designs would more appropriately and powerfully address these potential circumstances. AE is the typical biometrical model presumed based on twin correlation patterns across a number of disparate domains (i.e., M [rmz-2rdz] ∼ 0.003, SD=.20; (Hill et al., 2008). However, the possibly shifting patterns we observed in the potential contributions of C or D (and not just simplified to AE) suggest that one should consider development a life-long process, i.e., one biometrical model may not explain the whole of the adult lifespan within or across cognitive domains let alone other psychological and biomedical traits.
Summary and conclusions
While the etiology of cognitive abilities and cognitive aging has enjoyed much attention in the literature, much of that attention has focused on broad factors, principally general cognitive ability. In the current review, we considered age trends in heritability of specific cognitive abilities, as well as the profile of genetic and environmental factors contributing to cognitive aging to address ‘missing heritability’. Our findings based upon evaluating 27 reports in the literature suggest that age-sensitive domains – general memory, working memory, and executive functioning – show some modest evidence for increasing heritability with age, at least up to age 60, whereas verbal ability, spatial ability, perceptual speed, and possibly executive functioning show some evidence for declining heritability, typically after age 60. A second meta-analysis considered the gap between twin-based versus SNP-based heritability derived from population-based GWAS studies. Specifically, via MZ and DZ twin correlation ratios we agnostically re-evaluated biometrical models (ACE, ADE, AE) across age and by cognitive domain. Results modestly suggest that nonadditive genetic variance may become increasingly important with age, especially for verbal ability. If so, the lower SNP-based heritability estimates may result in part from uncaptured nonadditivity. Moreover, consistent with longitudinal twin studies of aging, as rearing environment becomes a more distal impact, proximal person-specific environmental influences strengthen, while increasing genetic variance may be partly a consequence of nonadditive genetic influences or possibly GE interplay (Finkel & Reynolds, 2009; Reynolds, et al., 2005; Reynolds, et al., 2014).Both meta-analyses focused on age, however, other moderators may well be important. Cohort factors may be instrumental in lessening or increasing individual differences in abilities, and hence heritability estimates may differ as a consequence (c.f., Baker et al., 1996; Heath et al., 1985). Access to higher educational and occupational opportunities as well as leisure time pursuits are obvious examples, among others.
Arguments that findings of twin and family studies have been superseded by newer molecular techniques lose sight of the value of twin studies to understanding of etiology beyond heritability (van Dongen et al., 2012). Moreover, we suggest from our investigations herein that biometrical analysis of twins over the life course may point to dynamic shifts from familial/shared environments (C) contributions to non-additive genetic effects (D) across development, and indeed GE interplay contributions. It is only by trying to unpack missing heritability differences across complementary designs that we may be able to more fully evaluate etiologies of stability and change in cognitive functioning across the full lifespan. Ideally this would be done using identical phenotyping (Plomin, Haworth, et al., 2013) and with close attention to developmental periods. Indeed, the value of rich phenotyping and opportunities to model GE interplay within a twin context increase understanding of genetic and environmental (co)actions (van Dongen, et al., 2012).
That said, we recognize that relying on scaled heritability estimates rather than genetic variance–as resorted to by necessity in the meta-analyses of available literature presented—may obscured iminutions or amplifications of genetic and environmental variance that provide clues to genetic and environmental influences (e.g., Reynolds, et al., 2014). However, this requires longitudinal studies that would employ continuity and invariance of measurement (or the possibility to model latent scale metrics). Although conducting the “cradle-to-grave” longitudinal studies that would help elucidate the continuity and emergence of genetic and environmental influences across the life course is likely unattainable, collaborations among existing studies can provide an approximation. One example, the consortium on Interplay of Genes and Environment across Multiple Studies (IGEMS; Pedersen et al., 2013), has created a sample of over 17,000 twins aged 25 to 102. Consortiums of this nature will be necessary to support a true-life course application of both behavior genetic and molecular genetic methods.
Supplementary Material
Supplemental table 1. Fit statistics (-2LL) of meta-analysis of heritability.
Supplemental table 2. Parameter estimates (standard errors) of meta-analysis of heritability.
Supplemental table 3. Fit statistics and criteria: meta analysis of MZ/DZ correlation ratios.
Acknowledgments
The authors gratefully acknowledge that this work was partially supported by the National Institute on Aging (R01 AG037985).
Footnotes
We note that while others refer to GxE as contributing to the missing heritability presumably this refers to GxC where C refers to the common environment, as Gx (nonshared) E leads to dissimilarity amongst relatives.
References
- Baker LA, Treloar SA, Reynolds CA, Heath AC, Martin NG. Genetics of Educational Attainment in Australian Twins: Sex Differences and Secular Changes. Behav Genet. 1996;26(2):89–102. doi: 10.1007/BF02359887. [DOI] [PubMed] [Google Scholar]
- Bergen SE, Gardner CO, Kendler KS. Age-Related Changes in Heritability of Behavioral Phenotypes over Adolescence and Young Adulthood: A Meta-Analysis. Twin Res Hum Genet. 2007;10:423–433. doi: 10.1375/twin.10.3.423. [DOI] [PubMed] [Google Scholar]
- Blacker D, Haines JL, Rodes L, Terwedow H, Go RC, Harrell LE, et al. Apoe-4 and Age at Onset of Alzheimer's Disease: The Nimh Genetics Initiative. Neurology. 1997;48(1):139–147. doi: 10.1212/wnl.48.1.139. [DOI] [PubMed] [Google Scholar]
- Bloom JS, Ehrenreich IM, Loo WT, Lite TL, Kruglyak L. Finding the Sources of Missing Heritability in a Yeast Cross. Nature. 2013;494(7436):234–237. doi: 10.1038/nature11867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boker S, Neale M, Maes H, Wilde M, Spiegel M, Brick T, et al. Openmx: An Open Source Extended Structural Equation Modeling Framework. Psychometrika. 2011;76(2):306–317. doi: 10.1007/s11336-010-9200-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouchard TJ. The Wilson Effect: The Increase in Heritability of Iq with Age. Twin Res Hum Genet. 2013;16(5):923–930. doi: 10.1017/thg.2013.54. [DOI] [PubMed] [Google Scholar]
- Bouchard TJ, McGue M. Familial Studies of Intelligence: A Review. Science. 1981;212:1055–1059. doi: 10.1126/science.7195071. [DOI] [PubMed] [Google Scholar]
- Chipuer HM, Rovine MJ, Plomin R. Lisrel Modeling: Genetic and Environmental Influences on IQ Revisited. Intelligence. 1990;14(1):11–29. [Google Scholar]
- Christiansen K, McGue M. Growing Old but Not Growing Apart: Twin Aimilarity in the Latter Half of the Lifespan. Behavior Genetics. 2013;43:1–12. doi: 10.1007/s10519-012-9559-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies G, Harris SE, Reynolds CA, Payton A, Knight HM, Liewald DC, et al. A Genome-Wide Association Study Implicates the Apoe Locus in Nonpathological Cognitive Ageing. Mol Psychiatry. 2014;19(1):76–87. doi: 10.1038/mp.2012.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies G, Tenesa A, Payton A, Yang J, Harris SE, Liewald D, et al. Genome-Wide Association Studies Establish That Human Intelligence Is Highly Heritable and Polygenic. Molecular Psychiatry. 2011;16(10):996–1005. doi: 10.1038/mp.2011.85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deary IJ, Yang J, Davies G, Harris SE, Tenesa A, Liewald D, et al. Genetic Contributions to Stability and Change in Intelligence from Childhood to Old Age. Nature. 2012;481(7384):212–215. doi: 10.1038/nature10781. [DOI] [PubMed] [Google Scholar]
- Eichler EE, Flint J, Gibson G, Kong A, Leal SM, Moore JH, et al. Missing Heritability and Strategies for Finding the Underlying Causes of Complex Disease. Nat Rev Genet. 2010;11(6):446–450. doi: 10.1038/nrg2809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Erlenmeyer-Kimling L, Jarvik LF. Genetics and Intelligence: A Review. Science. 1963;142:1477–1479. doi: 10.1126/science.142.3598.1477. [DOI] [PubMed] [Google Scholar]
- Falconer DS, Mackay TFC. Introduction to Quantitative Genetics. Longman; 1996. [Google Scholar]
- Finch CE, Kirkwood TB. Chance, Development, and Aging. Oxford University Press; 200. [Google Scholar]
- Finkel D, Reynolds CA. Behavioral Genetic Investigations of Cognitive Aging. In: Kim YK, editor. Handbook of Behavior Genetics. New York: Springer; 2009. pp. 101–112. [Google Scholar]
- Finkel D, Reynolds CA. Cognitive and Physical Aging Pathways: Contributions from Behavioral Genetics. In: Cavanaugh JC, Cavanaugh CK, editors. Aging in America: Volume 2: Physical and Mental Health. Denver, CO: Praeger Publishing; 2010. pp. 26–56. [Google Scholar]
- Finkel D, Reynolds CA, Larsson M, Gatz M, Pedersen NL. Both Odor Identification and Apoe-Epsilon4 Contribute to Normative Cognitive Aging. Psychol Aging. 2011;26(4):872–883. doi: 10.1037/a0023371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finkel D, Reynolds CA, McArdle JJ, Gatz M, Pedersen NL. Latent Growth Curve Analyses of Accelerating Decline in Cognitive Abilities in Late Adulthood. Developmental Psychology. 2003;39(3):535–550. doi: 10.1037/0012-1649.39.3.535. [DOI] [PubMed] [Google Scholar]
- Gatz M, Jang JY, Karlsson IK, Pedersen NL. Dementia: Genes, Environments, Interactions. In: Finkel D, Reynolds CA, editors. Behavior Genetics of Cognition across the Lifespan. Vol. 1. Springer; New York: 2014. pp. 201–231. [Google Scholar]
- Gusev A, Bhatia G, Zaitlen N, Vilhjalmsson BJ, Diogo D, Stahl EA, et al. Quantifying Missing Heritability at Known Gwas Loci. PLoS Genet. 2013;9(12):e1003993. doi: 10.1371/journal.pgen.1003993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton WD. The Moulding of Senescence by Natural Selection. J Theor Biol. 1966;12:12–45. doi: 10.1016/0022-5193(66)90184-6. [DOI] [PubMed] [Google Scholar]
- Heath AC, Berg K, Eaves LJ, Solaas MH, Corey LA, Sundet J, et al. Education Policy and the Heritability of Educational Attainment. Nature. 1985;314(6013):734–736. doi: 10.1038/314734a0. [DOI] [PubMed] [Google Scholar]
- Hemani G, Knott S, Haley C. An Evolutionary Perspective on Epistasis and the Missing Heritability. PLoS Genet. 2013;9(2):e1003295. doi: 10.1371/journal.pgen.1003295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill WG, Goddard ME, Visscher PM. Data and Theory Point to Mainly Additive Genetic Variance for Complex Traits. PLoS Genet. 2008;4(2):e1000008. doi: 10.1371/journal.pgen.1000008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honea RA, Cruchaga C, Perea RD, Saykin AJ, Burns JM, Weinberger DR, et al. Characterizing the Role of Brain Derived Neurotrophic Factor Genetic Variation in Alzheimer's Disease Neurodegeneration. PloS one. 2013;8(9):e76001. doi: 10.1371/journal.pone.0076001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Horn JL, Cattell RB. Age Differences in Primary Mental Ability Factors. J Gerontol. 1966;21(2):210–220. doi: 10.1093/geronj/21.2.210. [DOI] [PubMed] [Google Scholar]
- Horn JL, Cattell RB. Age Differences in Fluid and Crystallized Intelligence. Acta Psychol (Amst) 1967;26(2):107–129. doi: 10.1016/0001-6918(67)90011-x. [DOI] [PubMed] [Google Scholar]
- Hox J. In: Multilevel Analysis: Techniques and Applications. Kindle, editor. New York: Routledge; Taylor and Francis; 2010. [Google Scholar]
- Johansson B, Hofer SM, Allaire JC, Maldonado-Molina MM, Piccinin AM, Berg S, et al. Change in Cognitive Abilities in the Oldest Old: The Effects of Proximity to Death in Genetically Related Individuals over a 6-Year Period. Psychology and Aging. 2004;19:145–156. doi: 10.1037/0882-7974.19.1.145. [DOI] [PubMed] [Google Scholar]
- Johnson W, McGue M, Deary IJ. Normative Cognitive Aging. In: Finkel D, Reynolds CA, editors. Behavior Genetics of Cognition across the Lifespan. New York: Springer; 2014. pp. 169–200. [Google Scholar]
- Kambeitz JP, Bhattacharyya S, Kambeitz-Ilankovic LM, Valli I, Collier DA, McGuire P. Effect of Bdnf Val (66) Met Polymorphism on Declarative Memory and Its Neural Substrate: A Meta-Analysis. Neuroscience and biobehavioral reviews. 2012;36(9):2165–2177. doi: 10.1016/j.neubiorev.2012.07.002. [DOI] [PubMed] [Google Scholar]
- Kaprio J. Twins and the Mystery of Missing Heritability: The Contribution of Gene-Environment Interactions. J Intern Med. 2012;272(5):440–448. doi: 10.1111/j.1365-2796.2012.02587.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirkwood TBL, Cordell HJ, Finch CE. Speed-Bumps Ahead for the Genetics of Later-Life Diseases. Trends in Genetics. 2011;27(10):387–388. doi: 10.1016/j.tig.2011.07.001. [DOI] [PubMed] [Google Scholar]
- Koch L. Epigenetics: An Epigenetic Twist on the Missing Heritability of Complex Traits. Nat Rev Genet. 2014;15(4):218. doi: 10.1038/nrg3698. [DOI] [PubMed] [Google Scholar]
- Koots KR, Gibson JP. Realized Sampling Variances of Estimates of Genetic Parameters and the Difference between Genetic and Phenotypic Correlations. Genetics. 1996;143:1409–1416. doi: 10.1093/genetics/143.3.1409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lambert JC, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R, Bellenguez C, et al. Meta-Analysis of 74,046 Individuals Identifies 11 New Susceptibility Loci for Alzheimer's Disease. Nature genetics. 2013;45(12):1452–1458. doi: 10.1038/ng.2802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee T, Henry JD, Trollor JN, Sachdev P. Genetic Influences on Cognitive Functions in the Eldelry: A Selective Review of Twin Studies. Brain Research Review. 2010;64(1-13) doi: 10.1016/j.brainresrev.2010.02.001. [DOI] [PubMed] [Google Scholar]
- Lessov-Schlagger CN, Swan GE, Reed T, Wolf PA, Carmelli D. Longitudinal Genetic Analysis of Executive Function in Elderly Men. Neurobiology of Aging. 2007;28:1759–1768. doi: 10.1016/j.neurobiolaging.2006.07.018. [DOI] [PubMed] [Google Scholar]
- Liu DJ, Leal SM. Estimating Genetic Effects and Quantifying Missing Heritability Explained by Identified Rare-Variant Associations. Am J Hum Genet. 2012;91(4):585–596. doi: 10.1016/j.ajhg.2012.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maher B. Personal Genomes: The Case of the Missing Heritability. Nature. 2008;456(7218):18–21. doi: 10.1038/456018a. [DOI] [PubMed] [Google Scholar]
- Maki-Tanila A, Hill WG. Influence of Gene Interaction on Complex Trait Variation with Multilocus Models. Genetics. 2014;198(1):355–367. doi: 10.1534/genetics.114.165282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mandelman SD, Grigorenko EL. Bdnf Val66met and Cognition: All, None, or Some? A Meta-Analysis of the Genetic Association. Genes, Brain and Behavior. 2012;11(2):127–136. doi: 10.1111/j.1601-183X.2011.00738.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McArdle JJ, Ferrer-Caja E, Hamagami F, Woodcock RW. Comparative Longitudinal Structural Analyses of the Growth and Decline of Multiple Intellectual Abilities over the Life Span. Dev Psychol. 2002;38(1):115–142. [PubMed] [Google Scholar]
- McArdle JJ, Hamagami F, Meredith W, Bradway KP. Modeling the Dynamic Hypotheses of Gf-Gc Theory Using Longitudinal Life-Span Data. Learning and Individual Differences. 2000;12(1):53–79. [Google Scholar]
- McArdle JJ, Prescott CA, Hamagami F, Horn JL. A Contemporary Method for Developmental-Genetic Analyses of Age Changes in Intellectual Abilities. Developmental Neuropsychology. 1998;14:69–114. [Google Scholar]
- McCartney K, Harris MJ, Bernieri F. Growing up and Growing Apart: A Development Meta-Analysis of Twin Studies. Psychological Bulletin. 1990;107(2):226–237. doi: 10.1037/0033-2909.107.2.226. [DOI] [PubMed] [Google Scholar]
- Muthén LK, Muthén BO. Mplus User's Guide. Seventh. Los Angeles, CA: Muthén & Muthén; 1998-2012. [Google Scholar]
- Pedersen NL, Christensen K, Dahl AK, Finkel D, Franz CE, Gatz M, et al. Igems: The Consortium on Interplay of Genes and Environment across Multiple Studies. Twin Res Hum Genet. 2013;16(1):481–489. doi: 10.1017/thg.2012.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plomin R. Commentary: Missing Heritability, Polygenic Scores, and Gene-Environment Correlation. J Child Psychol Psychiatry. 2013;54(10):1147–1149. doi: 10.1111/jcpp.12128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plomin R, DeFries JC, Knopik VS, Neiderhiser JM. Behavioral Genetics. 6th. New York: Worth; 2013. [Google Scholar]
- Plomin R, Haworth CMA, Meaburn EL, Price TS, Davis OSP. Common DNA Markers Can Account for More Than Half of the Genetic Influence on Cognitive Abilities. Psychological Science. 2013;24(4):562–568. doi: 10.1177/0956797612457952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prescott CA. Using the Mplus Computer Program to Estimate Models for Continuous and Categorical Data from Twins. Behavior Genetics. 2004;34(1):17–40. doi: 10.1023/B:BEGE.0000009474.97649.2f. [DOI] [PubMed] [Google Scholar]
- Raz N. Aging of the Brain and Its Impact on Cognitive Performance: Integration of Structural and Functional Findings. In: Craik FIM, Salthouse TA, editors. The Handbook of Aging and Cognition. 2nd. Hillsdale, NJ: Erlbaum; 2000. pp. 1–90. [Google Scholar]
- Reynolds CA, Finkel D. Cognitive and Physical Aging: Genetic Influences and Gene-Environment Interplay. In: Schaie KW, Williis SL, editors. Handbook of the Psychology of Aging. 8th. Academic Press; 2014. in press. [Google Scholar]
- Reynolds CA, Finkel D, McArdle JJ, Gatz M, Berg S, Pedersen NL. Quantitative Genetic Analysis of Latent Growth Curve Models of Cognitive Abilities in Adulthood. Developmental Psychology. 2005;41:3–16. doi: 10.1037/0012-1649.41.1.3. [DOI] [PubMed] [Google Scholar]
- Reynolds CA, Finkel D, Zavala C. Gene by Environment Interplay in Cognitive Aging. In: Finkel D, Reynolds CA, editors. Behavior Genetics of Cognition across the Lifespan. Vol. 1. New York: Springer; 2014. pp. 169–199. [Google Scholar]
- Reynolds CA, Gatz M, Berg S, Pedersen NL. Genotype-Environment Interactions: Cognitive Aging and Social Factors. Twin Res Hum Genet. 2007;10(2):241–254. doi: 10.1375/twin.10.2.241. [DOI] [PubMed] [Google Scholar]
- Reynolds CA, Phillips D. Sciences. Elsevier; 2014. Genomics of Brain Aging - Twin Aging Encyclopedia of Biomedical. under review. [Google Scholar]
- Reynolds CA, Prince JA, Feuk L, Brookes AJ, Gatz M, Pedersen NL. Longitudinal Memory Performance During Normal Aging: Twin Association Models of Apoe and Other Alzheimer Candidate Genes. Behav Genet. 2006;36(2):185–194. doi: 10.1007/s10519-005-9027-6. [DOI] [PubMed] [Google Scholar]
- Reynolds CA, Zavala C, Gatz M, Vie L, Johansson B, Malmberg B, et al. Sortilin Receptor 1 Predicts Longitudinal Cognitive Change. Neurobiology of Aging. 2013;34(6):1710 e1711–1718. doi: 10.1016/j.neurobiolaging.2012.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ridge PG, Mukherjee S, Crane PK, Kauwe JS, Alzheimer's Disease, Genetics C. Alzheimer's Disease: Analyzing the Missing Heritability. PLoS One. 2013;8(11):e79771. doi: 10.1371/journal.pone.0079771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowe SJ, Rowlatt A, Davies G, Harris SE, Porteous DJ, Liewald DC, et al. Complex Variation in Measures of General Intelligence and Cognitive Change. PLoS One. 2013;8(12):e81189. doi: 10.1371/journal.pone.0081189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scarr S, McCartney K. How People Make Their Own Environments: A Theory of Genotype Greater Than Environment Effects. Child development. 1983;54(2):424–435. doi: 10.1111/j.1467-8624.1983.tb03884.x. [DOI] [PubMed] [Google Scholar]
- Schaie KW. Intellectual Development in Adulthood. In: Birren JE, Schaie KW, editors. Handbook of the Psychology of Aging. 4th. New York: Academic Press; 1996. pp. 266–286. [Google Scholar]
- Sham P. Statistics in Human Genetics. New York: John Wiley & Sons Inc; 1998. [Google Scholar]
- Trzaskowski M, Dale PS, Plomin R. No Genetic Influence for Childhood Behavior Problems from DNA Analysis. J Am Acad Child Adolesc Psychiatry. 2013;52(10):1048–1056 e1043. doi: 10.1016/j.jaac.2013.07.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Dongen J, Boomsma DI. The Evolutionary Paradox and the Missing Heritability of Schizophrenia. Am J Med Genet B Neuropsychiatr Genet. 2013;162B(2):122–136. doi: 10.1002/ajmg.b.32135. [DOI] [PubMed] [Google Scholar]
- van Dongen J, Slagboom PE, Draisma HH, Martin NG, Boomsma DI. The Continuing Value of Twin Studies in the Omics Era. Nature reviews Genetics. 2012;13(9):640–653. doi: 10.1038/nrg3243. [DOI] [PubMed] [Google Scholar]
- Visscher PM, Hill WG, Wray NR. Heritability in the Genomics Era--Concepts and Misconceptions. Nat Rev Genet. 2008;9(4):255–266. doi: 10.1038/nrg2322. [DOI] [PubMed] [Google Scholar]
- Yang J, Lee SH, Goddard ME, Visscher PM. Gcta: A Tool for Genome-Wide Complex Trait Analysis. Am J Hum Genet. 2011;88(1):76–82. doi: 10.1016/j.ajhg.2010.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang C, Pierce BL. Genetic Susceptibility to Accelerated Cognitive Decline in the Us Health and Retirement Study. Neurobiology of Aging. 2014;35(6):1512.e1511–1512.e1518. doi: 10.1016/j.neurobiolaging.2013.12.021. [DOI] [PubMed] [Google Scholar]
- Zhang G, Karns R, Sun G, Indugula SR, Cheng H, Havas-Augustin D, et al. Finding Missing Heritability in Less Significant Loci and Allelic Heterogeneity: Genetic Variation in Human Height. PLoS One. 2012;7(12):e51211. doi: 10.1371/journal.pone.0051211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zuk O, Hechter E, Sunyaev SR, Lander ES. The Mystery of Missing Heritability: Genetic Interactions Create Phantom Heritability. Proc Natl Acad Sci U S A. 2012;109(4):1193–1198. doi: 10.1073/pnas.1119675109. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental table 1. Fit statistics (-2LL) of meta-analysis of heritability.
Supplemental table 2. Parameter estimates (standard errors) of meta-analysis of heritability.
Supplemental table 3. Fit statistics and criteria: meta analysis of MZ/DZ correlation ratios.