

Abstract
Capsular polysaccharide A (CPSA) is a four-sugar repeating unit polymer found on the surface of the gut symbiont Bacteroides fragilis that has therapeutic potential in animal models of autoimmune disorders. This therapeutic potential has been credited to its zwitterionic character derived from a positively charged N-acetyl-4-aminogalactosamine (AADGal) and a negatively charged 4,6-O-pyruvylated galactose (PyrGal). In this report, using a fluorescent polyisoprenoid chemical probe, the complete enzymatic assembly of the CPSA tetrasaccharide repeat unit is achieved. The proposed pyruvyltransferase, WcfO, galactopyranose mutase, WcfM, and glycosyltransferases, WcfP and WcfN, encoded by the CPSA biosynthesis gene cluster were heterologously expressed and functionally characterized. Pyruvate modification, catalyzed by WcfO, was found to occur on galactose of the polyisoprenoid-linked disaccharide (AADGal-Gal), and did not occur on galactose linked to uridine diphosphate (UDP) or a set of nitrophenyl-galactose analogues. This pyruvate modification was also found to be required for the incorporation of the next sugar in the pathway N-acetylgalactosamine (GalNAc) by the glycosyltransferase WcfP. The pyruvate acetal modification of a galactose has not been previously explored in the context of a polysaccharide biosynthesis pathway, and this work demonstrates the importance of this modification to repeat unit assembly. Upon production of the polyisoprenoid-linked AADGal-PyrGal-GalNAc, the proteins WcfM and WcfN were found to work in concert to form the final tetrasaccharide, where WcfM formed UDP-galactofuranose (Galf) and WcfN transfers Galf to the AADGal-PyrGal-GalNAc. This work demonstrates the first enzymatic assembly of the tetrasaccharide repeat unit of CPSA in a sequential single pot reaction.
Keywords: capsular polysaccharide, undecaprenyl, bactoprenyl, pyruvyltransferase, in vitro reconstitution
Introduction
Capsular polysaccharide A (CPSA), produced by the Gram negative microfloral organism Bacteroides fragilis, plays an important role in mammalian immune system development,1,2 and has been suggested to have protective effects in model systems for autoinflammatory diseases, including multiple sclerosis.3-5 The polymer, isolated from B. fragilis, has also recently been used as a carbohydrate antigen to replace classic antigenic proteins, like cationic BSA and KLH, for vaccine development.6, 7 CPSA is made up of the four-sugar repeating unit →3) α-D-2-N-acetylamido-4-amino-galactopyranose (AADGal)-(1→4)-(β-D-galactofuranose (Galf)-(1→3)) α-D-N-acetylamido-galactopyranose (GalNAc)-(1→3)-(4,6-O-pyruvate)-β-D-galactopyranose (PyrGal)-(1→ (Figure 1a).8,9 Exceptional work has been done in understanding the biological mechanisms by which the relatively unique CPSA zwitterionic capsule is associated with the symbiotic effect of the organism on its mammalian host.10, 11, 12 Akin to how peptides are presented by major histocompatibility complex (MHC) molecules, CPSA fragments are internalized by antigen presenting cells, then chemically processed and displayed by MHC II molecules. Once displayed, naïve T-cells are activated and differentiate to Th1 helper cells, which promote a healthy balance between Th1 and Th2. Chemically masking the zwitterion of CPSA depletes biological function.
Figure 1. CPSA structure and proposed pathways for its production.

A) CPSA tetrasaccharide repeat unit structure B) Proposed routes for the incorporation of pyruvate in to the tetrasaccharide repeat unit
Genetic studies have demonstrated that the CPSA polymer is produced via a Wzy-dependent pathway encoded by a single genetic operon.13 In the typical Wzy-dependent pathway, each sugar is appended, one at-a-time, to a hydrophobic C55 bactoprenyl phosphate (BP) embedded in the inner membrane of the bacterial cell. Once the repeating unit is completed, it is then flipped to the periplasmic side of the inner membrane, polymerized by a Wzy polymerase, and then exported out of the cell.14-16 While the overall general pathway for these types of polysaccharides is well understood, the precise roles of each protein involved is not clearly defined.
One of the major challenges associated with the characterization of proteins involved in polysaccharide biosynthesis is the lack of clearly defined and easily quantifiable substrates. A recent advance that avoids many of the challenges associated with traditional methods is the development of optically active tagged isoprenoids that allow simple substrate and product tracking and quantification by reverse-phase high performance liquid chromatography (HPLC).17-19 These methods were applied to identify the protein WcfS as an initiating transferase that transfers AADGal-phosphate to BP, followed by WcfQ, which transfers galactose to the bactoprenyl diphosphate-linked AADGal (BPP-AADGal) to form the CPSA repeat unit precursor BPP-AADGal-Gal.
While the proteins responsible for forming the positively charged AADGal moiety and for incorporating it into the repeat unit have been identified,20 it is not clear how or when the negatively charged pyruvate moiety is formed. One could expect pyruvate incorporation to occur before, during or after repeat unit assembly (Figure 1b). Few pyruvyltransferases that form the pyruvate acetal have been characterized.21-23 In addition, none have been biochemically characterized in the context of a biosynthesis pathway for a complex polysaccharide.
Because the zwitterionic nature of CPSA is critical for its biological function, identifying the protein responsible, in the context of the biosynthesis pathway and its temporal position, is critical. Knowing how this material can be produced enzymatically provides alternatives to de novo chemical synthesis or isolation from the surface of the difficult to grow anaerobic microbe. Enzymatic methods, in particular, will also prove useful in the biochemical analysis of elusive flippases, polymerases and transport proteins downstream of repeat unit assembly.
Results
Expression of the proposed Bacteroides fragilis pyruvyltransferase
The CPSA biosynthesis gene cluster has one gene that emerges, that is homologous to pyruvyltransferase family members based on a pBLAST search on the encoded protein sequences.24 This gene, wcfO, encodes a 347 amino acid protein with less than 45% sequence identity to any other protein in the NCBI database, and has very low similarity to any biochemically characterized pyruvyltransferase. The closest similarity of a biochemically characterized 4,6-O-pyruvyltransferase is the 29% similar, 14% identical PssM protein from Rhizobium leguminosarum (Supporting Figure 1).25 However, since there were no other probable candidates in the CPSA biosynthesis gene cluster and there was an apparent homology to the pyruvyltransferase family of proteins, the wcfO gene was amplified from genomic DNA and inserted into a pET-24a vector for expression in E. coli BL-21(DE3) cells. As recently observed with other proteins encoded by the CPSA biosynthesis gene cluster, WcfO fractionated, upon cell lysis, with the bacterial cell membrane, even though there were no clear transmembrane domains or hydrophobic patches detected by sequence analysis (Supporting Figure 2).19 Unlike the glycosyltransferases encoded by the cluster, WcfO could be separated from the membrane fraction and purified to homogeneity (Figure 2). The total purified protein yield from a 500 mL expression culture was 1.31 mg.
Figure 2. WcfO isolated as membrane fraction and purified protein.
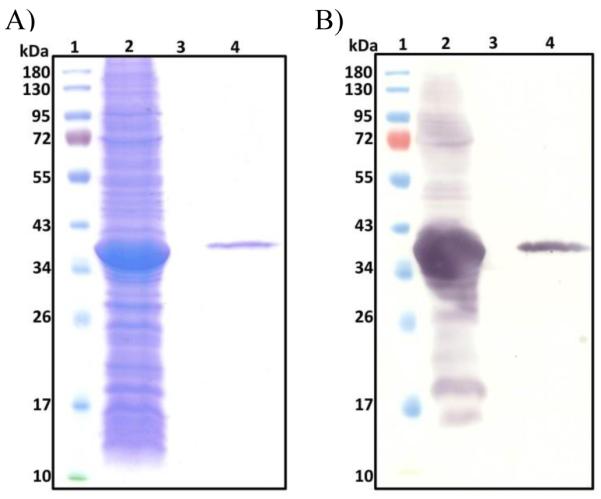
A) SDS-PAGE and B) Anti-His western blot analysis of WcfO membrane fraction and purified protein. Lane 1 protein ladder, 2 membrane fraction, 3. blank and 4. solubilized protein.
WcfO catalyzes the addition of pyruvate to isoprenoid-linked disaccharide
Since there was no precedent associated with an acetal forming pyruvyltransferase in a polysaccharide biosynthesis pathway, we envisioned three ways this moiety could be incorporated (Figure 1b). Either 1) the pyruvate would be transferred to galactose (Gal) linked to uridine diphosphate (UDP) that would then be added to the polyisoprenoid-linked precursor, 2) the pyruvate would be transferred to Gal after it is transferred to polyprenyl-linked AADGal, and the modification would be required for subsequent transferase reactions, or 3) the pyruvate is incorporated after the repeat unit is formed. The few 4,6-O-pyruvyltransferases that had been previously described used phosphoenolpyruvate (PEP) as the pyruvate donor, and were able to utilize an easily detectable nitroaromatic-sugar as the acceptor for the transferase.21 To test if the isolated WcfO was functional, α-p-nitrophenyl and β-o-nitrophenyl galactopyranoside were used as potential pyruvate acceptors. However, no change was observed in HPLC retention times of these sugars in the presence of WcfO and PEP (Supporting figure 3a). This suggested that either: WcfO was isolated as a nonfunctional protein, it was not a pyruvyltransferase or, unlike the previously characterized pyruvyltransferases, Gal alone was not enough for recognition by the protein. In addition, when WcfO was mixed with PEP and UDP-Gal, no product formation was observed by capillary electrophoresis (Supporting figure 3b), a high resolution technique that would be expected to readily distinguish between even very small changes in nucleotide-linked sugar structures.26, 27
In order to test the ability of WcfO to function on polyisoprenoid-linked AADGal-Gal, a fluorescent 2-nitrileanilinobactoprenyl monophosphate was prepared with four Z-configuration isoprene units (2CNA-B(4Z)P, Figure 3a) using conditions similar to previous reports.28 The 2CNA-B(4Z)P was then extended using WcfS and WcfQ with the appropriate sugar donors to form 2-nitrileanilino-bactoprenyl diphosphate-linked (2CNA-B(4Z)PP) AADGal-Gal as a potential fluorescent substrate for WcfO. When PEP was mixed with 2CNA-B(4Z)PP-AADGal-Gal a major shift in retention of 1.6 minutes was observed in the presence of WcfO (Figure 3b). The change in retention time was consistent with the introduction of the negatively charged pyruvate moiety. Similar results were observed with 2CNA-B(nZ)PP-AADGal-Gal with n=5 or 6 Z-configuration isoprene units, and analogues containing anthranilamide in the place of the 2-nitrileaniline (data not shown). It is important to note that no claims are made here towards the specific linkage position of each sugar. These are presumed based on the known structure of the CPSA repeat unit (Figure 1).8
Figure 3. WcfO functions with a bactoprenyl-linked AADGal substrate.

A) Scheme for the preparation of fluorescent 2-nitrileanilinobactoprenyl diphosphate-linked AADGal-Gal. WcfO was incubated with PEP and the disaccharide B) 2CNA-B(4Z)PP-AADGal-Gal or C) 2CNA-B(4Z)PP-AADGal-Glc. After 30 minutes the reaction mixture was analyzed by RP-HPLC with 35% n-propanol in 100 mM ammonium bicarbonate as the mobile phase. Control reactions with identical conditions but no enzyme are shown as dotted lines, reaction mixtures that contained WcfO are shown with solid lines. Both disaccharide substrates of WcfO were prepared using WcfQ, which can be used to incorporate Gal or Glc depending on reaction conditions. A new WcfO product with different retention time from substrate is observed with both disaccharide substrates and no detectable starting material remains.
Specificity of WcfO for sugar linker
It was clear from the above experiments that WcfO catalyzed a change in polyisoprenoid-linked sugar structure and was selective for the pyruvate acceptor, with no activity observed on Gal that was not linked to AADGal. In previous work, it was found that the glycosyltransferase WcfQ could be used to also transfer glucose (Glc) to AADGal.19 WcfO was tested with a 2CNA-B(4Z)PP-AADGal-Glc substrate using conditions identical to those described above with the Gal substrate. Surprisingly, while WcfO was selective for what the Gal was linked to, it was not selective for Gal over Glc (Figure 3c). In addition, the retention time of the WcfO product with Gal was identical to the retention time of the Glc product, suggesting similar structural characteristics, although again the precise linkages have not been identified. To test whether BPP-AADGal-Gal was the preferred substrate of WcfO and therefore the more likely natural substrate, a kinetic analysis was performed using the fluorescent disaccharides 2-amideaniline (2AA) B(6Z)PP-AADGal-Gal and 2AA-B(6Z)PP-AADGal-Glc. The anthranilamide analogues were used because of the superior fluorescent properties and ease of detection for small quantities of products formed in kinetic analyses. Using a timed quench assay with varying PEP concentrations, initial reaction rates were determined then fit to the Michaelis-Menten equation. Very little if any difference was observed in the catalytic efficiency when 2AA-B(6Z)PP-AADGal-Glc was used as a substrate in comparison to 2AA-B(6Z)PP-AADGal-Gal (Table 1). This data suggested that there was effectively no discrimination between Gal and Glc by WcfO. However, it should be noted that the kinetic analysis only focused on apparent kinetic constants associated with the pyruvate donor, with a relatively low concentration of polyisoprenoid. Varying concentrations of the polyisoprenoid substrate could be complicated a by interfacial interactions of the enzyme with surfactant micelles and substrate.29, 30
Table 1. Kinetic parameters with varying phosphoenolpyruvate concentration.
2-amideanilinobactoprenyl diphosphate linked AADGal-Gal and AADGal-Glc were prepared with concentrations of PEP varying from 0.1-5.0 mM. Reactions were quenched with 1-propanol at varying times then analyzed by HPLC to determine initial rates. Apparent kinetic constants with the 0.45 μM isoprenoid were determined by fitting to the Michaelis-Menten equation. Error shown is a reflection of the data fit.
| 2AA-B(6Z)PP-AADGal- Gal |
2AA-B(6Z)PP-AADGal- Glc |
|
|---|---|---|
| appKmPEP | 299 ± 49 μM | 291 ± 46 μM |
| appkcatPEP | 0.249 ± 0.010 s−1 | 0.231 ± 0.009 s−1 |
| appkcat/KmPEP | 833 ± 141 M−1 • s−1 | 795 ± 129 M−1 • s−1 |
Sugar transfer to the WcfO product is acceptor substrate selective
The above data suggested that WcfO could utilize Glc or Gal-containing isoprenoid-linked pyruvylated disaccharides as substrates, yet the published structure of CPSA clearly shows a 4,6-O-pyruvylated galactose.8 Since there was only limited discrimination by WcfO, it was possible that the next step in the pathway would only use one specific WcfO product as a substrate. As previously reported,19 WcfN and WcfP, the two remaining uncharacterized glycosyltransferases encoded by the CPSA biosynthesis locus, were prepared as membrane fractions from BL-21 (DE3) cells overexpressing each. WcfN in the presence of UDP-N-acetylgalactosamine (GalNAc) was ineffective for altering the retention of 2CNA-B(4Z)PP-AADGal-Gal or either of the pyruvylated WcfO products (Gal or Glc-linked) (Supporting Figure 4a-b). However, WcfP did catalyze a reaction between UDP-GalNAc and 2CNA-B(4Z)PP-AADGal-PyrGal (Figure 4a), but not with the Glc containing pyruvylated disaccharide (Figure 4b), or unpyruvylated disaccharide (data not shown). The 0.35 minute retention time shift was much smaller for the HexNAc sugar addition. E. coli BL-21 (DE3) membrane fractions from cells transformed with pET-24a that did not contain the WcfP gene also did not lead to a change in retention of the substrate (data not shown). UDP-N-acetylglucosamine (GlcNAc) was tested with WcfP, WcfN and each WcfO product, but no transfer was observed (Figure 4a-b, Supporting Figure 4a-b), suggesting that WcfP was selective for both the PyrGal acceptor and GalNAc donor substrates.
Figure 4. WcfP transfers UDP-GalNAc to pyruvylated disaccharide.

Reactions containing A) 2CNA-B(4Z)PP-AADGal-PyrGal or B) 2CNA-B(4Z)PP-AADGal-PyrGlc with 1) No sugar donor or enzyme, 2) WcfP and UDP-GalNAc or 3) WcfP and UDP-GlcNAc were analyzed by RP-HPLC with 32% 1-propanol in 100 mM ammonium bicarbonate as the mobile phase. Results confirmed the formation of product with WcfP and UDP-GalNAc only. * indicates the new peak formed and a dotted line is added to help mark the position of the starting material. Peaks at 2 minutes and below are primarily associated with the membrane fractions and are more prominent with the Glc modified disaccharide because the reaction scale was nearly a third for the Gal reactions. A shoulder can be noticed on the peaks in B2 and B3, it is unclear what leads to this other than the lower scale of the reaction, but it cannot be the addition of the HexNAc sugar as this would lead to a shift to a shorter retention time rather than a longer retention time.
WcfN completes assembly of the CPSA repeat unit
The final step in the assembly of the CPSA repeat unit was expected to be the addition of a galactofuranose (Galf) residue to the apparent pyruvylated trisaccharide product of WcfP. Only one glycosyltransferase, WcfN, remained uncharacterized from the CPSA biosynthesis gene cluster. UDP-Galf is not commercially available, so a chemically synthesized UDP-Galf31 was used with WcfN and 2CNA-B(4Z)PP-AADGal-PyrGal-GalNAc to verify the role of WcfN in the biosynthesis pathway (Figure 5a). WcfN catalyzed the formation of a new product in the presence of UDP-Galf but not UDP-galactopyranose (Galp). The change in retention of the product was 0.90 minutes, which was again consistent with the addition of a single furanose unit.
Figure 5. WcfN catalyzes the transfer of galactofuranose completing tetrasaccharide formation.

A) 2CNA-B(4Z)PP-AADGal-PyrGal-GalNAc was mixed with 1) No additional enzyme or sugar donor, 2) UDP-Galactopyranose and WcfN and 3) UDP-Galf and WcfN confirming the role of WcfN in the final step of repeat unit biosynthesis. B) HPLC analysis of 1) Pure UDP-Galf, 2) WcfM reaction with UDP-Galp, 3) Pure UDP-Galp. C) HPLC analysis of reactions containing 2CNA-B(4Z)PP-AADGal-PyrGal-GalNAc with 1) No enzyme or sugar donor, 2) UDP-Galf and WcfN, 3) UDP-Galp, WcfM and WcfN, 4) UDP-Galp and WcfN. D) Starting with bactoprenyl phosphate, enzymes were sequentially added leading to the formation of each intermediate product in the pathway and the single pot formation of the tetrasaccharide. HPLC with isoprenoid linked materials was performed with a mobile phase of 32% 1-Propanol in 100 mM ammonium bicarbonate. HPLC conditions for UDP-linked sugars used a mobile phase of 1.5% acetonitrile in 50mM triethylammonium acetate buffer (pH 6.8).
The protein WcfM was encoded by the CPSA biosynthesis gene cluster and had greater than 90% sequence similarity to biochemically characterized galactopyranose mutases.13, 32-35 Galactopyranose mutases have been difficult to work with as a synthetic tool, as typically only very low yields are obtained for the formation of UDP-Galf in vitro, therefore characterization is typically done on the reverse reaction. WcfM was expressed using methods similar to the other proteins in the CPSA biosynthesis pathway and was isolated (Supporting Figure 5). Consistent with other Galp mutases, WcfM was effective in converting UDP-Galp to UDP-Galf with relatively low yields (Figure 5b). WcfN coupled with WcfM in the presence of UDP-Galp led to the formation of a product with an identical retention time to the product of the reaction with the chemically synthesized UDP-Galf (Figure 5c), and no product was observed when WcfM was left out of the mixture.
Single pot synthesis of isoprene-linked tetrasaccharide
A key advantage of the enzymatic synthesis of materials is the possibility that one can take advantage of the specificity of these proteins to do multiple synthetic transformations in a single pot reaction, without purification of intermediates. To determine if the entire CPSA tetrasaccharide repeat unit could be assembled a set of sequential reactions were prepared in which first a WcfS, 2CNA-BP, and UDP-AADGal reaction was prepared then a small aliquot was analyzed by HPLC (Figure 5d). After confirmation that the reaction was complete WcfQ and UDP-Gal were added and the mixture was again analyzed by HPLC. These results were consistent with previous observations using this same set of enzymes.19, 20 Next WcfO, WcfP, WcfM and WcfN, along with sugar donors, were added stepwise to give the full tetrasaccharide in a single pot preparation (Figure 5d).
Mass characterization of polyprenol-linked products
A surprising challenge in the reconstitution of the CPSA biosynthesis pathway was the mass spectrometric (MS) characterization of the products formed throughout the pathway. Clear differences were observed in the fluorescence-based HPLC analysis of reactions containing 2CNA-bactoprenyl concentrations in the low micromolar range with HPLC injections as small as five pmoles of isoprenoid. However, in our hands, electrospray ionization (ESI)-MS characterization required as much as four nmoles injected into the spectrometer. MALDI-MS with anthracene and DHB matrices were unsuccessful on the polyprenyl diphosphate-linked materials (data not shown). We also found that triton-X-100 (TX-100), the primary surfactant used in these studies, led to serious problems with ESI-MS analysis. Replacing TX-100 with sodium cholate was a key change for improving detection of isolated products. Pyruvulated di-, tri- and tetrasaccharide from the WcfO, WcfP and WcfN reactions with sodium cholate as the surfactant were isolated by HPLC then subjected to negative ion-mode ESI-MS. The mass of the new WcfO products with either Glc or Gal were easily obtainable with as little as 0.1 nmoles of product and the masses were consistent with the addition of the pyruvate acetal (Figure 6a-b). Obtaining the mass on the trisaccharide product of WcfP was more challenging and required from 0.5-1.0 nmoles, yet the signal from these samples was several orders of magnitude lower on the same spectrometer used with the WcfO products. In addition, although the pyruvylated disaccharide was difficult to detect in these mixtures by HPLC, significant signal from contaminating starting material was observed by ESI-MS (Figure 6c). The pyruvylated disaccharide signal is likely due to the improved ESI-MS detection sensitivity to the smaller compound. Lastly, the final mass of the WcfN tetrasaccharide product was obtained, but required 1-4 nmoles of material for detection (Figure 6d). The signal was an order of magnitude lower than the pyruvylated trisaccharide. Although trisaccharide and disaccharide were not readily detected by HPLC from these isolated products, signal was observed by ESI-MS. Signal from the smaller compounds is likely again due to low levels of contamination, but are more prevalent than WcfN product due to enhanced relative detection sensitivity by ESI-MS.
Figure 6. ESI-MS confirmation of tetrasaccharide formation intermediates.

Negative ion mode ESI-MS analysis was performed on HPLC isolated products of A) WcfO with UDP-Gal, expected mass=1119.5, B) WcfO with UDP-Glc, expected mass=1119.5, C) WcfP expected mass=1322.6 and D) WcfN, 1484.6. All masses are within one unit of the expected mass. Note that even though the dominant product by HPLC in C and D was the tri and tetrasaccharide respectively, more signal is observed for the smaller contaminating material consistent with the decrease in sensitivity with the larger oligosaccharides.
Discussion
One of the key findings in this work was the establishment of pyruvylation as a required modification of the CPSA polysaccharide repeat unit precursor BPP-AADGal-Gal. The fact that WcfP is unable to utilize BPP-AADGal-Gal as a substrate and will only modify the pyruvylated disaccharide suggests that this modification must occur during repeat unit biosynthesis and cannot occur after assembly of the tetrasaccharide or after formation of the final polymer. In addition, the fact that WcfO is unable to modify galactose linked to UDP, but does function with polyisoprenoid-linked AADGal-Gal, suggests that, at minimum, some aspect of the bactoprenyl AADGal structure is important for substrate recognition. While it was surprising that Glc was accepted as effectively as Gal by WcfO, the discrimination by WcfP in the next step demonstrates that the structure of that Glc-modified disaccharide was sufficiently different from the Gal. It would be interesting to see if a pyruvylated AADGal-PyrGlc is found in other materials in B. fragilis, or if this is an artifact of the in vitro analysis.
One of the only examples of the biochemical characterization of a pyruvyltransferase utilized a purified enzyme from the yeast Schizosaccharomyces pombe. This protein, Pvg1, which has no sequence similarity to WcfO, was assayed with galactose α- and β- linked to nitrophenol. Pvg1 was found to only transfer pyruvate to β-linked nitrophenyl-Gal, and did not utilize any α-linked sugar.21 However, in this work we found that this selectivity for just the Gal unit is not universal for 4,6-O-pyruvyltransferases. WcfO was unable to modify either of the nitrophenol-linked substrates, or UDP-Gal under conditions where it was fully functional with 2CNA-BPP-AADGal-Gal. The only record of a biochemically characterized bacterial 4,6-O-pyruvyltransferase is the PssM protein from the Gram-negative bacterium Rhizobium leguminosarum.23 This protein has been shown to incorporate pyruvate from PEP into membrane components associated with the organism. However, there were few specifics as to what these membrane components were. This work suggests that the activity of many pyruvyltransferases will be inaccessible without the assembly of the polyprenyl-linked substrates.
Because CPSA has garnered a great deal of recent attention, a route for the chemical synthesis of the repeat unit has been developed.36 This route required over 20 synthetic manipulations from relatively inexpensive starting materials, and provides an excellent method for the preparation of large quantities of the tetrasaccharide. While this achievement is astounding, the biologically active polymer requires, at minimum, eight linked tetrasaccharide units.2 The tetrasaccharide alone, when unlinked to an isoprenoid, is also of limited value for working with the flippase, polymerase and transport proteins involved in the formation of the final polymer and export. The work provided here established a single pot method for the production of the tetrasaccharide repeat unit, which is linked to an isoprenoid and ready for use with downstream proteins.
Traditional methods to reconstitute oligosaccharide biosynthesis in vitro have been heavily dependent on radiolabeled materials for detection of products.37-39 Once the functional role of each protein is assigned, one way to characterize by MS has been to cleave the oligosaccharide, label with anthranilamide, then purify and analyze by MALDI-MS.37 Even without the radiolabel, UDP-linked sugars are relatively expensive and enzymatic synthesis of materials rarely can achieve grams to work with. Because of the expense and time associated with traditional methods, systems that require relatively small amounts of substrate are ideal. In this work reactions associated with the easily quantifiable 2-nitrileaniline bactoprenyl analogues could be done with as little as 2.5 pmoles of isoprenoid. We found that characterization of these types of compounds by ESI-MS required in some cases as much as 4 nmoles, and in previous work we found that 200 nmoles would be required for NMR analysis.17, 18 While, our work is dependent on the original structural characterizations of CPSA,8 it does provide a simple, sensitive, non-radiolabel based method for preparing these materials and analyzing the specific roles of proteins involved in their formation. The major challenge now is to enhance MS signal, as sensitivity of this technique clearly decreased as the oligosaccharide repeat unit grew larger.
Lastly, this work describes the final step in CPSA repeat unit biosynthesis, which requires a functional Galp mutase, WcfM, to form Galf that is then incorporated by WcfN to form the tetrasaccharide. Previously, it has been shown that coupling glycosyltransferases with the mutase has been effective in overcoming the low yield associated with Galp to Galf turnover.40-43 In this report, a large excess of UDP-Galp was used, so it is not clear how much this coupling benefits the low turnover of the WcfM reaction. We have now established the complete in vitro reconstitution of the repeat unit of CPSA using key fluorescent isoprenoid probes. This is the first complete synthesis of a repeat unit linked to these isoprenoids.
Methods
General Procedures
All HPLC was performed on an Agilent 1100 HPLC system equipped with diode array and fluorescence detectors. HPLC analysis was performed with a flow rate of 1 mL/min unless stated otherwise on either a C8 or C18 Agilent Eclipse XDB, 5μm, 4.6 × 150 mm column. PglF, WcfS, WcfR, WcfP, WcfN, and 2CNA bactoprenyl phosphates were prepared as described previously.19, 28 Total protein concentration in membrane fractions were measured using a Bradford assay with BSA as a standard.44 WcfO, WcfP and WcfN reactions were left unquenched for more than an hour, unless stated otherwise. All reactions were performed at room temperature 22-24 °C. Concentrations of isoprenoid were measured spectroscopically using the 2-nitrileaniline (2CNA) extinction coefficient ε340=2700 M−1 cm−1. Uridine diphosphate-linked sugar concentrations were based on the extinction coefficient of uridine (ε260=10,000 M−1 cm−1). Each reaction and HPLC analysis was performed three times with each repeat on separate days. Results were similar for multiple batches of prepared proteins.
WcfO and WcfM cloning and expression
Polymerase chain reaction (PCR) amplification of wcfO and wcfM was performed using B. fragilis genomic DNA (ATCC 25285) and the following primers: wcfO Forward: GCATGGAGGATCCATGAGGAAGATATTATTAACATATGGAGATATTAAAACC wcfO Reverse: GCATGGACTCGAGTGACATCATAAATTTATTACATATATTAATTAATC wcfM Forward: GCATGGAGGATCCATGAAAAAAAAATATGACTATCTAATTGTCGGAGCCGG wcfM Reverse: GCATGGACTCGAGTAAGTCACTATTTATAACTTTTTCCACAATCACATTC
The wcfO and wcfM PCR products were digested with BamHI and XhoI. Each was ligated into a pET-24a vector digested with the same restriction enzymes. Chemically competent E. coli DH5α cells were transformed with the ligated vector and kanamycin resistant colonies were selected. Plasmids were isolated from the kanamycin resistant colonies and sequenced to confirm introduction of each gene (Eurofins-Operon). Chemically competent E. coli BL-21(DE3) RIL cells (Agilent) were then transformed and positive colonies were selected by Kanamycin resistance.
WcfO expression and isolation
Cells containing WcfO encoding plasmids were cultured in 5 mL of Luria Broth (LB) overnight at 37° C then 1 mL was used to inoculate 0.5 L of LB for protein expression. Cells were grown to an OD600 of 0.6 and the temperature was decreased to 26.5° C for 30 min. Isopropylthiogalactoside (IPTG) was added at a final concentration of 1 mM. Cells were allowed to incubate with shaking for an additional three hours, then were harvested by centrifugation at 4° C for 20 min at 5000 relative centrifugal force (RCF), the supernatant was removed and cells were stored at −80° C for later use. Protein expressing cells were thawed in 20 mL of 50 mM Tris-HCl (pH=8.0) containing 200 mM NaCl and 20 mM imidazole then sonicated on ice for 3 minutes (total) with a pulse of one second on and one second off. Unbroken cells were removed by centrifugation at 5000 RCF for 15 minute at 4° C. The supernatant containing membrane and cytosolic components were separated at 150,000 RCF for one hour at 4° C. The pelleted membrane components were homogenized into 2 mL of 50 mM Tris-HCl (pH=8.0) 200 mM NaCl then 10-50 μL aliquots were stored at −80° C. The presence of the overexpressed protein was confirmed by SDS-PAGE and Western Blot analysis with an anti-His primary antibody to detect the vector encoded tag.
Isolation of WcfO from membrane fraction
Membrane fraction (1 mL) containing WcfO was homogenized in 20 mL of Tris-HCl (pH=8.0) supplemented with 1% Triton-X-100 (or 0.1% DDM) and 200 mM NaCl. The homogenized fraction was separated at 150,000 RCF for an hour at 4° C. The supernatant was mixed with 1 mL Ni-NTA Agarose (5 Prime) for one hour at 4° C then was poured through a column. The resin was washed with a solution of 50 mM Tris-HCl (pH=8.0), 50 mM imidazole, 200 mM NaCl (4 × 3 mL). After washing, 0.35 mL of elution buffer containing 50 mM Tris-HCl (pH=8.0), 500 mM imidazole, 200 mM NaCl was passed through the column. Another 1.75 mL of elution buffer was then passed through the column and collected five times to obtain purified protein. The purified protein concentration was determined to be 1.31 mg/mL using a Bradford assay. The presence of purified protein was confirmed by Western Blot analysis with an anti-His antibody, and purity was assessed by SDS-PAGE.
WcfM overexpression and isolation
Two 0.5 L batches of LB were inoculated with an overnight culture (1 mL each), and cells were grown at 37 °C with agitation till an OD600 of 0.6 was reached, upon which temperature was reduced to 26 °C. Protein expression was induced with 1 mM IPTG. Four hours after induction, the cells were harvested by centrifugation (5000 RCF, 10 min). The harvested cells were washed with 50 mM potassium phosphate buffer (pH 7.4), collected again by centrifugation (5000 RCF, 10 min), and pelleted cells were stored at −80 °C. Cells were resuspended in lysis buffer (50 mM potassium phosphate buffer, pH 7.4, 20 mM imidazole, 200 mM NaCl), lysed by sonication as above, and pelleted at 2800 RCF to remove cell debris. Membrane components were removed by centrifugation at 180000 RCF for one hour and the remaining supernatant was mixed 1 mL Ni-NTA resin. The column was washed with a solution of 50 mM Tris-HCl (pH=8.0), 50 mM imidazole, 200 mM NaCl (4 × 3 mL), then dialyzed into 50 mM phosphate buffer (pH=7.4) 200 mM NaCl. The purified mutase was stable to storage at −80° C in potassium phosphate buffer containing 15% glycerol. Purity and confirmation of protein identity was assessed by SDS-PAGE and anti-His Western Blot analysis.
WcfO polyisoprenoid-linked substrate assay
In the typical WcfO assay a solution was prepared containing 5 μM 2CN-B(4Z)PP-AADGal-Gal or 2CN-B(4Z)PP-AADGal-Glc, 10 mM phosphoenolpyruvate, 25 mM Bicine, 10 mM MgCl2, 1% Triton-X-100, 10% DMSO, and 10 μg/mL purified WcfO, and incubated for at least one hour. HPLC was performed on a reverse phase C18 column using isocratic 35% 1-propanol with 65% 100 mM ammonium bicarbonate. Injection sizes ranged from 1-4 μL. Starting material and product was detected by fluorescence at excitation 340 nm and emission 390 nm.
WcfO assay with α and β nitrophenyl sugars
A solution was prepared containing 1 mM α-p-nitrophenyl galactopyranoside or β-o-nitrophenyl galactopyranoside, 5 mM phosphoenolpyruvate, 25 mM Bicine, 10 mM MgCl2, 1% Triton-X-100, and 10 μg/mL purified WcfO, which was then analyzed by HPLC. HPLC analysis was performed on the same C18 column as above using a gradient method in which over 30 minutes 12 % 1-propanol 88 % 100 mM ammonium bicarbonate was raised to 100 % 1-propanol. Sugar was detected at an absorbance of 405 nm.
UDP-Gal WcfO assay
Assays were prepared with 10 μg/mL of WcfO containing membrane fraction or purified WcfO in 100 μL containing 25 mM Tris, 10 mM MgCl2, 1% Triton-X-100, 300 μM UDP-Gal, 10 mM phosphoenolpyruvate. The mixtures were incubated at room temperature for 4 hours and analyzed by capillary electrophoresis (CE) for product formation. After the incubation, 8 μL of the reaction components were loaded on the capillary by pressure injection and separated at 15 kV for 25 minutes through a 48.0 μm inner diameter capillary at a length of 40.2 cm with 25 mM sodium tetraborate running buffer (pH=9.26). The starting materials as well as any potential products covalently linked to UDP were detected at 260 nm. NAD+ was added as an internal standard to normalize electrophoretic mobility between analyses allowing us to account for electrophoretic drift from injection to injection.
WcfO kinetic assay
Reaction mixtures of 500 μL containing 0.45 μM 2AA-B(6Z)PP-AAD-Gal-Gal or 2AA-B(6Z)PP-AAD-Gal-Glc and varying concentrations of PEP from 0.1 mM to 5 mM were incubated with 9.5 nM WcfO. Aliquots were removed at time intervals of 15 seconds over 1.5 minutes, and quenched with 1-propanol. The quenched aliquots were then analyzed by HPLC with fluorescence detection at excitation 350 nm and emission 450 nm. The column used was the same C18 described above, and the mobile phase was 50 % 1-propanol in 100 mM ammonium bicarbonate. The product turnover was calculated based on peak integrals and the initial rate was determined. Initial rates were fit to the Michaelis-Menten equation to determine apparent kinetic constants appkcatPEP and appKmPEP. Fits and error were based all data from seven PEP concentrations performed three times.
WcfP and WcfN assays
Assays for the second glycosyltransferase were prepared with 1 μL of WcfP containing membrane fraction (0.93 mg/mL total protein concentration) or WcfN (0.72 mg/mL total protein concentration) in a 10 μL solution containing 5 μM 2CN-B(4Z)PP-AADGal-PyrGal or 2.1 μM 2CN-B(4Z)PP-AADGal-PyrGlc, 1% Triton, 10 mM MgCl2, 10% DMSO, 25 mM Bicine (pH=8.3), and 3.5 mM UDP-GalNAc or UDP-GlcNAc. After 1 hour the reaction was analyzed by HPLC at 32 % 1-propanol, 68 % 100 mM ammonium bicarbonate with a C8 column.
WcfN assays
Assays for the final glycosyltransferase were prepared with 1 μL of WcfN membrane fraction (0.72 mg/mL total protein concentration) in a 10 μL solution containing 5 μM 2CN-B(4Z)PP-AADGal-PyrGal-GalNAc, 1 % Triton, 10 mM MnCl2, 10 % DMSO, 25 mM Bicine (pH=8.3) and 2.5 mM UDP Galf. After 30 minutes the reaction was analyzed by HPLC using conditions identical to the WcfP assays.
WcfM assays
WcfM assays were performed in 100 mM potassium phosphate buffer (pH 7.4) with 20 mM sodium dithionite. 3 μM WcfM (based on the extinction coefficient of WcfM) was incubated with 3.5 mM UDP-Galp in the buffer mix. After 10 minutes the reaction was analyzed on a reverse phase C18 column. The mobile phase used was 1.5% acetonitrile in 50 mM triethylammonium acetate buffer (pH 6.8), with a flow rate of 0.5 ml/min.33 Product formation was monitored at an absorbance of 260 nm.
WcfM/WcfN coupled assay
In these assays 1 μL of membrane fraction of WcfN (0.72 mg/mL final total protein centration) was incubated in a 10 μL solution with 5 μM 2CN-B(4Z)PP-AADGal-PyrGal-GalNAc, 1 % Triton, 10 mM MnCl2, 10 % DMSO, 25 mM Bicine (pH=8.3), 20 mM sodium dithionite, 3 μM WcfM and 2.5 mM UDP Galp. Product was analyzed by HPLC as described for WcfN alone.
Single pot sequential reaction
An HPLC sample was prepared as described below and was allowed to incubate for one hour before HPLC injection of 5 μL 0.5-1 hour after each enzyme addition. The reaction mixture started in 200 μL of 25 mM Bicine (pH=8.3), 1 % Triton-X-100, 10 mM MgCl2, 10 mM MnCl2, 5 μM of 2CN-B(4Z)P, 500 μM UDP-AADGal, 0.48 mg/mL total protein WcfS membrane fraction. Next 3 μL of WcfQ membrane fraction (stock concentration 6.9 mg/mL) and 2 μL of UDP-Gal (50 mM) were added, then HPLC analyzed. Next, 1 μL of PEP (1M) and 5 μL of purified WcfO (stock concentration 1.31 mg/mL) were added. After HPLC analysis, 2.6 μL of UDP-GalNAc (115 mM) and 2 μL of WcfP (stock concentration 9.3 mg/mL) were added. Finally, after HPLC analysis of the tetrasaccharide product 2.8 μL of UDP-Galf (105 mM) was added along with 2 μL of WcfN (stock concentration 7.2 mg/mL).
Characterization of products using MS
The products of WcfO, WcfP and WcfN were prepared using 3.5 mM sodium cholate in the place of Triton X-100, then were isolated by HPLC. Typical reactions contained 10-100 nmoles of 2CNA-B(4Z)P starting material. Concentrations of WcfO PyrGal and PyrGlc products for MS analysis in HPLC fractions were 15.9 μM and 12.3 μM. WcfP product in the HPLC fraction was 13.1 μM. WcfN product obtained after concentrating the sample was 42 μM. MS was performed on 10-100 μL of each sample on a Thermo VELOS Pro Dual-Pressure Linear Ion Trap using ESI introduced into the capillary with a 0.200 mL/min flow rate of acetonitrile. Negative mode was used with a capillary temperature of 300 °C and a spray voltage of 4.00 kV.
2CNA-B(4Z)PP-AADGal-PyrGal
Formula: C54H81N4O17P2− Expected mass: 1119.5 obtained 1119.7
2CNA-B(4Z)PP-AADGal-PyrGlc
Formula: C54H81N4O17P2− Expected mass: 1119.5 obtained 1119.6
2CNA-B(4Z)PP-AADGal-PyrGal-GalNAc
Formula: C62H94N5O22P2− Expected mass: 1322.6 obtained 1322.8
2CNA-B(4Z)PP-AADGal-PyrGal-GalNAc-Galf
Formula: C68H104N5O27P2− Expected mass: 1484.6 obtained 1484.1
Supplementary Material
ACKNOWLEDGMENT
We thank T. Lowary for providing UDP-galactofuranose. We thank V.Amabo, C. Martinez and J. Hazel for help preparing the nitrileaniline and anthranilamide analogues. We also thank R. Jew for editing and critique of this manuscript.
Funding Sources
This work was supported by a National Institutes of Health AREA grant R15GM100402 (JMT), R15GM114773 (JMT) and NSF instrumentation grant 1337873 (UNC-Charlotte Chemistry).
Footnotes
Supporting Information. Sequence analysis, WcfM protein analysis gels, and WcfN data with disaccharide, are provided in supporting information. The Supporting Information is available free of charge on the ACS Publications website
Author Contributions
The manuscript was written through contributions of all authors. All authors have given approval to the final version of the manuscript.
REFERENCES
- [1].Avci FY, Kasper DL. How bacterial carbohydrates influence the adaptive immune system. Annu Rev Immunol. 2010;28:107–130. doi: 10.1146/annurev-immunol-030409-101159. [DOI] [PubMed] [Google Scholar]
- [2].Mazmanian SK, Kasper DL. The love-hate relationship between bacterial polysaccharides and the host immune system. Nat Rev Immunol. 2006;6:849–858. doi: 10.1038/nri1956. [DOI] [PubMed] [Google Scholar]
- [3].Ochoa-Reparaz J, Wang Y, Miecarz DW, Begum-Haque S, Kasper DL, Kasper LH. Oral Treatment with Purified Polysaccharide A (PSA) of Human Commensal Bacteroides fragilis Protects Against Experimental CNS Demyelinating Disease. Neurology. 2010;74:A562–A562. doi: 10.1038/mi.2010.29. [DOI] [PubMed] [Google Scholar]
- [4].Ochoa-Reparaz J, Mielcarz DW, Wang Y, Begum-Haque S, Dasgupta S, Kasper DL, Kasper LH. A polysaccharide from the human commensal Bacteroides fragilis protects against CNS demyelinating disease. Mucosal Immunology. 2010;3:487–495. doi: 10.1038/mi.2010.29. [DOI] [PubMed] [Google Scholar]
- [5].Chung HC, Kasper DL. Microbiota-stimulated immune mechanisms to maintain gut homeostasis. Current Opinion in Immunology. 2010;22:455–460. doi: 10.1016/j.coi.2010.06.008. [DOI] [PubMed] [Google Scholar]
- [6].Trabbic KR, De Silva RA, Andreana PR. Elucidating Structural Features of an Entirely Carbohydrate Cancer Vaccine Construct Employing Circular Dichroism and Fluorescent Labeling. Medchemcomm. 2014;5:1143–1149. doi: 10.1039/C4MD00038B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].De Silva RA, Wang Q, Chidley T, Appulage DK, Andreana PR. Immunological response from an entirely carbohydrate antigen: design of synthetic vaccines based on Tn-PS A1 conjugates. J Am Chem Soc. 2009;131:9622–9623. doi: 10.1021/ja902607a. [DOI] [PubMed] [Google Scholar]
- [8].Baumann H, Tzianabos AO, Brisson JR, Kasper DL, Jennings HJ. Structural elucidation of two capsular polysaccharides from one strain of Bacteroides fragilis using high-resolution NMR spectroscopy. Biochemistry. 1992;31:4081–4089. doi: 10.1021/bi00131a026. [DOI] [PubMed] [Google Scholar]
- [9].Tzianabos AO, Pantosti A, Baumann H, Brisson JR, Jennings HJ, Kasper DL. The capsular polysaccharide of Bacteroides fragilis comprises two ionically linked polysaccharides. J Biol Chem. 1992;267:18230–18235. [PubMed] [Google Scholar]
- [10].Cobb BA, Kasper DL. Zwitterionic capsular polysaccharides: the new MHCII-dependent antigens. Cell Microbiol. 2005;7:1398–1403. doi: 10.1111/j.1462-5822.2005.00591.x. [DOI] [PubMed] [Google Scholar]
- [11].Cobb BA, Wang O, Tzianabos AO, Kasper DL. Polysaccharide processing and presentation by the MHCII pathway. Cell. 2004;117:677–687. doi: 10.016/j.cell.2004.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Duan J, Kasper DL. Regulation of T cells by gut commensal microbiota. Curr Opin Rheumatol. 2011;23:372–376. doi: 10.1097/BOR.0b013e3283476d3e. [DOI] [PubMed] [Google Scholar]
- [13].Coyne MJ, Tzianabos AO, Mallory BC, Carey VJ, Kasper DL, Comstock LE. Polysaccharide biosynthesis locus required for virulence of Bacteroides fragilis. Infect Immun. 2001;69:4342–4350. doi: 10.1128/IAI.69.7.4342-4350.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [14].Whitfield C. Biosynthesis and assembly of capsular polysaccharides in Escherichia coli. Annual Review of Biochemistry. 2006;75:39–68. doi: 10.1146/annurev.biochem.75.103004.142545. [DOI] [PubMed] [Google Scholar]
- [15].Woodward R, Yi W, Li L, Zhao G, Eguchi H, Sridhar PR, Guo H, Song JK, Motari E, Cai L, Kelleher P, Liu X, Han W, Zhang W, Ding Y, Li M, Wang PG. In vitro bacterial polysaccharide biosynthesis: defining the functions of Wzy and Wzz. Nat Chem Biol. 2010;6:418–423. doi: 10.1038/nchembio.351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Guo H, Yi W, Song JK, Wang PG. Current Understanding on Biosynthesis of Microbial Polysaccharides. Current Topics in Medicinal Chemistry. 2008;8:11. doi: 10.2174/156802608783378873. [DOI] [PubMed] [Google Scholar]
- [17].Lujan DK, Stanziale JA, Mostafavi AZ, Sharma S, Troutman JM. Chemoenzymatic synthesis of an isoprenoid phosphate tool for the analysis of complex bacterial oligosaccharide biosynthesis. Carbohydr Res. 2012;359:44–53. doi: 10.1016/j.carres.2012.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [18].Mostafavi AZ, Lujan DK, Erickson KM, Martinez CD, Troutman JM. Fluorescent probes for investigation of isoprenoid configuration and size discrimination by bactoprenol-utilizing enzymes. Bioorg Med Chem. 2013;21:5428–5435. doi: 10.1016/j.bmc.2013.06.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [19].Troutman JM, Sharma S, Erickson KM, Martinez CD. Functional identification of a galactosyltransferase critical to Bacteroides fragilis Capsular Polysaccharide A biosynthesis. Carbohydr Res. 2014;395:19–28. doi: 10.1016/j.carres.2014.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Mostafavi AZ, Troutman JM. Biosynthetic assembly of the Bacteroides fragilis capsular polysaccharide A precursor bactoprenyl diphosphate-linked acetamido-4-amino-6-deoxygalactopyranose. Biochemistry. 2013;52:1939–1949. doi: 10.1021/bi400126w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Yoritsune K, Matsuzawa T, Ohashi T, Takegawa K. The fission yeast Pvg1p has galactose-specific pyruvyltransferase activity. FEBS Lett. 2013;587:917–921. doi: 10.1016/j.febslet.2013.02.016. [DOI] [PubMed] [Google Scholar]
- [22].Marzocca MP, Harding NE, Petroni EA, Cleary JM, Ielpi L. Location and cloning of the ketal pyruvate transferase gene of Xanthomonas campestris. J Bacteriol. 1991;173:7519–7524. doi: 10.1128/jb.173.23.7519-7524.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Ivashina TV, Ksenzenko VN. Exopolysaccharide Biosynthesis in Rhizobium leguminosarum: From Genes to Functions. 2012.
- [24].Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic Local Alignment Search Tool. Journal of Molecular Biology. 1990;215:7. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- [25].Ivashina TV, Fedorova EE, Ashina NP, Kalinchuk NA, Druzhinina TN, Shashkov AS, Shibaev VN, Ksenzenko VN. Mutation in the pssM gene encoding ketal pyruvate transferase leads to disruption of Rhizobium leguminosarum bv. viciae-Pisum sativum symbiosis. J Appl Microbiol. 2010;109:731–742. doi: 10.1111/j.1365-2672.2010.04702.x. [DOI] [PubMed] [Google Scholar]
- [26].Lam JS, Kneidinger B, Larocque S, Brisson JR, Cadotte N. Biosynthesis of 2-acetamido-2,6-dideoxy-L-hexoses in bacteria follows a pattern distinct from those of the pathways of 6-deoxy-L-hexoses. Biochemical Journal. 2003;371:989–995. doi: 10.1042/BJ20030099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Olivier NB, Imperiali B. Crystal structure and catalytic mechanism of PglD from Campylobacter jejuni. J Biol Chem. 2008;283:27937–27946. doi: 10.1074/jbc.M801207200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Troutman JM, Erickson KM, Scott PM, Hazel JM, Martinez CD, Dodbele S. Tuning the production of variable length, fluorescent polyisoprenoids using surfactant-controlled enzymatic synthesis. Biochemistry. 2015;54:2817–2827. doi: 10.1021/acs.biochem.5b00310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Guerin ME, Kordulakova J, Schaeffer F, Svetlikova Z, Buschiazzo A, Giganti D, Gicquel B, Mikusova K, Jackson M, Alzari PM. Molecular recognition and interfacial catalysis by the essential phosphatidylinositol mannosyltransferase PimA from mycobacteria. J Biol Chem. 2007;282:20705–20714. doi: 10.1074/jbc.M702087200. [DOI] [PubMed] [Google Scholar]
- [30].Jain MK, Yuan W, Gelb MH. Competitive inhibition of phospholipase A2 in vesicles. Biochemistry. 1989;28:4135–4139. doi: 10.1021/bi00436a002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Poulin MB, Lowary TL. Methods to study the biosynthesis of bacterial furanosides. Methods Enzymol. 2010;478:389–411. doi: 10.1016/S0076-6879(10)78019-8. [DOI] [PubMed] [Google Scholar]
- [32].Nassau PM, Martin SL, Brown RE, Weston A, Monsey D, McNeil MR, Duncan K. Galactofuranose Biosynthesis in Escherichia coli K-12: Identification and Cloning of UDP-Galactopyranose Mutase. Journal of Bacteriology. 1996;178:6. doi: 10.1128/jb.178.4.1047-1052.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Zhang Q. a. L., H. Studies of UDP-Galactopyranose Mutase from Escherichia coli: An Unusual Role of Reduced FAD in Its Catalysis. Journal of American Chemical Society. 2000;122:6. [Google Scholar]
- [34].Richards MR, Lowary TL. Chemistry and biology of galactofuranose-containing polysaccharides. Chembiochem. 2009;10:1920–1938. doi: 10.1002/cbic.200900208. [DOI] [PubMed] [Google Scholar]
- [35].Soltero-Higgin M, Carlson EE, Gruber TD, Kiessling LL. A unique catalytic mechanism for UDP-galactopyranose mutase. Nat Struct Mol Biol. 2004;11:539–543. doi: 10.1038/nsmb772. [DOI] [PubMed] [Google Scholar]
- [36].Pragani R, Seeberger PH. Total Synthesis of the Bacteroides fragilis Zwitterionic Polysaccharide A1 Repeating Unit. Journal of American Chemical Society. 2011;133:102–107. doi: 10.1021/ja1087375. [DOI] [PubMed] [Google Scholar]
- [37].Glover KJ, Weerapana E, Imperiali B. In vitro assembly of the undecaprenylpyrophosphate-linked heptasaccharide for prokaryotic N-linked glycosylation. Proc Natl Acad Sci U S A. 2005;102:14255–14259. doi: 10.1073/pnas.0507311102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [38].Han W, Cai L, Wu B, Li L, Xiao Z, Cheng J, Wang PG. The wciN gene encodes an alpha-1,3-galactosyltransferase involved in the biosynthesis of the capsule repeating unit of Streptococcus pneumoniae serotype 6B. Biochemistry. 2012;51:5804–5810. doi: 10.1021/bi300640b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Brown S, Zhang YH, Walker S. A revised pathway proposed for Staphylococcus aureus wall teichoic acid biosynthesis based on in vitro reconstitution of the intracellular steps. Chem Biol. 2008;15:12–21. doi: 10.1016/j.chembiol.2007.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Lee R, Monsey D, Weston A, Duncan K, Rithner C, McNeil M. Enzymatic Synthesis of UDP-Galactofuranose and an Assay for UDP-Galactopyranose Mutase Based on High-Performance Liquid Chromatography. Analytical Biochemistry. 1996;242:7. doi: 10.1006/abio.1996.0419. [DOI] [PubMed] [Google Scholar]
- [41].Guan S, Clarke AJ, Whitfield C. Functional analysis of the galactosyltransferases required for biosynthesis of D-galactan I, a component of the lipopolysaccharide O1 antigen of Klebsiella pneumoniae. J Bacteriol. 2001;183:3318–3327. doi: 10.1128/JB.183.11.3318-3327.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [42].Koplin R, Brisson JR, Whitfield C. UDP-galactofuranose Precursor Required for Formation of the Lipopolysaccharide O Antigen of Klebsiella pneumoniae Serotype O1 Is Synthesized by the Product of the rfbDKPO1 Gene. Journal of Biological Chemistry. 1997;272:8. doi: 10.1074/jbc.272.7.4121. [DOI] [PubMed] [Google Scholar]
- [43].Poulin MB, Nothaft H, Hug I, Feldman MF, Szymanski CM, Lowary TL. Characterization of a bifunctional pyranose-furanose mutase from Campylobacter jejuni 11168. J Biol Chem. 2010;285:493–501. doi: 10.1074/jbc.M109.072157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [44].Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1006/abio.1976.9999. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.