

Abstract
Di(2-ethylhexyl) phthalate (DEHP) is a ubiquitous environmental pollutant used worldwide as a plasticizer and solvent in many formulations. Based on available toxicological data, it has been classified as toxic for reproduction and as an endocrine disruptor. Despite this, ecotoxicological studies in aquatic wildlife organisms are still scarce. In the present work, the toxic molecular alterations caused by DEHP in aquatic larvae of the midge Chironomus riparius have been studied, by analyzing the transcriptional activity of genes related to some vital cellular pathways, such as the ribosomal machinery (rpL4, rpL13), the cell stress response (hsc70, hsp70, hsp40, hsp27), the ecdysone hormone pathway (EcR), the energy metabolism (GAPDH), and detoxication processes (CYP4G). Environmentally relevant concentrations (10−3 to 105 μg/L) and exposure conditions (24 to 96 h) have been tested, as well as the toxic effects after DEHP withdrawal. Although the compound caused no mortality, significant changes were detected in almost all the studied biomarkers: e.g. strong repression of hsp70; general inhibition of EcR; GAPDH activity loss in long exposures; among others. Our data show a general transcriptional downregulation that could be associated with an adaptive response to cell damage. Besides, the activity of the compound as an ecdysone antagonist and its delayed effects over almost all the biomarkers analyzed are described as novel toxic targets in insects.
Introduction
Phthalates or phthalic acid esters (PAEs) are a family of man-made chemicals widely used since 1930s as plasticizers in the manufacture and processing of plastic materials, and which also appear in the composition of hundreds of consumer products, such as paints, detergents, adhesives, solvents, lubricants, insecticides, cosmetics and personal care products, among others [1,2]. World production of PAEs has grown rapidly in recent decades and, to date, 6–8 million tons of these chemicals are consumed worldwide each year [3]. Among all of them, di(2-ethylhexyl) phthalate (DEHP) has been the most commonly used, with an approximate annual consumption at present of 1–2 million tons [4,5].
Given that PAEs are not chemically bound to the polymeric matrix, they can gradually enter the environment by losses during their manufacturing, storage, use, and disposal [6,7], and can leach, migrate or evaporate into indoor air and atmosphere, soil, water, and a variety of materials such as foodstuff or medical devices, among others [1,8–10]. These characteristics, along with their physico-chemical properties, have turned PAEs into ubiquitous environmental pollutants that represent serious risks to human and environmental health.
Based on its toxicological profile and along with other phthalates, DEHP is blacklisted as a priority substance by the European Union [11,12], and several international regulatory agencies [3,8,13]. This chemical has also been included in different regulations concerning water quality, in which agencies have established the highest concentrations in water that are not expected to involve a significant risk to the majority of species in a specific environment, or to people [12,14,15]. Moreover, the presence of DEHP or other PAEs in different consumer products (e.g. toys, foodstuff) has been banned or restricted in recent years [16–18], and the presence of DEHP in drinking water has been severely limited [19–21].
Because of the large variety of toxic effects described to date, DEHP is classified as toxic for reproduction [22,23], Endocrine Disrupting Chemical (EDC) [24], and possibly carcinogenic to humans [5]. The environmental fate and toxicokinetics of DEHP has been reviewed extensively [1,3,8,25]. In its main degradation pathway, DEHP hydrolyzes to mono-ethylhexyl phthalate (MEHP) and follows subsequent glucuronid conjugation, but the formation rate and fate of MEHP in the environment is not known [26]. Differences in detoxication capabilities among aquatic species lead to variable bioaccumulation rates, although it is known that invertebrates are less able to break down DEHP [27]. MEHP reprotoxic effects have been reported in studies on mammals, but there are no other data on ecotoxicological properties of MEHP available [26]. Despite the increase in recent years in the amount of data available on the environmental toxicity of DEHP, research on its ecotoxicological effects in aquatic invertebrates and benthic wildlife organisms is still limited.
Since water constitutes the main vehicle for the dispersion of DEHP, aquatic ecosystems are especially sensitive to the presence of the compound. Although the compound in water tends to bind to sediments and suspended particles, a small amount persists dissolved in the water column [27]. Sediments integrate time-space effects of surface water pollution and represent a serious hazard to benthic and pelagic communities, so the ability of the compound to accumulate in such material makes it essential the evaluation of its toxic effects on species in that specific environment [26]. In this regard, among all freshwater benthic invertebrates, chironomid larvae constitute one of the most ubiquitous and abundant, and represent an important link in the food chain, capable of incorporating pollutants accumulated in sediments in which they grow and feed on. In fact, Chironomus midges are internationally used as model organisms in standardized environmental toxicity protocols [28–33], and have been selected as suitable organisms for research on the capacity of xenobiotics to cause endocrine disruption [34].
Complementary to other scientific works with more classical toxicity endpoints (e.g. survival, growth, immobilization, life-cycle, etc.), the use of molecular targets for ecotoxicity assessments (e.g. gene or enzyme activity) has increase in recent years in Chironomus riparius, as they have demonstrated to be effective biomarkers for the early detection of cellular stress responses and chemical toxicity, and constitute an important approach to achieve time and cost-effective tests for larger-scale evaluations. As an example, in the last few years different Chironomus species have served to assess transcriptional alterations caused by exposure to phthalates and other xenobiotics, biocides, metals, and nanoparticles, among other environmental stressors (e.g. [35–40] and references therein). It has been described the modulation of genes encoding ribosomal proteins, heat-shock proteins, hemoglobins, the ecdysone receptor, the estrogen-related receptor, alcohol dehydrogenase, calponin, and serine-type endopeptidase (e.g. [41–45] and references therein). Although it has been shown that expression levels of some of these molecular biomarkers are physiologically modulated throughout the development [42], available data demonstrate that gene expression is more sensitive to toxicant exposure than life cycle endpoints, underlining that the transcriptional changes can be harnessed to diagnose the exposure and effects of environmental chemicals in ecotoxicity testing and environmental risk assessments [46–48].
To determine accurate responsive genes which could be used as reliable biomarkers in the ecotoxicological risk assessment of DEHP, our objective in the present study was to analyze its molecular effects on aquatic larvae of the model organism C. riparius. We selected environmentally relevant concentrations and different exposure scenarios and evaluated the transcriptional activity of genes related to several crucial cell systems: the ribosomal machinery (rpL4, rpL13); the cell stress response (hsc70, hsp70, hsp40, hsp27); the ecdysone hormone pathway (EcR); the energy metabolism (GAPDH); and detoxication processes (CYP4G). We also assessed the enzyme activity of glutathione S-transferase (GST).
Materials and methods
Test animals
The experimental animals were aquatic larvae from the non-biting midge Chironomus riparius. Larvae used were reared under standard laboratory conditions, according to toxicity testing guidelines [32,33,49]. They were grown from egg masses in polyethylene tanks (500 mL) with aqueous culture medium (0.5 mM CaCl2, 1 mM NaCl, 1 mM MgSO4, 0.1 mM NaHCO3, 0.025 mM KH2PO4, 0.01 mM FeCl3) supplemented with nettle leaves, commercial fish food, and cellulose tissue. Cultures were maintained under constant aeration at 20°C and standard light-dark periods 16:8. Experiments were carried out using exclusively fourth instar larvae, and the larval stage was determined based on the size of head capsule [49].
Exposure conditions and survival tests
Solutions of di(2-ethylhexyl) phthalate (DEHP, CAS No. 117-81-7) (Sigma-Aldrich, USA) were dissolved in analytical grade ethanol to provide a stock concentration of 107 μg/L. The test solutions were constructed in culture medium at 0.01% ethanol. This was the final percentage of ethanol present in the solvent controls used in the experiment and preliminary tests demonstrated that induced no effects on the organisms in any of the selected endpoints. The nominal concentrations of DEHP ranged from 10−3 to 105 μg/L, and included both higher and lower levels than those described in the literature for drinking water resources and aquatic ecosystems [1,3,8,19,20,26].
Larval survival was studied in all DEHP concentrations for 24-h exposures, and at the four lowest doses (10−3 to 1 μg/L) for longer experiments. The delayed toxicity (24+24 h), consisting of 24-h exposure followed by DEHP withdrawal and additional 24 hours in fresh medium, was also evaluated. Groups of 20 larvae were selected randomly and exposed to aqueous solutions of DEHP, without sediment. Larvae were not fed during the experiments, and survival rates were calculated after 24, 48, 72, or 96 h. For each experimental condition, four independent experiments were performed and groups of five surviving larvae were randomly selected, stored at -80°C, and used for RNA or protein extraction.
RNA and protein isolation
TRIzol Reagent (Invitrogen, Germany) was used to extract total RNA from frozen larvae, following the manufacturer's instructions. Samples were treated with RNase-free DNase (Roche, Germany) for 90 min, and an organic extraction with phenol-chloroform was completed. Following precipitation using isopropyl alcohol (0.5 v/v) and washing with 70% ethanol, RNA was resuspended in DEPC water, quantified by absorption spectroscopy (BioPhotometer, Eppendorf, Germany), and stored at -80°C.
Total protein content was obtained after homogenization in 0.5 mL of Tris–EDTA buffer (40 mM Tris, 1 mM EDTA, pH 7.8) with 7x complete EDTA-Free protease inhibitor (Roche) with a subsequent centrifugation [42], and was quantified with BCA Protein Assay Reagent (Thermo Fisher Scientific, USA), according to the manufacturer's instructions. Protein samples were stored at -20°C.
Reverse Transcription Polymerase Chain Reaction (RT- PCR)
Gene expression analyses were carried out only at low DEHP doses (10−3 to 1 μg/L). Semi-quantitative RT-PCR was used to evaluate the mRNA expression profile of selected genes. Reverse transcription was performed with 1 μg of the isolated RNA. Oligo(dT) primer (Invitrogen) was used with the M-MLV enzyme (Invitrogen) following the manufacturer’s instructions. Suites of primers were selected to specifically target selected genes (Table 1). PCR was performed in a MiniOpticon Thermocycler (Bio-Rad, USA), according to [42]. For all samples, the initial cDNA content and the amplification curves were adjusted in the PCR protocols, to prevent the ulterior saturation of DNA bands and subsequent quantification errors. The amplified PCR products were run in a 9% acrylamide gel at 60 V for 3 hours in 1x TGE buffer (40 mM Tris-Cl (pH 8.5), 200 mM glycine, and 2.5 mM EDTA), visualized after ethidium bromide staining and quantified with Chemigenius3 (Syngene, USA), using GeneSnap 6.05 and GeneTools 3.06 software. Actin and 26S were used as reference genes to normalize the fluorescence of bands, and a second normalization was carried out to calculate the expression levels of DEHP-exposed samples in relation to control conditions. To minimize technical errors, three replicates were carried out for each experiment.
Table 1. Primers used for RT-PCR amplification of the genes studied in C. riparius.
Forward (F) and reverse (R) sequences, base pair (bp) length of the obtained fragments and origin of sequences are provided. References to the original published sequences can be found in [38,42].
| Gene | Description | DNA sequence (5′-3′) | Fragment size (bp) |
|---|---|---|---|
| actin | Actin protein | F GATGAAGATCCTCACCGAACG | 201 |
| R CGGAAACGTTCATTACCG | |||
| 26S | 26S ribosomal ribonucleic acid | F TTCGCGACCTCAACTCATGT | 220 |
| R CCGCATTCAAGCTGGACTTA | |||
| rpL4 | Ribosomal protein L4 | F AACGCTTCAGAGCTGGACGTGG | 149 |
| R ATTCATCTTGTGTACGCTCATTG | |||
| rpL13 | Ribosomal protein L13 | F AAGCTGCTTTCCCAAGAC | 351 |
| R TTGGCATAATTGGTCCAG | |||
| hsc70 | 70 kDa heat-shock cognate protein | F CGTGCTATGACTAAGGACAA | 239 |
| R GCTTCATTGACCATACGTTC | |||
| hsp70 | 70 kDa heat-shock protein | F CATGTGAACGAGCCAAGAGA | 274 |
| R TTGCCACAGAAGAAATCTTG | |||
| hsp40 | 40 kDa heat-shock protein | F TACGTGACGCTAGAGGAAA | 131 |
| R TTCCAGCCCGGCTT | |||
| hsp27 | 27 kDa heat-shock protein | F TCCTCGTGCTTGCC | 202 |
| R CAAGGATGGCTTCCA | |||
| EcR | Ecdysone receptor | F AGACGGTTATGAACAGCC | 240 |
| R CGAGCCATGCGCAACATC | |||
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase | F GGTATTTCATTGAATGATCACTTTG | 110 |
| R TAATCCTTGGATTGCATGTACTTG | |||
| CYP4G | Cytochrome p450 family 4 subfamily G | F GACATTGATGAGAATGATGTTGGTG | 340 |
| R TAAGTGGAACTGGTGGGTACAT |
Glutathione S-Transferase (GST) activity
Two representative DEHP concentrations were selected to evaluate the GST activity: 1 and 103 μg/L. For each experiment, five control larvae and five treated larvae were collected after DEHP treatments. Three replicates were performed and each sample was run in duplicate wells. Total protein was quantified with BCA Protein Assay Reagent (Thermo Fisher Scientific), and 25 μg of total protein were used for the enzyme assay. The GST activity was assessed spectrophotometrically with the kit GST (Sigma-Aldrich) in a JASCO V-530 spectrophotometer (JASCO, Japan), following [42].
Data analysis
SPSS® Statistics 22 software (IBM, USA) was used for statistical analysis. Normality (Shapiro-Wilk’s test) and homoscedasticity (Levene’s test) of data were checked. The normalized levels of transcripts and the GST enzyme function were analysed with ANOVA, followed by Games Howell’s or Bonferroni’s post hoc tests, when appropriate. The Kruskal-Wallis’ test was used when data were not homogeneous or normally distributed, and the differences between pairs were stablished using Mann-Whitney’s tests. Significant differences were stablished at p < 0.05.
Results
Larval survival
A wide range of concentrations (10−3 to 105 μg/L) was selected to assess larval mortality after 24-h exposure to DEHP. Within this range, levels found in natural exposure scenarios and also exposure concentrations evaluated in laboratory studies were tested [2,9,26,27,50,51]. For longer treatments (48 to 96 h), the four lowest concentrations were selected. Larval survival was not affected under any of the conditions analyzed. Although in 24-h exposures no significant mortality was observed, reaching survival rates very close to 100%, all experiments in these conditions showed a generalized loss of mobility and coloring of individuals in concentrations above 1 μg/L. These effects were not observed in longer exposures to lower concentrations.
Expression profile of ribosomal genes
The ability of DEHP to alter the ribosomal machinery was evaluated by means of transcriptional analysis of genes encoding ribosomal proteins L4 and L13. As shown in Fig 1A–1C, no effects were detected for the rpL4 gene in none of the studied conditions. However, the rpL13 gene was affected by the xenobiotic in all exposure scenarios, confirming the slight downregulation (not significant) detected in 24 and 72 h (Fig 1D and 1G) as statistically significant at 48 and 96 h (Fig 1E and 1H). Particularly relevant was the clear upregulation of rpL13 in the delayed toxicity experiments (24-h exposure to DEHP plus 24 h in fresh culture medium) (Fig 1F), showing the ability of DEHP to stimulate the transcriptional activity of this gene time after removal from culture medium.
Fig 1.

Effects of DEHP treatments on the relative expression of genes coding for ribosomal proteins: rpL4 (A-C), and rpL13 (D-H). Each bar is the mean ± SE obtained from three independent experiments, each with three replicates. Values are expressed as fold changes with respect to the control. X-axis values range from 10−3 to 1 μg/L. *Significant differences (p ≤ 0.05) as compared to control cultures.
Alteration of the heat-shock transcriptional response
The expression profile of constitutive (hsc70) and inducible (hsp70, hsp40, and hsp27) heat-shock genes were analyzed to evaluate the interactions of DEHP on the cellular stress response. Although no significant changes were detected for the heat-shock cognate gene hsc70, as shown in Fig 2A–2C, significant transcriptional alterations caused by DEHP were found for all inducible heat-sock genes. The compound induced a clear dose- and time-dependent downregulation of the hsp70 gene in exposures over 24 h, reaching values of 70% below control in 96-h treatments (Fig 2E, 2G and 2H). Earlier effects were detected in the genes coding for the 40 and 27 kDa proteins, with a repression of hsp40 at the highest dose (Fig 2I) and the overexpression of hsp27 at the lowest ones (Fig 2L). For these two genes, 48-h exposures led to increased transcriptional levels comparing with control samples (Fig 2J and 2M), unlike that observed for hsp70. Delayed toxicity tests revealed a strong overexpression of hsp70 (up to 5-fold) (Fig 2F), and a slight but steady transcriptional induction of hsp40 (Fig 2K).
Fig 2.

Effects of DEHP treatments on the relative expression of different heat-shock genes in C. riparius fourth instar larvae: hsc70 (A-C), hsp70 (D-H), hsp40 (I-K), and hsp27 (L-N). Each bar is the mean ± SE obtained from three independent experiments, each with three replicates. Values are expressed as fold changes with respect to the control. X-axis values range from 10−3 to 1 μg/L. *Significant differences (p ≤ 0.05) as compared to control cultures.
Modulation of the ecdysone receptor levels
The endocrine disrupting effect of DEHP and its ability to behave as a hormone antagonist was evaluated by means of the analysis of the transcriptional levels of the gene encoding the ecdysone receptor (EcR). Like the effects previously described for the hsp70 gene, DEHP exposures led to a time-dependent downregulation of EcR, with significant reductions of the transcriptional activity in each exposure time (48 to 96 h) for all the studied concentrations (between 30% and 70% below control, respectively), as shown in Fig 3A, 3B, 3D and 3E. However, contrary to that observed for the hsp70 gene, it is noteworthy that the withdrawal of the compound after 24-h treatments triggered a significant repression of the EcR gene 24 h later (Fig 3C), thus reaching transcriptional values about 40% below control in the delayed toxicity studies.
Fig 3. Effects of DEHP treatments on the relative expression of the ecdysone receptor gene (EcR) in C. riparius fourth instar larvae.

Each bar is the mean ± SE obtained from three independent experiments, each with three replicates. Values are expressed as fold changes with respect to the control. X-axis values range from 10−3 to 1 μg/L. *Significant differences (p ≤ 0.05) as compared to control cultures.
Detoxication and energy metabolism
The expression pattern of the CYP4G gene, as well as the enzyme activity of GST, provided us data related to the detoxication activities in C. riparius, whereas the transcriptional levels of GAPDH showed the possible toxic interaction between DEHP and the energy metabolism of exposed larvae. The xenobiotic clearly repressed the activity of CYP4G, even in the lowest concentrations at the shortest exposure time (24 h) (Fig 4A), and this decline remained significant for longer experiments (Fig 4B and 4D), with values close to 50% below control in 96-h treatments (Fig 4E). It is important to note that the repression produced by DEHP after 24 h was not only completely reverted after its removal but turned into a significant overexpression in the delayed toxicity assays (Fig 4C). The other detoxication pathway analyzed, represented by the GST enzyme (Fig 5), showed no effects in the 24-h acute exposures but was reduced after 48 h in the concentrations studied (1 and 103 μg/L), with identical effects in both 48-h continuous exposure experiments and delayed toxicity tests.
Fig 4.

Effects of DEHP treatments on the relative expression of genes involved in the detoxication metabolism in C. riparius fourth instar larvae: CYP4G (A-E), and GAPDH (F-J). Each bar is the mean ± SE obtained from three independent experiments, each with three replicates. Values are expressed as fold changes with respect to the control. X-axis values range from 10−3 to 1 μg/L. *Significant differences (p ≤ 0.05) as compared to control cultures.
Fig 5. Glutathione S-transferase activity in C. riparius fourth instar larvae after exposure to DEHP (1 and 103 μg/L) for 24, 48, and 24+24 hours.
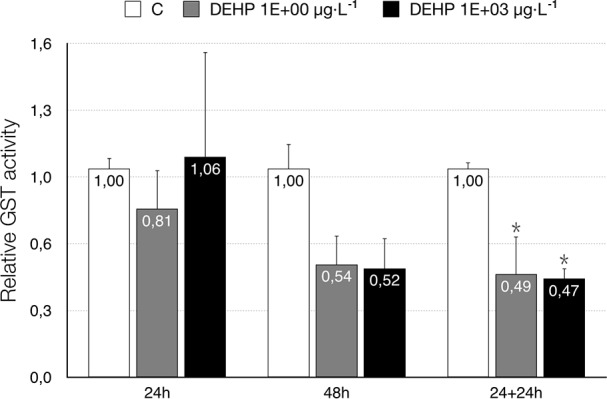
Each bar is the mean ± SE obtained from three independent experiments, each with three replicates. Values are expressed as fold changes with respect to the control. *Significant differences (p ≤ 0.05) as compared to control cultures.
Finally, DEHP had a time-dependent effect on the transcriptional activity of the GAPDH gene (Fig 4F–4J), leading to a significant reduction in 72-h treatments (Fig 4I) and an almost absolute repression in 96-h exposures (Fig 4J), with values of about 90% below the control. No delayed toxicity was detected for this gene (Fig 4H) in none of the studied conditions.
Discussion
DEHP is by far the most commonly used plasticizer. Its high production volume and its many applications worldwide make this chemical a ubiquitous pollutant, allowing the emergence of numerous possible exposure scenarios. The scientific community has frequently discussed the ability of the compound to produce adverse effects in humans. Its toxic and carcinogenic effects have been well described in experimental animals (especially rodents) [5,26,50] and plants [52]; however, its ability to induce similar effects in humans is still controversial [53]. In contrast to the abundance of studies focused ultimately on human health, there is much less information in scientific literature concerning the ecotoxicological profile of DEHP, particularly in invertebrates.
MEHP is the primary biodegradation product of DEHP and has proven to be toxic in studies with mammals. It is therefore reasonable to believe that MEHP could cause toxic effects also to other species like birds, fish, frogs etc. [26] However, very scarce information is available and, particularly in invertebrates, there are no previous works assessing the toxic effects of this DEHP metabolite. In this sense, further research is needed to evaluate the consequences of MEHP exposure on invertebrate species.
Survival analysis
Exposures to DEHP spanned from 24 to 96 hours and comprised a wide range of concentrations (10−3 to 105 μg/L), although the five highest concentrations were tested only for short-term (24h) survival experiments. This conditions include DEHP levels both above and (especially) below those described in other scientific works concerning drinking water resources or aquatic environments [8,9,26,51]. Interestingly, the survival rate of C. riparius larvae was not significantly affected in any of the conditions tested, which contrasts with the significant mortality caused by butyl benzyl phthalate (BBP) in experiments performed under identical conditions, where the LC50 was established at 2.7·104 μg/L [42]. These results are in accordance with the lack of significant mortalities detected for other invertebrate species (Hyalella azteca, Chironomus tentans, Lumbriculus variegatus) [54,55], even in long-term toxicity tests (Dahpnia magna, Chironomus plumosus, Palamonetes pugio, Mytilus edulis) [56–62].
Although no mortality was detected under any exposure condition, 24-h exposure to concentrations above 1 μg/L led to a generalized loss of mobility and coloring of individuals, proportionally to increased DEHP concentrations. Though not the subject of our study, larval mobility is considered a parameter equivalent to death rate in some ecotoxicological assays [63]. Similarly, larvae discoloration could be used as an early indicator of the toxicity of a compound [64], given that loss of red pigmentation, typically associated with hemoglobin, is a representative characteristic of larvae exposed to hypoxia-induced stress conditions [65]. The ability of DEHP to reduce the transcriptional activity of different hemoglobin genes has been previously described in C. tentans [44,45]. Although these genes were not included among the biomarkers selected for this work, the observed effects bring an interesting target for future studies regarding the toxicity of the xenobiotic.
Transcriptional changes on the expression of ribosomal protein genes
Genes encoding ribosomal proteins have evidenced to be sensitive biomarkers in C. riparius larvae exposed to different stressors ([38] and references therein). The stability shown by both rpL4 and rpL13 in 24-h exposures, confirms that the ribosomal machinery does not seem to be an early target of the toxic effect of low doses of the xenobiotic, in accordance with the results described by [66] for the ribosomal genes rpS3, rpS6, rpL11, rpL13, and rpL15 in a higher DEHP concentration. Our results demonstrate the capacity of DEHP to alter the activity of genes encoding ribosomal proteins by means of a slight but significant downregulation of rpL13, similarly to that observed in a previous work with the phthalate BBP [38]. It should be noted that removal of the compound resulted in a clear overexpression of the rpL13 gene, which could indicate that the organism increases its ribosomal activity (protein synthesis) trying to compensate for the damage induced previously by the xenobiotic, as described previously under other stressful conditions (e.g. [67]).
Alterations on the cellular stress response
The 70 kDa gene family includes both constitutive (hsc70) and inducible (hsp70) members, which share many common structural characteristics but present quite different expression patterns. While the constitutive forms are expressed at stable and relatively high levels under any condition, inducible heat-shock genes present a very low transcriptional profile under normal conditions, but their expression and subsequent translation of their products increase rapidly in response to a plethora of stress signals [68,69]. Our results showed no effects in hsc70 or hsp70 transcriptional levels after 24-h exposure to DEHP concentrations up to 1 μg/L. A previous work in C. tentans [45] found a clear induction of both genes after 24 h, at concentrations from 5·102 μg/L. This was partially consistent with another research in C. riparius [41] where DEHP led to the upregulation of hsp70 at concentrations between 103 and 105 μg/L, although hsc70 remained unaltered. In our study, longer exposures to the xenobiotic induced time and dose-dependent changes in the transcriptional profile of hsp70, with a strong and significant downregulation for all DEHP concentrations at 72–96 hours (up to 70% below control). It has been described that downregulation of hsp70 levels leads to increased sensitivity towards apoptosis-inducing agents, induces differentiation and cell death in cancer cells [70], and diminishes cell survival [71], although the strong repression detected in our study (together with those detected for rpL13, EcR, CYP4G, and GAPDH) could be the result of a general protective cell response consistent in maintaining a general low transcriptional profile in the presence of a sustained damage [72]. It is important to note the absence of changes in the activity of hsc70 in our experimental conditions, which reinforces the idea of its constitutive presence and confirms the toxic role of DEHP in the specific transcriptional alterations observed for the other genes.
The HSP40 protein is important for protein folding/unfolding, translation, translocation, and degradation, as it stimulates the ATP-ase activity of the HSP70 proteins [73]. In addition, the HSP40/HSP70 chaperone complex controls specific processes at distinct locations within the cell (e.g. cell cycle, cell differentiation, or apoptosis), including the progression of certain pathologies (e.g. oncogenesis, viral infections) [70]. Thus, the slight repression of hsp40 gene after 24-h exposure could compromise the effective course of HSP70 role, while in 48-h treatments the gene was slightly overexpressed, therefore promoting this chaperone activity. Our results clearly contrast with previous data [74] in which short-term exposures to three concentrations of DEHP (1, 10, and 30 μg/L) induced significant increases (up to 4-fold) in the expression of hsp40 and hsp90.
The HSP27 protein belongs to the small molecular weight heat shock proteins (sHSPs) family, and plays two major roles in response to stressful stimuli: (1) preserves the regular functioning of cells through remodelling and stabilization of the cytoskeleton, as well as facilitating the proper refolding or removal of defective proteins; and (2) prevents apoptosis by interfering with caspase activation in both mitochondrial dependent and independent pathways, and also lowering the levels of reactive oxygen species [75]. The chaperone activity of HSP27 prevents the formation of aggregates of denatured or improperly folded proteins [76], which can serve as a pro-apoptotic signal [77]. Such misfolded forms are processed by the endoplasmic reticulum (ER), which is responsible for the structural maturation of proteins by means of two different mechanisms that try to avoid ER stress: the unfolded protein response (UPR), which increases the folding capacity; and the ER-associated degradation (ERAD), which leads to protein removal [78]. It has been recently described that DEHP can trigger the ER stress response [79], leading to an adaptive response (inhibition of cell proliferation, cell cycle delay) rather than a pro-apoptotic one [80]. Previous works in C. riparius have detected a variety of hsp27 transcriptional responses under temperature or xenobiotic-induced stress ([42,81] and references therein). In the present study, the hsp27 gene tended to a significant overexpression in treatments from 24 to 48 hours, that could counteract the apoptotic signals derived from hsp70 downregulation.
Thereby, taken together, our results could suggest that DEHP is somehow altering protein folding and thus activating an adaptive response that slows down metabolism, trying to buy enough time to correct errors before progressing with normal cell cycle. Additionally, in vertebrates the HSPs are involved in stabilizing and activating the steroid hormone receptor [82], so if similar functions are performed in arthropods, changes in the transcriptional activity of these genes, particularly hsp70, could result in the alteration of the ecdysone-mediated hormone pathway by affecting the hormone receptor [83].
Effects over the ecdysone hormone pathway
Phthalates have been identified as EDCs in humans and also in mammalian models [84], causing adverse effects on reproduction and development. Although it has been described that EcR transcriptional peaks occur naturally during larval development [85,86], different studies have demonstrated the ability of some xenobiotics to modulate the expression levels of the receptor, working as ecdysone agonists/antagonists and ultimately influencing the development of the organism (e.g. [42,87,88]). Moreover, although endocrine systems in invertebrates differ drastically from vertebrates, it is worth mentioning that ecdysteroid hormones in insects belong to the family of steroid hormones, and that the ecdysone receptor belongs to the superfamily of nuclear hormone receptors that includes estrogens, androgens, thyroid hormone, retinoic acid and glucocorticoid receptors, among others [41]. Consequently, the ability of the EcR gene to detect endocrine disrupting activities in invertebrates may be a useful tool in an attempt to prevent environmental risks derived from EDCs, and also in predicting possible interferences with the vertebrate hormone system. In a previous work in C. riparius [41], 24-h exposure to DEHP did not alter the transcriptional levels of EcR in concentrations up to 104 μg/L, although a slight significant repression was detected at 105 μg/L. Results obtained in the present work demonstrate that longer exposures (48 to 96 hours) also lead to a significant downregulation of this gene, although at concentrations that are up to eight orders of magnitude below than those reflected in that previous study. Therefore, our data suggest that DEHP interferes with the endocrine function acting as an ecdysone antagonist, both at short exposures to high doses, or at long exposures to low doses. Data obtained for 48-h exposures are similar to those obtained with BBP [42], which could indicate that under certain conditions this antagonistic effect could be common to several members of the phthalate family.
Variations in energy and xenobiotic metabolism
Classic targets in ecotoxicity testing (e.g. survival, growth, reproduction rate) ultimately reflect changes in the energy metabolism of organisms [89]. Under this assumption, the transcriptional profile of the gene encoding GAPDH enzyme has been studied in the present work, as this protein plays a key role in energy production throughout glycolysis, although it is also involved in other functions at multiple subcellular compartments [90]. Precisely because of its many functions and its constant expression, GAPDH has been widely used as a reference gene in RT-PCR quantification, although it is known that it can be unstable under various experimental conditions (e.g. [91]). This fact makes necessary to check the stability of GAPDH under every experimental condition, thus avoiding errors caused by incorrect normalization. Our results show a stable behavior of the gene up to 48-h exposure to DEHP, but a significant drop in its transcriptional levels after 72 hours. It should be emphasized the almost total absence of GAPDH activity in 96-h experiments (up to 99% below control), which could be consistent with the hypothesis about the ability of the xenobiotic to slow down metabolism as an adaptive response to DEHP toxic effects.
In addition to their detoxication functions, P450 enzymes are also involved in the synthesis of ecdysteroids and juvenile hormones, with key roles in insect growth, development, and reproduction [92]. Thus, the response of some cytochromes seems to establish relations between exposure to some chemicals and the endocrine function [93]. Alterations in the transcriptional activity of different CYP genes have been described previously in Chironomus larvae exposed to a variety of xenobiotics ([42] and references therein). Our experiments let us detect a significant repression of the CYP4G gene in all the tested concentrations, even at the shortest DEHP exposures. This downregulation was concomitant with the significant low levels detected for EcR after 48 to 96-h exposure to DEHP, which could reflect the previously described association between the enzyme activity, the ecdysone synthesis and, by extension, the hormone receptor levels.
In many species, the expression levels of GST genes and the GST enzyme activity can be boosted significantly following exposure to xenobiotics, suggesting that they are involved in adaptive responses to chemical stress. Studies on the insect GSTs have focused primarily on their role in conferring resistance to insecticides [94]. As an example in chironomids, there have been described GST alterations in the gene activity [95] or the enzyme function [42,96,97] in response to the presence of xenobiotics. In this work, we have evaluated only two representative DEHP concentrations (1 and 103 μg/L) and it was found that this phthalate inhibits GST enzyme activity to about 50% at both doses in 48-h experiments, similarly to that observed for CYP4G transcriptional rates. This could suggest that DEHP restricts the ability of C. riparius larvae to maintain normal detoxication rates, decreasing the biotransformation capacity of both phase I and phase II metabolism enzymes.
Delayed toxicity studies
These experiments showed that the compound can cause adverse effects to the selected targets even after its withdrawal. Although some biomarkers that were not altered in 24-h exposures remained unchanged after 24 h of recovery (rpL4, hsc70, GAPDH), other genes showed a significant upregulation once the compound was removed from the culture medium (rpL13, hsp70, CYP4G). It could be hypothesized that the end of the DEHP injury allows the organism to recover normal metabolic rates, therefore being necessary to reinforce the ribosomal machinery and the protein folding/checking systems. Moreover, in the knowledge that high hsp70 levels lead to increased resistance against apoptosis [70], its upregulation could be compensating this protective role, implemented during the presence of the xenobiotic by high levels of the hsp27 gene. It is also possible that the significant rise in hsp40 levels is directly related to the increased activity of the HSP40/HSP70 chaperone complex. It should be highlighted that the removal of DEHP revealed surprising findings about the endocrine disrupting capacity of this phthalate. Although the transcriptional activity of the EcR gene was not altered in 24-h exposures to DEHP, its removal led to a significant repression of this gene when larvae were maintained an additional 24 h in fresh culture medium. This behavior differs from the rest of genes analyzed in this work and appears to confirm a specific effect of the xenobiotic on the endocrine system of C. riparius, acting as an antagonist of the insect steroid hormone.
Conclusions
The present study indicates that DEHP, the most commonly used plasticizer in the world, induces transcriptional alterations in exposed larvae of C. riparius. These toxic effects occur after short acute exposures, even at concentrations lower than those detected in environmental samples or permitted by different international regulations for drinking water. The use of biomarker genes constitutes a useful tool for the early detection of toxic effects, especially for those cellular pathways that have been evolutionary-conserved across different species. It is of particular interest to see how some of the toxic effects are detected long after the larvae have ceased to be exposed to the compound, which makes some of the DEHP toxic properties usually go unnoticed in the acute exposures of classical toxicity tests. Given the importance of this non-biting midge in the trophic chains of aquatic ecosystems, and the ability of the compound to bioaccumulate, it is advisable to continue investigating the different toxic effects of this xenobiotic, especially in natural populations. In addition, the appearance of effects at such low concentrations poses a potential worldwide risk to human and environmental health.
Supporting information
Acute exposures (24 to 96h) to a wide range of DEHP concentrations (1 ng/L to 0.1 g/L) caused no mortality in C. riparius larvae but led to a loss of mobility and coloring, and to a general decrease in the transcriptional activity of the studied genes.
(TIF)
Acknowledgments
This research was supported by Ministerio de Economía y Competitividad (Spain) contract grant number CTM2015-64913-R. The authors thank Dr. A. Hidalgo and Dr. M. Casanova (CNIC, Madrid, Spain) for their help with spectrophotometry equipment used in enzyme activity tests.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This research was supported by Ministerio de Economía y Competitividad (Spain) contract grant number CTM2015-64913-R (http://www.idi.mineco.gob.es).
References
- 1.Staples CA, Peterson DR, Parkerton TF, Adams WJ. The environmental fate of phthalate esters: A literature review. Chemosphere. 1997;35: 667–749. [Google Scholar]
- 2.Oehlmann J, Schulte-Oehlmann U, Kloas W, Jagnytsch O, Lutz I, Kusk KO, et al. A critical analysis of the biological impacts of plasticizers on wildlife. Philos Trans R Soc Lond B Biol Sci. 2009;364: 2047–62. 10.1098/rstb.2008.0242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Net S, Sempéré R, Delmont A, Paluselli A, Ouddane B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ Sci Technol. 2015;49: 4019–35. 10.1021/es505233b [DOI] [PubMed] [Google Scholar]
- 4.Halden RU. Plastics and health risks. Annu Rev Public Health. 2010;31: 179–94. 10.1146/annurev.publhealth.012809.103714 [DOI] [PubMed] [Google Scholar]
- 5.IARC. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans Volume 101 Some Chemicals Present in Industrial and Consumer Products, Food and Drinking-water. Lyon: International Agency for Research on Cancer; 2012. [PMC free article] [PubMed] [Google Scholar]
- 6.Staples CA. Phthalate Esters [Internet]. Staples CA, editor. Berlin, Heidelberg: Springer-Verlag; 2003. [Google Scholar]
- 7.Cadogan DF, Papez M, Poppe AC, Scheubel J. Assessment of the release, occurrence and possible effects of plasticizers in the environment. Prog Rubber Plast Technol. Brussels: European Council for Plasticisers and Intermediates; 1993;10: 1–19. [Google Scholar]
- 8.Gao D-W, Wen Z-D. Phthalate esters in the environment: A critical review of their occurrence, biodegradation, and removal during wastewater treatment processes. Sci Total Environ. 2016;541: 986–1001. 10.1016/j.scitotenv.2015.09.148 [DOI] [PubMed] [Google Scholar]
- 9.Clark K, Cousins IT, Mackay D, Yamada K. Observed Concentrations in the Environment The Handbook of Environmental Chemistry Vol3, Part Q. Berlin: Springer-Verlag; 2003. pp. 125–177. [Google Scholar]
- 10.Schettler T. Human exposure to phthalates via consumer products. Int J Androl. 2006;29: 134–9–5. [DOI] [PubMed] [Google Scholar]
- 11.ECHA. Member State Committee Support Document for Identification fo Bis(2-ethylhexyl)phthalate (DEHP) as a Substance of Very High Concern. Helsinki: European Chemicals Agency; 2008. [Google Scholar]
- 12.EU. Directive 2013/39/EU of the European Parliament and of the Council of 12 August 2013 amending Directives 2000/60/EC and 2008/105/EC as regards priority substances in the field of water policy. Luxembourg: Official Journal of the European Union, 24 August 2013, L226/1; 2013.
- 13.ATSDR. Support Document to the 2013 Priority List of Hazardous Substances [Internet]. Atlanta: U.S. Agency for Toxic Substances and Disease Registry; 2014. Available: http://www.atsdr.cdc.gov/SPL/resources/ATSDR_2013_SPL_Support_Document.pdf
- 14.EPA. Appendix A: Water Quality Standards Regulation. Water Quality Standards Handbook: Second Edition Washington: U.S. Environmental Protection Agency; 1994. [Google Scholar]
- 15.OECD. Surface Water Quality Regulation in EECCA Countries: Directions for Reform [Internet]. Paris: Organisation for Economic Co-operation and Development; 2008. Available: http://www.oecd.org/env/outreach/41832129.pdf [Google Scholar]
- 16.CPSC. Consumer Product Safety Improvement Act of 2008. Bethesda: Consumer Product Safety Commission; 2008. [Google Scholar]
- 17.EU. Directive 2005/84/EC of the European Parliament and of the Council of 14 December 2005 relating to restrictions on the marketing and use of certain dangerous substances and preparations (phthalates in toys and childcare articles). Luxembourg: Official Journal of the European Union, 27 December 2005, L344; 2005.
- 18.EU. Commission Directive 2007/19/EC of 30 March 2007 relating to plastic materials and articles intended to come into contact with food or foodstuffs. Luxembourg: Official Journal of the European Union, 31 March 2007, L91; 2007.
- 19.WHO. Di(2-ethylhexyl)phthalate in Drinking-water. Geneva: World Health Organization; 2003. [Google Scholar]
- 20.EPA. 2012 Edition of the Drinking Water Standards and Health Advisories. Washington: U.S. Environmental Protection Agency; 2012. [Google Scholar]
- 21.EU. Directive 2008/105/EC of the European Parliament and of the Council of 16 December 2008 on environmental quality standards in the field of water policy. Luxembourg: Official Journal of the European Union, 16 December 2008, L226/1; 2008.
- 22.EU. Regulation (EC) No 1272/2008 of the European Parliament and of the Council, of 16 December 2008, on classification, labelling and packaging of substances and mixtures, amending and repealing Directives 67/548/EEC and 1999/45/EC, and amending Regulation (E. Luxembourg: Official Journal of the European Union, 31 December 2008, L353; 2008.
- 23.EU. Commission Directive 2001/59/CE of 6 August 2001 relating to the classification, packaging and labelling of dangerous substances. Luxembourg: Official Journal of the European Union, 6 August 2001, L225/1; 2001.
- 24.EC. Towards the establishment of a priority list of substances for further evaluation of their role in endocrine disruption. Delft: European Commission DG ENV; 2000. [Google Scholar]
- 25.Koch HM, Preuss R, Angerer J. Di(2-ethylhexyl)phthalate (DEHP): human metabolism and internal exposure–an update and latest results. Int J Androl. 2006;29: 155–65–5. [DOI] [PubMed] [Google Scholar]
- 26.ECB. European Union Risk Assessment Report—bis(2-ethylhexyl)phthalate (DEHP). Luxembourg: European Chemicals Bureau; 2008. [Google Scholar]
- 27.OEHHA. Toxicological Profile for Di-(2-ethylhexyl) Phthalate (DEHP). Sacramento: California Environmental Protection Agency; 2009. [Google Scholar]
- 28.EPA. Ecological Effects Test Guidelines. OPPTS 850.1790 Chironomid Sediment Toxicity Test. Washington: U.S. Environmental Protection Agency; 1996.
- 29.EPA. Ecological Effects Test Guidelines. OPPTS 850.1735 Whole Sediment Acute Toxicity Invertebrates, Freshwater. Washington: U.S. Environmental Protection Agency; 1996.
- 30.OECD. Test No. 235: Chironomus sp., Acute Immobilisation Test. Paris: Organisation for Economic Co-operation and Development; 2011.
- 31.OECD. Test No. 233: Sediment-Water Chironomid Life-Cycle Toxicity Test Using Spiked Water or Spiked Sediment. Paris: Organisation for Economic Co-operation and Development; 2010.
- 32.OECD. Test No. 218: Sediment-Water Chironomid Toxicity Test Using Spiked Sediment. Paris: Organisation for Economic Co-operation and Development; 2004.
- 33.OECD. Test No. 219: Sediment-Water Chironomid Toxicity Test Using Spiked Water. Paris: Organisation for Economic Co-operation and Development; 2004.
- 34.OECD. Detailed review paper on aquatic arthropods in life cycle toxicity tests with an empashis on developmental, reproductive and endocrine disruptive effects OECD Series on Testing and Assessment. Number 55 Paris: Organisation for Economic Co-operation and Development; 2006. [Google Scholar]
- 35.Ozáez I, Morcillo G, Martínez-Guitarte J-L. Ultraviolet filters differentially impact the expression of key endocrine and stress genes in embryos and larvae of Chironomus riparius. Sci Total Environ. 2016;557–558: 240–247. 10.1016/j.scitotenv.2016.03.078 [DOI] [PubMed] [Google Scholar]
- 36.Aquilino M, Sánchez-Argüello P, Martínez-Guitarte J-L. Vinclozolin alters the expression of hormonal and stress genes in the midge Chironomus riparius. Aquat Toxicol. 2016;174: 179–87. 10.1016/j.aquatox.2016.03.001 [DOI] [PubMed] [Google Scholar]
- 37.Morales M, Planelló R, Martínez-Paz P, Herrero O, Cortés E, Martínez-Guitarte JL, et al. Characterization of Hsp70 gene in Chironomus riparius: expression in response to endocrine disrupting pollutants as a marker of ecotoxicological stress. Comp Biochem Physiol C Toxicol Pharmacol. Elsevier Inc.; 2011;153: 150–8. 10.1016/j.cbpc.2010.10.003 [DOI] [PubMed] [Google Scholar]
- 38.Herrero Ó, Planelló R, Morcillo G. The ribosome biogenesis pathway as an early target of benzyl butyl phthalate (BBP) toxicity in Chironomus riparius larvae. Chemosphere. 2016;144: 1874–84. 10.1016/j.chemosphere.2015.10.051 [DOI] [PubMed] [Google Scholar]
- 39.Lee S-W, Choi J. Multi-level ecotoxicity assay on the aquatic midge, Chironomus tentans (Diptera, Chironomidae) exposed to octachlorostyrene. Environ Toxicol Pharmacol. 2009;28: 269–74. 10.1016/j.etap.2009.05.004 [DOI] [PubMed] [Google Scholar]
- 40.Park K, Kwak I-S. The effect of temperature gradients on endocrine signaling and antioxidant gene expression during Chironomus riparius development. Sci Total Environ. 2014;470–471: 1003–11. 10.1016/j.scitotenv.2013.10.052 [DOI] [PubMed] [Google Scholar]
- 41.Planelló R, Herrero O, Martínez-Guitarte JL, Morcillo G. Comparative effects of butyl benzyl phthalate (BBP) and di(2-ethylhexyl) phthalate (DEHP) on the aquatic larvae of Chironomus riparius based on gene expression assays related to the endocrine system, the stress response and ribosomes. Aquat Toxicol. 2011;105: 62–70. 10.1016/j.aquatox.2011.05.011 [DOI] [PubMed] [Google Scholar]
- 42.Herrero Ó, Planelló R, Morcillo G. The plasticizer benzyl butyl phthalate (BBP) alters the ecdysone hormone pathway, the cellular response to stress, the energy metabolism, and several detoxication mechanisms in Chironomus riparius larvae. Chemosphere. 2015;128: 266–77. 10.1016/j.chemosphere.2015.01.059 [DOI] [PubMed] [Google Scholar]
- 43.Park K, Kwak I-S. Molecular effects of endocrine-disrupting chemicals on the Chironomus riparius estrogen-related receptor gene. Chemosphere. Elsevier Ltd; 2010;79: 934–41. 10.1016/j.chemosphere.2010.03.002 [DOI] [PubMed] [Google Scholar]
- 44.Anderson TD, Jin-Clark Y, Begum K, Starkey SR, Zhu KY. Gene expression profiling reveals decreased expression of two hemoglobin genes associated with increased consumption of oxygen in Chironomus tentans exposed to atrazine: a possible mechanism for adapting to oxygen deficiency. Aquat Toxicol. 2008;86: 148–56. 10.1016/j.aquatox.2007.10.015 [DOI] [PubMed] [Google Scholar]
- 45.Lee S-M, Lee S-B, Park C-H, Choi J. Expression of heat shock protein and hemoglobin genes in Chironomus tentans (Diptera, chironomidae) larvae exposed to various environmental pollutants: a potential biomarker of freshwater monitoring. Chemosphere. 2006;65: 1074–81. 10.1016/j.chemosphere.2006.02.042 [DOI] [PubMed] [Google Scholar]
- 46.Marinković M, de Leeuw WC, Ensink WA, de Jong M, Breit TM, Admiraal W, et al. Gene expression patterns and life cycle responses of toxicant-exposed chironomids. Environ Sci Technol. 2012;46: 12679–86. 10.1021/es3033617 [DOI] [PubMed] [Google Scholar]
- 47.Lee S-W, Park K, Hong J, Choi J. Ecotoxicological evaluation of octachlorostyrene in fourth instar larvae of Chironomus riparius (Diptera, Chironomidae). Environ Toxicol Chem. 2008;27: 1118–27. 10.1897/07-219.1 [DOI] [PubMed] [Google Scholar]
- 48.Herrero O, Planelló R, Gómez-Sande P, Aquilino M, Morcillo G. Evaluation of the toxic effects of phthalates on natural populations of Chironomus riparius (Diptera): implications for ecotoxicity studies. Rev Toxicol. 2014;31: 176–186. [Google Scholar]
- 49.EPA. Methods for Measuring the Toxicity and Bioaccumulation of Sediment-associated Contaminants with Freshwater Invertebrates. Second Edition Washington: U.S. Environmental Protection Agency; 2000. [Google Scholar]
- 50.NTP-CERHR. NTP-CERHR monograph on the potential human reproductive and developmental effects of di (2-ethylhexyl) phthalate (DEHP). [Internet]. Bethesda: U.S. National Institutes of Health; 2006. Available: http://www.ncbi.nlm.nih.gov/pubmed/19407857 [PubMed] [Google Scholar]
- 51.ATSDR. Toxicological Profile for Di(2-ethylhexyl)phthalate (DEHP). Atlanta: U.S. Agency for Toxic Substances and Disease Registry; 2002. [PubMed] [Google Scholar]
- 52.Herrero O, Pérez Martín JM, Fernández Freire P, Carvajal López L, Peropadre A, Hazen MJ. Toxicological evaluation of three contaminants of emerging concern by use of the Allium cepa test. Mutat Res. 2012;743: 20–4. 10.1016/j.mrgentox.2011.12.028 [DOI] [PubMed] [Google Scholar]
- 53.FDA. Safety Assessment of Di (2-ethylhexyl) phthalate (DEHP) Released from PVC Medical Devices. Rockville: U.S. Food and Drug Administration; 2001. [Google Scholar]
- 54.Call DJ, Markee TP, Geiger DL, Brooke LT, VandeVenter FA, Cox DA, et al. An assessment of the toxicity of phthalate esters to freshwater benthos. 1. Aqueous exposures. Environ Toxicol Chem. 2001;20: 1798–1804. [PubMed] [Google Scholar]
- 55.Call DJ, Cox DA, Geiger DL, Genisot KI, Markee TP, Brooke LT, et al. An assessment of the toxicity of phthalate esters to freshwater benthos. 2. Sediment exposures. Environ Toxicol Chem. 2001;20: 1805–1815. [PubMed] [Google Scholar]
- 56.Knowles CO, McKee MJ, Palawski DU. Chronic effects of DI-2-ethylhexyl phthalate on biochemical composition, survival and reproduction of Daphnia magna. Environ Toxicol Chem. 1987;6: 201–208. [Google Scholar]
- 57.Brown D, Thompson SE. Phthalates and the aquatic environment: Part I The effect of di-2-ethylhexyl phthalate (DEHP) and di-isodecyl phthalate (DIDP) on the reproduction of of Daphnia magna and observations on their bioconcentration. Chemosphere. 1982;11: 417–426. [Google Scholar]
- 58.Brown D, Thompson R. Phthalates and the aquatic environment: Part II The bioconcentration and depuration of di-2-ethylhexyl phthalate (DEHP) and di-isodecyl phthalate (DIDP) in mussels (Mytilus edulis). Chemosphere. 1982;11: 427–435. [Google Scholar]
- 59.Brown D, Croudace CP, Williams NJ, Shearing JM, Johnson PA. The effect of phthalate ester plasticisers tested as surfactant stabilised dispersions on the reproduction of the Daphnia magna. Chemosphere. 1998;36: 1367–1379. [Google Scholar]
- 60.Adams WJ, Heidolph BB. Short-cut chronic toxicity estimates using Daphnia magna In: Cardwell RD, Purdy R, Bahner RC, editors. Aquatic Toxicology and Hazard Assessment. Philadelphia: American Society for Testing and Materials; 1985. pp. 87–103. [Google Scholar]
- 61.Streufert JM, Jones JR, Sanders HO. Toxicity and biological effects of phthalate esters on midges (Chironomus plumosus). Trans Missouri Acad Sci. Commission of the European Communities; 1980;14: 33–40. [Google Scholar]
- 62.Laughlin RB, Neff JM, Hrung YC, Goodwin TC, Giam CS. The effects of three phthalate esters on the larval development of the grass shrimp Palaemonetes pugio (Holthuis). Water Air Soil Pollut. Kluwer Academic Publishers; 1978;9: 323–336. [Google Scholar]
- 63.OECD. 40 Years of Chemical Safety at the OECD: Quality and Efficiency. Paris: Organisation for Economic Co-operation and Development; 2011. [Google Scholar]
- 64.Majumdar TN, Gupta A. Acute and chronic toxicity of copper on aquatic insect Chironomus ramosus from Assam, India. J Environ Biol. 2012;33: 139–42. Available: http://www.ncbi.nlm.nih.gov/pubmed/23033657 [PubMed] [Google Scholar]
- 65.MacDonald MM, Warne AL, Stock NL, Mabury S a, Solomon KR, Sibley PK. Toxicity of perfluorooctane sulfonic acid and perfluorooctanoic acid to Chironomus tentans. Environ Toxicol Chem. 2004;23: 2116–23. [DOI] [PubMed] [Google Scholar]
- 66.Park K, Kwak I-S. Gene expression of ribosomal protein mRNA in Chironomus riparius: effects of endocrine disruptor chemicals and antibiotics. Comp Biochem Physiol C Toxicol Pharmacol. Elsevier Inc.; 2012;156: 113–20. 10.1016/j.cbpc.2012.05.002 [DOI] [PubMed] [Google Scholar]
- 67.Shi Y, Xu X, Zheng X, Lu Y. Responses of growth inhibition and antioxidant gene expression in earthworms (Eisenia fetida) exposed to tetrabromobisphenol A, hexabromocyclododecane and decabromodiphenyl ether. Comp Biochem Physiol C Toxicol Pharmacol. 2015;174–175: 32–8. 10.1016/j.cbpc.2015.06.005 [DOI] [PubMed] [Google Scholar]
- 68.Lindquist S, Craig EA. The heat-shock proteins. Annu Rev Genet. 1988;22: 631–77. 10.1146/annurev.ge.22.120188.003215 [DOI] [PubMed] [Google Scholar]
- 69.Bukau B, Horwich AL. The Hsp70 and Hsp60 chaperone machines. Cell. 1998;92: 351–66. Available: http://www.ncbi.nlm.nih.gov/pubmed/9476895 [DOI] [PubMed] [Google Scholar]
- 70.Mayer MP, Bukau B. Hsp70 chaperones: cellular functions and molecular mechanism. Cell Mol Life Sci. 2005;62: 670–84. 10.1007/s00018-004-4464-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kiang JG, Tsokos GC. Heat shock protein 70 kDa: molecular biology, biochemistry, and physiology. Pharmacol Ther. 1998;80: 183–201. Available: http://www.ncbi.nlm.nih.gov/pubmed/9839771 [DOI] [PubMed] [Google Scholar]
- 72.Peterson JSK, Bain LJ. Differential gene expression in anthracene-exposed mummichogs (Fundulus heteroclitus). Aquat Toxicol. 2004;66: 345–55. 10.1016/j.aquatox.2003.10.005 [DOI] [PubMed] [Google Scholar]
- 73.Qiu X-B, Shao Y-M, Miao S, Wang L. The diversity of the DnaJ/Hsp40 family, the crucial partners for Hsp70 chaperones. Cell Mol Life Sci. 2006;63: 2560–70. 10.1007/s00018-006-6192-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Park K, Kwak I-S. Characterization of heat shock protein 40 and 90 in Chironomus riparius larvae: effects of di(2-ethylhexyl) phthalate exposure on gene expressions and mouthpart deformities. Chemosphere. Elsevier Ltd; 2008;74: 89–95. 10.1016/j.chemosphere.2008.09.041 [DOI] [PubMed] [Google Scholar]
- 75.Concannon CG, Gorman AM, Samali A. On the role of Hsp27 in regulating apoptosis. Apoptosis. 2003;8: 61–70. Available: http://www.ncbi.nlm.nih.gov/pubmed/12510153 [DOI] [PubMed] [Google Scholar]
- 76.Bryantsev AL, Kurchashova SY, Golyshev SA, Polyakov VY, Wunderink HF, Kanon B, et al. Regulation of stress-induced intracellular sorting and chaperone function of Hsp27 (HspB1) in mammalian cells. Biochem J. 2007;407: 407–417. 10.1042/BJ20070195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Soldatenkov VA, Dritschilo A. Apoptosis of Ewing’s sarcoma cells is accompanied by accumulation of ubiquitinated proteins. Cancer Res. 1997;57: 3881–5. Available: http://www.ncbi.nlm.nih.gov/pubmed/9307264 [PubMed] [Google Scholar]
- 78.Bukau B, Weissman J, Horwich A. Molecular chaperones and protein quality control. Cell. 2006;125: 443–51. 10.1016/j.cell.2006.04.014 [DOI] [PubMed] [Google Scholar]
- 79.Sun X, Lin Y, Huang Q, Shi J, Qiu L, Kang M, et al. Di(2-ethylhexyl) phthalate-induced apoptosis in rat INS-1 cells is dependent on activation of endoplasmic reticulum stress and suppression of antioxidant protection. J Cell Mol Med. 2015;19: 581–94. 10.1111/jcmm.12409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Peropadre A, Fernández Freire P, Pérez Martín JM, Herrero Ó, Hazen MJ. Endoplasmic reticulum stress as a novel cellular response to di (2-ethylhexyl) phthalate exposure. Toxicol In Vitro. 2015;30: 281–287. 10.1016/j.tiv.2015.10.009 [DOI] [PubMed] [Google Scholar]
- 81.Martín-Folgar R, Martínez-Guitarte J-L. Cadmium alters the expression of small heat shock protein genes in the aquatic midge Chironomus riparius. Chemosphere. 2016;169: 485–492. 10.1016/j.chemosphere.2016.11.067 [DOI] [PubMed] [Google Scholar]
- 82.Gillesby BE, Zacharewski TR. Exoestrogens: Mechanisms of action and strategies for identification and assessment. Environ Toxicol Chem. 1998;17: 3–14. [Google Scholar]
- 83.Oehlmann J, Schulte-Oehlmann U. Endocrine disruption in invertebrates. Pure Appl Chem. 2003;75: 2207–2218. [Google Scholar]
- 84.Lyche JL, Gutleb AC, Bergman A, Eriksen GS, Murk AJ, Ropstad E, et al. Reproductive and developmental toxicity of phthalates. J Toxicol Environ Health B Crit Rev. 2009;12: 225–49. 10.1080/10937400903094091 [DOI] [PubMed] [Google Scholar]
- 85.Riddiford LM, Cherbas P, Truman JW. Ecdysone receptors and their biological actions. Vitam Horm. 2000;60: 1–73. [DOI] [PubMed] [Google Scholar]
- 86.Planelló R, Herrero Ó, Gómez-Sande P, Ozáez I, Cobo F, Servia MJ. Ecdysone-Related Biomarkers of Toxicity in the Model Organism Chironomus riparius: Stage and Sex-Dependent Variations in Gene Expression Profiles. Escriva H, editor. PLoS One. Public Library of Science; 2015;10: e0140239 10.1371/journal.pone.0140239 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Ozáez I, Martínez-Guitarte JL, Morcillo G. The UV filter benzophenone 3 (BP-3) activates hormonal genes mimicking the action of ecdysone and alters embryo development in the insect Chironomus riparius (Diptera). Environ Pollut. 2014;192: 19–26. 10.1016/j.envpol.2014.04.038 [DOI] [PubMed] [Google Scholar]
- 88.Nair PMG, Choi J. Modulation in the mRNA expression of ecdysone receptor gene in aquatic midge, Chironomus riparius upon exposure to nonylphenol and silver nanoparticles. Environ Toxicol Pharmacol. Elsevier B.V.; 2012;33: 98–106. 10.1016/j.etap.2011.09.006 [DOI] [PubMed] [Google Scholar]
- 89.Servia MJ, Péry ARR, Heydorff M, Garric J, Lagadic L. Effects of copper on energy metabolism and larval development in the midge Chironomus riparius. Ecotoxicology. 2006;15: 229–40. 10.1007/s10646-005-0054-0 [DOI] [PubMed] [Google Scholar]
- 90.Tristan C, Shahani N, Sedlak TW, Sawa A. The diverse functions of GAPDH: views from different subcellular compartments. Cell Signal. 2011;23: 317–23. 10.1016/j.cellsig.2010.08.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bustin SA. Why the need for qPCR publication guidelines?–The case for MIQE. Methods. 2010;50: 217–26. 10.1016/j.ymeth.2009.12.006 [DOI] [PubMed] [Google Scholar]
- 92.Feyereisen R. Insect P450 enzymes. Annu Rev Entomol. 1999;44: 507–33. 10.1146/annurev.ento.44.1.507 [DOI] [PubMed] [Google Scholar]
- 93.Le Goff G, Hilliou F, Siegfried BD, Boundy S, Wajnberg E, Sofer L, et al. Xenobiotic response in Drosophila melanogaster: sex dependence of P450 and GST gene induction. Insect Biochem Mol Biol. 2006;36: 674–82. 10.1016/j.ibmb.2006.05.009 [DOI] [PubMed] [Google Scholar]
- 94.Che-Mendoza A, Penilla RP, Rodríguez DA. Insecticide resistance and glutathione S-transferases in mosquitoes: A review. African J Biotechnol. 2009;8: 1386–97. Available: http://www.academicjournals.org/Ajb/abstracts/abs2009/20Apr/Che-Mendoza et al.htm [Google Scholar]
- 95.Nair PMG, Chung IM. Alteration in the expression of antioxidant and detoxification genes in Chironomus riparius exposed to zinc oxide nanoparticles. Comp Biochem Physiol B Biochem Mol Biol. 2015;190: 1–7. 10.1016/j.cbpb.2015.08.004 [DOI] [PubMed] [Google Scholar]
- 96.Lee S-B, Choi J. Multilevel evaluation of nonylphenol toxicity in fourth-instar larvae of Chironomus riparius (Diptera, Chironomidae). Environ Toxicol Chem. 2006;25: 3006–14. Available: http://www.ncbi.nlm.nih.gov/pubmed/17089725 [DOI] [PubMed] [Google Scholar]
- 97.Somparn A, Iwai CB, Noller B. Potential use of acetylcholinesterase, glutathione-S-transferase and metallothionein for assessment of contaminated sediment in tropical chironomid, Chironomus javanus. J Environ Biol. 2015;36: 1355–9. Available: http://www.ncbi.nlm.nih.gov/pubmed/26688973 [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Acute exposures (24 to 96h) to a wide range of DEHP concentrations (1 ng/L to 0.1 g/L) caused no mortality in C. riparius larvae but led to a loss of mobility and coloring, and to a general decrease in the transcriptional activity of the studied genes.
(TIF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.