

Abstract
The long noncoding (lnc) RNA H19 was involved in the tumorigenesis of many types of cancer. However, the role of H19 in the tumorigenesis of colon cancer has not been fully illustrated. Recent studies suggested a potential relationship between H19 and vitamin D receptor (VDR) signaling. Considering the pivotal role of VDR signaling in the colon epithelium both physiologically and pathologically, the correlation between H19 and VDR signaling may have an important role in the development of colon cancer. In this study, the correlation between H19 and vitamin D receptor (VDR) signaling and the underlying mechanisms in colon cancer were investigated both in vitro and in vivo. The results suggested that VDR signaling was able to inhibit the expression of H19 through regulating C-Myc/Mad-1 network. H19, on the other hand, was able to inhibit the expression of VDR through micro RNA 675-5p (miR-675-5p). Furthermore, H19 overexpression induced resistance to the treatment with 1,25(OH)2D3 both in vitro and in vivo. Together, these results suggested that H19 overexpression might be one of the mechanisms underlying the development of resistance to the treatment with 1,25(OH)2D3 in the advanced stage of colon cancer.
Introduction
Colorectal cancer is one of the most common cancers in both men and women. About 140,000 new cases of colorectal cancer are expected in the USA and it remains the third leading cause of death from cancer in 2014 [1]. Although the 5-year mortality rate of colon cancer has slightly declined in recent decades, there is still a pressing need to identify new prognostic factors and potential therapeutic targets for this disease [2], [3].
LncRNAs, with length >200 nucleotides, have been considered as a new kind of gene expression regulator in recent years [4], [5]. LncRNA H19, firstly reported in 1991 by Bartolomei, is considered to play a critical role in the progression of multiple types of cancer [6], [7], [8], [9], [10]. H19 overexpression has been reported to be related with a poor prognosis in both bladder and gastric cancer [11], [12]. However, the mechanism underlying the tumorigenic role of H19 in the development of colon cancer remains to be illustrated.
Vitamin D receptor (VDR) signaling has been reported to be able to attenuate the initiation and development of multiple types of cancer including colorectal cancer [13]. VDR signaling was able to attenuate the proliferation and migration of colon cancer cells via multiple mechanisms including inhibiting Wnt/β-catenin pathway [14]. However, colon cancer cells at advanced stage often showed less sensitive or even resistant to the treatment of 1,25(OH)2D3 (the most active form of vitamin D in the human body) and its analogs [15], [16]. Investigating the mechanisms underlying the development of resistance to VDR signaling and restoring the effects of 1,25(OH)2D3 remains a promising method for seeking new therapeutic targets for colon cancer [17].
Recently, lnc RNA profiling of VDR−/− mice showed that H19 might be involved in the role of VDR signaling in the development of epidermal diseases [18]. Actually, H19 has been reported to be a regulator of multiple signaling pathways by regulating the expression of vital proteins including RB, Cbl-b, RUNX and so on, through miR-675-5p, which is transcribed from the first exon of H19 [19], [20], [21]. Considering the pivotal role of H19 and VDR signaling in the development of multiple types of cancer including colon cancer, we set out to investigate the potential correlation and the underlying mechanism between H19 and VDR signaling and the role of this correlation in the pathogenesis of colon cancer.
The effect of VDR signaling on the expression of H19 was investigated by treating colon cancer cells with increasing doses of 1,25(OH)2D3. The mechanism underlying the effect of 1,25(OH)2D3 was investigated by western-blotting, EMSA and ChiP assays. The correlation between the expression of H19 and VDR was further validated in colon cancer tissues and paired adjacent normal tissues collected from 24 colon cancer patients and 6 colon cancer cell lines. The effect of H19 overexpression on the expression level of VDR was investigated by Western-blotting and real-time PCR utilizing a H19 expressing plasmid. The role of the correlation between H19 and VDR signaling in the pathogenesis of colon cancer cells was then investigated by testing the sensitivity of colon cancer cells to the treatment of 1,25(OH)2D3 with or without H19 overexpression both in vitro and in vivo. The results suggested that H19 expression was inversely correlated with the expression of VDR in colon cancer tissues and colon cancer cell lines. VDR signaling was able to down-regulate the expression of H19 by inhibiting the C-Myc/Mad-1 network. H19, however, was able to down-regulate the expression of VDR by transcribing miR-675-5p. Colon cancer cells overexpressing H19 showed resistance to the treatment of 1,25(OH)2D3 both in vitro and in vivo, indicating the pivotal role of H19 in the development of resistance to vitamin D treatment in advanced colon cancer cells.
Taken together, these results suggest that H19 overexpression might be one of the mechanisms underlying the development of resistance to vitamin D treatment in advanced colon cancer cells and this may provide potential therapeutic targets for the treatment of colon cancer.
Materials and Methods
Patients and Samples
Between September 2010 and April 2013, 24 patients with colon cancer who underwent surgeries at Peking University First Hospital were included in this study. This study was approved by the institutional review board at Peking University First Hospital and conducted in accordance with the principles of the Declaration of Helsinki. After informed consent had been obtained, samples of colon cancer tissues and paired adjacent normal tissues were collected immediately after resection and stored in a −80°C environment until further analysis.
Cell Culture
HT-29 and DLD-1 cells were purchased from ATCC (American Type Culture Collection, USA) and maintained at 37°C in a culture medium composed of Dulbecco's modified Eagle's medium (DMEM) with 4.5 mg/ml glucose, 50 U/ml penicillin, 50 U/ml streptomycin, 25 mmol/l HEPES, and 10% fetal bovine serum (FBS) as previously described. 1,25(OH)2D3 (Sigma Aldrich, USA) diluted with ethanol was added every 24 h and equal quantities of ethanol were used as control.
Quantitative Real-Time PCR for Detection of H19 and VDR
Total RNA was extracted using the TRIzol one-step extraction method (TRIzol reagent; Invitrogen, USA) and reverse transcribed into cDNA using kit (Thermo Fisher, USA) according to the manufacturer's recommendations. Quantitative Real-time PCR analysis was performed in a 25 μl final reaction volume using the TaqMan® Universal PCR Master Mix (Applied Biosystems, USA) according to the manufacturer's recommendations. All reactions were run in triplicate with 7500 real-time PCR System (Applied Biosystems, USA). RNA relative expression was calculated as fold change using the comparative threshold cycle (CT) method (2−ΔΔCT) with GAPDH serving as the internal control gene. The primers and probes are listed 5′-3′ as follows: GAPDH: F, CAG TCA GCC GCA TCT TCT TTT; R, GTG ACC AGG CGC CCA ATA C; GAPDH probe: TAMRA-CGT CGC CAG CCG AGC CAC A-BHQ2; H19: F, AAT CGG CTC TGG AAG GTG AA; R, CTG CTG TTC CGA TGG TGT CTT; H19 probe, TAMRA-CTA GAG GAA CCA GAC CTC ATC AGC CCA AC-BHQ1; VDR: F, GGA GGC CTT GAA GGA CAG TCT; R, CTC CAC CAT CAT TCA CAC GAA; VDR probe: TAMRA-TAC TCC GAC TTC TGC CAG TTC CGG C-BHQ2.
Quantitative Real-Time PCR for Detection of miR-675-5p
MicroRNA was extracted using the miRNeasy Mini Kit (Qiagen, German) and reverse transcribed into cDNA using the kit (QuantiMir™ RT Kit, USA) according to the manufacturer's recommendations. Real-time PCR analysis was performed in a 25 μl final reaction volume using the TaqMan® Universal PCR Master Mix (Applied Biosystems, USA) according to the manufacturer's recommendations. All reactions were run in triplicate with 7500 real-time PCR System (Applied Biosystems, USA). RNA relative expression was calculated as fold change using the comparative threshold cycle (CT) method (2−ΔΔCT) with U6 serving as the internal control gene. The primers and probes are listed 5′-3′ as follows: miR-675-5p: F, ATG CTG TGG TGC GGA GAG G; R, TAT GGT TGT TCA CGA CTC CTT CAC; miR-675-5P probe: TAMRA-GTG TCA CCA GAC ATA CCA ACC TAT CCC-BHQ1; U6: F, ATT GGA ACG ATA CAG AGA AGATT; R, GGA ACG CTT CAC GAA TTT G; probe: TAMARA-TGC GCA AGG ATG ACA CGC A-BHQ1.
Western Blot Assay
Total protein was extracted using the method described previously with little modifications [22]. Nuclear protein was extracted using NucBuster™ protein extraction kit (Novagen, German) following the manufacturer's recommendations. The concentrations of protein were determined using BCA kit (ThermoFisher, USA). Then, the extracts containing equal quantities of protein (30 μg) were electrophoresed in 6% or 10% polyacrylamide gel. Subsequently, the separated proteins were transferred onto a PVDF membrane. The membrane was then blocked for 1 h (5% bovine serum albumin (BSA) in TBS-Tween 20 buffer) at room temperature and then incubated overnight at 4°C with rabbit anti-VDR monoclonal antibody (1:1000 dilution, Abcam, USA), rabbit anti-Mad-1 monoclonal antibody (1:100 dilution, Santa Cruz), rabbit anti-C-Myc monoclonal antibody (1:1000 dilution, CST, USA), rabbit anti-Histone H3 monoclonal antibody (1:1000 dilution, Abcam, USA) and rabbit anti-GAPDH monoclonal antibody (1:1000 dilution, CST, USA). The membranes were subsequently incubated at room temperature for 1 h with corresponding secondary antibodies (1:10,000 dilution, CST, USA) and blots were developed with ECL detection reagents (Millipore, USA). Images were collected utilizing Syngene GeneGenius gel imaging system (Syngene, UK) following the manufacturer's instructions.
Electrophoretic Mobility Shift Assay (EMSA)
Nuclear protein extracts were prepared using the NucBuster™ protein extraction kit (Novagen, German) following the manufacturer's recommendations. Briefly, 106 cells were washed with PBS and re-suspended in cytoplasmic extraction reagent and centrifuged for 5 min at 16,000×g. The supernatant (cytosolic extract) was removed and the pellet re-suspended in nuclear extraction reagent. Nuclear extracts were obtained by centrifugation at 16,000×g for 10 min. The nuclear protein extracts were kept frozen at −80°C at a concentration of 1 mg/ml until used. DNA-protein binding assays were carried out by DIG Gel Shift Kit, 2nd Generation (Roche, Germany). Double stranded complementary oligonucleotides containing the C-Myc binding sites were synthesized and end-labeled with DIG: 5′-CGA GCG CAG TGG CGC ATG GCT GTA ATC CCA-3′. Binding reactions were carried out at room temperature in binding buffer using 30 fmol DIG end-labeled target DNA and 5 mg of nuclear extract. Competition assays were performed by adding 125-fold excess of unlabeled probe before the labeled probe. Assays were electrophoresed onto native 8% polyacrylamide gels and transferred onto a positively charged nylon membrane. Transferred DNAs were cross-linked to the membrane at 1200 × 100 μJ/cm2 and detected using anti-digoxigenin-AP and the chemiluminescent substrate CSPD.
Chromatin Immunoprecipitation (ChiP) Assays
ChiP assays were performed using Pierce™ Magnetic ChiP Kit (ThermoFisher, USA) following the manufacturer's recommendations. The antibodies used were listed as follows: rabbit anti-C-Myc monoclonal antibody (CST, USA), rabbit anti-Mad-1 monoclonal antibody (Santa Cruz, USA) and mouse anti-VDR monoclonal antibody (Santa Cruz, USA). Real-time PCR was performed using human genomic DNA as the standard and normalizing the specific antibody signal to the input signal as described previously. The primers designed according to the sequence of the promoter of H19 containing the E-box used for real-time PCR are listed 5′-3′as follows: F, CCC ACA TGC CAC GGA ATC; R, TCC TCT CAT CTC CCC AAC CTT.
Transfection of DNA Constructs and/or RNA Oligonucleotides
Transfection was performed with Lipofectamine 3000 (Life Invitrogen, USA) following the manufacturer's instructions. Cells were cultured on 12-well plates (Corning, USA). 1 μg of DNA construct (H19 or empty vector, GenePharma, China) and/or 20 pmol RNA oligonucleotides (miR-675-5p mimics or inhibitors, GenePharma, China) or 1.5 μl Lipofectamine 3000 were pre-incubated in 25 μl of Opti-MEM, respectively. Then, two solutions were mixed and incubated for 5 min at room temperature and the mixture was added to each well. After incubation for 3 h at 37°C, 1 ml of DMEM containing 10% FBS and no antibiotics was added to each well. Subsequently, media were replaced with normal growth media 24 h after transfection. The sequences of the micro RNA mimics, inhibitors and their corresponding negative controls are listed as follows: miR-675-5p mimics: 5′-UGGUG CGGAGAGGGCCCACAGUG-3′; the negative control for miR-675-5p mimics: 5′-UUCUCCGAACGUGUCACGU-3′; miR-75-5p inhibitors: 5′-CAC UGU GGG CCC UCU CCG CAC CA-3′; the negative control for miR-675-5p inhibitors: 5′-CAG UAC UUU UGU GUA GUA CAA-3′. The total protein was collected 72 h after transfection using the method as described above.
Luciferase Activity Assay
The part of VDR 3′-UTR containing the putative binding site and the 5′ and 3′ flanking sequence was amplified by a pair of primers (F: 5′-GAC GTT CTA GAT GAG TCA TGA TCT CCC TGC C-3′, R: 5′-GAC GTT CTA GAT ACC CTA CAT CAC GGA ACC C-3′) and subcloned into pGL3-control vector (Promega, USA) immediately downstream of the luciferase gene to form the pGL3-VDR-3′-UTR construct. Point mutations in seed sequence were generated by PCR to form the pGL3-VDR-3′-UTR-MUT construct. 1 × 105 cells were plated in 24-well plates for 24 h. 0.4 μg of pGL3 constructs plus 0.07 μg of pRL-CMV plasmid-expressing renilla luciferase (Promega) were transfected in combination with 60 pmol of either a negative control RNA oligonucleotide or miR-675 mimics/inhibitors (GenePharma, Shanghai, China) using Lipofectamine 3000 (Invitrogen). After 48 h, luciferase activity was measured using the Dual Luciferase Reporter Assay System (Promega, USA). Firefly luciferase activity was normalized to renilla luciferase activity for each transfected well. The results were obtained from three independent experiments and each one was performed in triplicate.
Generation of Stable Cell Lines
To establish cells stably overexpressing H19, HT-29 and DLD-1 cells were transfected with pcDNA 3.1 (+)-H19 (GenePharma, China) or pcDNA 3.1(+) empty vector (GenePharma, China) as control using Lipofectamine 3000 and cells were allowed to recover for 48 h. Cell were then incubated with medium containing G418 (Sigma Aldrich, USA) (1.8 mg/ml for HT-29 and 1 mg/ml for DLD-1) for at least 4 weeks. The expression level of H19 of clones obtained were investigated and the clones with highest expression level of H19 were used in the studies both in vitro and in vivo.
MTT Proliferation Assays
A total of 1000 cells stably expressing H19 or empty vector (mock) were seeded in each well of a 96-well plate and 100 nM 1,25(OH)2D3 was added every 24 h. After 5 days, the cells were incubated in 50 μl of 0.1 mg/ml solution of 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) (Sigma-Aldrich, USA) at 37°C for 4 h and then lysed in 150 μl of dimethyl sulfoxide (DMSO) at room temperature for 30 min. The absorbance in each well was measured at 580 nm with Synergy H2 microplate reader (Bio Tek Instruments, USA). Experiments were performed in triplicates and repeated at least three times.
Migration Assays
Cell migration assays were assessed with transwell chambers (8.0 μm pore size, Corning, USA). Briefly, 5 × 104 cells stably expressing H19 or empty vector in 200 μl DMEM medium without serum were added in the upper chamber, while 600 μl DMEM medium with 20% FBS was placed in the bottom of wells. 1,25(OH)2D3, at 100 nM, was added to the basolateral parts of the transwell system every 24 h and the plates were incubated in 5% CO2 atmosphere at 37°C for 48 h. The cells that migrated to the opposite side of the membrane were fixed and stained with hematoxylin and eosin and the number of invading cells was counted under the microscope (Olympus, Japan). Experiments were performed in triplicates and repeated three times.
TOP: FOP Flash Assays
The TOP Flash and FOP Flash plasmids used in this study were purchased from Millipore (USA) and the pRLSV40 plasmid (Promega, USA) containing the Renilla reniformis luciferase gene (RLuc) under the control of the SV40 virus promoter (kindly provided by Professor Bu in the Peking University First Hospital Central Laboratory) was used as control. The expressing plasmids pcDNA 3.1(+)-H19 and pcDNA 3.1(+) empty vector were purchased from Genepharma (China). Cells were transfected with 500 ng H19 or empty vector, 500 ng TOP Flash or FOP Flash vector, and 12 ng RLuc using Lipofectamine 3000.12 h after transfection, cells were incubated with indicated doses of 1,25(OH)2D3 or equal quantities of ethanol for 48 and the analysis of Luc and RLuc activities was performed using the Dual Luciferase Reporter Assay System (Promega, USA) and the Luc and Rluc activities were measured with Synergy H2 microplate reader (Bio Tek Instruments, USA). The Luc activity was then normalized to RLuc activity.
Immunofluorescence
Cells cultured on slides were treated as indicated above. Cellular localization of β-catenin was assessed by immunofluorescence as described previously [23]. Briefly, cells were rinsed with PBS and then fixed with acetone at −20°C for 5 min. Cells were then rinsed in PBS followed by blocking with 1% BSA for 2 h at room temperature. Subsequently, cells were incubated with 5 mg/mL mouse monoclonal anti-β-catenin (ThermoFisher, USA) overnight at 4°C. After being washed with PBS, filters were incubated with goat anti-mouse IgG conjugated to Alexa555 (Molecular Probes, USA) in 1% BSA for 1 h at room temperature. After being washed with PBS, cells were stained with DAPI. After being washed with PBS, cells were mounted using the Prolong Gold anti-fade reagent (Molecular Probes, USA) and stored at 4°C in dark until analyzed. The fluorescence was visualized under Fluoview 1000 confocal microscope (Olympus, Japan).
Tumorigenesis in Nude Mice
This study was performed following the guidelines of the China Laboratory Animal Management Committee and the study has been approved by the institute review board at Peking University First Hospital. Male nude mice (3 weeks old) were purchased from Vital River Inc. (Beijing, China) and raised in the containment unit of the Laboratory Animal Center at the Peking University First Hospital. The mice were allowed to adapt to the environment for 1 week before any treatment. DLD-1 cells (2 × 106 cells/mouse, 0.2 ml PBS) with stably transfection with H19 or empty vector were subcutaneously injected into the flank fat pads of 4-week-old male nude mice. 1,25(OH)2D3, diluted with propylene glycol (PG), was injected i.p. at 0.5 μg/kg body weight every twice day and equal quantities of PG were used as negative control. Tumor volume was monitored every 3 days with the method as described previously [25]. The mice were euthanized 3 weeks after injection and the tumors were removed and weighed.
Statistical Analysis
The results were expressed as mean ± standard error of the mean (SEM) and analyzed using a Student t tests for unpaired data and ANOVA to compare groups whenever required (GraphPad Prism version 5.0, CA). P < .05 was used to indicate statistical significance.
Results
1,25(OH)2D3 Down-Regulates the Expression Level of H19 in Colon Cancer Cell Lines
DLD-1 and HT-29 cells were incubated with increasing doses of 1,25(OH)2D3 (1, 10, 100 nM) for 48 h before the expression level of H19 was investigated by qRT-PCR. The results suggested that 1,25(OH)2D3 down-regulated the expression level of H19 in both DLD-1 and HT-29 cells significantly in a dose-dependent manner (P < .05) (Figure 1A). The results of Western blot suggested that 1,25(OH)2D3 increased VDR signaling in a dose-dependent manner, featured by increased level of VDR, which validated the increased VDR signaling elicited by 1,25(OH)2D3 (Figure 1B).
Figure 1.

1,25(OH)2D3 decreased the expression of H19 in a dose-dependent manner. A, The expression level of H19 after treatment with increasing doses of 1,25(OH)2D3 for 48 h. B, The level of VDR after treatment with increasing doses of 1,25(OH)2D3 for 48 h. Cells treated with equal quantities of ethanol served as control (*P < .05, vs control). All experiments were performed in triplicate and repeated at least three times. Results were expressed as mean ± SEM (n = 3).
VDR Signaling Inhibits the Expression of H19 by Regulating the C-Myc/mad-1 Network
Previous reports have illustrated the role of VDR as a master regulator of C-Myc/Mad-1 network in tongue cancer cells [24]. The expression of H19 has been reported to be regulated by the C-Myc/Mad-1network and the C-Myc binding E-box has also been validated in the promoter of H19 [26]. Taken these previous studies into consideration, the role of C-Myc/Mad-1 in the inhibitory effect of VDR signaling on the expression of H19 was further investigated. The results suggested that 1,25(OH)2D3 decreased the level of C-Myc and increased the nucleus level of Mad-1 in a dose-dependent manner, indicating a potential role of C-Myc/Mad-1 network underlying the effect of VDR signaling on the expression level of H19 (Figure 2, A and B). EMSA assays suggested that 1,25(OH)2D3 significantly inhibited the level of C-Myc conjugated with E-box sequence in the promoter region of H19 (Figure 2C). ChiP assays suggested that 1,25(OH)2D3 blocked the binding of C-Myc and increased the binding of Mad-1 with the E-box region in the promoter of the H19 gene in a dose-dependent manner (Figure 2D). These results suggested that VDR signaling inhibited the expression of H19 by regulating the C-Myc/Mad-1 network.
Figure 2.

VDR signaling inhibits the expression of H19 by regulating the C-Myc/Mad-1 network in colon cancer cells. A, The level of C-Myc after treatment with increasing doses of 1,25(OH)2D3 for 48 h. B, The nucleus level of Mad-1 after treatment with increasing doses of 1,25(OH)2D3 for 48 h. Cells treated with equal quantities of ethanol served as control. C, Nuclear protein of DLD-1 cells was collected 48 h after treatment with increasing doses of 1,25(OH)2D3 (0,1,10,100 nM) and EMSA assays were performed. 1,25(OH)2D3 inhibited the level of C-Myc conjugated with E-box sequence in the promoter region of H19 in a dose-dependent manner. D, ChiP and Re-ChiP assays were performed in DLD-1 cells after treatment with increasing doses of 1,25(OH)2D3 for 48 h as described in the section of materials and method. 1,25(OH)2D3 decreased the binding of C-Myc and increased the binding of Mad-1 to the E-box in the promoter of H19 as assessed by real-time PCR following CHiP assays. Re-ChiP assays were performed by immunoprecipitation with an anti-VDR antibody followed by immunoprecipitation of C-Myc or Mad-1 and the results suggested that 1,25(OH)2D3 decreased the association of C-Myc with VDR and increased the association of Mad-1 with VDR in a dose-dependent manner (*P < .05, vs control). All experiments were performed in triplicate and repeated at least three times. Results were expressed as mean ± SEM (n = 3).
The Correlation Between the Expression of VDR and H19 in Primary Colon Cancer Tissues and Colon Cancer Cell Lines
The expression level of VDR and H19 was investigated in 24 independent colon cancer tissues and their paired adjacent normal tissues and the results suggested that the expression of H19 was negatively correlated with the expression of VDR in colon cancer tissues (r = 0.3221, P < .05) (Figure 3A). The expression level of H19 was also found to be negatively correlated with the expression level of VDR in 6 colon cancer cell lines (r = 0.3782, P < .05) (Figure 3B). Together, these results validated the negative correlation between the expression of H19 and VDR signaling in colon cancer.
Figure 3.
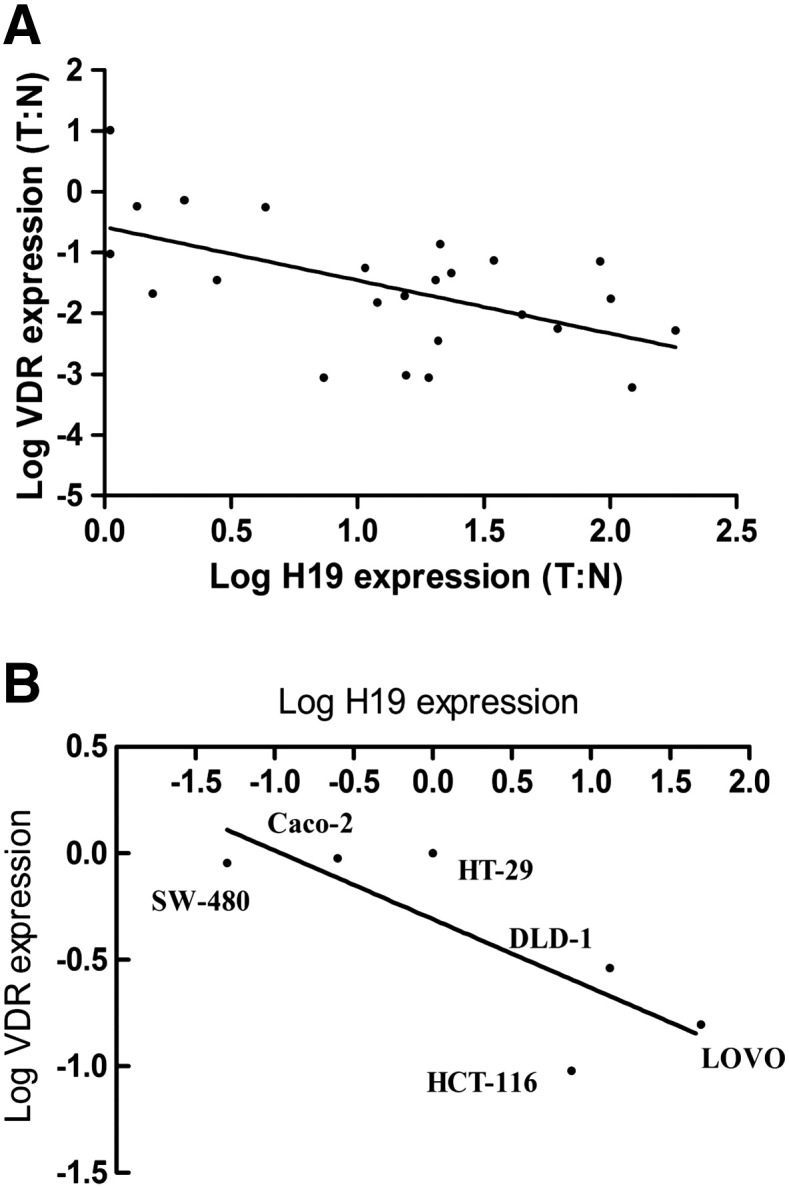
Negative correlation between the expression of VDR and H19 in primary colon cancer tissues and colon cancer cell lines. A, Negative correlation between the expression of VDR and H19 in primary colon cancer tissues (r = 0.3221, P < .05). B, Negative correlation between the expression of VDR and H19 in 6 colon cancer cells lines (r = 0.3221, P < .05).
H19 Overexpression Inhibited the Expression of VDR Through miR-675-5p
MiR-675-5p, transcribed from the first exon of H19, has been reported to mediate the function of H19 in multiple kinds of tissues. The above mentioned negative correlation between the expression of VDR and H19 bought the possible action of H19 on the expression of VDR into our attention. A potential binding site of miR-675-5p was identified in the 3′UTR of VDR mRNA utilizing the online prediction system “Targetscan” (Figure 4C). Transfection with H19 expressing plasmid significantly increased the level of H19 in both cell lines (Supplementary Figure 1). H19 overexpression significantly inhibited the expression of VDR in both cell lines (Figure 4A). This inhibitory effect was significantly although not totally abrogated by co-transfecting miR-675-5p inhibitors (Figure 4B). Transfection with miR-675-5p mimics significantly increased the level of miR-675-5p (Supplementary Figure 2) and down-regulated the expression of VDR in both cell lines (Figure 4D). Luciferase assays suggested that transfection with miR-675-5p mimics significantly decreased the relative luciferase activity of the pGL3-VDR-3′UTR construct in both cell lines, transfection with miR-675-5p inhibitors, however, significantly increased the luciferase activity in both cell lines (Figure 4E). These results suggested that H19 overexpression induced decreased expression of VDR through miR-675-5p.
Figure 4.

H19 overexpression decreased the expression of VDR through miR-675-5p. A, Total protein was collected 72 h after transfection with H19 expressing vector or empty vector (mock) and western blotting was performed as described in the section of materials and method. H19 overexpression significantly decreased the expression of VDR in both cell lines. B, Total protein was collected 72 h after transfection with H19 or empty vector (mock) and miR-675-5p inhibitors or negative control oligonucleotides and western blotting was performed as described in the section of materials and method. MiR-675-5p inhibitors significantly attenuated the inhibitory effect of H19 overexpression on the expression of VDR. C, Predicted binding site of miR-675-5p in the 3′UTR of VDR mRNA. D, Total protein was collected 72 h after transfection with miR-675-5p mimics or negative control oligonucleotides and western blotting was performed as described in the section of materials and method. MiR-675-5p mimics (MiR-675) significantly decreased the expression of VDR in both cell lines compared with negative control (Mock). E, pGL3-VDR-3’UTR construct or pGL3-VDT-3′UTR-MUT construct was transfected into both cell lines with either miR-675-5p mimics or inhibitors with negative control RNA oligo nucleotides relatively serving as control. Luciferase activity was collected 48 h after transfection respectively. MiR-675-5p mimics induced significantly decreased luciferase activity compared with control (P < .05). MiR-675-5p-inhibitors induced significantly increased luciferase activity compared with control (P < .05). Results were expressed as mean ± SEM (n = 3). (*P < .05, vs control).
H19 Overexpression Abrogated the Inhibitory Effect of 1,25(OH)2D3 on the Proliferation and Migration of Colon Cancer Cells
1,25(OH)2D3 has been reported to inhibit the proliferation and metastasis of colon cancers cells. However, colon cancer cells with advanced malignancy have been reported to be resistant to 1,25(OH)2D3. Based on the results mentioned above, the effect of H19 overexpression on the effect of VDR signaling on the colon cancer cells was further investigated. The results suggested that, H19 overexpression abrogated the inhibitory effect of 1,25(OH)2D3 on the proliferation and migration of the 1,25(OH)2D3 sensitive cells HT-29 and DLD-1 (Figure 5, A and B).
Figure 5.

H19 overexpression abrogated the inhibitory effect of 1,25(OH)2D3 on the proliferation, migration and the β-catenin/TCF transcriptional activity in colon cancer cells. A, 1,25(OH)2D3 significantly inhibited the proliferation of DLD-1 and HT-29 cells which were transfected with empty vector. The inhibitory effect didn't take place in cells transfected with H19. B, 1,25(OH)2D3 significantly inhibited the migration of DLD-1 and HT-29 cells transfected with empty vector featured by decreased cell numbers in transwell assays. The inhibitory effect didn't take place in cells transfected with H19. C, DLD-1 were transfected with either mock or H19 expressing vector and the transcriptional activities of β-catenin/TCF complexes were measured after transfection with either the TOP Flash or FOP Flash reporter vector and treatment with 100 nM 1,25(OH)2D3 or equal quantities of ethanol. H19 overexpression abrogated the inhibitory effect of 1,25(OH)2D3 on the transcriptional activity of β-catenin/TCF complexes in DLD-1 cells. D, Immunofluorescence of β-catenin in mock and H19 cells after treatment with either 100 nM 1,25(OH)2D3 or equal quantities of ethanol for 48 h. Nucleus were stained with DAPI. The redistribution of β-catenin from the nucleus to the plasma membrane induced by 1,25(OH)2D3 in mock cells did not take place in cells with H19 overexpression (*P < .05, vs control). All experiments were performed in triplicate and repeated at least three times. Results were expressed as mean ± SEM (n = 3).
The inhibition of Wnt/β-catenin pathway mediated by VDR signaling has been reported as one of the major mechanisms underlying the antineoplastic effect of 1,25(OH)2D3 on colon cancer cells. Thus, the transcriptional level of β-catenin was investigated in DLD-1 cells after treatment with 1,25(OH)2D3 with or without H19 overexpression utilizing TOP: FOP flash assays. The results suggested that H19 overexpression significantly abrogated the inhibitory effect of 1,25(OH)2D3 on the β-catenin/TCF transcriptional activity (Figure 5C). The results of immunofluorescence was also in accordance with the results of TOP: FOP flash assays, indicating that the redistribution of β-catenin from the nucleus to the plasma membrane induced by 1,25(OH)2D3 in mock cells did not take place in cells with H19 overexpression.
H19 Overexpression Induced Resistance to 1,25(OH)2D3 In Vivo
DLD-1 cells stably expressing H19 or empty vector were subcutaneously injected into nude mice. Mice were treated with indicated doses of 1,25(OH)2D3 or equal quantities of propylene glycol (PG) as control and tumor growth was monitored every 3 days. After 3 weeks, mice were euthanized and tumors were weighed. The results suggested that 1,25(OH)2D3 significantly inhibited the growth of tumor in DLD-1/vector transfected mice, which did not take place in DLD-1/H19 transfected mice (Figure 6A). No significant difference was observed between the weight of tumors collected from mice transfected with DLD-1/H19 plus treatment with 1,25(OH)2D3 and mice transfected with DLD-1/H19 treated with propylene glycol. The growth curve suggested that tumors grew faster in mice transfected with DLD-1/H19 compared with DLD-1/vector transfected mice. Furthermore, the growth curve suggested that tumors in mice transfected with DLD-1/H19 were resistant to the treatment with 1,25(OH)2D3 (Figure 6B). The results of qRT-PCR and western blot also validated the negative correlation between H19 and VDR signaling in vivo (Figure 6, C and D). Together, these results suggested that H19 overexpression induced resistance to the treatment of 1,25(OH)2D3 in DLD-1 cells in vivo.
Figure 6.

H19 overexpression induced resistance to the treatment with 1,25(OH)2D3 in vivo. A, 1,25(OH)2D3 significantly inhibited the proliferation of cells stably expressing empty vector. The inhibitory effect didn't take place in cells stably expressing H19. The weight of tumors of mice/H19 cells were significantly increased compared with tumors of mice/mock cells (*P < .05, vs Mock + PG). B, The tumor growth curves of DLD-1 cells with stable overexpression of vector or H19 and treatment with 1,25(OH)2D3 or PG showed the same results as cell proliferation assays in vitro. H19 overexpression abrogated the inhibitory effect of 1,25(OH)2D3 on the proliferation of DLD-1 cells. C, Total RNA of tumors of mice/mock cells treated with either 1,25(OH)2D3 or PG was collected and real-time PCR was performed to measure the expression level of H19. Treatment with 1,25(OH)2D3 significantly inhibited the expression level of H19 in vivo. D, Total protein of tumors of mice/mock and mice/H19 treated with PG was collected and western blotting was performed to investigate the expression of VDR. H19 overexpression significantly inhibited the expression of VDR in vivo (*P < .05, vs control). All experiments were performed in triplicate and repeated at least three times. Results were expressed as mean ± SEM (n = 3).
Discussion
H19 overexpression has been found to be related with the initiation and development of multiple kinds of cancer and miR-675-5p, transcribed from the first exon of H19, was found to be the main mechanism underlying the effect of H19 [27], [28]. Song et al. reported that there was more than 8 fold increased expression of H19 in gastric tumor tissues compared with paired normal tissues [29]. Increased expression of H19 has also been reported to be significantly related with early recurrence of bladder cancer and could be considered as a predictive marker for poor prognosis [14]. Recently, Zhou et al. reported that plasma H19 expression enabled the differentiation of early stage gastric cancer from controls with AUC of 0.877; sensitivity, 85.5%; specificity, 80.1% [30]. However, the mechanism underlying the role of H19 in the tumorigenesis of colon cancer remains to be elucidated.
Recently, lnc RNA profiling of VDR−/− mice revealed the potential correlation between H19 and VDR signaling [21]. Considering the emerging role of vitamin D as an important target for the development of novel therapy for colon cancer, investigating the correlation between H19 and VDR signaling and the underlying mechanisms may provide new insights into the development of therapeutic approaches for colon cancer. In the current study, the correlation between H19 and VDR signaling was investigated in colon cancer tissues and colon cancer cell lines and the mechanism and implication of this correlation was further investigated both in vitro and in vivo.
The results suggested that 1,25(OH)2D3 down-regulated the expression of H19 in a dose-dependent manner in both cell lines, indicating the inhibitory effect of VDR signaling on the expression of H19. The results suggested that 1,25(OH)2D3 blocked the binding of C-Myc and increased the binding of Mad-1 with the E-box region of the promoter of the H19 gene in a dose-dependent manner via VDR signaling. Together, these results indicate that VDR signaling is able to inhibit the expression of H19 through regulating C-Myc/Mad-1network.
H19 was found to be significantly increased in tumor tissues compared with normal tissues and VDR was found to be significantly decreased in tumor tissues compared with normal tissues and a negative correlation between the expression of H19 and VDR was observed in 24 independent colon cancer tissue samples. This negative correlation was also confirmed by the results collected from 6 colon cancer cell lines.
Considering this negative correlation and the plethora of proteins regulated by H19-miR-675-5p axis, the possible inhibitory effect of H19 overexpression on VDR was further investigated in this study. The results of western blot and luciferase assays indicated that H19 overexpression was able to induce decreased expression of VDR through miR-675-5p.
In the recent decades, plethora of studies have revealed the potential of 1,25(OH)2D3 and its analogs as a new therapy for multiple types of cancer including colorectal cancer [31], [32]. The mechanisms of the anti-neoplastic effect of 1,25(OH)2D3 can be very complex including inhibiting Wnt/β-catenin pathway, inhibiting the epithelial to mesenchymal transition (EMT) process, regulating the cell cycle and so on [33], [34], [35]. However, colon cancer cells at advanced stage often showed resistance to the treatment of 1,25(OH)2D3, due to the decreased expression of VDR with the development of cancer [36]. Currently, the mechanism underlying the decreased expression of VDR in advanced stage of colon cancer remains to be elucidated.
The results mentioned above in the current study indicated that H19 overexpression might be one of the mechanisms underlying the decreased expression of VDR in colon cancer cells and we set out to test the implication of this negative correlation between H19 and VDR signaling in the proliferation and migration of colon cancer cells both in vitro and in vivo. The results suggested that HT-29 and DLD-1 cells overexpressing H19 lost sensitivity to the treatment of 1,25(OH)2D3 compared with cells transfected with empty vector. 1,25(OH)2D3, at 100 nM, failed to inhibit the proliferation and migration of HT-29 and DLD-1 cells overexpressing H19. The results of TOP:FOP assays and immunofluorescence suggested that H19 overexpression abrogated the inhibitory effect of 1,25(OH)2D3 on the transcriptional activity and nuclear level of β-catenin, which might be one of the mechanisms underlying the development of increased tumorigenesis elicited by H19 overexpression.
The negative correlation between H19 and VDR signaling was further validated utilizing tumor xenograft model. The results suggested that 1,25(OH)2D3 significantly inhibited the proliferation and the expression level of H19 in DLD-1 cells stably expressing empty vector. DLD-1 cells stably expressing H19, however, showed resistance to the treatment of 1,25(OH)2D3 and the expression level of VDR was also significantly decreased in DLD-1cells overexpressing H19. These results were in accordance with the results collected in vitro, indicating that H19 overexpression might be an important mechanism underlying the development of resistance to the treatment of 1,25(OH)2D3 in the development of colon cancer.
In conclusion, this study illustrates the negative correlation between H19 and VDR signaling. VDR signaling was able to inhibit the expression of H19 by regulating the C-Myc/Mad-1 network. H19 overexpression, on the other hand, was able to down-regulate the expression of VDR through miR-675-5p. This correlation might play a vital role in the development of resistance to 1,25(OH)2D3 in the advanced stage of colon cancer and bring new insight into the development of potential therapeutic targets for colon cancer.
Conflict of Interest Statement
There is no conflict of interest.
Acknowledgements
We would like to thank Professor Yu Qi and Shen-shen Kong for their advice and technical assistance in the animal experiment.
Footnotes
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.neo.2016.10.007.
Contributor Information
Yucun Liu, Email: yucunliu@bjmu.edu.cn.
Pengyuan Wang, Email: wangpengyuan2014@126.com.
Appendix A. Supplementary data
Supplementary figures
References
- 1.Rebecca Siegel MPH, Ma J, Zou Z, Jemal A. Cancer statistics. CA Cancer J Clin. 2014;64:9–29. doi: 10.3322/caac.21208. [DOI] [PubMed] [Google Scholar]
- 2.Fearnhead NS, Wilding JL, Bodmer WF. Genetics of colorectal cancer: hereditary aspects and overview of colorectal tumorigenesis. Br Med Bull. 2002;64:27–43. doi: 10.1093/bmb/64.1.27. [DOI] [PubMed] [Google Scholar]
- 3.Wolpin BM, Mayer RJ. Systemic treatment of colorectal cancer. Gastroenterology. 2008;134:1296–1310. doi: 10.1053/j.gastro.2008.02.098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ponting CP, Oliver PL, Reik W. Evolution and functions of long noncoding RNAs. Cell. 2009;136:629–641. doi: 10.1016/j.cell.2009.02.006. [DOI] [PubMed] [Google Scholar]
- 5.Whitehead J, Pandey GK, Kanduri C. Regulation of the mammalian epigenome by long noncoding RNAs. Biochim Biophys Acta. 2009;1790:936–947. doi: 10.1016/j.bbagen.2008.10.007. [DOI] [PubMed] [Google Scholar]
- 6.Bartolomei MS, Zemel S, Tilghman SM. Parental imprinting of the mouse H19 gene. Nature. 1991;351:153–155. doi: 10.1038/351153a0. [DOI] [PubMed] [Google Scholar]
- 7.Tabano S, Colapietro P, Cetin I, Grati FR, Zanutto S, Mandò C, Antonazzo P, Pileri P, Rossella F, Larizza L. Epigenetic modulation of the IGF2/H19 imprinted domain in human embryonic and extra-embryonic compartments and its possible role in fetal growth restriction. Epigenetics. 2010;4:313–324. doi: 10.4161/epi.5.4.11637. [DOI] [PubMed] [Google Scholar]
- 8.Byun HM, Wong HL, Birnstein EA, Wolff EM, Liang G, Yang AS. Examination of IGF2 and H19 loss of imprinting in bladder cancer. Cancer Res. 2007;67:10753–10758. doi: 10.1158/0008-5472.CAN-07-0329. [DOI] [PubMed] [Google Scholar]
- 9.Berteaux N, Lottin S, Monté D, Pinte S, Quatannens B, Coll J, Hondermarck H, Curgy JJ, Dugimont T, Adriaenssens E. H19 mRNA-like noncoding RNA promotes breast cancer cell proliferation through positive control by E2F1. J Biol Chem. 2005;280:29625–29636. doi: 10.1074/jbc.M504033200. [DOI] [PubMed] [Google Scholar]
- 10.Kim SJ, Park SE, Lee C, Lee SY, Jo JH, Kim JM, Oh YK. Alterations in promoter usage and expression levels of insulin-like growth factor-II and H19 genes in cervical carcinoma exhibiting biallelic expression of IGF-II. Biochim Biophys Acta. 2002;1586:307–315. doi: 10.1016/s0925-4439(01)00109-0. [DOI] [PubMed] [Google Scholar]
- 11.Ariel I, Sughayer M, Fellig Y, Pizov G, Ayesh S, Podeh D, Libdeh B, Levy C, Birman T, Tykocinski M. The imprinted H19 gene is a marker of early recurrence in human bladder carcinoma. Mol Pathol. 2000;53:320–323. doi: 10.1136/mp.53.6.320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Li H, Yu B, Li J, Su L, Yan M, Zhu Z, Liu B. Overexpression of lncRNA H19 enhances carcinogenesis and metastasis of gastric cancer. Oncotarget. 2014;30:2318–2329. doi: 10.18632/oncotarget.1913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Feldman D, Krishnan AV, Swami S, Giovannucci E, Feldman BJ. The role of vitamin D in reducing cancer risk and progression. Nat Rev Cancer. 2014;14(5):342–357. doi: 10.1038/nrc3691. [DOI] [PubMed] [Google Scholar]
- 14.Pálmer HG, González-Sancho JM, Espada J, Berciano MT, Puig I, Baulida J, Quintanilla M, Cano A, de Herreros AG, Lafarga M. Vitamin D(3) promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of beta-catenin signaling. J Cell Biol. 2001;154(2):369–387. doi: 10.1083/jcb.200102028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kállay E, Bareis P, Bajna E, Kriwanek S, Bobber E, Toyokuni S, Cross HS. Vitamin D receptor activity and prevention of colonic hyperproliferation and oxidative stress. Food Chem Toxicol. 2002;40:1191–1196. doi: 10.1016/s0278-6915(02)00030-3. [DOI] [PubMed] [Google Scholar]
- 16.Pálmer HG, Larriba MJ, García JM, Ordóñez-Morán P, Peña C, Peiró S, Puig I, Rodríguez R, de la Fuente R, Bernad A. The transcription factor SNAIL represses vitamin D receptor expression and responsiveness in human colon cancer. Nat Med. 2004;10:917–919. doi: 10.1038/nm1095. [DOI] [PubMed] [Google Scholar]
- 17.Bhatia V, Falzon M. Restoration of the anti-proliferative and anti-migratory effects of 1,25-dihydroxyvitamin D by silibinin in vitamin D-resistant colon cancer cells. Cancer Lett. 2015;362(2):199–207. doi: 10.1016/j.canlet.2015.03.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jiang YJ, Bikle DD. LncRNA profiling reveals new mechanism for VDR protection against skin cancer formation. J Steroid Biochem Mol Biol. 2014;144(Pt A):87–90. doi: 10.1016/j.jsbmb.2013.11.018. [DOI] [PubMed] [Google Scholar]
- 19.Tsang WP, Ng EK, Ng SS, Jin H, Yu J, Sung JJ, Kwok TT. Oncofetal H19-derived miR-675 regulates tumor suppressor RB in human colorectal cancer. Carcinogenesis. 2010;31(3):350–358. doi: 10.1093/carcin/bgp181. [DOI] [PubMed] [Google Scholar]
- 20.Vennin C, Spruyt N, Dahmani F, Julien S, Bertucci F, Finetti P, Chassat T, Bourette RP, Le Bourhis X, Adriaenssens E. H19 non coding RNA-derived miR-675 enhances tumorigenesis and metastasis of breast cancer cells by downregulating c-Cbl and Cbl-b. Oncotarget. 2015;6(30):29209–29223. doi: 10.18632/oncotarget.4976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Liu G, Xiang T, Wu QF, Wang WX. Long Noncoding RNA H19-Derived miR-675 Enhances Proliferation and Invasion via RUNX1 in Gastric Cancer Cells. Oncol Res. 2016;23(3):99–107. doi: 10.3727/096504015X14496932933575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ma TY, Iwamoto GK, Hoa NT, Akotia V, Pedram A, Boivin MA, Said HM. TNF-alpha-induced increase in intestinal epithelial tight junction permeability requires NF-kappa B activation. Am J Physiol Gastrointest Liver Physiol. 2004;286(3):G367–G376. doi: 10.1152/ajpgi.00173.2003. [DOI] [PubMed] [Google Scholar]
- 23.Ye D, Guo S, Al-Sadi R, Ma TY. MicroRNA regulation of intestinal epithelial tight junction permeability. Gastroenterology. 2011;141(4):1323–1333. doi: 10.1053/j.gastro.2011.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Salehi-Tabar R, Nguyen-Yamamoto L, Tavera-Mendoza LE, Quail T, Dimitrov V, An BS, Glass L, Goltzman D, White JH. Vitamin D receptor as a master regulator of the c-MYC/MXD1 network. Proc Natl Acad Sci U S A. 2012;109(46):18827–18832. doi: 10.1073/pnas.1210037109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lagadec C, Meignan S, Adriaenssens E, Foveau B, Vanhecke E, Romon R, Toillon RA, Oxombre B, Hondermarck H, Le Bourhis X. TrkA overexpression enhances growth and metastasis of breast cancer cells. Oncogene. 2009;28:1960–1970. doi: 10.1038/onc.2009.61. [DOI] [PubMed] [Google Scholar]
- 26.Barsyte-Lovejoy D, Lau SK, Boutros PC, Khosravi F, Jurisica I, Andrulis IL, Tsao MS, Penn LZ. The c-Myc oncogene directly induces the H19 noncoding RNA by allele-specific binding to potentiate tumorigenesis. Cancer Res. 2006;66(10):5330–5337. doi: 10.1158/0008-5472.CAN-06-0037. [DOI] [PubMed] [Google Scholar]
- 27.Raveh E, Matouk IJ, Gilon M, Hochberg A. The H19 Long non-coding RNA in cancer initiation, progression and metastasis—a proposed unifying theory. Mol Cancer. 2015;14(1):184. doi: 10.1186/s12943-015-0458-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Matouk IJ, Halle D, Raveh E, Gilon M, Sorin V, Hochberg A. The role of the oncofetal H19 lncRNA in tumor metastasis: orchestrating the EMT-MET decision. Oncotarget. 2016;7(4):3748–3765. doi: 10.18632/oncotarget.6387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Song H, Sun W, Ye G, Ding X, Liu Z, Zhang S, Xia T, Xiao B, Xi Y, Guo J. Long non-coding RNA expression profile in human gastric cancer and its clinical significances. J Transl Med. 2013;11:225. doi: 10.1186/1479-5876-11-225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zhou X, Yin C, Dang Y, Ye F, Zhang G. Identification of the long non-coding RNA H19 in plasma as a novel biomarker for diagnosis of gastric cancer. Sci Rep. 2015;5:11516. doi: 10.1038/srep11516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Yang SW, Tsai CY, Pan YC, Yeh CN, Pang JH, Takano M, Kittaka A, Juang HH, Chen TC, Chiang KC. MART-10, a newly synthesized vitamin D analog, represses metastatic potential of head and neck squamous carcinoma cells. Drug Des Devel Ther. 2016;10:1995–2002. doi: 10.2147/DDDT.S107256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Dou R, Ng K, Giovannucci EL, Manson JE, Qian ZR, Ogino S. Vitamin D and colorectal cancer: molecular, epidemiological and clinical evidence. Br J Nutr. 2016;115(9):1643–1660. doi: 10.1017/S0007114516000696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Johnson AL, Zinser GM, Waltz SE. Vitamin D3-dependent VDR signaling delays ron-mediated breast tumorigenesis through suppression of β-catenin activity. Oncotarget. 2015;6(18):16304–16320. doi: 10.18632/oncotarget.4059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Chen S, Zhu J, Zuo S, Ma J, Zhang J, Chen G, Wang X, Pan Y, Liu Y, Wang P. 1,25(OH)2D3 attenuates TGF-β1/β2-induced increased migration and invasion via inhibiting epithelial-mesenchymal transition in colon cancer cells. Biochem Biophys Res Commun. 2015;468(1–2):130–135. doi: 10.1016/j.bbrc.2015.10.146. [DOI] [PubMed] [Google Scholar]
- 35.Jensen SS, Madsen MW, Lukas J, Binderup L, Bartek J. Inhibitory effects of 1alpha,25-dihydroxyvitamin D(3) on the G(1)-S phase-controlling machinery. Mol Endocrinol. 2001;15(8):1370–1380. doi: 10.1210/mend.15.8.0673. [DOI] [PubMed] [Google Scholar]
- 36.Giardina C, Madigan JP, Tierney CA, Brenner BM, Rosenberg DW. Vitamin D resistance and colon cancer prevention. Carcinogenesis. 2012;33(3):475–482. doi: 10.1093/carcin/bgr301. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary figures