

Abstract
AIM
To examine the effect of high doses of vitamin C (VitC) on ex vivo human platelets (PLTs).
METHODS
Platelet concentrates collected for therapeutic or prophylactic transfusions were exposed to: (1) normal saline (control); (2) 0.3 mmol/L VitC (Lo VitC); or (3) 3 mmol/L VitC (Hi VitC, final concentrations) and stored appropriately. The VitC additive was preservative-free buffered ascorbic acid in water, pH 5.5 to 7.0, adjusted with sodium bicarbonate and sodium hydroxide. The doses of VitC used here correspond to plasma VitC levels reported in recently completed clinical trials. Prior to supplementation, a baseline sample was collected for analysis. PLTs were sampled again on days 2, 5 and 8 and assayed for changes in PLT function by: Thromboelastography (TEG), for changes in viscoelastic properties; aggregometry, for PLT aggregation and adenosine triphosphate (ATP) secretion in response to collagen or adenosine diphosphate (ADP); and flow cytometry, for changes in expression of CD-31, CD41a, CD62p and CD63. In addition, PLT intracellular VitC content was measured using a fluorimetric assay for ascorbic acid and PLT poor plasma was used for plasma coagulation tests [prothrombin time (PT), partial thrombplastin time (PTT), functional fibrinogen] and Lipidomics analysis (UPLC ESI-MS/MS).
RESULTS
VitC supplementation significantly increased PLTs intracellular ascorbic acid levels from 1.2 mmol/L at baseline to 3.2 mmol/L (Lo VitC) and 15.7 mmol/L (Hi VitC, P < 0.05). VitC supplementation did not significantly change PT and PTT values, or functional fibrinogen levels over the 8 d exposure period (P > 0.05). PLT function assayed by TEG, aggregometry and flow cytometry was not significantly altered by Lo or Hi VitC for up to 5 d. However, PLTs exposed to 3 mmol/L VitC for 8 d demonstrated significantly increased R and K times by TEG and a decrease in the α-angle (P < 0.05). There was also a fall of 20 mm in maximum amplitude associated with the Hi VitC compared to both baseline and day 8 saline controls. Platelet aggregation studies, showed uniform declines in collagen and ADP-induced platelet aggregations over the 8-d study period in all three groups (P > 0.05). Collagen and ADP-induced ATP secretion was also not different between the three groups (P > 0.05). Finally, VitC at the higher dose (3 mmol/L) also induced the release of several eicosanoids including thromboxane B2 and prostaglandin E2, as well as products of arachidonic acid metabolism via the lipoxygenases pathway such as 11-/12-/15-hydroxyicosatetraenoic acid (P < 0.05).
CONCLUSION
Alterations in PLT function by exposure to 3 mmol/L VitC for 8 d suggest that caution should be exerted with prolonged use of intravenous high dose VitC.
Keywords: Platelet function, Thromboelastography, Flow cytometry, Platelet lipidomics, Vitamin C
Core tip: High dose intravenous vitamin C (VitC) is often used by Complementary and Alternate Medicine practitioners for a variety of ailments. Moreover, use of high dose VitC by mainstream physicians as an adjunct in the treatment of sepsis, sepsis induced acute lung injury, cancer and burns is on the rise. However, there is no information on the impact of these high doses VitC on normal platelet (PLT) function. Prolonged exposure of ex vivo PLTs to high doses of VitC altered some PLT functions as assessed by thromboelastography. However, short term exposure (< 8 d) or low dose exposure had almost no impact on PLT function.
INTRODUCTION
Platelets (PLTs) are central to physiologic processes involved in hemostasis and thrombosis[1]. While an immune function of PLTs has been described in the literature[2], recent studies point to an incompletely understood role for PLTs in a myriad of host immune responses. These studies point to altered PLT function in numerous disease states including inflammation, acute respiratory distress syndrome, atherosclerosis and cancer[3-5].
L-ascorbic acid is the reduced form of vitamin C (VitC). It is a water soluble molecule with strong antioxidant properties[6,7]. PLTs possess Na+-dependent VitC transporters (SVCT2) and this allows them to actively transport VitC intracellularly[8]. Although normal VitC concentrations in plasma are 50-80 μmol/L, PLTs can hold up to 4 mmol/L of intracellular VitC[9]. This is 50-80 fold higher than circulating VitC concentrations in plasma[10]. Studies have shown that VitC plays several roles in platelet functions, including reduction of reactive oxygen species[11], inhibition of expression of the pro-inflammatory CD40 ligand (CD40L)[12], inhibition of thromboxane B2 formation[13] and stimulation of prostaglandin E1 production[14]. This underscores the important role of VitC for normal platelet metabolic functions.
While VitC at normal physiological concentrations is critical for PLT function, there is virtually no information on the impact of high concentrations of VitC on PLT function. High dose intravenous VitC was predominantly used by Complementary and Alternate Medicine (CAM) practitioners. However, there has been a recent trend to use high dose intravenous VitC to treat many chronic, untreatable or intractable disease states. At the present time, high dose intravenous VitC is often used as an adjunct in the treatment of sepsis, sepsis induced acute lung injury, cancer, iron deficiency in hemodialysis patients and even in the burn protocol[15-19]. A few studies have reported that high dose intravenous VitC has complications in those with renal impairment or glucose 6 phosphate dehydrogenase deficiency. But, in general, its use appears relatively safe in multiple published randomized clinical trials. Yet, since the scale of such use is on the rise, it is vital that the safety of high dose VitC be examined in greater detail. To address this need, we examined the effect of exposing human PLTs to high doses of VitC. The doses used in this study were typically those reported in many of the recently completed randomized clinical trials[15-19]. We also used PLTs under ex vivo conditions for these studies. These PLTs were primarily collected for therapeutic or prophylactic transfusions and stored appropriately. We examined the effect of high doses VitC on a variety of PLT functions, both at rest and following activation, over an 8 d period.
MATERIALS AND METHODS
Platelet concentrate preparation
Platelet concentrates (PCs) were prepared by Virginia Blood Services (Richmond, VA) following standard operating procedures. Briefly, freshly collected, whole blood was centrifuged at low speed (soft spin 1500 × g) to separate platelet rich plasma (PRP). PRP was subjected to a second centrifugation (hard spin 5000 × g), then all but 50 mL of supernatant plasma was removed to concentrate the PLTs. The PLTs were re-suspended in residual plasma and stored with agitation at 22 °C-24 °C for 8 d at the Virginia Commonwealth University Transfusion Medicine Center.
Experimental design and study groups
PCs were treated with one of three additives: Normal saline (control); 0.3 mmol/L VitC (Lo VitC); or 3 mmol/L VitC (Hi VitC) as final concentrations. We used 6-10 PC’s per treatment arm. The VitC additive was preservative-free buffered ascorbic acid in water (Ascor L500, McGuff Pharmaceuticals, Santa Ana, CA), pH 5.5 to 7.0 adjusted with sodium bicarbonate and sodium hydroxide. Prior to supplementation, an initial baseline sample was collected at the blood supplier facility and transported to participating laboratories for analysis. PCs that passed standard screening tests were transported to the Virginia Commonwealth University Transfusion Medicine Center and sampled again on days 2, 5 and 8.
Sample processing
PLT samples were collected using sterile technique and processed. An initial PLT count was obtained and a portion of the sample was used to obtain platelet poor plasma (PPP) by centrifugation at 2000 × g for 10 min. The resultant PPP was then used to adjust the sample platelet concentration to 230-270 × 103/ μL (Adj. PRP).
Platelet pH and ascorbate analysis
An aliquot of the unadjusted PC was used for pH determination. For ascorbate determination, Adj. PRP (500 μL) was pelleted by centrifugation; washed with room temperature phosphate buffered saline; deproteinized in 100 μL of cold 20% trichloroacetic acid followed by addition of 100 μL of cold 0.2% dithiothreitol to prevent oxidation. Platelet lysates were vortexed and centrifuged at 10000 g for 10 min 4 °C. The supernatants were stored at -80 °C for batch analysis. Total ascorbate was assessed using a Tempol-OPDA based fluorescence end-point assay as previously described[20].
Plasma coagulation tests
Aliquots of PPP were assayed for prothrombin time (PT), activated partial thromboplastin time, and functional fibrinogen using the Stago STA Compact Coagulation Analyzer (Diagnostica Stago Inc., Parsippany, NJ) according to manufacturer’s instructions.
Measurement of platelet function
Viscoelastic properties measurement: The viscoelastic properties of PRP were measured in duplicate on a thromboelastography analyzer [thromboelastography (TEG) 5000, Haemonetics Corp., Braintree, maximum amplitude (MA)] using published methods[21]. Briefly, 30 μL of 0.2 mmol/L CaCl2 and 330 μL of PRP were loaded into the TEG cup sequentially and test parameters (i.e., R, K, α and MA) recorded.
Platelet aggregation and secretion: PLT aggregation in response to 2 μg/mL collagen or 10 μmol/L adenosine diphosphate (ADP) stimulation was measured by optical density using PRP. Simultaneously, the associated PLT adenosine triphosphate (ATP) secretion was measured via luminescence using Chrono-LumeTM reagent. Respective PPP aliquots of each sample were used as blanks. All runs were done in duplicate on a Chrono-log Series 500 aggregometer (Chrono-Log Corp., Havertown, PA) according to manufacturer’s instructions.
Platelet flow cytometry
Reagents: Human thrombin (T7009), Gly-Pro-Arg-Pro (GPRP) tetra-peptide inhibitor of fibrin polymerization, and serum bovine albumin were all obtained from Sigma-Aldrich (St. Louis, MO); phosphate-buffered saline and formalin from Beckman Coulter (Fullerton, CA); fluorescein isothiocynate conjugated CD41a (CD41a-FITC) and CD-31 (CD31-FITC), and phycoerythrin conjugated CD62p (CD62p-PE) and CD63 (CD63-PE) were obtained from BD Biosciences (San Jose, CA); ADP from Chrono-Log Corp (Havertown, PA).
Procedure: Adj. PRP was diluted (1:10) using sterile saline (0.9% NaCl). Eight tubes were prepared per sample; 1 unstained, 3 CD62p-PE (alone, thrombin 0.5 U/mL, or ADP 10 μmol/L), 3 CD63-PE (alone, thrombin 0.5 U/mL, or ADP 10 μmol/L), and 1 containing the corresponding isotypes-matched monoclonal antibodies mixture (negative control). CD41a-FITC and/or CD31-FITC were used to set the platelets gate for acquisition. GPRP (0.5 mmol/L) was added prior to thrombin activation to inhibit fibrin polymerization. All tubes were processed the same day on the Accuri C6 flow cytometer (BD Biosciences, San Jose, CA). The assay was performed under the following conditions: Fluidics: Medium; forward scatter threshold: 30000; and 20000 events were collected in a preset platelet gate using standard methods including CD41a and/or CD31 as global platelet markers. The collected data were analyzed using FlowJo version 7.6.5. (Ashland, OR). Results were expressed in mean fluorescence intensity units for CD41 and in percentages for other markers of activation.
Lipidomics analysis
Eicosanoids were analyzed as previously described[22-24]. Quantitative analysis of the lipids in the ethanolic extracts was carried out using UPLC ESI-MS/MS as described with minor modifications[25,26]. Briefly, to 200 μL of plasma, LCMS grade ethanol containing 10 ng of each internal standard was added (1 mL). The samples were mixed using a bath sonicator followed by incubation overnight at -20 °C for lipid extraction. Following incubation, the insoluble fraction was precipitated by centrifuging at 12000 g for 20 min and the supernatant was transferred into a new glass tube. The lipid extracts were then dried under vacuum and reconstituted in LCMS grade 50:50 EtOH:dH2O (100 μL) for eicosanoid quantitation via UPLC ESI-MS/MS analysis.
Statistical analysis
Statistical analysis was performed using SAS 9.3 and GraphPad Prism 6.0 (GraphPad Software, San Diego, CA, United States) by Bernard J Fisher, Division of Pulmonary Disease and Critical Care Medicine, Department of Internal Medicine, Virginia Commonwealth University, Richmond, Virginia and Bassem M. Mohammed, Department of Pharmacotherapy and Outcomes Science, Virginia Commonwealth University, Richmond, Virginia. Data are expressed as mean ± SE. Results were compared by one-way ANOVA and the post hoc Tukey test to identify specific differences between groups. Statistical significance was confirmed at a P value of < 0.05.
RESULTS
VitC exposure increased intracellular PLT VitC concentrations during storage
Freshly isolated PLTs contain high concentrations of intracellular VitC (1.23 ± 0.09 mmol/L). These concentrations are about 20 fold higher than the typical plasma levels of 50-80 μmol/L VitC. In freshly isolated PLTs on day 0, intracellular platelet VitC concentrations were not significantly different between the groups (Figure 1). By day 2, PLTs from VitC supplemented bags had significantly higher VitC levels (3.2 mmol/L for Lo VitC and 15.7 mmol/L for Hi VitC) compared to saline (1.2 mmol/L, P < 0.05). VitC content of PLTs observed at day 2 did not significantly change throughout the rest of the storage period in all three groups. This suggests that PLTs, when exposed to high concentrations of VitC, have the capacity to store VitC intracellularly at concentrations that are significantly higher than that observed at normal plasma levels.
Figure 1.
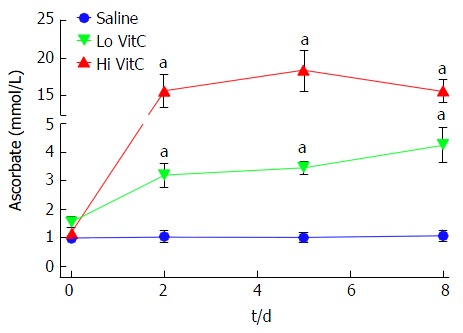
Vitamin C exposure increased intracellular platelet vitamin C concentrations during storage. PLTs supplemented with Lo/Hi VitC had significantly higher intracellular levels of VitC (3.2 mmol/L for Lo VitC and 15.7 mmol/L for Hi VitC) compared to saline controls (1 mmol/L), (n = 10/group, aP < 0.05). VitC: Vitamin C; PLTs: Platelets.
VitC exposure was associated with pH drop on day 8
Baseline pH was initially identical between the three groups. There was a slow, but comparable drop in pH in the three groups until day 5. However, in the PC exposed to Lo/Hi VitC supplementation, there was a further significant decrease in pH between day 5 and day 8 (Figure 2, P < 0.05).
Figure 2.
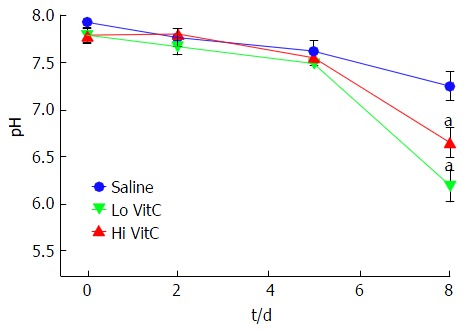
Vitamin C exposure was associated with pH drop on day 8. The addition of Lo/Hi VitC did not alter the pH of the platelet concentrate. Changes in pH were comparable throughout day 5 of storage. Both Lo/Hi VitC were associated with a further significant decrease in pH between day 5 and day 8 (n = 6/group, aP < 0.05). VitC: Vitamin C; PLTs: Platelets.
VitC exposure did not alter coagulation pathways in PLTs
VitC supplementation did not significantly change PT and PTT values which gradually increased in all three groups (Figure 3A and B). On similar lines, functional fibrinogen levels also did not differ between the groups over the 8 d and remained within a clinically relevant range (Figure 3C).
Figure 3.
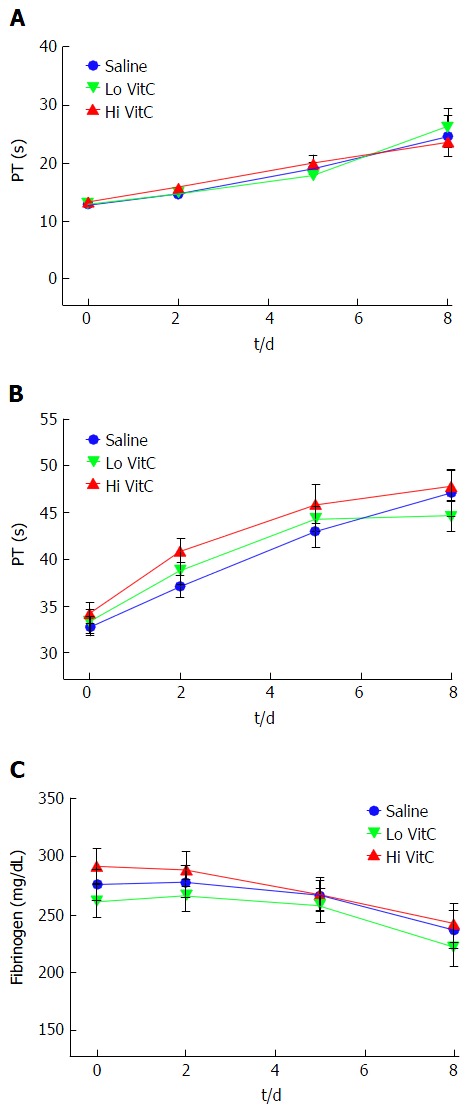
Vitamin C exposure did not alter coagulation pathways in platelets. PT, PTT and Fibrinogen were performed to detect major impacts on the intrinsic, extrinsic and common coagulation pathways. Using platelet poor plasma we observed no significant changes across the saline, Lo- and Hi VitC groups in the PT (A), PTT (B), and Fibrinogen (C) profiles over storage (n =10/group). VitC: Vitamin C; PT: Prothrombin time; PTT: Partial thrombplastin time.
High VitC exposure impacts PLT function on day 8
Over the first 5 d, the addition of Hi or Lo VitC had no deleterious impact on any of the TEG parameters compared to the saline control (Figure 4). However, on day 8 significant differences were associated with extended storage in Hi VitC. Specifically, R and K times were extended in the Hi VitC group when compared to saline group (Figure 4A and B, P < 0.05). In agreement with K-time data, a decrease in α-angle was observed in the Hi VitC group on day 8 (Figure 4C, P < 0.05). On day 8, there was a fall of 20 mm in MA associated with the Hi VitC compared to both baseline and day 8 saline controls (Figure 4D, P < 0.05).
Figure 4.
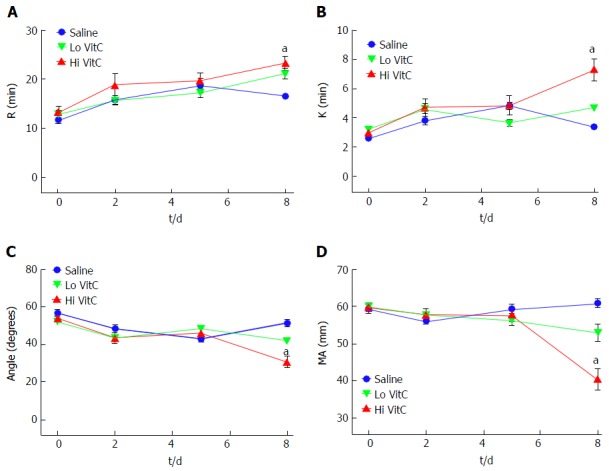
High vitamin C exposure impacts platelet function on day 8. No significant differences were observed between saline, Lo VitC and Hi VitC throughout the standard 5-d storage period in any of the TEG parameters analyzed (n = 10/group). On day-8, R- (A) and K- (B) times were extended in the Hi VitC group compared to saline (n = 10, aP < 0.05). Similar changes were observed for the α-angle (C) on day-8 (n = 10, aP < 0.05). The Hi VitC group was also associated with a significantly lower MA (D) on day-8 compared to the saline group (n = 10, aP < 0.05). VitC: Vitamin C; TEG: Thromboelastography.
Platelet aggregation studies, showed uniform declines in Collagen and ADP-induced platelet aggregations over the study period in all three groups (Figure 5A and B). In addition, Lo/Hi VitC addition did not alter Collagen and ADP-induced ATP secretion throughout the study (Figure 5C and D). Furthermore, flow cytometric analysis of CD62 and CD63 expression profiles, showed that VitC supplementation had no effect on basal CD62p and CD63 expression during storage (Figure 6A and D). Following ADP (Figure 6B) or thrombin (Figure 6C) stimulation, the CD62 expression showed a steady decrease that was not significantly different across the three groups (except for Hi VitC vs saline on day 8, P < 0.05). The expression profiles for CD63 differed depending on whether the PC were stimulated with ADP or thrombin (Figure 6E and F). However, the observed flow cytometric analysis changes were not significant across the three groups over 8 d.
Figure 5.
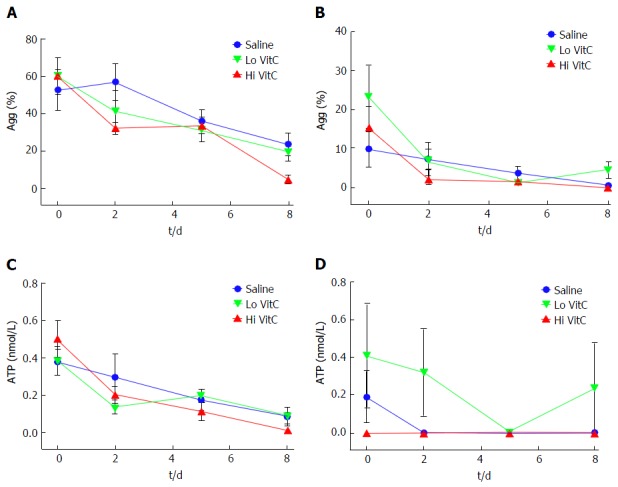
Vitamin C exposure did not affect agonist-stimulated aggregation or adenosine triphosphate secretion by platelets. Using adj. PRP aliquots, addition of Lo/Hi VitC did not alter Collagen-induced PLTs aggregation (A) and ATP secretion (C) as well as ADP-induced PLTs aggregation (B) and ATP secretion (D) when compared to saline controls (n = 10/group). VitC: Vitamin C; PLTs: Platelets; ADP: Adenosine diphosphate; PRP: Platelet rich plasma; ATP: Adenosine triphosphate.
Figure 6.
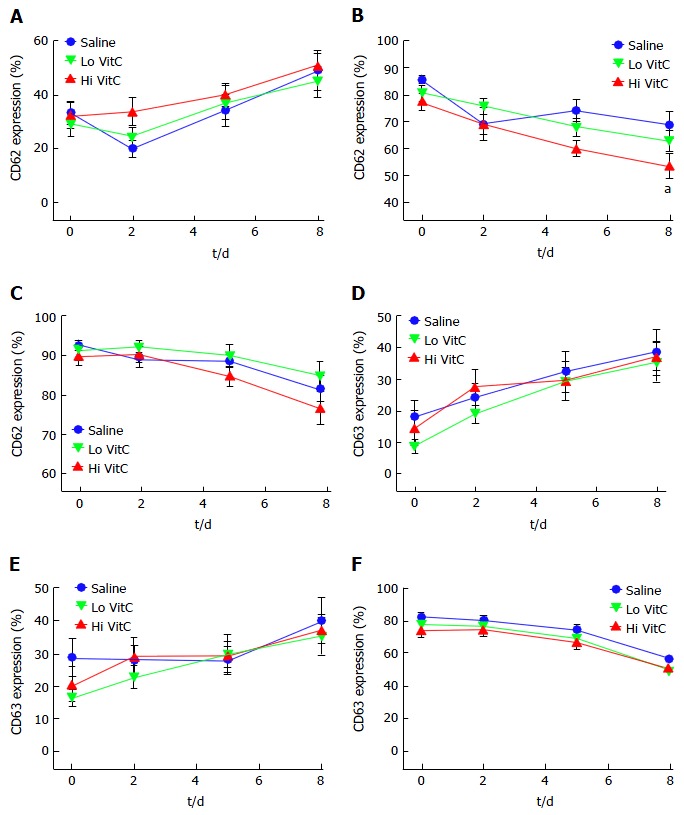
Vitamin C exposure did not significantly alter basal or agonist-stimulated CD62 or CD63 expression profiles. Flow cytometry showed that basal expression of both CD62 (A) and CD63 (D) did not differ with the addition of Lo/Hi VitC. ADP-stimulated CD62 (B) expression was lower on day 8 in the Hi VitC group (n = 10/group, aP < 0.05), but CD63 (E) expression was not. Thrombin-stimulated CD62 (C) and CD63 (F) expression also did not differ across the three groups (n = 10/group). VitC: Vitamin C; ADP: Adenosine diphosphate.
Effects of VitC exposure on eicosanoids metabolism in PLTs
Eicosanoids analysis was carried out on days 0, 2, 5 and 8 using aliquots of the PPP fraction of each sample. The levels of the free polyunsaturated fatty acids: Arachidonic acid (AA), eicosapentanoic acid (EPA), and docosahexanoic acid (DHA) did not differ significantly across the three groups throughout the study period (Figure 7A-C). Interestingly, free AA levels showed an initial drop from baseline in the three groups, and then remained unchanged through day 8. Formation of 11-/12-/15-HETE, products of AA metabolism via the lipoxygenases pathway showed a gradual increase over storage time (Figure 7D-F). The magnitudes of changes were not significantly different between the Lo VitC and the saline groups. However, levels of 11-HETE was significantly higher at days 5 and 8 in the Hi VitC group when compared to saline (Figure 7D, P < 0.05). Hi VitC supplementation significantly augmented 12-HETE levels on days 2, 5 and 8 (Figure 7E, P < 0.05). With respect to 15-HETE, only day 8 levels were significantly higher in the Hi VitC group compared to saline (Figure 7F, P < 0.05). On similar lines, thromboxane B2 (TXB2) (the stable metabolite of TXA2) and prostaglandin E2 (PGE2), products of AA metabolism via the cyclooxygenases pathway were not significantly different between the Lo VitC and saline group. However, Hi VitC supplementation was associated with significantly higher TXB2 levels on days 5 and 8 (Figure 7G, P < 0.05); and significantly higher PGE2 levels on days 2 and 8 compared to saline (Figure 7H, P < 0.05).
Figure 7.
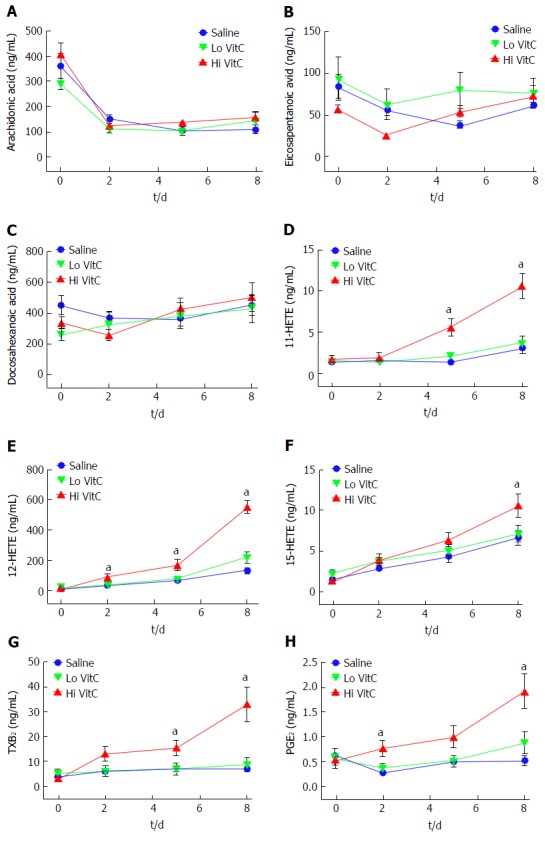
Hi vitamin C, but not Lo vitamin C exposure, was associated with significant changes in the eicosanoid profile over time. Addition of Lo/Hi VitC did not affect the levels of the PUFA: AA (A), EPA (B) and DHA (C) in comparison to saline controls (n = 10/group). Addition of Hi VitC was associated with significantly higher levels of 11-HETE (D) on days 5 and 8 (n = 10/group, aP < 0.05); 12-HETE (E) on days 2, 5 and 8 (n = 10/group, aP < 0.05); and 15-HETE (F) on day 8 (n = 10/group, aP < 0.05). TXB2 (G) was also significantly higher in the Hi VitC group on days 5 and 8 (n = 10/group, aP < 0.05). In addition, PGE2 levels (H) were significantly higher on days 2 and 8 in the Hi VitC group (n = 10/group, aP < 0.05). VitC: Vitamin C; PUFA: Polyunsaturated fatty acids; AA: Arachidonic acid; EPA: Eicosapentanoic acid; DHA: Docosahexanoic acid; TXB2: Thromboxane B2; PGE2: Prostaglandin E2.
DISCUSSION
VitC is one of the most enduring and popular alternative medical treatments sought after. Beyond its oral use to treat scurvy, parenteral VitC has been used by CAM practitioners for more than 6 decades[27-29]. The most controversial use of high dose VitC as a cancer treatment was promoted by the Nobel Laureate, Cameron et al[30,31]. Recently published evidence has demonstrated that intravenous, but not oral administration of VitC produces pharmacologic plasma concentrations of VitC[32]. This has elucidated possible mechanisms of action of intravenous VitC and for the first time made therapeutic effects, biologically plausible[33]. In the past few years this therapeutic option has been implemented most often as adjunct therapy in diverse conditions such as sepsis, infections, autoimmune diseases and cancers[15-19]. The basis for use of high dose intravenous VitC has been established in pre-clinical studies in which VitC modulated coagulopathies in disease states. For example, Swarbeck et al[34] showed that VitC attenuates plasminogen activator inhibitor-1 expression and release in an in vitro model of sepsis. On similar lines Secor et al[35] showed that VitC reduces mouse platelet aggregation and surface P-selectin expression in an ex vivo model of sepsis. However, to date, no studies have directly examined the effect of high doses of VitC on human PLT function. To address this, we asked the question whether exposure to high doses of VitC, as would normally be observed with high dose intravenous VitC therapy, have any effect on PLT function.
In our study we found that PLTs exposed to VitC rapidly accumulated millimolar quantities of VitC as early as day 2 and maintained these levels throughout the study period (Figure 1). Savini et al[8] showed that human PLTs possess the VitC transporter SVCT2, which enable PLTs to increase intracellular levels of VitC. Importantly, while PLTs typically have approximately 4 mmol/L intracellular VitC in normal plasma, exposure to 3 mmol/L VitC increased intracellular VitC levels to > 15 mmol/L. This is significant, especially in cardiovascular pathologies since PLT activation and aggregation are modulated by reactive oxygen species[36]. VitC can alter the oxidative state of PLTs and inhibit the expression of CD40L, a transmembrane protein with pro-inflammatory and pro-thrombotic properties[12]. Indeed, oral administration of VitC has been reported to reduce arterial stiffness and platelet aggregation[37].
VitC did not alter the pH of the PC throughout the standard 5 day storage period when compared to saline controls (Figure 2). Unlike the saline treated PLTs whose pH stayed in the neutral range on day 8, there was a significant drop in pH in the VitC treated PLTs. However, it is unlikely that these pH changes would significantly alter blood pH due to presence of the carbonic acid-bicarbonate buffer, the phosphate buffer system, which consists of phosphoric acid (H3PO4) in equilibrium with dihydrogen phosphate ion (H2PO4-) and H+ and hemoglobin that play an important role in regulating the pH of the blood.
Our results show that the changes in PT, PTT and fibrinogen were comparable across the three groups throughout the study period and that exposure to high doses of VitC did not adversely impact these parameters (Figure 3). Only a few studies have employed TEG to evaluate functionality of human PLTs in the PC’s[38-41]. However, unlike previous studies that adjusted PRP counts using freshly thawed PPP, we used same sample PPP to make adjustments. While TEG parameters across the three groups were comparable, some critical differences were observed on day 8 in the Hi VitC treated PLTs (Figure 4). The Hi VitC treated group showed a prolonged R-time, and a reduced MA as compared to the saline controls. Also, the Hi VitC groups had significantly delayed kinetics which was evidenced by prolonged K-times and reduced α-angle. It is unclear at this time why these changes occurred and future mechanistic approaches are needed to explain this finding. However, the corollary from these studies is that this inhibitory effect of VitC may have deleterious clinical implications if high dose VitC therapy is instituted for 8 d or more, or if plasma VitC levels are maintained at 3 mmol/L or higher for long periods of time.
Platelet aggregation studies show that Lo/Hi VitC addition did not alter the baseline or collagen and ADP-induced platelet aggregations (Figure 5A and B) and collagen and ADP-induced ATP secretion throughout the study (Figure 5C and D). Flow cytometry showed that VitC supplementation had no effect on basal CD62p and CD63 expression during storage (Figure 6A and D). Following ADP (Figure 6B) or thrombin (Figure 6C) stimulation, the CD62 expression showed a steady decrease that was not significantly different across the three groups (except for Hi VitC). Lo/Hi VitC induced changes in CD63 expression with ADP or thrombin (Figure 6E and F) were not significant over 8 d. These results imply that exposure of normal PLTs to high concentrations of VitC has virtually no impact on agonist induced platelet aggregation under these conditions.
Although we did not observe differences in the levels of free AA, EPA and DHA in the plasma of stored PCs, exposure to Hi VitC was associated with a significant increase in the levels of PGE2, TXB2, 11-, 12- and 15-HETE (Figure 7). Some of these free fatty acids (FFAs) have roles in host defense against potential pathogenic or opportunistic microorganisms. Indeed, there is extensive literature demonstrating the antibacterial effects of various free fatty acids from a wide range of biological sources including plants, animals and algae[42]. Whilst their antibacterial mode of action is still poorly understood, studies have shown that their prime target is the cell membrane where FFAs disrupt the electron transport chain and oxidative phosphorylation. Besides interfering with cellular energy production, FFA also inhibit enzyme activity, impair nutrient uptake, and participate in the generation of peroxidation and auto-oxidation degradation products or direct lysis of bacterial cells. While intravenous VitC has been shown to reduce bacterial burden and improve survival in pre-clinical models of sepsis[43-45] it remains to be determined whether the mechanism involves the induction of these FFAs.
As discussed above, both TXB2 and PGE2 levels were significantly higher in PLTs exposed to Hi VitC (Figure 7). Along with possible bacteriostatic effects, there are other potentially beneficial effects associated with induction of these metabolites. For example, a recent study by Bruegel et al[46] demonstrated that reduced release of 11-HETE, PGE2 and TXB2 was associated with increased disease severity and poor prognosis in septic patients. PGE2 plays a dual role balancing PLTs response by stimulation or suppression; and is more generally involved in fine tuning the pro-/anti-inflammatory response[47]. While TXA2 production is associated with PLTs activation, recent data have supported a protective role of TXA2 via its inhibitory regulation of iNOS in the vasculature. In this regard, TXA2 was found to overcome vascular hypo-responsiveness and help maintaining the vascular tone[48]. The increased production of 12- and 15-HETEs observed in Hi VitC treated bags may also be a protective mechanism against the significantly increased TXA2[49,50]. In sum, exposure of PLTs to high doses of VitC alters endogenous production of lipid mediators by PLTs. These mediators could have unappreciated, yet far reaching impacts on not just PLTs function but on the entire circulatory system.
We recognize that our studies had a few limitations. PLTs in storage bags are not in their normal physiologic environment. They do not interact with endothelial cells or other cell types in these storage bags; they are highly concentrated; and also have access only to a finite amount of nutrients[51]. Accumulation of products of metabolism in the storage bags and other factors associated with platelets storage beyond 5 d could impact the system buffering capacity leading to the drop in pH observed in our study. This change in pH combined with the closed nature of the ex vivo system could also account for the observed effects. A second limitation is that in vitro testing of stored PLTs has limitations. While some PLTs functions are lost during storage, others may be recovered in vivo following transfusion. As suggested by Cardigan et al[51], changes observed in stored PLTs might not necessarily abrogate in vivo hemostatic activities. Whether storage of PCs in VitC truly affects hemostatic activities under in vivo conditions remains to be determined as a future endeavor.
COMMENTS
Background
Vitamin C (VitC) is a key modulator of platelet (PLT) function. Platelets store high intracellular concentrations of VitC, which then modify its oxidative state and play a role in its ability to aggregate. High dose intravenous VitC is increasingly being used both by Complementary and Alternate Medicine practitioners and by licensed medical practitioners as adjunct therapy for wide ranging diseases including sepsis, sepsis induced acute lung injury, multiple cancers, iron deficiency in hemodialysis patients and burns. However, there is no information on the impact of high dose VitC on normal PLT function. To address this need, the authors examined the effect of exposing ex vivo human PLTs to high doses of VitC.
Research frontiers
It is well known that VitC is required for normal platelet function. While pre-clinical studies have examined changes in PLT function in disease and the impact of VitC on these functions, no studies have examined PLT function in the presence of such high doses of VitC.
Innovations and breakthroughs
This is the first study to evaluate ex vivo PLT function in the presence of high concentrations of VitC. The innovative approach to use PLT storage bags afforded a reproducible system that allowed for gauging the temporal effects of high doses of VitC on PLT function.
Applications
This study advises moderate levels of caution regarding the extended use of high doses of intravenous VitC. While these high doses have no deleterious impact on PLT function in the short term (up to 5 d), there appear to be unanticipated effects on PLT function as assessed by thromboelastography (TEG) after 8 d of continuous exposure.
Terminology
TEG, is a hemostatic assay that measures the viscoelastic properties (physical) of whole blood clot formation under low shear stress. It shows the interaction of platelets with the coagulation cascade (aggregation, clot strengthening, fibrin cross linking and fibrinolysis).
Peer-review
This is an interesting paper and is worth to be considered for publication.
Footnotes
Manuscript source: Invited manuscript
Specialty type: Critical care medicine
Country of origin: United States
Peer-review report classification
Grade A (Excellent): 0
Grade B (Very good): B, B
Grade C (Good): C
Grade D (Fair): 0
Grade E (Poor): 0
Conflict-of-interest statement: To the best of the authors’ knowledge, no conflict of interest exists.
Data sharing statement: Dataset available from the corresponding author (ramesh.natarajan@vcuhealth.org).
Peer-review started: July 29, 2016
First decision: September 2, 2016
Article in press: November 2, 2016
P- Reviewer: Liu PY, Li W, Schattner MA S- Editor: Kong JX L- Editor: A E- Editor: Li D
References
- 1.Marcus AJ, Safier LB. Thromboregulation: multicellular modulation of platelet reactivity in hemostasis and thrombosis. FASEB J. 1993;7:516–522. doi: 10.1096/fasebj.7.6.8472890. [DOI] [PubMed] [Google Scholar]
- 2.Morrell CN. Immunomodulatory mediators in platelet transfusion reactions. Hematology Am Soc Hematol Educ Program. 2011;2011:470–474. doi: 10.1182/asheducation-2011.1.470. [DOI] [PubMed] [Google Scholar]
- 3.Thomas MR, Storey RF. The role of platelets in inflammation. Thromb Haemost. 2015;114:449–458. doi: 10.1160/TH14-12-1067. [DOI] [PubMed] [Google Scholar]
- 4.Yadav H, Kor DJ. Platelets in the pathogenesis of acute respiratory distress syndrome. Am J Physiol Lung Cell Mol Physiol. 2015;309:L915–L923. doi: 10.1152/ajplung.00266.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mezouar S, Frère C, Darbousset R, Mege D, Crescence L, Dignat-George F, Panicot-Dubois L, Dubois C. Role of platelets in cancer and cancer-associated thrombosis: Experimental and clinical evidences. Thromb Res. 2016;139:65–76. doi: 10.1016/j.thromres.2016.01.006. [DOI] [PubMed] [Google Scholar]
- 6.Naidu KA. Vitamin C in human health and disease is still a mystery? An overview. Nutr J. 2003;2:7. doi: 10.1186/1475-2891-2-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Figueroa-Méndez R, Rivas-Arancibia S. Vitamin C in Health and Disease: Its Role in the Metabolism of Cells and Redox State in the Brain. Front Physiol. 2015;6:397. doi: 10.3389/fphys.2015.00397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Savini I, Catani MV, Arnone R, Rossi A, Frega G, Del Principe D, Avigliano L. Translational control of the ascorbic acid transporter SVCT2 in human platelets. Free Radic Biol Med. 2007;42:608–616. doi: 10.1016/j.freeradbiomed.2006.11.028. [DOI] [PubMed] [Google Scholar]
- 9.Padayatty SJ, Levine M. Vitamin C: the known and the unknown and Goldilocks. Oral Dis. 2016;22:463–493. doi: 10.1111/odi.12446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Levine M, Padayatty SJ, Espey MG. Vitamin C: a concentration-function approach yields pharmacology and therapeutic discoveries. Adv Nutr. 2011;2:78–88. doi: 10.3945/an.110.000109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Olas B, Wachowicz B. Resveratrol and vitamin C as antioxidants in blood platelets. Thromb Res. 2002;106:143–148. doi: 10.1016/s0049-3848(02)00101-9. [DOI] [PubMed] [Google Scholar]
- 12.Pignatelli P, Sanguigni V, Paola SG, Lo Coco E, Lenti L, Violi F. Vitamin C inhibits platelet expression of CD40 ligand. Free Radic Biol Med. 2005;38:1662–1666. doi: 10.1016/j.freeradbiomed.2005.02.032. [DOI] [PubMed] [Google Scholar]
- 13.Ho PP, Walters CP, Sullivan HR. Biosynthesis of thromboxane B2: assay, isolation, and properties of the enzyme system in human platelets. Prostaglandins. 1976;12:951–970. doi: 10.1016/0090-6980(76)90129-5. [DOI] [PubMed] [Google Scholar]
- 14.Srivastava KC. Ascorbic acid enhances the formation of prostaglandin E1 in washed human platelets and prostacyclin in rat aortic rings. Prostaglandins Leukot Med. 1985;18:227–233. doi: 10.1016/0262-1746(85)90022-8. [DOI] [PubMed] [Google Scholar]
- 15.Tanaka H, Matsuda T, Miyagantani Y, Yukioka T, Matsuda H, Shimazaki S. Reduction of resuscitation fluid volumes in severely burned patients using ascorbic acid administration: a randomized, prospective study. Arch Surg. 2000;135:326–331. doi: 10.1001/archsurg.135.3.326. [DOI] [PubMed] [Google Scholar]
- 16.Fowler AA, Syed AA, Knowlson S, Sculthorpe R, Farthing D, DeWilde C, Farthing CA, Larus TL, Martin E, Brophy DF, et al. Phase I safety trial of intravenous ascorbic acid in patients with severe sepsis. J Transl Med. 2014;12:32. doi: 10.1186/1479-5876-12-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Levy TE. Vitamin C, Infectious Diseases, and Toxins: Curing the Incurable. Philadelphia: Xlibris; 2002. [Google Scholar]
- 18.Riordan NH, Riordan HD, Casciari JJ. Clinical and experimental experiences with intravenous vitamin C. J Orthomolecular Med. 2000;15:201–203. Available from: http://orthomolecular.org/library/jom/2000/articles/2000-v15n04-p201.shtml. [Google Scholar]
- 19.Hoffer LJ, Levine M, Assouline S, Melnychuk D, Padayatty SJ, Rosadiuk K, Rousseau C, Robitaille L, Miller WH. Phase I clinical trial of i.v. ascorbic acid in advanced malignancy. Ann Oncol. 2008;19:1969–1974. doi: 10.1093/annonc/mdn377. [DOI] [PubMed] [Google Scholar]
- 20.Mohammed BM, Fisher BJ, Huynh QK, Wijesinghe DS, Chalfant CE, Brophy DF, Fowler AA, Natarajan R. Resolution of sterile inflammation: role for vitamin C. Mediators Inflamm. 2014;2014:173403. doi: 10.1155/2014/173403. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.White NJ, Contaifer D, Martin EJ, Newton JC, Mohammed BM, Bostic JL, Brophy GM, Spiess BD, Pusateri AE, Ward KR, et al. Early hemostatic responses to trauma identified with hierarchical clustering analysis. J Thromb Haemost. 2015;13:978–988. doi: 10.1111/jth.12919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wijesinghe DS, Allegood JC, Gentile LB, Fox TE, Kester M, Chalfant CE. Use of high performance liquid chromatography-electrospray ionization-tandem mass spectrometry for the analysis of ceramide-1-phosphate levels. J Lipid Res. 2010;51:641–651. doi: 10.1194/jlr.D000430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wijesinghe DS, Brentnall M, Mietla JA, Hoeferlin LA, Diegelmann RF, Boise LH, Chalfant CE. Ceramide kinase is required for a normal eicosanoid response and the subsequent orderly migration of fibroblasts. J Lipid Res. 2014;55:1298–1309. doi: 10.1194/jlr.M048207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wijesinghe DS, Chalfant CE. Systems-Level Lipid Analysis Methodologies for Qualitative and Quantitative Investigation of Lipid Signaling Events During Wound Healing. Adv Wound Care (New Rochelle) 2013;2:538–548. doi: 10.1089/wound.2012.0402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Blaho VA, Buczynski MW, Brown CR, Dennis EA. Lipidomic analysis of dynamic eicosanoid responses during the induction and resolution of Lyme arthritis. J Biol Chem. 2009;284:21599–21612. doi: 10.1074/jbc.M109.003822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Wijesinghe DS, Mayton EK, Mietla JA, Mukherjee A, Wu J, Fang X, Chalfant CE. Characterization of lysophosphatidic acid subspecies produced by autotaxin using a modified HPLC ESI-MS/MS method. Anal Methods. 2011;3:2822–2828. doi: 10.1039/C1AY05459G. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Klenner FR. Observations on the dose and administration of ascorbic acid when employed beyond the range of a vitamin in human pathology. J App Nutr. 1971;23:61–88. [Google Scholar]
- 28.Riordan NH, Riordan HD, Meng X, Li Y, Jackson JA. Intravenous ascorbate as a tumor cytotoxic chemotherapeutic agent. Med Hypotheses. 1995;44:207–213. doi: 10.1016/0306-9877(95)90137-x. [DOI] [PubMed] [Google Scholar]
- 29.Calleja HB, Brooks RH. Acute hepatitis treated with high doses of vitamin C. Report of a case. Ohio State Med J. 1960;56:821–823. [PubMed] [Google Scholar]
- 30.Cameron E, Pauling L. Supplemental ascorbate in the supportive treatment of cancer: Prolongation of survival times in terminal human cancer. Proc Natl Acad Sci USA. 1976;73:3685–3689. doi: 10.1073/pnas.73.10.3685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Cameron E, Pauling L. Supplemental ascorbate in the supportive treatment of cancer: reevaluation of prolongation of survival times in terminal human cancer. Proc Natl Acad Sci USA. 1978;75:4538–4542. doi: 10.1073/pnas.75.9.4538. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Padayatty SJ, Sun H, Wang Y, Riordan HD, Hewitt SM, Katz A, Wesley RA, Levine M. Vitamin C pharmacokinetics: implications for oral and intravenous use. Ann Intern Med. 2004;140:533–537. doi: 10.7326/0003-4819-140-7-200404060-00010. [DOI] [PubMed] [Google Scholar]
- 33.Levine M, Espey MG, Chen Q. Losing and finding a way at C: new promise for pharmacologic ascorbate in cancer treatment. Free Radic Biol Med. 2009;47:27–29. doi: 10.1016/j.freeradbiomed.2009.04.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Swarbreck SB, Secor D, Ellis CG, Sharpe MD, Wilson JX, Tyml K. Effect of ascorbate on plasminogen activator inhibitor-1 expression and release from platelets and endothelial cells in an in-vitro model of sepsis. Blood Coagul Fibrinolysis. 2015;26:436–442. doi: 10.1097/MBC.0000000000000273. [DOI] [PubMed] [Google Scholar]
- 35.Secor D, Swarbreck S, Ellis CG, Sharpe MD, Tyml K. Ascorbate reduces mouse platelet aggregation and surface P-selectin expression in an ex vivo model of sepsis. Microcirculation. 2013;20:502–510. doi: 10.1111/micc.12047. [DOI] [PubMed] [Google Scholar]
- 36.Iuliano L, Colavita AR, Leo R, Praticò D, Violi F. Oxygen free radicals and platelet activation. Free Radic Biol Med. 1997;22:999–1006. doi: 10.1016/s0891-5849(96)00488-1. [DOI] [PubMed] [Google Scholar]
- 37.Wilkinson IB, Megson IL, MacCallum H, Sogo N, Cockcroft JR, Webb DJ. Oral vitamin C reduces arterial stiffness and platelet aggregation in humans. J Cardiovasc Pharmacol. 1999;34:690–693. doi: 10.1097/00005344-199911000-00010. [DOI] [PubMed] [Google Scholar]
- 38.Bontekoe IJ, van der Meer PF, de Korte D. Determination of thromboelastographic responsiveness in stored single-donor platelet concentrates. Transfusion. 2014;54:1610–1618. doi: 10.1111/trf.12515. [DOI] [PubMed] [Google Scholar]
- 39.Ostrowski SR, Bochsen L, Windeløv NA, Salado-Jimena JA, Reynaerts I, Goodrich RP, Johansson PI. Hemostatic function of buffy coat platelets in additive solution treated with pathogen reduction technology. Transfusion. 2011;51:344–356. doi: 10.1111/j.1537-2995.2010.02821.x. [DOI] [PubMed] [Google Scholar]
- 40.Svendsen MS, Rojkjaer R, Kristensen AT, Salado-Jimena JA, Kjalke M, Johansson PI. Impairment of the hemostatic potential of platelets during storage as evaluated by flow cytometry, thrombin generation, and thrombelastography under conditions promoting formation of coated platelets. Transfusion. 2007;47:2057–2065. doi: 10.1111/j.1537-2995.2007.01430.x. [DOI] [PubMed] [Google Scholar]
- 41.Reddoch KM, Pidcoke HF, Montgomery RK, Fedyk CG, Aden JK, Ramasubramanian AK, Cap AP. Hemostatic function of apheresis platelets stored at 4°C and 22°C. Shock. 2014;41 Suppl 1:54–61. doi: 10.1097/SHK.0000000000000082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Desbois AP, Smith VJ. Antibacterial free fatty acids: activities, mechanisms of action and biotechnological potential. Appl Microbiol Biotechnol. 2010;85:1629–1642. doi: 10.1007/s00253-009-2355-3. [DOI] [PubMed] [Google Scholar]
- 43.Fisher BJ, Seropian IM, Kraskauskas D, Thakkar JN, Voelkel NF, Fowler AA, Natarajan R. Ascorbic acid attenuates lipopolysaccharide-induced acute lung injury. Crit Care Med. 2011;39:1454–1460. doi: 10.1097/CCM.0b013e3182120cb8. [DOI] [PubMed] [Google Scholar]
- 44.Fisher BJ, Kraskauskas D, Martin EJ, Farkas D, Wegelin JA, Brophy D, Ward KR, Voelkel NF, Fowler AA, Natarajan R. Mechanisms of attenuation of abdominal sepsis induced acute lung injury by ascorbic acid. Am J Physiol Lung Cell Mol Physiol. 2012;303:L20–L32. doi: 10.1152/ajplung.00300.2011. [DOI] [PubMed] [Google Scholar]
- 45.Gaut JP, Belaaouaj A, Byun J, Roberts LJ, Maeda N, Frei B, Heinecke JW. Vitamin C fails to protect amino acids and lipids from oxidation during acute inflammation. Free Radic Biol Med. 2006;40:1494–1501. doi: 10.1016/j.freeradbiomed.2005.12.013. [DOI] [PubMed] [Google Scholar]
- 46.Bruegel M, Ludwig U, Kleinhempel A, Petros S, Kortz L, Ceglarek U, Holdt LM, Thiery J, Fiedler GM. Sepsis-associated changes of the arachidonic acid metabolism and their diagnostic potential in septic patients. Crit Care Med. 2012;40:1478–1486. doi: 10.1097/CCM.0b013e3182416f05. [DOI] [PubMed] [Google Scholar]
- 47.Dolegowska B, Lubkowska A, De Girolamo L. Platelet lipidomic. J Biol Regul Homeost Agents. 2012;26:23S–33S. [PubMed] [Google Scholar]
- 48.Yamada T, Fujino T, Yuhki K, Hara A, Karibe H, Takahata O, Okada Y, Xiao CY, Takayama K, Kuriyama S, et al. Thromboxane A2 regulates vascular tone via its inhibitory effect on the expression of inducible nitric oxide synthase. Circulation. 2003;108:2381–2386. doi: 10.1161/01.CIR.0000093194.21109.EC. [DOI] [PubMed] [Google Scholar]
- 49.Mais DE, Saussy DL, Magee DE, Bowling NL. Interaction of 5-HETE, 12-HETE, 15-HETE and 5,12-diHETE at the human platelet thromboxane A2/prostaglandin H2 receptor. Eicosanoids. 1990;3:121–124. [PubMed] [Google Scholar]
- 50.Fonlupt P, Croset M, Lagarde M. 12-HETE inhibits the binding of PGH2/TXA2 receptor ligands in human platelets. Thromb Res. 1991;63:239–248. doi: 10.1016/0049-3848(91)90287-7. [DOI] [PubMed] [Google Scholar]
- 51.Cardigan R, Turner C, Harrison P. Current methods of assessing platelet function: relevance to transfusion medicine. Vox Sang. 2005;88:153–163. doi: 10.1111/j.1423-0410.2005.00618.x. [DOI] [PubMed] [Google Scholar]