

Abstract
Research over the past 15 years has established that defective primary cilia play a fundamental role in the pathogenesis of various genetic and acquired human diseases now termed ciliopathies. A new study, published in this issue of EMBO reports, expands the spectrum of cilia‐related diseases by uncovering a role for primary cilia in preventing cardiovascular disease. Dinsmore and Reiter show that lack of endothelial cilia in the aorta results in accelerated development of atherosclerotic lesions in a mouse model of atherosclerosis on a high‐fat, high‐cholesterol diet. Loss of endothelial cilia further increases inflammatory gene expression and reduces vasoprotective eNOS activity 1. In summary, this interesting study identifies a role for aortic endothelial cilia to protect against atherosclerosis.
Subject Categories: Cell Adhesion, Polarity & Cytoskeleton; Molecular Biology of Disease
Virtually all mammalian cell types carry primary cilia, little microtubule‐based organelles that project from the cell surface and serve as antennae to receive signals from the microenvironment. Cilia express various receptors and signaling molecules to locally restrict, inhibit, or promote signaling evoked through hormonal, chemical, and mechanical stimuli (reviewed in 2). They control a variety of cellular programs including cell differentiation, proliferation as well as directed cell migration and polarization. Mutations of genes involved in the formation and function of cilia have been linked to several disorders ranging from cystic kidney disease and retinopathies to hydrocephalus, diabetes, obesity, and complex malformations and syndromes 3.
… the study […] provides important insight into the role of cilia on endothelial cells and may open a new field of research in vascular biology
Cilia have been described on endothelial cells in the past. They project into the lumen of the blood vessel and are thought to be exposed to mechanical strain through the blood stream (reviewed in 4). Recent data showed that endothelial cilia are essential for the developmental vascular integrity in zebra fish 5. However, the study by Dinsmore and Reiter could not recapitulate these findings in mice, which may be explained by differences in the approach or species‐specific characteristics. Endothelial‐specific deletion of the intraflagellar transport (IFT) protein Ift88 clearly removed endothelial cilia in adult mice without affecting vascular development. Dinsmore and Reiter quantified the non‐uniform distribution of cilia in the adult aorta confirming previous findings that cilia are enriched at vascular branch points and sites of high curvature, known predilection sites of atherosclerotic plaques 6. Therefore, a role for endothelial cilia in inducing anti‐atherogenic programs could be envisioned 7. Indeed, the study shows that loss of cilia on endothelial cells in the aorta predisposes a mouse model of artherosclerosis toward the development of the disease 1. These findings are exciting and potentially clinically relevant. The authors further performed gene expression analyses, which showed upregulation of pro‐inflammatory genes in aortas lacking cilia. These results may open a new avenue for research into cilia‐controlled anti‐inflammatory signaling. Inflammation also plays a major role in the pathogenesis of atherosclerosis.
In response to shear stress, endothelial cells generate a number of soluble factors that lead to relaxation of smooth muscle cells and vasodilation. The most prominent of these endothelial‐derived relaxing factors is nitric oxide (NO), which beyond vasodilation is vasoprotective by interfering with the proliferation of smooth muscle cells, inflammation, and platelet function. Interestingly, the ciliopathy protein polycystin‐2 has been suggested to act as a ciliary mechanosensor responding to shear stress in a cilia‐dependent manner by increasing the production of nitric oxide 8. However, aortic endothelial cells typically form cilia in regions of the aorta where shear stress is absent, which is consistent with a report demonstrating that laminar shear stress induces ciliary disassembly 9. Therefore, the role of polycystin‐2 in endothelial cells remains unclear. Mutations in PKD2 encoding for polycystin‐2 are responsible for approximately 15% of autosomal‐dominant polycystic kidney disease (ADPKD) cases 3. It has long been known that ADPKD patients can display cardiovascular abnormalities. Investigating potential mechanisms for the protective effect of endothelial cilia, Dinsmore and Reiter analyzed Pkd2 expression in vivo. Besides the known expression of Pkd2 in cilia, they found a large pool of extra‐ciliary Pkd2 protein in the aortic endothelium. In addition, following the idea that cilia and Pkd2 might be required for the activation of endothelial nitric oxide synthase (eNOS), they demonstrate decreased phosphorylation of eNOS in mice lacking endothelial cilia. These results cannot ultimately clarify the functional contribution of endothelial cilia in the protection from atherosclerosis but suggest that loss of endothelial cilia is a pro‐atherogenic event.
Figure 1. In the aorta, primary cilia are predominantly found in regions exposed to disturbed laminar flow matching with predilection sites of atherosclerosis.
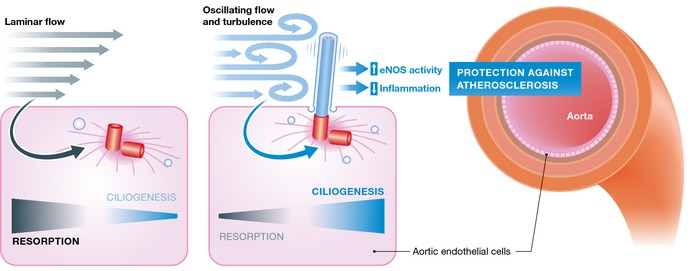
The study by Dinsmore and Reiter suggests that cilia may suppress pro‐atherogenic factors and protect against atherosclerotic lesions.
However, there are caveats. Dinsmore and Reiter abrogated endothelial ciliogenesis through genetic deletion of the IFT protein Ift88. This approach does not necessarily link the protective effects by loss of Ift88 to cilia. Ift88 has been shown to contribute to cilia‐independent functions such as transport of various proteins in the secretory pathway. To rule out that cilia‐independent effects may promote susceptibility to atherogenesis in Ift88 mutants, the authors recapitulated some of the findings by genetic deletion of Kif3a, a motor protein required for cilia formation. While the data provided are clear and the models well established, these results do not necessarily rule out cilia‐independent effects. Firstly, Kif3a is in the same complex with Ift88 and may also be involved in cilia‐independent transport mechanisms. Secondly and more importantly, the authors did not demonstrate a detrimental role for Kif3a‐mediated cilia loss in the atherogenesis model, indicating that the interesting results need to be interpreted with some caution. Furthermore, deletion of cilia may cause a rather unspecific damage to endothelial cells and repair defects, which may then generate loci of minor resistance to atherogenic events. Thus, while the study by Dinsmore and Reiter is exciting and potentially of clinical relevance, further studies are required to confirm the contribution of ciliary signaling in the protection against endothelial dysfunction and atherosclerosis.
The study by Dinsmore and Reiter provides important insight into the role of cilia on endothelial cells and may open a new field of research in vascular biology. However, it also brings up new questions. What are the mechanisms that restrict ciliogenesis to vascular branch points and sites of high curvature? Do ciliated endothelial cells play a similar role in smaller arteries that are typically affected by atherosclerosis? How does aging affect the formation and presence of endothelial cilia? These and more questions will need to be tackled in the future, but the current study raises hope that therapeutic approaches through the control of ciliogenesis may be helpful in the prevention or treatment of atherosclerosis.
See also: C Dinsmore & JF Reiter (February 2016)
References
- 1. Dinsmore C, Reiter JF (2016) EMBO Rep. 17: 156–166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Gerdes JM, Davis EE, Katsanis N (2009) Cell 137: 32–45 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Hildebrandt F, Benzing T, Katsanis N (2011) N Engl J Med 364: 1533–1543 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Egorova AD, van der Heiden K, Poelmann RE, Hierck BP (2012) Differentiation 83: S56–S61 [DOI] [PubMed] [Google Scholar]
- 5. Kallakuri S, Yu JA, Li J, Li Y, Weinstein BM, Nicoli S, Sun Z (2015) J Am Soc Nephrol 26: 864–875 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Van der Heiden K, Hierck BP, Krams R, de Crom R, Cheng C, Baiker M, Pourquie MJ, Alkemade FE, DeRuiter MC, Gittenberger‐de Groot AC et al (2008) Atherosclerosis 196: 542–550 [DOI] [PubMed] [Google Scholar]
- 7. Egorova AD, Khedoe PP, Goumans MJ, Yoder BK, Nauli SM, ten Dijke P, Poelmann RE, Hierck BP (2011) Circ Res 108: 1093–1101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. AbouAlaiwi WA, Takahashi M, Mell BR, Jones TJ, Ratnam S, Kolb RJ, Nauli SM (2009) Circ Res 104: 860–869 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Iomini C, Tejada K, Mo W , Vaananen H, Piperno G (2004) J Cell Biol 164: 811–817 [DOI] [PMC free article] [PubMed] [Google Scholar]