

Abstract
The dispersal patterns of mosquito vectors are important drivers of vector-borne infectious disease dynamics and understanding movement patterns is pivotal to devise successful intervention strategies. Here, we investigate the dispersal patterns of two globally important mosquito vectors, Aedes albopictus and Culex quinquefasciatus, by marking naturally-occurring larvae with stable isotopes (13C or 15N). Marked individuals were captured with 32 CDC light trap, 32 gravid trap, and 16 BG Sentinel at different locations within two-kilometer radii of six larval habitats enriched with either 13C or 15N. In total, 720 trap nights from July to August 2013 yielded a total of 32,140 Cx. quinquefasciatus and 7,722 Ae. albopictus. Overall, 69 marked female mosquitoes and 24 marked male mosquitoes were captured throughout the study period. The distance that Cx. quinquefasciatus females traveled differed for host-seeking and oviposition-seeking traps, with females seeking oviposition sites traveling further than those seeking hosts. Our analysis suggests that 41% of Cx. quinquefasciatus females that were host-seeking occurred 1–2 kilometer from their respective natal site, while 59% remained within a kilometer of their natal site. In contrast, 59% of Cx. quinquefasciatus females that were seeking oviposition sites occurred between 1–2 kilometer away from their larval habitat, while 15% occurred > 2 kilometer away from their natal site. Our analysis estimated that approximately 100% of Ae. albopictus females remained within 1 km of their respective natal site, with 79% occurring within 250m. In addition, we found that male Ae. albopictus dispersed farther than females, suggesting male-biased dispersal in this Ae. albopictus population. This study provides important insights on the dispersal patterns of two globally relevant vector species, and will be important in planning next generation vector control strategies that mitigate mosquito-borne disease through sterile insect techniques, novel Wolbachia infection, and gene drive strategies.
Author summary
Resolving patterns of mosquito dispersal across landscapes is a critical step toward the development of effective control strategies that mitigate vector-borne disease transmission and its public health burden. Here, we used a recently developed technique involving the enrichment of aquatic larval habitat with stable isotopes to mark male and female mosquitoes of two important vector species, Aedes albopictus and Culex quinquefasciatus. We show that the patterns of dispersal were fundamentally different between these urban vectors. Culex quinquefasciatus dispersed much further than Aedes albopictus. In addition, male Aedes albopictus dispersed further than female mosquitoes. Our study suggests that infectious disease agents transmitted by Culex quinquefasciatus will be more difficult to control locally than those transmitted by Aedes albopictus. Our results on sex-biased dispersal in Aedes albopictus have implications for next-generation vector control strategies that rely on the release of sterile or sterilizing males to control mosquito populations. Finally, our study continues to show the utility of the stable-isotope marking technique to study mosquito movement.
Introduction
Understanding patterns of mosquito dispersal is paramount to the development of effective vector control strategies that mitigate vector-borne disease transmission and its public health burden [1]. The spatial scales of vector control techniques are optimized with information on adult mosquito dispersal. For instance, programs that utilize insecticides must calibrate the spatial scale of insecticide treatment to adult dispersal distances for maximal success [2, 3]. The field implementation of next generation vector control technologies that involve the release of autocidal, sterile, or Wolbachia-infected individuals must account for the likely dispersal distance of released individuals [4, 5], and adjust the number and geographic spread of release sites to cover target areas. At larger scales, mosquito dispersal may facilitate the spread of infectious diseases into new regions, shaping geographical patterns of disease emergence and our ability to predict and contain the spread of infectious agents [6].
Mosquitoes disperse to find resting sites, mates, nectar sources, blood sources, and oviposition sites [7, 8]. The distribution of these resources in the environment is an important modulator of mosquito movement. For instance, Edman et al. [9] demonstrated that the probability of dispersal of female Aedes aegypti (Linnaeus) is greater from homes that have fewer potential oviposition sites. Maciel-De-Freitas et al. [10] demonstrated that Aedes albopictus (Skuse) released in forests preferentially dispersed over a kilometer to urban areas to feed on humans, while those released within urban environments were generally sedentary. Additionally, patterns of dispersal may vary between species in the same environment. Many studies suggest that Ae. aegypti [11–14] and Ae. albopictus [15–18] are short dispersing mosquitoes, generally moving less than a few hundred meters. In contrast, other vectors like Culex pipiens (Linnaeus) [19] and Cx. quinquefasciatus (Say) [20–23] mosquitoes may disperse several kilometers. Landscape features can impact mosquito dispersal patterns by serving as corridors that promote mosquito movement. In addition, weather conditions also impact mosquito dispersal distance and direction. For instance, Lapointe [21] demonstrated that Cx. quinquefasciatus in Hawaii moved along roads in the landscape and dispersed predominately in the direction of prevailing winds. Finally, some studies have suggested that mosquito dispersal patterns vary between sexes [24], while others suggest that they are generally similar [25, 26].
Dispersal in mosquitoes has been investigated using mark-release-recapture designs in which adult mosquitoes are marked with dusts, dyes, paints, trace elements, and radioactive isotopes [27]. Typically, these marked adult individuals are released at a specific point and then subsequently recaptured at other sites. These techniques can be highly invasive, tedious, and require the rearing and marking of large quantities of adults. While utilized widely, these methods may alter the behavior of the mosquito, introducing artifacts in data that skew dispersal patterns. In addition, the artificial release of these insects augments local populations, potentially increasing the capacity for those populations to transmit infectious disease. Ideal protocols for adult mosquito mark-recapture studies should involve a marker that is environmentally safe, cost-effective, easy to use, and does not inhibit normal vector biology [27].
Stable isotopes offer safe and useful biological tracers as they occur naturally in the environment, do not decay, and are non-toxic. In a recent study, Hamer et al. [28] developed a stable isotope method to mark naturally-occurring mosquitoes during their larval stage. The laboratory experiments from this study suggested life-long retention of the marker with no apparent impact on morphology or survival. There are several advantages to using stable isotopes to mark mosquitoes during dispersal studies. One principal advantage is the ability to mark naturally occurring immature mosquitoes in aquatic habitat by treatment with isotopically-enriched material. The larval habitat of mosquitoes, such as Culex sp. and Aedes sp., is typically confined (often in artificial containers in urban settings) and thus easily enriched with stable isotopes during larval development. Hamer et al. [28] also showed that there is no evidence of transgenerational marking and that the isotopic retention was higher in 15N-enriched adults (δ15N = +500 ‰ at 55 days post emergence) than 13C-enriched mosquitoes (δ13C = +100 ‰ at 55 days post emergence). Hamer et al. [19] implemented this method to study the dispersal of Cx. pipiens, the primary vector for West Nile virus (WNV), in suburban Chicago, Illinois. That study enriched Culex larvae in productive catch basins with 15N. A survey of over 30,000 mosquitoes revealed 12 15N-marked individuals, yielding a mean distance traveled of 1.15km.
Here, we use a similar stable isotope marking protocol to investigate the dispersal of two important mosquito vectors, Cx. quinquefasciatus and Ae. albopictus, in central Texas, a severely neglected arbovirus hotspot [29]. Cx. quinquefasciatus is accepted as the principal vector of WNV in much of the southern United States [30–32], and is a globally important vector of human filariasis [33], St. Louis encephalitis virus [34], and avian malaria [35]. Ae. albopictus is now a globally invasive mosquito that originated in southeast Asia. This species is a competent vector for numerous arboviruses, including dengue virus (DENV) [36], chikungunya virus (CHIKV) [37–39], yellow fever virus (YFV) [40], and potentially Zika virus (ZIKV) [41], which has recently emerged across the Americas. Given the cosmopolitan distributions of Cx. quinquefasciatus and Ae. albopictus, an improved understanding of dispersal behavior and distance will improve the management of these important vectors globally.
Materials and methods
Stable isotope enrichment
From July 1st to August 31st 2013, two sites were treated with stable isotopes, one site with 15N-potassium nitrate (30° 36' 16.83"N, 96° 19' 34.29"W) and the other site with 13C-glucose (30° 36' 11.196"N, 96° 19' 48.021"W). Enrichment sites were separated by approximately 0.5km and each site consisted of three black tubs (i.e. artificial containers), 30 (width) x 50 (length) by 20 cm (height), filled with approximately three liters of water, and allowed mosquitoes to naturally develop within the environment. The initial treatment concentration was 2.0 mg of isotope per liter of water. Every third week, one container from each enrichment site was disposed of; new water was added and again enriched with the initial treatment. Culex egg rafts from wild females were laid directly on the water and a Whatman filter paper was taped to the side of the tub to allow Ae. albopictus oviposition. The filter paper containing Aedes eggs were dried for 48 hours and then submerged in the tub water to allow hatching. Because the larval habitats were confined, there was no concern of downstream enrichment of the surrounding environment. The containers were consistently monitored for any evaporation, exploitation, and rainfall events causing overflow. Containers were inspected for egg rafts, larvae, and pupae every three days, under the assumption that there would be new pupae every 48-72h. We used the counts of pupae to estimate the number of mosquitoes of each species emerging from enriched containers over the study period. Subsamples of 10-4th instars and pupae were collected and identified as Cx. quinquefasciatus or Ae. albopictus, and were submitted for stable isotope analysis to confirm enrichment.
Larvicide
During the mark-capture study, we implemented a larvicide program by treating larval habitat of Culex and Aedes mosquitoes with methoprene products (Altosid). One justification for larvicide was to ensure that our project would not result in a net increase in mosquito productivity in a region with a potential for arbovirus transmission. Even though we were allowing a few containers to produce mosquitoes, we would remove many more mosquitoes from the landscape by implementing a larvicide program in addition to our intensive adult mosquito trapping. The second justification for conducting a larvicide program was to reduce the total un-marked mosquito population in the study region. With this larvicide program we increased the probability of detecting an isotopically-enriched pool (i.e. marked pool) by reducing the population of un-marked mosquitoes in the study area.
Prior to the start of the stable isotope amendment, we surveyed containers in the mark-capture study region (213.5 hectares) for the presence of water and immature mosquitoes. On June 26th and July 12th 2013, habitat containers containing water were treated with either a Altosid 7-gram water-soluble packet (30-day submerged residual activity; 4.25% S-methoprene) or 3.5-grams of the granular formula (up to 21-day residual control; 4.25% S-methoprene). The stagnant margins of a 1.1 km creek running through the residential study area was also treated with Altosid extended release briquette (150-day residual control; 2.1% S-methoprene). A combination of 14 briquettes, 425 g of the granular formula and 53 water-soluble packets were distributed to various containers holding water and the creek on June 26, 2013. On July 12, 2013, these same habitat containers and the creek were treated again with 6 briquettes, 1,000 g of granular formula and 133 water-soluble packets. Water habitat was treated with these various products of different formulations according to the Altosid label. The active ingredient in Altosid, S-methoprene is an insect growth inhibitor that does not influence the oviposition behavior of Culex and Aedes mosquitoes [42, 43]. Therefore, it is unlikely that the Altosid treatment resulted in changes in female mosquito dispersal that is in part modulated by oviposition behavior.
Adult mosquito trapping
Mosquitoes were trapped from May to September 2013 in College Station, Texas. Three types of mosquito traps were used for this experiment: 32 CDC gravid traps (The John Hock Company, Gainesville, Florida), 32 CDC miniature light traps (BioQuip Products, Inc., Rancho Dominguez, CA), and 16 BG-Sentinel traps (Biogents, Regensburg, Germany) were set weekly (Figs 1–8). Gravid traps were baited with organically enriched water made by infusing rabbit pellets with water, leaving in the sun for about six days, and then deploying in gravid trap tubs. Light and BG traps were baited with CO2 by placing a cooler with 2 kg of dry ice next to the traps. The BG traps were also baited with the BG-Lure. All traps were set in the evening and collected the next morning and each trap location was visited once per week during the study. Trap locations were placed in all directions from the enrichment sites (Figs 1–8), with a goal of an even distribution of the three trap types in the cardinal directions. The density of traps was highest closest to the release point and the exact location of traps was dictated by receiving permission from private homeowners. The closest mosquito trap was 26.6 m and the furthest was 2.16 km from the 15N enrichment site. The mosquito trap nearest to the 13C enrichment site was at 27.7 m and the furthest was 2.46 km away. The mean trap distance for 13C and 15N was 0.96 km and 0.95 km, respectively. Mosquitoes were identified to species and sex based on morphological keys [44, 45]. Approximately half of the individual female Cx. quinquefasciatus and Ae. albopictus mosquitoes were placed in pools of up to 4 individuals and prepared for stable isotope testing following the same general protocol of Hamer et al [28].
Fig 1. Map of mark-capture study region in College Station, Texas demonstrating the 15N-enrichment site and the spatial distribution of enriched female Cx. quinquefasciatus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The satellite image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. The figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
Fig 8. Map of mark-capture study region in College Station, Texas demonstrating the 13C-enrichment site and the spatial distribution of enriched male Ae. albopictus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
Weather data were collected using an existing weather station located at the Texas A&M University Turfgrass Research Laboratory approximately 3 km northwest from the 13C and 15N enrichment sites. The weather station recorded hourly data for temperature, wind speed, wind direction, and precipitation. From the weather observation data, East-West and North-South components of wind speed and direction vector were calculated for each month, and total season [19].
Stable isotope analysis
Fourth instar larvae, pupae, and adult mosquitoes were stored at -40°C and processed for stable isotope analysis by drying and crimping each sample. Mass was estimated based on previously recorded data. Samples were dried at 50°C for 18–24 h, encapsulated into tin capsules that were crimped into a spherical shape, placed into a 96-well plate arranged to include standards, and submitted for stable isotope carbon (13C/12C) and nitrogen (15N/14N) analysis at the Stable Isotope Geosciences Facility, Texas A&M University, College Station, Texas. Initial samples, which required a shorter turn-around period in order to facilitate enrichment activities, were sent to Isotech Laboratories Inc., Champaign, Illinois. Texas A&M samples were analyzed using a Carlo Erba NA 1500 Series 2 Elemental Analyzer (EA) attached to a ThermoFinnigan Conflo III and a ThermoFinnigan Delta Plus XP isotope ratio mass spectrometer (IRMS). Briefly the EA combusts the samples at 1,200°C and the combusted samples passed through two reactors to convert the nitrogen oxides generated in the oxidation reactor to N2 gas. The resulting CO2 and N2 gases are chromatographically separated and then analyzed on the IRMS.
The peak areas of sample mass-to-charge ratios 28 (N2) and 44 (CO2) of a combusted sample are converted to total mass of nitrogen and carbon, respectively, using an intra-run calibration that allows for the calculation of the total nitrogen and carbon content. Raw sample δ15N and δ13C measurements are converted to the Air and VPDB isotopic scales, respectively, through an intra-run, two-point calibration of ~1 mg of L-glutamic acid standards with known isotopic values (USGS 40: δ15N = -4.52‰ Air, δ13C = -26.39‰ VPDB and USGS 4: δ15N = 47.57‰ Air, δ13C = 37.63‰ VPDB). Results are presented in standard delta (δ) notation: δX = [(Rsample/Rstandard)-1] x 1,000, where R is the ratio of the heavy to light stable isotope in the sample and standard. Internal laboratory standards (approximately every 12 unknowns) are used to measure analytical precision. For natural abundance samples, the analytical uncertainty is ± 0.20‰ for δ13C and δ15N. For labeled samples whose isotopic value extends beyond the range of the USGS 40 and 41 calibration standards the uncertainty will increase the more positive the value.
Data analyses
To quantify Mean Distance Traveled (MDT) for each of our captured marked mosquito, we used a formula outlined by Silver [1] with a correction factor accounting for different trap densities in each annulus [19, 46]. We modeled the probability that species-specific pools composed of 2–4 female mosquitoes were marked by stable isotope enrichment with a general linear mixed model assuming a binomial error distribution. Models were implemented in the lme4 [47] package in program R. We added isotope (15N or 13C), week of capture (8 levels), and location of trap (78 groups) as random effects in the model. The number of individuals per pool was incorporated into the model by the offset function in program R. For Cx. quinquefasciatus, models included 4,098 observations. Fixed effects included distance from trap location, trap type (host seeking trap like a BG-Sentinel or CDC light trap, or an oviposition trap), direction of dispersal (four levels of 90° portions with due North set at 0° from source locations), and an interaction between distance and trap type. We discriminated on the basis of Akaike Information Criteria corrected for small sample size (AICc) among a set of candidate models including a full model with all fixed effects, 5 nested models of the full model, and an intercept only model. For Ae. albopictus, models included 1,372 observations. Given the small dispersal distances and lower recapture rate, we only included distance from larval habitat source as a fixed effect in a model and compared it to an intercept only model based on AICc.
We tested for differences between the sexes in the relationship between the probability of marked pools and distance from the larval source habitat with a sex*distance interaction. This modeling strategy also used a general linear mixed model assuming a binomial error distribution implemented in lme4 package, with the same random error structure described above. We tested the interaction effect with a parametric bootstrap of the log-likelihood ratio between a model that included the interaction in addition to the main effects and a model that included only the main effects. The bootstrap was implemented in the R package “pbkrtest” [48].
We estimated an index of relative density of marked mosquitoes within consecutive annuli centered on the enrichment site based on predictions from the best-fit models for each species, using modified equations from Morris et al [46]. We assume that model predictions of the probability of pool enrichment represent the probability of detecting a dispersing mosquito. The probability of detecting a dispersing individual is assumed to be directly proportional to the number of dispersing mosquitoes (i.e. marked mosquitoes) and the trap effort within an annulus (i.e. the number of traps operated across trap type), and inversely proportional to the area of the annulus (see Eq 1).
| (1) |
We solve the equation for the estimated number of dispersing mosquitoes and interpret it as an index of the density of dispersing mosquitoes in our study. Relative density was estimated as the value for the number of dispersing mosquitoes of a given annulus divided by the sum of those values across all annuli.
Results
Stable isotope enrichment
Mosquito larvae were collected from treated containers and had a mean enrichment of 1,130.7±914.8 (n = 20) and 226.7±305.5 (n = 16) for δ15N and δ13C, respectively. We estimated that our larval habitats produced 1,240 Cx. quinquefasciatus and 1,003 Ae. albopictus from July 1st to August 31st 2013 by counting the number of pupae present in the enriched larval habitat every 48–72 hours. This number was used to estimate recapture rates (see below). Throughout the manuscript, we assume a 1:1 sex ratio in the adult population that emerged from the enriched larval habitat. A total of 298 and 482 larvae subsampled throughout the field season were collected and identified to be Ae. albopictus (13C n = 234, 15N n = 64) and Cx. quinquefasciatus (13C n = 157, 15N n = 325), respectively.
Adult mosquito trapping
We collected a total of 71,962 female mosquitoes between May and September, of which 32,140 were Cx. quinquefasciatus (44.7%) and 7722 were Ae. albopictus (10.7%). A total of 2,758 female pools and 331 male samples were analyzed for the presence of enriched 15N and 13C. Of these, 69 (2.5%) and 24 (7.3%) female and male pools were enriched with a stable isotope, respectively. Eight enriched female and 2 male pools were captured in light traps, 29 female and 7 male pools were captured in gravid traps and 32 female and 15 male pools were captured in BG-Sentinel traps.
Weather data is reported in the supplementary online material (S1 Table). Throughout the study period, wind speed ranged from 0.45 m/s to 7.16 m/s with a net wind speed of 1.84 m/s. Wind direction ranged from -23.8 to 325.7 with a net wind direction of 150.1 vector degrees, equating to a south by southeast wind direction. Cumulative rainfall from June to August 2013 is 71.9 mm.
Cx. quinquefasciatus dispersal
A total of 2,066 female Cx. quinquefasciatus pools (8,002 individuals) were analyzed for presence of stable isotope enrichment. Of those tested, 12 were enriched with 15N with a mean δ15N of 1,273.6±530.0 ‰ (Figs 1 and 9A). The mean δ15N of unenriched female Cx. quinquefasciatus mosquito pools was 9.4±0.1 ‰. We estimate a re-capture rate of 2.9% for Cx. quinquefasciatus that emerged from 15N. The mean distance traveled (MDT) for female Cx. quinquefasciatus marked with 15N was 0.4 km. The closest trap with a captured marked mosquito was 26.6 m and the furthest was 596.7 m from the 15N enrichment site (Fig 1). Of the 2,066 female Cx. quinquefasciatus pools analyzed, 28 were enriched with 13C with a mean δ13C of 23.1±5.4 ‰ (Figs 2 and 9A). Mean δ13C of unenriched female Cx. quinquefasciatus pools was -22.1±0.1 ‰. We estimated a re-capture rate of 10.0% for Cx. quinquefasciatus that emerged from 13C. The MDT for female Cx. quinquefasciatus that emerged from 13C was 1.0 km. The nearest trap with a marked mosquito was 27.7 m and the furthest was 1.9 km from the 13C enrichment site (Fig 2).
Fig 9. Map of mark-capture study region in College Station, Texas demonstrating the 13C and 15N-enrichment larval habitat and the locations of captured of marked Cx. quinquefasciatus females (A), and Cx. quinquefasciatus males (B).

Trap symbols that are black captured zero marked pools and numbers next to symbols in legend represents the number of marked pools captured. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
Fig 2. Map of mark-capture study region in College Station, Texas demonstrating the 13C-enrichment site and the spatial distribution of enriched female Cx. quinquefasciatus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
A total of 161 male Cx. quinquefasciatus pools (632 individuals) were analyzed for stable isotope enrichment. Of these, one was enriched with 15N with a mean δ15N of 685.8 ‰ (Figs 3 and 9B). The mean δ15N of unenriched male Cx. quinquefasciatus mosquito pools was 8.1±0.2 ‰. The estimated re-capture rate was 0.2% for male Cx. quinquefasciatus that emerged from the 15N enrichment site. The MDT for male Cx. quinquefasciatus that emerged from 15N was 0.3 km. The only trap with a captured marked mosquito was 64.1 m from the 15N enrichment site (Fig 3). Of the 161 male Cx. quinquefasciatus pools tested, nine were enriched with 13C with a mean δ13C of 58.2±10.9 ‰ (Figs 4 and 9B). The mean δ13C of unenriched male Cx. quinquefasciatus mosquito pools was -22.3±0.2 ‰. The estimated re-capture rate was 3.2% for male Cx. quinquefasciatus that emerged from the 13C enrichment site. The MDT for male Cx. quinquefasciatus that emerged from 13C enrichment site was 1.2 km. The nearest trap with a captured marked mosquito was 0.84 km and the furthest was 1.7 km from the 13C enrichment site (Fig 4). Mean distance traveled and sample sizes for total and marked pools for both sexes of Cx. quinquefasciatus are summarized in Table 1.
Fig 3. Map of mark-capture study region in College Station, Texas demonstrating the 15N-enrichment site and the spatial distribution of enriched male Cx. quinquefasciatus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
Fig 4. Map of mark-capture study region in College Station, Texas demonstrating the 13C-enrichment site and the spatial distribution of enriched male Cx. quinquefasciatus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
Table 1. Mean distance traveled (MDT) in kilometers +/- standard error for Cx. quinquefasciatus and Ae. albopictus based on sex and stable isotope enrichment type.
| Sex | Isotope | Species | MDT (km) | N pools | N marked pools |
|---|---|---|---|---|---|
| Female | 13C | Cx. quinquefasciatus | 1.0 ± 0.4 | 2,066 | 28 |
| Ae. albopictus | 0.4 ± 0.0 | 692 | 13 | ||
| 15N | Cx. quinquefasciatus | 0.4 ± 0.0 | 2,066 | 12 | |
| Ae. albopictus | 0.3 ± 0.0 | 692 | 16 | ||
| Male | 13C | Cx. quinquefasciatus | 1.2 ± 0.2 | 161 | 9 |
| Ae. albopictus | 1.1 ± 0.1 | 170 | 12 | ||
| 15N | Cx. quinquefasciatus | 0.3 ± 0.0 | 161 | 1 | |
| Ae. albopictus | 0.3 ± 0.0 | 170 | 2 |
The best-fit model predicting the probability of a marked pool of Cx. quinquefasciatus female mosquitoes incorporated an interaction between distance from the enrichment site and trap type (weight = 0.56). The best-fit model was 2.8-times more likely than a nested model that excluded the interaction term (ΔAICc = 2.1, weight = 0.20), 5.1-times more likely than a model that only included distance from source (ΔAICc = 3.3, weight = 0.14), and 7.0-times more likely than the full model that also included direction of dispersal as a fixed effect (ΔAICc = 3.9, weight = 0.08; Table 2). In general, models with a fixed effect of direction and the intercept only model (ΔAICc = 10.7, weight ≈ 0) fit the data poorly (Table 2). The interaction in the best-fit model suggested that the probability of capture declined with distance, but the decline was steeper for pools of individuals from host seeking relative to oviposition traps. Cx. quinquefasciatus males dispersed slightly further than females, however the interaction effect between sex and distance was not significant (p = 0.18; parametric bootstrap of the log-likelihood ratio).
Table 2. Summary of nested models and corresponding Akaike information criteria.
| Models | AICc | ΔAICc | DF | Weight |
|---|---|---|---|---|
| a) Culex quinquefasciatus | ||||
| dist, trap type, dist*trap type | 442.4 | 0 | 7 | 0.56 |
| dist, trap type | 444.5 | 2.1 | 6 | 0.20 |
| dist | 445.6 | 3.3 | 5 | 0.11 |
| dist, trap type, direction, dist*trap type | 446.3 | 3.9 | 10 | 0.08 |
| dist, trap type, direction | 448.3 | 5.9 | 9 | 0.03 |
| dist, direction | 448.9 | 6.5 | 8 | 0.02 |
| intercept-only | 453.0 | 10.7 | 4 | 0.00 |
| trap type | 453.7 | 11.4 | 5 | 0.00 |
| direction | 455.2 | 12.8 | 7 | 0.00 |
| b) Aedes albopictus | ||||
| dist | 202.4 | 0 | 5 | 0.89 |
| dist,direction | 206.6 | 4.2 | 8 | 0.11 |
| direction | 239.1 | 36.7 | 7 | 0.00 |
| intercept-only | 251.6 | 49.2 | 4 | 0.00 |
*dist = distance from larval habitat, trap type has 2 levels (host-seeking or oviposition), direction has four levels (northwest, northeast, southwest, southeast), “*” denotes an interaction
The analysis of relative density corrected for area and trap effort estimated that 41% of Cx. quinquefasciatus females that were host-seeking occurred between 1–2 km from their respective natal site (Fig 10A), while 59% remained within a kilometer of their natal larval habitat. In contrast, 59% of Cx. quinquefasciatus females that were seeking oviposition sites occurred between 1–2 km away from their natal larval habitat, while 15% occurred greater than 2 km away from their natal site (Fig 10B). Only 26% remained within 1 km of their natal site (Fig 10B).
Fig 10. Estimates of the relative density of marked female mosquitoes corrected for trap effort and annulus area for each 250 meter annulus in the analysis.
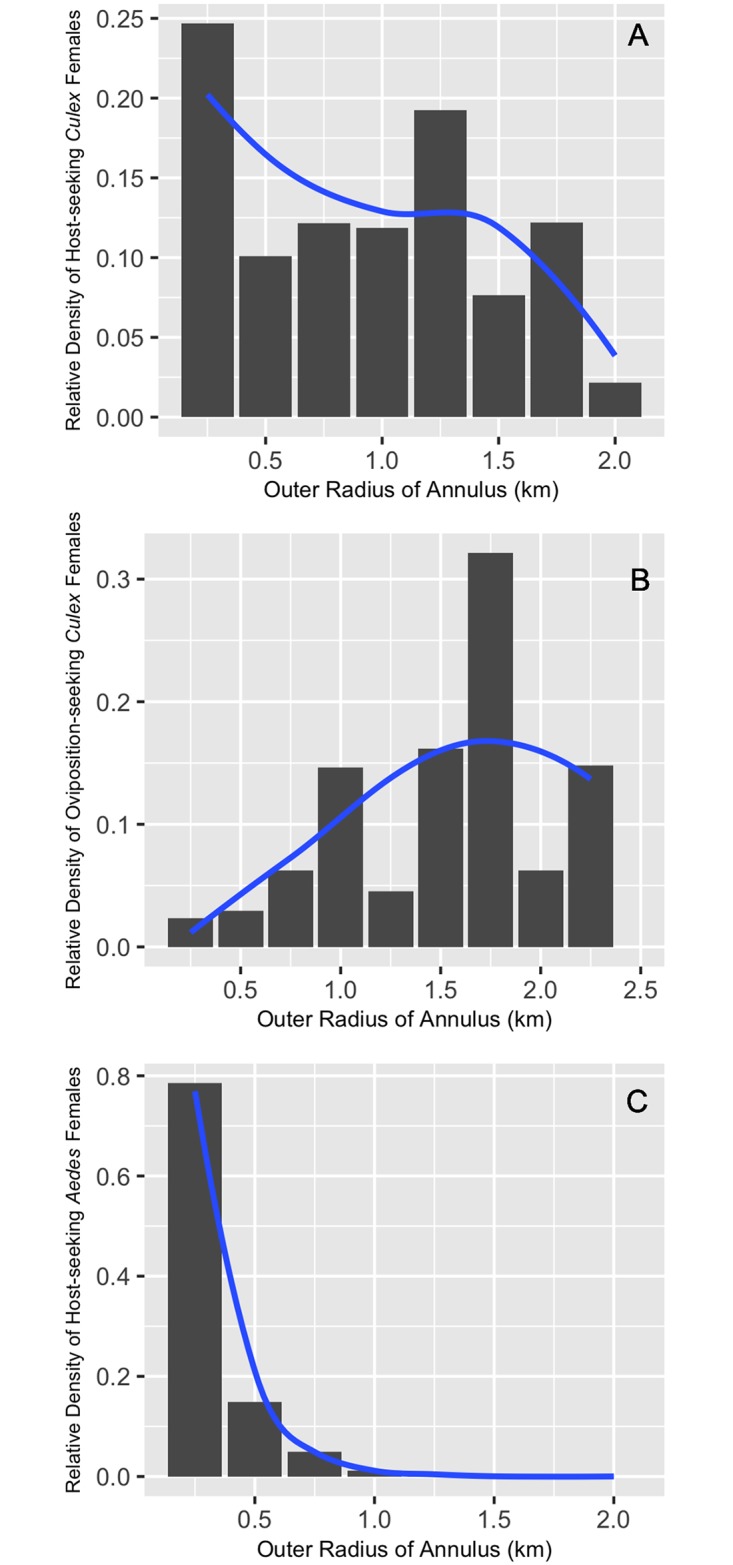
Blue lines represent approximate average curves for the various estimates of relative density across the annuli. The curve was estimated with the smoother function in ggplot2 (geom_smooth). Subpanels A, B, and C correspond to estimated densities of host-seeking Cx. quinquefasciatus females, oviposition-seeking Cx. quinquefasciatus females, and host-seeking Ae. albopictus females, respectively.
Ae. albopictus dispersal
A total of 692 female Ae. albopictus pools (2,535 individuals) were tested for the presence of 15N and 13C. Of those tested, 16 were enriched with 15N with a mean δ15N of 1,388.3±278.0 ‰ (Figs 5 and 11A). The mean δ15N of unenriched female Ae. albopictus mosquito pools was 10.5±0.1 ‰. The estimated recapture rate was 18.0%, for Ae. albopictus females that emerged from 15N enriched sites. The MDT for 15N marked female Ae. albopictus was 0.3 km (Table 1). The closest trap with a captured marked mosquito was 26.6 m and the furthest was 737.5 m from the 15N enrichment site (Fig 5). Of the 692 female Ae. albopictus pools analyzed for stable isotopes, 13 were enriched with 13C with a mean δ13C of 72.5±29.0 ‰ (Figs 6 and 11A). The mean δ13C of unenriched female Ae. albopictus mosquito pools was -23.0±0.1 ‰. The estimated re-capture rate of 13C marked females was 3.8%. The MDT of female Ae. albopictus that emerged from 13C enrichment site was 0.4 km (Table 1). The nearest trap with a captured marked mosquito was 45.3 m and the furthest was 656.2 m from the 13C enrichment site (Fig 6).
Fig 5. Map of mark-capture study region in College Station, Texas demonstrating the 15N-enrichment site and the spatial distribution of enriched female Ae. albopictus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
Fig 11. Map of mark-capture study region in College Station, Texas demonstrating the 13C and 15N-enrichment larval habitat and the locations of captured of marked Ae. albopictus females (A), and Ae. albopictus males (B).

Trap symbols that are black captured zero marked pools and numbers next to symbols in legend represents the number of marked pools captured. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
Fig 6. Map of mark-capture study region in College Station, Texas demonstrating the 13C-enrichment site and the spatial distribution of enriched female Ae. albopictus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
A total of 170 male Ae. albopictus pools (671 individuals) were analyzed for stable isotope enrichment. Of these, two were enriched with 15N with a mean δ15N of 818.8±245.9 ‰ (Figs 7 and 11B). The mean δ15N of unenriched Ae. albopictus mosquito pools was 9.5±0.2 ‰. The estimated re-capture rate was 2.3%. The MDT for male Ae. albopictus that emerged from 15N was 0.3km (Table 1). The only trap with captured marked mosquitoes was 33.5 m from the 15N enrichment site (Fig 7). Of the 170 male Ae. albopictus pools tested for stable isotopes, 12 were enriched with 13C and a mean δ13C of 89.5±8.0 ‰ (Figs 8 and 11B). The mean δ13C of unenriched male Ae. albopictus pools was -23.5±0.1 ‰. The estimated re-capture rate was 3.5%. The MDT for male Ae. albopictus that emerged from the 13C enrichment site was 1.1 km (Table 2). The nearest trap location with a captured marked individual was 314.1 m and the furthest was 1.9 km from the 13C enrichment site (Fig 8). Mean distance traveled and sample sizes for total and marked pools for both sexes of Ae. albopictus are summarized in Table 1.
Fig 7. Map of mark-capture study region in College Station, Texas demonstrating the 15N-enrichment site and the spatial distribution of enriched male Ae. albopictus captures.

Numbers next to symbols in legend represents the number of marked pools captured at each location. The map image was published by the USDA Aerial Photography Field Office as part of the National Agriculture Imagery Program (NAIP), and downloaded as a GIS file. Figure was produced using ArcGIS 10.2 (Esri, Redlands, CA).
The model predicting the probability of enrichment in female Ae. albopictus pools as a function of distance from the source (weight = 0.89) better fit the data than models that included direction of dispersal (Table 2) and the intercept-only model (ΔAICc = 49.2, weight ≈ 0; Table 2). The sex of individual Aedes albopictus interacted with distance from the source to affect the probability of an enriched pool (p = 0.001; parametric bootstrap of the log-likelihood ratio), with model parameters suggesting that that males disperse farther than females (Fig 12).
Fig 12. Estimates of the relative density of marked female and male Ae. albopictus corrected for trap effort and annulus area for each 250-meter annulus in the analysis from a model that explored an interaction between sex and distance on the probability of capture (see methods).
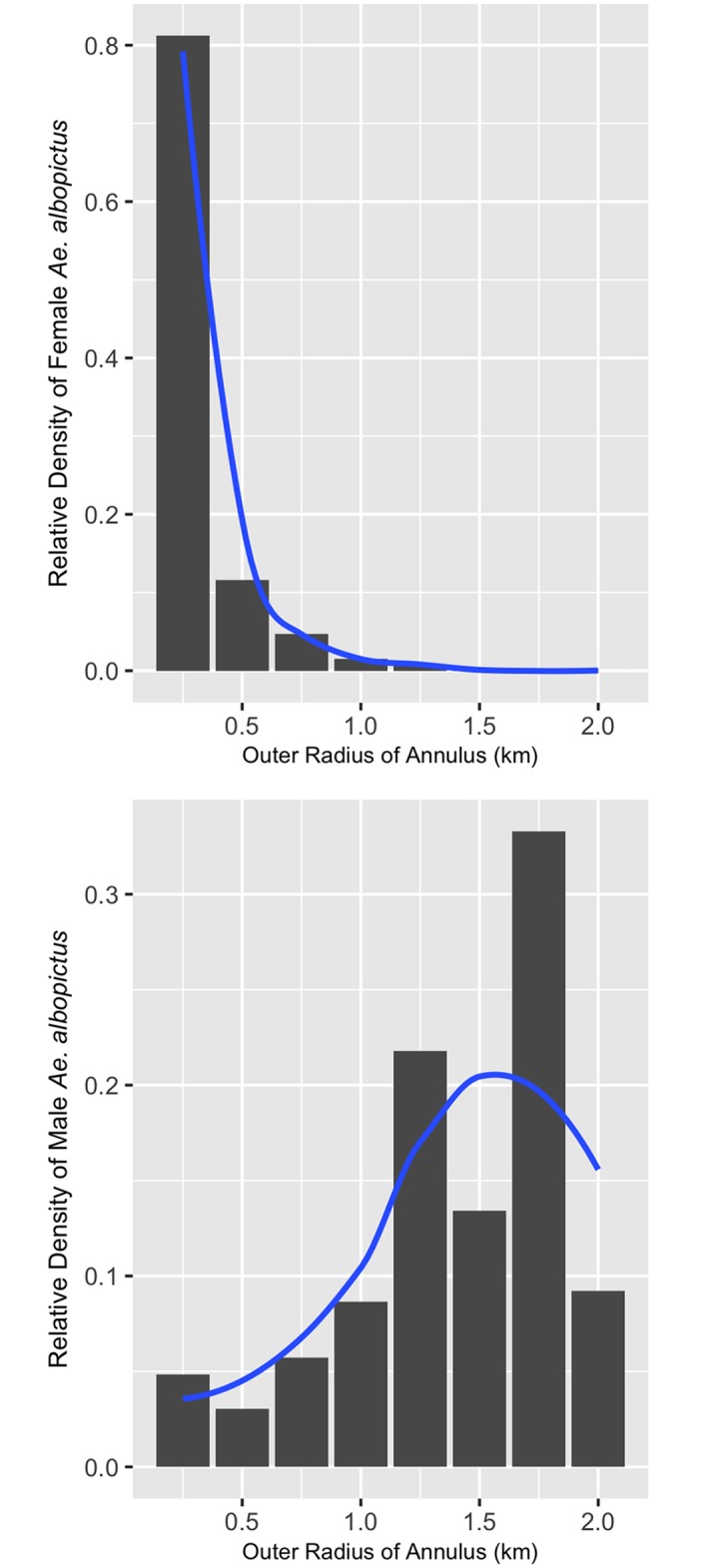
Blue lines represent approximate average curves for the various estimates of relative density across the annuli. The curve was estimated with a smoother function in ggplot2 (geom_smooth).
The analysis of relative density corrected for area and trap effort estimated that 100% of Ae. albopictus females that were host-seeking occurred within 1 km from their respective natal site (Fig 10C), with 79% occurring within 250m.
Discussion
Our study quantified the dispersal patterns of two medically important mosquito vectors, Cx. quinquefasciatus and Ae. albopictus, for both sexes at the same location, simultaneously. Thus our protocol, rare in the mosquito dispersal literature [8], is particularly powerful for the direct comparison of dispersal patterns between mosquito species and sexes. Such comparisons are integral to the development of effective vector control strategies by guiding the optimization of spatial scale for intervention campaigns that may target different vector-pathogen systems. Here, we discuss these results and focus on how they may influence the development of general vector control strategies to control emerging mosquito-borne pathogens.
Our data reveal that the dispersal patterns of Cx. quinquefasciatus and Ae. albopictus, two mosquitoes whose global ranges broadly overlap, are fundamentally different. Similar to other studies, we show that many Cx. quinquefasciatus females disperse 1–2 kilometers, while most Ae. albopictus females remain within 300m of the larval habitat from which they emerged. Long-distance dispersal decreases the efficacy of strategies to contain outbreaks and undermines intervention strategies [3]. In addition, long-distance dispersal increases the spatial scope of vector control efforts, and increases the odds that successfully treated areas may be reinfested from non-treated areas. Our data suggest that outbreaks of mosquito-borne disease will be inherently more difficult to control if vectored by Cx. quinquefasciatus relative to Ae. albopictus and other short distance dispersers like Ae. aegypti. This highlights the potential for Cx. quinquefasciatus and other closely related Culex mosquitoes to drive the emergence of vector-borne infectious disease for pathogens when their vectorial capacity is high.
We find evidence that male Ae. albopictus dispersed farther than females. Previous studies have found no difference between the dispersal distances amongst the sexes in Ae. aegypti, or that females tended to disperse farther than males. Inbreeding avoidance and the reduction in competition among kin for local resources may drive the evolution of sex-biased dispersal [49, 50], as dispersing individuals are less likely to mate with a relative or to compete for mates and other resources with brothers or sisters. It may be more advantageous for males to disperse from natal sites when females are philopatric. This may explain why we observed a strong bias toward long-distance male dispersal in Ae. albopictus compared to Cx. quinquefasciatus. Future studies should continue to address sex-biased dispersal in mosquitoes. The stable isotope protocol employed here may be particularly useful as it allows males and females to disperse naturally from their natal site. This facilitates comparison between sexes among individuals that are emerging from the same place at approximately the same time, reducing the potential for landscape features or weather patterns to influence mosquito dispersal and produce artifacts. With the advent of genetically-modified sterile or autocidal male mosquitoes as next-generation vector control strategies, understanding male dispersal patterns among target mosquito species in different environmental and ecological contexts will be crucial to the field application and ultimate success of these techniques [51].
While our data has broad applicability to vector behavior and control, it is particularly relevant to Texas, a region with a relative high burden of mosquito-borne viruses and a relatively small mosquito control community [29] arboviorus transmission that is woefully understudied. Texas is endemic with WNV since 2002 and large epidemics of WNV neuroinvasive disease occurred in Dallas, TX in 2012 [52] and Houston, TX in 2014 [53]. Previously, Ae. albopictus in the United States was primarily regarded as a nuisance mosquito. However due to epidemic dengue transmission in parts of the Texas, Florida, and Hawaii [36, 54, 55] and the introduction of CHIKV to the United States in 2014 [55], public health officials are being encouraged to develop effective control strategies for Ae. albopictus and Ae. aegypti. Indeed, the first case of autochthonous CHIKV in Texas occurred in 2015 in Cameron County (Texas Department of State Health Services). In addition, the ZIKV epidemic in Latin America and its probable future emergence in Texas have only increased the urgency to devise integrated vector control strategies toward these mosquito vectors. Such efforts will mitigate the potential for these diseases to establish in Texas, and as a result reduce the potential for unimpeded human movement to spread mosquito-borne disease within the United States.
Isotopic enrichment of larvae is simple, effective, relatively inexpensive, and can be achieved in the field. Based on the limited controlled studies, stable isotope marking of the insect does not inhibit the growth or normal biology, and offers life-long retention. However, the method has some limitations. The stable isotope analysis is expensive and can range from $5 to much higher per sample depending on the isotope, facility, and the turn-around time. In the current study we analyzed a total of 3,089 at $6 per sample for a total cost of $18,534. In addition, we analyzed 20 immature mosquitoes for 15N and another 20 for 13C which was $1,200 ($30 per sample for rush turn-around time at Isotech Laboratories Inc.). Additionally, this technique requires a long delay between mosquito collection in the field and eventual stable isotope analysis. From the time of sample receipt, stable isotope labs vary in their turn-around time of results from about 1 week to 20 weeks. Extreme attention to detail is necessary for sample and database management to ensure the stable isotope results are accurately matched to the field data. Continued use of stable isotope analysis in both enrichment and natural abundance study could eventually reduce the cost of this technique and technological advances in the technique could offer more rapid turnaround. Temporal dispersal measurements are often difficult to obtain using this technique, as mosquitoes are not released at a single time point. As a result of this study design, it is difficult to quantify the effects that abiotic factors such as temperature, precipitation, wind, and humidity have on dispersal when using stable isotopes as a marker. This is due to the challenge of determining the exact age of the marked mosquito or the date of the dispersal event. One other challenge with most studies of mosquito dispersal is that the trap design can influence the results of the study; Guerra et al. [8] found a positive correlation between the radius of the experimental area with traps and the mean distance traveled. In the present study, we deployed traps out to about 2 km but the density in each annuli was not constant and the trap types were not balanced among annuli, which could have influenced our results. Stable isotope marking has recently been demonstrated for mosquito vectors of malaria in Tanzania [56] and studies are underway to use this technique not to study dispersal per se, but to evaluate the success of a novel intervention strategy. This study in Tanzania and the current study in Texas reveal the diverse ways in which stable isotopes can be used as useful tools to study of mosquito biology in diverse settings.
Supporting information
(DOCX)
Acknowledgments
We are grateful to the residents of College Station, Texas that granted us to permission to trap mosquitoes on their property. We appreciate support from the Brazos County Health Department. We also thank our team of graduate and undergraduate students that worked on this project as both field and lab technicians: Chris Madeira, Michael Sanders, Andrew Golnar, Chris Tarrand, Sarah Noe, and Tim Foreman. Bret Collier provided valuable discussion and guidance on modeling mark-capture data. Texas A&M University's Department of Soil and Crop Sciences provided weather station data. We appreciate the constructive reviews provided by two anonymous reviewers.
Data Availability
Mosquito dispersal data from this study are available from the Oak Trust (Digital Repository) through Texas A&M University. Data are available at: http://oaktrust.library.tamu.edu/handle/1969.1/158586.
Funding Statement
This study was funded by the Mosquito Research Foundation and the Department of Entomology, Texas A&M University (GLH). The open access publishing fees for this article have been covered by the Texas A&M University Open Access to Knowledge Fund (OAKFund), supported by the University Libraries and the Office of the Vice President for Research. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Silver JB. Mosquito Ecology: Field Sampling Methods. 3rd ed New York: Springer; 2008. [Google Scholar]
- 2.Garcia-Rejon J, Lorono-Pino MA, Farfan-Ale JA, Flores-Flores L, Rosado-Paredes ED, Rivero-Cardenas N, et al. Dengue virus-infected Aedes aegypti in the home environment. Am J Trop Med Hyg. 2008;79(6):940–50. [PubMed] [Google Scholar]
- 3.Killeen GF, Knols BG, Gu W. Taking malaria transmission out of the bottle: implications of mosquito dispersal for vector-control interventions. The Lancet Infect Dis. 2003;3(5):297–303. [DOI] [PubMed] [Google Scholar]
- 4.Bellini R, Albieri A, Balestrino F, Carrieri M, Porretta D, Urbanelli S, et al. Dispersal and survival of Aedes albopictus (Diptera: Culicidae) males in Italian urban areas and significance for sterile insect technique application. J Med Entomol. 2010;47(6):1082–91. [DOI] [PubMed] [Google Scholar]
- 5.Harris AF, Nimmo D, McKemey AR, Kelly N, Scaife S, Donnelly CA, et al. Field performance of engineered male mosquitoes. Nat Biotechnol. 2011;29(11):1034–7. 10.1038/nbt.2019 [DOI] [PubMed] [Google Scholar]
- 6.Venkatesan M, Rasgon JL. Population genetic data suggest a role for mosquito-mediated dispersal of West Nile virus across the western United States. Mol Ecol. 2010;19(8):1573–84. 10.1111/j.1365-294X.2010.04577.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Service MW. Mosquito (Diptera: Culicidae) dispersal—the long and short of it. J Med Entomol. 1997;34(6):579–88. [DOI] [PubMed] [Google Scholar]
- 8.Guerra CA, Reiner RC, Perkins TA, Lindsay SW, Midega JT, Brady OJ, et al. A global assembly of adult female mosquito mark-release-recapture data to inform the control of mosquito-borne pathogens. Parasite Vector. 2014;7 Artn 276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Edman JD, Scott TW, Costero A, Morrison AC, Harrington LC, Clark GG. Aedes aegypti (Diptera: Culicidae) movement influenced by availability of oviposition sites. J Med Entomol. 1998;35(4):578–83. [DOI] [PubMed] [Google Scholar]
- 10.Maciel-de-Freitas R, Neto RB, Goncalves JM, Codeco CT, Lourenco-de-Oliveira R. Movement of dengue vectors between the human modified environment and an urban forest in Rio de Janeiro. J Med Entomol. 2006;43(6):1112–20. [DOI] [PubMed] [Google Scholar]
- 11.Russell RC, Webb CE, Williams CR, Ritchie SA. Mark-release-recapture study to measure dispersal of the mosquito Aedes aegypti in Cairns, Queensland, Australia. Med Vet Entomol. 2005;19(4):451–7. 10.1111/j.1365-2915.2005.00589.x [DOI] [PubMed] [Google Scholar]
- 12.Harrington LC, Scott TW, Lerdthusnee K, Coleman RC, Costero A, Clark GG, et al. Dispersal of the dengue vector Aedes aegypti within and between rural communities. Am J Trop Med Hyg. 2005;72(2):209–20. [PubMed] [Google Scholar]
- 13.Muir LE, Kay BH. Aedes aegypti survival and dispersal estimated by mark-release-recapture in northern Australia. Am J Trop Med Hyg. 1998;58(3):277–82. [DOI] [PubMed] [Google Scholar]
- 14.Maciel-de-Freitas R, Codeco CT, Lourenco-de-Oliveira R. Daily survival rates and dispersal of Aedes aegypti females in Rio de Janeiro, Brazil. Am J Trop Med Hyg. 2007;76(4):659–65. [PubMed] [Google Scholar]
- 15.Bonnet DD, Worcester DJ. The dispersal of Aedes albopictus in the territory of Hawaii. Am J Trop Med. 1946;26(4):465–76. [DOI] [PubMed] [Google Scholar]
- 16.Marini F, Caputo B, Pombi M, Tarsitani G, della Torre A. Study of Aedes albopictus dispersal in Rome, Italy, using sticky traps in mark-release-recapture experiments. Med Vet Entomol. 2010;24(4):361–8. 10.1111/j.1365-2915.2010.00898.x [DOI] [PubMed] [Google Scholar]
- 17.Niebylski ML, Craig GB Jr. Dispersal and survival of Aedes albopictus at a scrap tire yard in Missouri. J Am Mosq Control Assoc. 1994;10(3):339–43. [PubMed] [Google Scholar]
- 18.Lacroix R, Delatte H, Hue T, Reiter P. Dispersal and survival of male and female Aedes albopictus (Diptera: Culicidae) on Reunion Island. J Med Entomol. 2009;46(5):1117–24. [DOI] [PubMed] [Google Scholar]
- 19.Hamer GL, Anderson TK, Donovan DJ, Brawn JD, Krebs BL, Gardner AM, et al. Dispersal of adult Culex mosquitoes in an urban West Nile virus hotspot: A mark-capture study incorporating stable isotope enrichment of natural larval habitats. Plos Neglect Trop Dis. 2014;8(3). ARTN e2768. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Schreiber ET, Mulla MS, Chaney JD, Dhillon MS. Dispersal of Culex quinquefasciatus from a Dairy in Southern California. J Am Mosq Control Assoc. 1988;4(3):300–4. [PubMed] [Google Scholar]
- 21.Lapointe DA. Dispersal of Culex quinquefasciatus (Diptera: Culicidae) in a Hawaiian rain forest. J Med Entomol. 2008;45(4):600–9. [DOI] [PubMed] [Google Scholar]
- 22.Reisen WK, Milby MM, Meyer RP, Pfuntner AR, Spoehel J, Hazelrigg JE, et al. Mark release recapture studies with Culex mosquitoes (Diptera, culicidae) in Southern California. J Med Entomol. 1991;28(3):357–71. [DOI] [PubMed] [Google Scholar]
- 23.Fussell E. Dispersal studies on radioactive-tagged Culex quinquefasciatus Say. Mosq News. 1964;24:422–6. [Google Scholar]
- 24.Trpis M, Hausermann W. Dispersal and other population parameters of Aedes aegypti in an African village and their possible significance in epidemiology of vector-borne diseases. Am J Trop Med Hyg. 1986;35(6):1263–79. [DOI] [PubMed] [Google Scholar]
- 25.Harrington LC, Vermeylen F, Jones JJ, Kitthawee S, Sithiprasasna R, Edman JD, et al. Age-dependent survival of the dengue vector Aedes aegypti (Diptera: Culicidae) demonstrated by simultaneous release-recapture of different age cohorts. J Med Entomol. 2008;45(2):307–13. [DOI] [PubMed] [Google Scholar]
- 26.Maciel-De-Freitas R, Codeco CT, Lourenco-De-Oliveira R. Body size-associated survival and dispersal rates of Aedes aegypti in Rio de Janeiro. Med Vet Entomol. 2007;21(3):284–92. 10.1111/j.1365-2915.2007.00694.x [DOI] [PubMed] [Google Scholar]
- 27.Hagler JR, Jackson CG. Methods for marking insects: Current techniques and future prospects. Annu Rev Entomol. 2001;46:511–43. 10.1146/annurev.ento.46.1.511 [DOI] [PubMed] [Google Scholar]
- 28.Hamer GL, Donovan DJ, Hood-Nowotny R, Kaufman MG, Goldberg TL, Walker ED. Evaluation of a stable isotope method to mark naturally-breeding larval mosquitoes for adult dispersal studies. J Med Entomol. 2012;49(1):61–70. Epub 2012/02/09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hamer GL. Heterogeneity of mosquito (Diptera: Culicidae) control community size, research productivity, and arboviral diseases across the United States. J Med Entomol. 2016; [DOI] [PubMed] [Google Scholar]
- 30.Molaei G, Andreadis TG, Armstrong PM, Bueno R, Dennett JA, Real SV, et al. Host feeding pattern of Culex quinquefasciatus (Diptera: Culicidae) and its role in transmission of West Nile virus in Harris County, Texas. Am J Trop Med Hyg. 2007;77(1):73–81. [PubMed] [Google Scholar]
- 31.Zinser M, Ramberg F, Willott E. Culex quinquefasciatus (Diptera: Culicidae) as a potential West Nile virus vector in Tucson, Arizona: Blood meal analysis indicates feeding on both humans and birds. J Insect Sci. 2004;4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Richards SL, Mores CN, Lord CC, Tabachnick WJ. Impact of extrinsic incubation temperature and virus exposure on vector competence of Culex pipiens quinquefasciatus Say (Diptera: Culicidae) for West Nile virus. Vector Borne and Zoonotic Dis. 2007;7(4):629–36. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Bockarie MJ, Pedersen EM, White GB, Michael E. Role of vector control in the global program to eliminate lymphatic filariasis. Annu Rev Entomol. 2009;54:469–87. 10.1146/annurev.ento.54.110807.090626 [DOI] [PubMed] [Google Scholar]
- 34.Mitchell CJ, Gubler DJ, Monath TP. Variation in infectivity of Saint Louis encephalitis viral strains for Culex pipiens quinquefasciatus (Diptera: Culicidae). J Med Entomol. 1983;20(5):526–33. [DOI] [PubMed] [Google Scholar]
- 35.LaPointe DA, Goff ML, Atkinson CT. Comparative susceptibility of introduced forest dwelling mosquitoes in Hawai'i to avian malaria, Plasmodium relictum. J Parasitol. 2005;91(4):843–9. 10.1645/GE-3431.1 [DOI] [PubMed] [Google Scholar]
- 36.Effler PV, Pang L, Kitsutani P, Vorndam V, Nakata M, Ayers T, et al. Dengue fever, Hawaii, 2001–2002. Emerg Infect Dis. 2005;11(5):742–9. 10.3201/eid1105.041063 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Martin E, Moutailler S, Madec Y, Failloux AB. Differential responses of the mosquito Aedes albopictus from the Indian Ocean region to two chikungunya isolates. BMC Ecol. 2010;10:8 10.1186/1472-6785-10-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pages F, Peyrefitte CN, Mve MT, Jarjaval F, Brisse S, Iteman I, et al. Aedes albopictus mosquito: the main vector of the 2007 Chikungunya outbreak in Gabon. PLoS One. 2009;4(3):e4691 10.1371/journal.pone.0004691 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Bonilauri P, Bellini R, Calzolari M, Angelini R, Venturi L, Fallacara F, et al. Chikungunya virus in Aedes albopictus, Italy. Emerg Infect Dis. 2008;14(5):852–4. 10.3201/eid1405.071144 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Miller BR, Mitchell CJ, Ballinger ME. Replication, tissue tropisms and transmission of yellow fever virus in Aedes albopictus. Trans R Soc Trop Med Hyg. 1989;83(2):252–5. [DOI] [PubMed] [Google Scholar]
- 41.Wong PS, Li MZ, Chong CS, Ng LC, Tan CH. Aedes (Stegomyia) albopictus (Skuse): a potential vector of Zika virus in Singapore. PLoS Negl Trop Dis. 2013;7(8):e2348 10.1371/journal.pntd.0002348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ritchie SA, Long S. Does S-methoprene affect oviposition by Aedes aegypti in an ovitrap? J Am Mosq Control Assoc. 2003;19(2):170–1. [PubMed] [Google Scholar]
- 43.Beehler JW, Mulla MS. Effect of the insect growth regulator methoprene on the ovipositional behavior of Aedes aegypti and Culex quinquefasciatus. J Am Mosq Control Assoc. 1993;9(1):13–6. [PubMed] [Google Scholar]
- 44.Fox M, Wesson D. Key to the Medically Important Mosquitoes of Louisiana. Centers for Disease Control and Prevention; 2006.
- 45.Darsie RF, Ward RA. Identification and Geographic Distribution of the Mosquitoes of North America, North of Mexico. 2nd ed: University Press of Florida; 2004. [Google Scholar]
- 46.Morris CD, Larson VL, Lounibos LP. Measuring mosquito dispersal for control programs. J Am Mosq Control Assoc. 1991;7(4):608–15. [PubMed] [Google Scholar]
- 47.Bates D, Machler M, Bolker BM, Walker SC. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67:1–48. [Google Scholar]
- 48.Halekoh U, Hojsgaard S. Kenward-Roger approximation and parametric bootstrap methods for tests in linear mixed models—the R Package pbkrtest. J Stat Softw. 2014;59:1–32.26917999 [Google Scholar]
- 49.Perrin N, Mazalov VV. Local competition, inbreeding, and the evolution of sex-biased dispersal. Am Nat. 2000;155(1):116–27. 10.1086/303296 [DOI] [PubMed] [Google Scholar]
- 50.Pusey AE. Sex-biased dispersal and inbreeding avoidance in birds and mammals. Trends Ecol Evol. 1987;2(10):295–9. 10.1016/0169-5347(87)90081-4 [DOI] [PubMed] [Google Scholar]
- 51.Winskill P, Carvalho DO, Capurro ML, Alphey L, Donnelly CA, McKemey AR. Dispersal of engineered male Aedes aegypti mosquitoes. PLoS Negl Trop Dis. 2015;9(11):e0004156 10.1371/journal.pntd.0004156 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chung WM, Buseman CM, Joyner SN, Hughes SM, Fomby TB, Luby JP, et al. The 2012 West Nile encephalitis epidemic in Dallas, Texas. J Am Med Assoc. 2013;310(3):297–307. Epub 2013/07/19. [DOI] [PubMed] [Google Scholar]
- 53.Randle YH, Freeman CB, Jackson M, Reyna M, Debboun M. 2014: A record-breaking year for West Nile virus positive mosquito pools in Harris county and the City of Houston, Texas. US Army Med Dep J. 2016;3–16:1–8. [PubMed] [Google Scholar]
- 54.Trout A, Baracco G, Rodriguez M, Barber J, Leal A, Radke E, et al. Locally acquired dengue Key West, Florida, 2009–2010. Morb Mortal Wkly Rep. 2010;59:577–81. [PubMed] [Google Scholar]
- 55.Brunkard JM, Robles Lopez JL, Ramirez J, Cifuentes E, Rothenberg SJ, Hunsperger EA, et al. Dengue fever seroprevalence and risk factors, Texas-Mexico border, 2004. Emerg Infect Dis. 2007;13(10):1477–83. 10.3201/eid1310.061586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Opiyo MA, Hamer GL, Lwetoijera DW, Auckland LD, Majambere S, Okumu FO. Using stable isotopes of carbon and nitrogen to mark wild populations of Anopheles and Aedes mosquitoes in South-Eastern Tanzania. PLoS One. 2016;11(7). ARTN e0159067. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
Data Availability Statement
Mosquito dispersal data from this study are available from the Oak Trust (Digital Repository) through Texas A&M University. Data are available at: http://oaktrust.library.tamu.edu/handle/1969.1/158586.