

Abstract
Objectives
Rotator cuff tears are among the most frequent upper extremity injuries. Current treatment strategies do not address the poor quality of the muscle and tendon following chronic rotator cuff tears. Hypoxia-inducible factor-1 alpha (HIF-1α) is a transcription factor that activates many genes that are important in skeletal muscle regeneration. HIF-1α is inhibited under normal physiological conditions by the HIF prolyl 4-hydroxylases (PHDs). In this study, we used a pharmacological PHD inhibitor, GSK1120360A, to enhance the activity of HIF-1α following the repair of a chronic cuff tear, and measured muscle fibre contractility, fibrosis, gene expression, and enthesis mechanics.
Methods
Chronic supraspinatus tears were induced in adult rats, and repaired 28 days later. Rats received 0 mg/kg, 3 mg/kg, or 10 mg/kg GSK1120360A daily. Collagen content, contractility, fibre type distribution and size, the expression of genes involved in fibrosis, lipid accumulation, atrophy and inflammation, and the mechanical properties of the enthesis were then assessed two weeks following surgical repair.
Results
At two weeks following repair, treatment groups showed increased muscle mass but there was a 15% decrease in force production in the 10 mg/kg group from controls, and no difference between the 0 mg/kg and the 3 mg/kg groups. There was a decrease in the expression of several gene transcripts related to matrix accumulation and fibrosis, and a 50% decrease in collagen content in both treated groups compared with controls. Additionally, the expression of inflammatory genes was reduced in the treated groups compared with controls. Finally, PHD inhibition improved the maximum stress and displacement to failure in repaired tendons.
Conclusions
GSK1120360A resulted in improved enthesis mechanics with variable effects on muscle function. PHD inhibition may be beneficial for connective tissue injuries in which muscle atrophy has not occurred.
Cite this article: J. P. Gumucio, M. D. Flood, A. Bedi, H. F. Kramer, A. J. Russell, C. L. Mendias. Inhibition of prolyl 4-hydroxylase decreases muscle fibrosis following chronic rotator cuff tear. Bone Joint Res 2017;6:57–65. DOI: 10.1302/2046-3758.61.BJR-2016-0232.R1.
Keywords: Rotator cuff, Fibrosis, Myosteatosis
Article focus
We used a translationally relevant rat model to assess the ability of a pharmacological inhibitor of Hypoxia-inducible factor (HIF) prolyl 4-hydroxylases, GSK1120360A, to reduce fibrosis and improve healing following a rotator cuff tear.
Key messages
Inhibition of HIF prolyl 4-hydroxylases improved mechanical properties of the enthesis.
Inhibition of HIF prolyl 4-hydroxylases reduced fibrosis without improving contractile function of muscles.
Strengths and limitations
Strength: Detailed analysis of changes in muscle fibre contractile function and size, muscle extracellular matrix accumulation and gene expression, and enthesis mechanics.
Limitation: We only measured properties at a single time point and did not evaluate changes over time.
Introduction
Tears of the rotator cuff are one of the most frequent and debilitating joint injuries, and are often associated with fibrosis, muscle atrophy, and fat infiltration within and around muscle fibres, collectively referred to as ‘fatty degeneration’.1 Surgical repair of torn cuff muscles does not decrease fatty degeneration, and in some cases lipid infiltration and fibrosis continue to persist despite successful repair of the torn tendon.1 Fibrosis can increase the passive stiffness of the muscle extracellular matrix (ECM),2 and lead to increased shearing and damage of the muscle fibre sarcolemma.3 The combination of increased tissue stiffness and membrane damage can render the muscle more prone to further injury and re-tear, which complicates rehabilitation and prevents the full restoration of function.4 Identifying approaches that reduce fibrosis and promote muscle regeneration are therefore likely to speed up the recovery and improve the quality of life in patients who undergo surgical repair of chronically torn rotator cuff muscles.
Hypoxia-inducible factor-1α (HIF-1α) is a transcription factor that regulates the cellular and molecular responses to hypoxia, or low oxygen conditions. Under normoxic conditions, HIF-1α is hydroxylated on two proline residues by the HIF-prolyl 4-hydroxylases (PHDs), which then lead to the subsequent ubiquitination and rapid degradation of HIF-1α.5 In conditions of hypoxia, there is a reduction in the enzymatic activity of PHDs, which then allows HIF-1α to accumulate, translocate to the nucleus and direct the expression of target genes.5 HIF-1α induces the expression of several genes which have beneficial effects on the regeneration and protection of many organ systems including neural tissue, heart, lung, and skeletal muscle.6 In a skeletal muscle ischaemic injury model, compared with normoxic conditioned muscles, preconditioning of muscles in a hypoxic environment improved contractile force production, calcium influx, and recovery after an ischaemic insult.7 While hypoxia has been shown to positively affect the regeneration of several different types of tissues, inducing functional hypoxia in vivo can be difficult to achieve, which has led to the development of pharmacological inhibitors of PHD.6 In renal and cardiac injury models, treatment with PHD inhibitors reduced inflammation and fibrosis, and improved tissue regeneration.8,9 However, the effects of PHD inhibition on the treatment of skeletal muscle injuries are not known.
As chronically torn rotator cuff muscles often experience substantial inflammation and fibrosis, and the use of PHD inhibitors has shown promise in the amelioration of these conditions in other tissue injury models, our objective was to test the ability of a PHD inhibitor, GSK1120360A,9 to enhance rotator cuff regeneration following a chronic tear. We hypothesised that the targeted inhibition of PHD using GSK1120360A would decrease fibrosis, increase muscle fibre and specific force production, and increase tendon to bone strength in chronically torn rotator cuff muscles that underwent surgical repair. To test this hypothesis, we treated rats with GSK1120360A following the repair of chronically torn rotator cuff muscles, and measured changes in supraspinatus muscle collagen content, contractility, fibre type distribution and size, the expression of genes involved in fibrosis, lipid accumulation, atrophy and inflammation, and the mechanical properties of the enthesis two weeks following surgical repair.
Materials and Methods
Animals and sample size calculations
This study was approved by the University of Michigan Institutional Animal Care and Use Committee. Adult male retired breeder Sprague Dawley rats, approximately nine months of age, were used in this study. This age was selected as rats at this age display a healing response that is more similar to humans than younger rats, and male rats were selected in order to draw comparative conclusions with previous studies which have used this gender. Rats were randomly assigned to one of three groups: vehicle-treated controls; 3 mg/kg of GSK1120360A; or 10 mg/kg of GSK1120360A. These doses were selected based on pilot testing in muscle injury models in rats. To detect a 30% difference in muscle fibre-specific force between control and treatment groups, using a power of 0.80 and α = 0.05, required seven for each group. A total of ten rats were used in each group. One rat in the control group, two in the 3 mg/kg group and three in the 10 mg/kg group were lost to complications prior to completing the study.
Surgeries
Chronic full-thickness tears of bilateral supraspinatus muscles were performed as previously described.10,11 Rats were anaesthetised with a mixture of 2% isoflurane in oxygen and the surgical area was shaved and thoroughly scrubbed with chlorhexidine gluconate. A deltoid-splitting transacromial technique was used to reveal the supraspinatus tendon, which was then removed from its insertion on the humerus and placed in sterile silicone surgical tubing (PharMed BPT, Akron, Oklahoma) that was secured using a modified Mason-Allen stitch using 5-0 Ethibond suture (Ethicon, Somerville, New Jersey). The silicone tubing was used to prevent adhesion of the tendon end to the surrounding connective tissue. The deltoid was closed using 5-0 chromic gut (Ethicon) and the skin was closed with 4-0 Vicryl (Ethicon) and GLUture (Abbott Laboratories, Abbott Park, Illinois). Ampicillin (20 mg/kg, subcutaneous) was administered as a post-operative antibiotic, and buprenorphine (0.05 mg/kg) was given for analgesia during the post-operative period. Rats were closely monitored for signs of distress or infection.
At four weeks following the induction of supraspinatus tears, the torn tendons were repaired.10,11 Beginning three days before repair surgery, rats were given GSK1120360A via oral gavage in the described doses, and this was repeated daily until the conclusion of the experiment. Rats were anaesthetised and prepared as described above. The tendon was carefully liberated from the surgical tubing, and a modified Mason-Allen stitch using 5-0 two-arm Ethibond suture (Ethicon) was placed in the free tendon end. Bone tunnels in a crossing pattern were drilled into the humeral head using a 0.7 mm Kirschner-wire (gSource LLC, Emerson, New Jersey) 2 mm lateral to the articular surface at the anterior and posterior margins of the cuff footprint. Suture ends were passed through the tunnels and tied over the metaphyseal cortex to anatomically reattach the supraspinatus tendon to its native footprint. The deltoid and skin were closed in the same manner as the previous procedure, and post-operative antibiotics and analgesic medications were administered as described above. Free weight-bearing and cage activity were allowed, and rats were monitored for signs of distress or infection.
At two weeks after repair surgery, rats were anaesthetised with sodium pentobarbital (50 mg/kg), and supraspinatus muscles were harvested and weighed. This time point was selected as it reflects the intermediate stages of regeneration, when potential therapies are best able to be evaluated to determine accelerated healing.10 The proximal third of the left supraspinatus was prepared for single-fibre contractility. The left humerus was transected at the midshaft and the muscle-tendon-bone unit was used for measurement of tendon mechanical properties. The proximal two-thirds of the right supraspinatus was finely minced and prepared for biochemical measurements. The distal third was placed in Tissue-Tek (Sakura Finetek USA Inc., Torrance, California) and snap frozen in liquid nitrogen-cooled isopentane and prepared for immunohistochemistry. Rats were then humanely euthanised by anaesthetic overdose and induction of a bilateral pneumothorax.
GSK1120360A administration
GSK1120360A (N-{[1-(2-cyclopropylethyl)-6-fluoro-4-hydroxy-2-oxo-1, 2-dihydro-3-quinolinyl] carbonyl} glycine) was synthesised by GlaxoSmithKline (Research Triangle Park, Piedmont, North Carolina). GSK1120360A is a potent and specific inhibitor of the HIF PHD enzymes and shows little to no activity against the collagen PHD enzymes.9 The lyophilised drug was dissolved in 1% hydroxypropyl methylcellulose plus 0.1% Tween-80, and based on previous studies,9 administered to rats in doses of 0 mg/kg, 3 mg/kg, or 10 mg/kg via oral gavage. A total of 2 mL was administered daily to rats, starting three days before repair surgery, and continued until the day of tissue harvest.
Soluble protein content
Because swelling can occur in injured tissues, we measured the protein content of muscle tissues. Then 50 mg of finely minced muscle was placed in 500 µL of Total Protein Extraction Reagent (Thermo Fisher Scientific Inc., Rockford, Illinois) with 1:200 protease inhibitor cocktail (Thermo Fisher). Samples were homogenised, vortexed vigorously at 4º C, centrifuged at 10000 × g for ten minutes, and the supernatant was collected and stored at -80º C. Protein concentration was determined using a Bicinchoninic Acid protein assay (Thermo Fisher) and measured in a SpectraMax microplate reader (Molecular Devices, Sunnyvale, California). Relative protein concentration was determined by normalising the concentration from the BCA assay by the wet mass of muscle tissue.
Permeabilised fibre contractility
Single-fibre contractility was performed as previously described.10,12,13 The cross-sectional area (CSA) of fibres was measured, and the fibre was immersed in a high-calcium solution to yield maximum isometric force (Fo). To obtain specific force (sFo), Fo was normalised to CSA.
Histology
Muscle fibre type analysis was performed on supraspinatus muscles as described.10,12,13 Fibre type distribution and size measurements from immunohistochemical images were performed on ImageJ (USNIH, Bethesda, Maryland).
Hydroxyproline content
Hydroxyproline, which is a commonly used marker of collagen content, was measured in muscle tissue as previously described10,14 and normalised to the dry mass of the muscle tissue.
Gene expression
Measurements were performed as previously described.10,12,13 Total RNA was isolated from supraspinatus muscles using a miRNeasy Kit (Qiagen, Valencia, California), reverse transcribed to complementary DNA using the RT2 First Strand Kit (Qiagen) and amplified in a CFX96 Real-Time thermal cycler (Bio-Rad, Hercules, California). Primer sets were purchased from Qiagen. Expression of mRNA transcripts was normalised to the stable housekeeping gene β-actin, using the methods of Schmittgen and Livak.15 Statistical analysis, as described below, was performed on the 2-∆Ct values. A list of measured transcripts is provided in Table I.
Table I.
List of genes used for real-time polymerase chain reaction in this study
| Gene | RefSeq | Name | Function |
|---|---|---|---|
| Actb | NM_031144 | Actin, beta | Housekeeping |
| Agpat6 | NM_001047849 | 1-acylglycerol-3-phosphate O-acyltransferase 6 (GPAT4) | Lipid synthesis |
| Atg16l1 | NM_001108809 | ATG16 autophagy related 16-like 1 (S. cerevisiae) | Autophagy/protein breakdown |
| Atg5 | NM_001014250 | ATG5 autophagy related 5 homolog (S. cerevisiae) | Autophagy/protein breakdown |
| Becn1 | NM_053739 | Beclin 1, autophagy related | Autophagy/protein breakdown |
| Cd11b | NM_012711 | Integrin, alpha M | M1 macrophage marker |
| Cd68 | NM_001031638 | Cd68 molecule | Pan-macrophage marker |
| Cebpa | NM_012524 | CCAAT/enhancer binding protein (C/EBP), alpha | Adipogenic transcription factor |
| Col1a2 | NM_053356 | Collagen, type I, alpha 2 | Extracellular matrix |
| Dgat1 | NM_053437 | Diacylglycerol O-acyltransferase homolog 1 (mouse) | Lipid synthesis |
| Emr1 | NM_001007557 | EGF-like module containing, mucin-like, hormone receptor-like 1 (F4/80) | Pan-macrophage marker |
| Fbxo32 | NM_133521 | Atrogin-1 | Protein breakdown |
| Fsp1 | NM_012618 | S100 calcium-binding protein A4 | Fibroblast marker |
| Fsp27 | NM_001024333 | Cell death-inducing DFFA-like effector c | Lipid droplet formation |
| Cd168 | NM_012964 | Hyaluronan mediated motility receptor | M2 macrophage marker |
| Il10 | NM_012854 | Interleukin 10 | Anti-inflammatory cytokine |
| Il1b | NM_031512 | Interleukin 1 beta | Pro-inflammatory cytokine |
| Il6 | NM_012589 | Interleukin 6 | Pro-inflammatory cytokine |
| Mmp14 | NM_031056 | Matrix metallopeptidase 14 (membrane-inserted) | Matrix protease |
| Mmp2 | NM_031054 | Matrix metallopeptidase 2 | Matrix protease |
| Mmp9 | NM_031055 | Matrix metallopeptidase 9 | Matrix protease |
| Myh3 | NM_012604 | Myosin, heavy chain 3, skeletal muscle, embryonic | Muscle contractile protein |
| Pdgfra | NM_012802 | Platelet derived growth factor receptor, alpha polypeptide | Fibroblast marker |
| Plin | NM_013094 | Perilipin 1 | Lipid droplet protein |
| Plin5 | NM_001134637 | Perilipin 5 | Lipid droplet protein |
| Atgl | NM_001108509 | Adipose triglyceride lipase | Lipid breakdown |
| Pparg | NM_013124 | Peroxisome proliferator-activated receptor gamma | Adipogenic transcription factor |
| Fat/Cd36 | XM_575338 | Similar to fatty acid translocase/CD36 | Fatty acid transporter |
| Scx | NM_001130508 | Scleraxis | Fibroblast transcription factor |
| Timp1 | NM_053819 | TIMP metallopeptidase inhibitor 1 | Inhibitor of matrix proteases |
| Timp2 | NM_021989 | TIMP metallopeptidase inhibitor 2 | Inhibitor of matrix proteases |
| Tnmd | NM_022290 | Tenomodulin | Marker of mature tenocytes/fibroblasts |
| Trim63 | NM_080903 | MuRF-1, Tripartite motif-containing 63 | Protein breakdown |
| Vegfa | NM_031836 | Vascular endothelial growth factor A | Angiogenesis |
| Vps34 | NM_022958 | Phosphoinositide-3-kinase, class 3 | Autophagy/protein breakdown |
Biomechanical testing
Testing of the repaired enthesis interface was performed using a modified ElectroForce ELF3200 uniaxial testing system (Bose Corp., Eden Prairie, Minnesota) equipped with a 100 N load cell (Omega Engineering Inc., Stamford, Connecticut), as previously described.10 The tendon was preloaded to 1.0 N, and two cameras (JAI CM-200 MCL; JAI Inc., San Jose, California) mounted 90º apart captured the front and side views of the tendon, and was then used to calculate tendon CSA by fitting width and length measurements to an ellipse. The specimen was then stretched until failure at a rate of 10 mm/sec, and the maximum displacement to yield, maximum load to yield, peak stress and peak stiffness were determined from testing.
Statistical analysis
Data are presented as mean and standard error and were analysed in a blinded fashion. Differences between groups were tested using a one-way analysis of variance (ANOVA) (α = 0.05) followed by Fisher’s Least Significant Difference (LSD) post hoc sorting. All analysis was performed in Prism 6.0 (GraphPad, La Jolla, California).
Results
At two weeks following repair, muscle mass in the 3 mg/kg group was 18% greater than in controls, and 12% greater in the 10 mg/kg group compared with controls (Fig. 1a) but there was no dose-dependent increase in muscle mass. There were no changes between groups for relative protein content (Fig. 1b). There were few changes in muscle fibre type distribution between groups, with the exception of the percentage of hybrid type fast IIB/IIX fibres, which exhibited a greater than 50% increase in frequency in the 10 mg/kg group compared with the 0 mg/kg and 3 mg/kg groups (Fig. 1c). There were also very few differences in muscle fibre CSA between groups (Fig. 1d), with a 53% increase in type IIX fast fibre CSA in the 10 mg/kg group compared with the 0 mg/kg group, and no differences were observed between the two treatment groups.
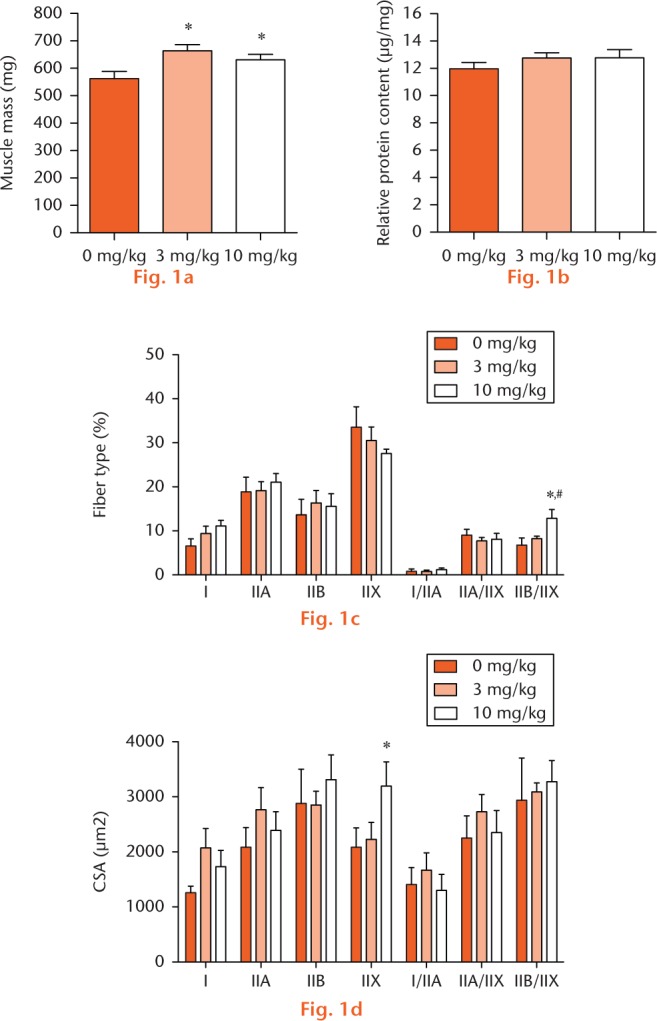
Muscle mass, protein content and fibre type distribution and size: (a) muscle mass of supraspinatus muscles, (b) relative protein content of supraspinatus muscles, (c) muscle fibre type distribution and (d) muscle fibre type cross-sectional area (CSA) from 0 mg/kg, 3 mg/kg, and 10 mg/kg groups. Values are mean and standard error. Differences between groups were tested using a one-way analysis of variance (α = 0.05) followed by Fisher’s Least Significant Difference post hoc sorting. *, significantly different from 0 mg/kg (p < 0.05).
For fibre contractility, there were no significant differences between muscle fibre CSA (Fig. 2a). However, there was a 25% decrease in Fo for the 10 mg/kg group compared with controls (Fig. 2b), but no differences between the 3 mg/kg group and controls (Fig. 2b). Additionally, there was no differences in sFo between the 0 mg/kg and 3 mg/kg groups (Fig. 2c), but the 10 mg/kg group exhibited a 15% and 10% decrease in sFo compared with the 0 mg/kg and 3 mg/kg groups, respectively (Fig. 2c). With regards to muscle fibrosis, there was roughly a 50% reduction in hydroxyproline content in the treated groups compared with the 0 mg/kg group, but no dose-dependent decrease in fibrosis between treatment groups (Fig. 3a). Representative wheat germ agglutinin stained images, that identify the ECM, from each group are presented in Figure 3b.
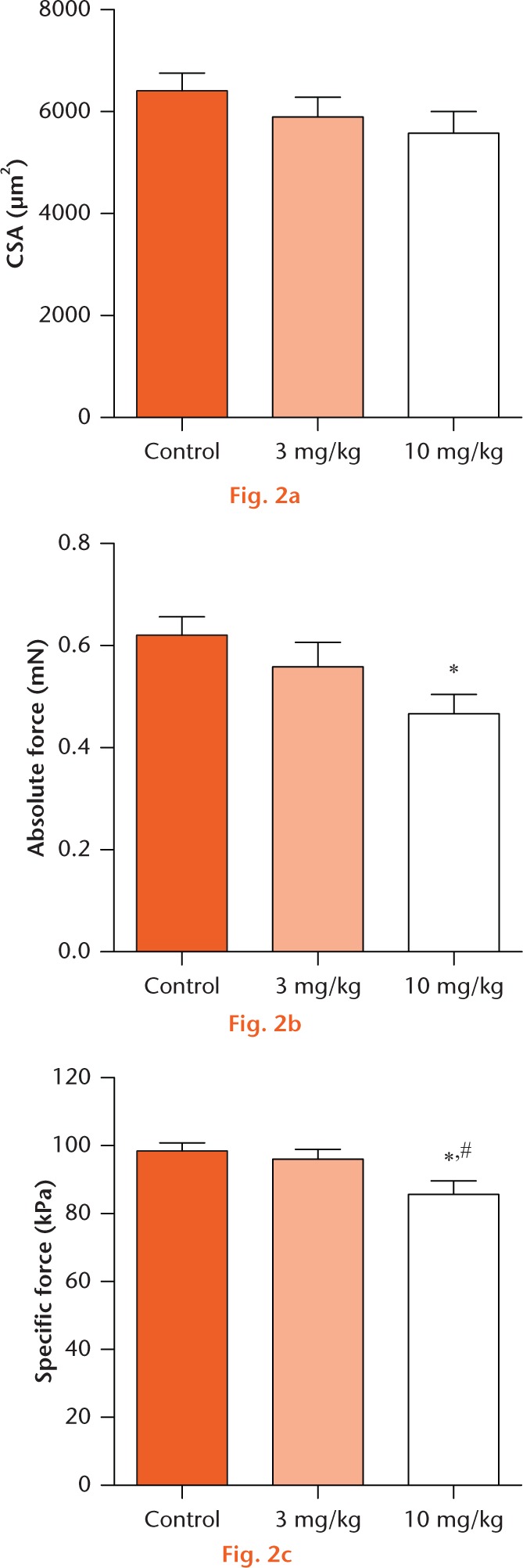
Permeabilised muscle fibre size and contractility: (a) permeabilised fibre cross-sectional area (CSA), (b) maximum isometric force (Fo) and (c) specific force (sFo) of 0 mg/kg, 3 mg/kg, and 10 mg/kg groups. Values are mean and standard error. Differences between groups were tested using a one-way analysis of variance (α = 0.05) followed by Fisher’s Least Significant Difference post hoc sorting. *, significantly different from 0 mg/kg; #, significantly different from 3 mg/kg (p < 0.05).

Hydroxyproline (Hyp) content and wheat germ agglutinin (WGA) stained muscle sections: (a) Hyp content from 0 mg/kg, 3 mg/kg, and 10 mg/kg groups. Values are mean and standard error. Differences between groups were tested using a one-way analysis of variance (α = 0.05) followed by Fisher’s Least Significant Difference post hoc sorting. *, significantly different from 0 mg/kg (p < 0.05). (b) Representative WGA stained images to identify extracellular matrix left to right from 0 mg/kg, 3 mg/kg, and 10 mg/kg groups (50mm).
There was a reduction in the expression of several genes involved in muscle fibrosis in the treated groups compared with controls (Fig. 4a), including collagen 1α2 (COL1α2), matrix metalloproteinase-2 (MMP2), matrix metalloproteinase-14 (MMP14), TIMP1, TIMP2, Tenomodulin, and fibroblast-specific protein-1 (FSP-1). For genes related to inflammation (Fig. 4b), there was a significant decrease in the pro-inflammatory interleukin IL-6 in the treated groups compared with controls. There was also an increase in the expression of the anti-inflammatory interleukin, IL-10, in the 10 mg/kg group compared with 0 mg/kg, but no differences from 3 mg/kg. There was a reduction in the pan-macrophage marker, CD68, in the 3 mg/kg group compared with controls, but no change in the 10 mg/kg group from other groups. Lastly, the anti-inflammatory macrophage marker, CD168, was elevated in the treated groups compared with controls.

Gene expression in muscles from 0 mg/kg, 3 mg/kg, and 10 mg/kg groups. Transcripts related to (a) extracellular matrix accumulation and maintenance, (b) inflammation and macrophage accumulation, (c) lipid synthesis and storage, and (d) muscle atrophy and autophagy. Values are mean and standard error. Differences between groups were tested using a one-way analysis of variance (α = 0.05) followed by Fisher’s Least Significant Difference post hoc sorting. *, significantly different from 0 mg/kg; #, significantly different from 3 mg/kg (p < 0.05).
For genes related to lipid accumulation and storage (Fig. 4c), there was a reduction in the expression of several genes in the treated groups compared with controls, including peroxisome proliferator-activated receptor gamma, CCAAT/enhancer binding protein alpha, perilipin 1, and fat-specific protein 27 (FSP27). Additionally, there was an induction in the expression of several genes in the treated groups compared with controls, including glycerol-3-phosphate acyltransferase 4 (GPAT4), diacylglycerol O-acyltransferase 1 (DGAT1), adipose triglyceride lipase, and perilipin 5. PHD inhibitor treatment also had an effect on the expression of genes related to muscle atrophy and remodelling (Fig. 4d). There was an increase in the expression of several genes in the treated groups compared with controls, including the E3 ubiquitin ligases, atrogin-1 and MuRF-1, vascular endothelial growth factor, and the autophagy-related transcripts, beclin-1, Vps34, and Atg16L1. The autophagy-related transcript Atg5 was also significantly reduced in the 10 mg/kg compared with the 3 mg/kg group, although both treated groups were upregulated compared with the 0 mg/kg group.
Finally, we measured the biomechanical properties of repaired shoulders. There was a reduction in tendon CSA in the 3 mg/kg group compared with the 0 mg/kg group, but no other significant changes in CSA (Fig. 5a). There were no changes in absolute load to failure (Fig. 5b), but displacement to failure was elevated in the treated groups compared with controls (Fig. 5c). There was a significant increase in peak stiffness for the 3 mg/kg group compared with the 0 mg/kg group but no differences between the 10 mg/kg group and 0 mg/kg group, nor were there any dose-dependent changes in stiffness between treatment groups (Fig. 5d). This was also the case for maximum stress to failure, where the 3 mg/kg group was increased from 0 mg/kg, and no other differences were measured (Fig. 5e).

Enthesis biomechanical failure testing values in 0 mg/kg, 3 mg/kg, and 10 mg/kg groups: (a) tendon cross-sectional area (CSA), (b) maximum load to failure, (c) maximum displacement to failure, (d) peak stiffness, and (e) maximum stress. Values are mean and standard error. Differences between groups were tested using a one-way analysis of variance (α = 0.05) followed by Fisher’s Least Significant Difference post hoc sorting. *, significantly different from 0 mg/kg (p < 0.05).
Discussion
Currently our ability to successfully treat chronic rotator cuff tears is limited by the poor quality of the muscle following repair. In particular, chronic inflammation and fibrosis play a large role in increasing the probability of failure of structurally successful repairs.1,2 Due to the lack of viable treatment options to treat muscle and tendon quality following chronic rotator cuff tears,1 and the potential for the activation of HIF-1α in augmenting muscle recovery,6 the use of PHD inhibitors holds attractive therapeutic potential to treat these injuries. In this study, the use of the PHD inhibitor, GSK1120360A, dramatically reduced fibrosis and inflammation within torn rotator cuff muscles, and increased the stiffness and stress to failure of the enthesis, with the 3 mg/kg dose overall demonstrating the most favourable changes.
Decreasing fibrosis in chronic tears of the rotator cuff has the potential to improve both functional recovery and patient quality of life, as fibrosis leads to muscle stiffness and a greater susceptibility to re-tear.2,4 In this study, following systemic administration of GSK1120360A, there was a 50% decrease in muscle hydroxyproline content after rotator cuff repair. This reduction in hydroxyproline content corresponded with lower levels of expression of type I collagen, MMP2, MMP14, and the fibroblast proliferation markers, tenomodulin and FSP-1. PHD inhibition was also associated with an improvement in mechanical properties of the enthesis. For both doses of GSK1120360A, there was an improvement in displacement to yield, a clinically important parameter for predicting long-term successful repair. The 3 mg/kg group also had an increase in peak stiffness and maximum stress to yield, suggesting an improvement in the organisation and quality of the tendon ECM. The inhibition of PHD may therefore improve the treatment of chronic rotator cuff tears not only by improving muscle recovery by reducing fibrosis, but also by promoting the development of a strong, mature enthesis.
Inflammation and lipid accumulation are associated with rotator cuff tears and are thought to contribute to impaired regeneration and recovery.1 Although the use of non-steroidal anti-inflammatory drugs or cox-2 inhibitor anti-inflammatory medications inhibits rotator cuff regeneration,16 we previously reported that the combined inhibition of the cyclooxygenase (COX)-1, COX-2 and 5-lipoxygenase pathways with licofelone decreased fibrosis, and increased load to failure of the enthesis.10 In the current study, inhibition of PHD reduced the expression of the pro-inflammatory cytokine IL-6 and increased the expression of the anti-inflammatory cytokine IL-10. Macrophages, which are important regulators of inflammation, also accumulate after rotator cuff tear,12,13 and PHD inhibition increased the expression of the anti-inflammatory M2 macrophage marker CD168. Increased inflammation in many models of skeletal muscle disease can also be caused by excess availability of lipid. In the case of type 2 diabetes mellitus, the accumulation of fatty acids and lipid intermediates within muscle fibres leads to an increase in the expression of several pro-inflammatory genes that have both local and systemic effects,17 and increasing the expression of triglyceride synthesis and storage enzymes reduces this inflammation.18 In the current study, the triglyceride storage enzymes, GPAT4 and DGAT1, were elevated in the PHD inhibitor-treated group. Additionally, perilipin 1 and FSP27, which are markers of pro-inflammatory lipid droplets and are highly upregulated following rotator cuff tear,12,13 were downregulated following PHD inhibition. PHD inhibition also resulted in an increase in perilipin 5 expression, which indicates lipid within the cell is likely being used for metabolic purposes rather than the synthesis of pro-inflammatory signalling molecules.19 Overall, these results indicate that PHD inhibition may be beneficial to long-term muscle recovery by reducing prolonged inflammation following chronic rotator cuff tears.
In addition to severe inflammation and fibrosis, chronic atrophy is a hallmark of rotator cuff tears. The two most common biochemical pathways that regulate muscle fibre atrophy are the ubiquitin-proteasome pathway and the autophagy pathway.20 For the ubiquitin-proteasome pathway, the E3 ubiquitin ligases atrogin-1 and MuRF-1 direct the polyubiquitination and subsequent degradation of damaged proteins through the 26S proteasome.20 Autophagy is a biochemical pathway in which organelles and proteins are targeted to the autolysosome for breakdown.20 There are several important components of the autophagy pathway, but Atg16L1, Atg5, beclin-1, and Vps34 are some of the key components of this process, and are commonly used as markers of autophagy.13,20 For both pathways, proteins are broken down into amino acids which are eventually returned to the cytosol to be used to synthesise new proteins.20 Although these processes are linked to atrophy and are thought to be detrimental to muscle function, they serve as important contributors to muscle amino acid availability and protein turnover. We and others have previously demonstrated that chronic rotator cuff tears are associated with increased expression of transcripts related to autophagy, but not E3 ubiquitin ligases atrogin-1 and MuRF-1, which are key rate-limiting enzymes in skeletal muscle protein degradation.13,21,22 In the current study, the E3 ubiquitin ligases atrogin-1 and MuRF-1, and the autophagy transcripts, Atg16L1, Atg5, beclin-1, and Vps34, were elevated in both groups that received the PHD inhibitor compared with control muscles. Despite the increase in expression of these transcripts, with the exception of a modest increase in type IIX muscle fibre CSA in the 10 mg/kg group, we did not observe changes in muscle fibre CSA or relative protein content with GSK1120360A treatment. HIF-1α can directly stimulate autophagy, which is thought to contribute to cell survival in hypoxia by reducing energy demands of the tissue.23 The relationship between HIF-1α accumulation and autophagy also likely influenced muscle fibre force production, with the 3 mg/kg dose of GSK1120360A demonstrating the ability to increase autophagy without decreasing force production, while the 10 mg/kg dose led to excessive autophagy that decreased force production. Combined, these results suggest that inhibition of PHD activity at the 3 mg/kg dose was able to reduce fibrosis and inflammation without negatively affecting muscle function or quality.
There are several limitations in this study. We use a rat model of chronic rotator cuff tear and repair due to similar anatomical features to human shoulders, but the rat model does not develop fatty degeneration to the extent observed in humans.13,24,25 A single time point of two weeks following repair was chosen to evaluate intermediate changes in regeneration, and did not assess the effect of PHD inhibition on muscle function and biochemical measures at different times following repair. We also did not evaluate an acute tear and repair model, which might be informative. Given that we did not use a genetic model of HIF-1α manipulation, we were unable to determine the effects of the inhibitor on tendon to bone healing versus direct muscle regeneration. For our single-fibre contractility measurements, we only measured the contractility of type II fibres. However, we demonstrate in this study and elsewhere13 that rat rotator cuff muscles contain primarily type II fibres, and there are no appreciable differences in sFo between type II isoforms.26 Despite these limitations, we believe that this study provided important insight into the role of PHD in the regeneration of chronically torn rotator cuff muscles following repair.
Fibrosis, lipid accumulation, and inflammation are debilitating to long-term functional recovery following rotator cuff tear. Current treatment strategies for patients with massive chronic tears do not reverse fatty degeneration, and persistent atrophy and fibrosis can increase the susceptibility of the repaired muscle to re-tear. Although PHD inhibition reduced fibrosis and inflammation, and improved mechanical properties of the bone-tendon interface, the treatment did not result in improvements in all of our functional measures and biomarkers. This study served as an important step in our understanding of the mechanisms behind muscle atrophy, inflammation, and fibrosis following rotator cuff tears. Although further studies are necessary, we identified HIF-1α as a potentially important pharmacological target for the treatment of chronic fibrotic and inflammatory muscle diseases, and it may be beneficial for acute connective tissue injuries in which muscle atrophy has not yet occurred.
Footnotes
Author Contribution: J. P. Gumucio: Study design, Data collection, Data analysis, Writing paper.
M. D. Flood: Data collection, Data analysis.
A. Bedi: Study design.
H. F. Kramer: Study design.
A. J. Russell: Study design, Writing the paper.
C. L. Mendias: Study design, Data analysis, Writing the paper.
ICMJE Conflicts of Intrest: H. F. Kramer and A. J. Russell are employees at, and stockholders of, GlaxoSmithKline, who own a patent on the drug reported in this work.
Funding Statement
The authors would like to acknowledge technical assistance from S. Roche and E. Lynch.
The study was funded by GlaxoSmithKline, and J. P. Gumucio received fellowship support from the National Institutes of Health (F31-AR065931). A. Bedi reports funding received from Arthrex, SLACK, and Springer which is not related to this article.
References
- 1. Gladstone JN, Bishop JY, Lo IKY, Flatow EL. Fatty infiltration and atrophy of the rotator cuff do not improve after rotator cuff repair and correlate with poor functional outcome. Am J Sports Med 2007;35:719-728. [DOI] [PubMed] [Google Scholar]
- 2. Sato EJ, Killian ML, Choi AJ, et al. Skeletal muscle fibrosis and stiffness increase after rotator cuff tendon injury and neuromuscular compromise in a rat model. J Orthop Res 2014;32:1111-1116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Ramaswamy KS, Palmer ML, van der Meulen JH, et al. Lateral transmission of force is impaired in skeletal muscles of dystrophic mice and very old rats. J Physiol 2011;589:1195-1208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Lieber RL, Ward SR. Cellular mechanisms of tissue fibrosis. 4. Structural and functional consequences of skeletal muscle fibrosis. Am J Physiol Cell Physiol 2013;305:C241-C252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Myllyharju J. Prolyl 4-hydroxylases, key enzymes in the synthesis of collagens and regulation of the response to hypoxia, and their roles as treatment targets. Ann Med 2008;40:402-417. [DOI] [PubMed] [Google Scholar]
- 6. Bernhardt WM, Wiesener MS, Weidemann A, et al. Involvement of hypoxia-inducible transcription factors in polycystic kidney disease. Am J Pathol 2007;170:830-842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kohin S, Stary CM, Howlett RA, Hogan MC. Preconditioning improves function and recovery of single muscle fibers during severe hypoxia and reoxygenation. Am J Physiol Cell Physiol 2001;281:C142-C146. [DOI] [PubMed] [Google Scholar]
- 8. Kapitsinou PP, Jaffe J, Michael M, et al. Preischemic targeting of HIF prolyl hydroxylation inhibits fibrosis associated with acute kidney injury. Am J Physiol Renal Physiol 2012;302:F1172-F1179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Bao W, Qin P, Needle S, et al. Chronic inhibition of hypoxia-inducible factor prolyl 4-hydroxylase improves ventricular performance, remodeling, and vascularity after myocardial infarction in the rat. J Cardiovasc Pharmacol 2010;56:147-155. [DOI] [PubMed] [Google Scholar]
- 10. Oak NR, Gumucio JP, Flood MD, et al. Inhibition of 5-LOX, COX-1, and COX-2 increases tendon healing and reduces muscle fibrosis and lipid accumulation after rotator cuff repair. Am J Sports Med 2014;42:2860-2868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Davis ME, Stafford PL, Jergenson MJ, et al. Muscle fibers are injured at the time of acute and chronic rotator cuff repair. Clin Orthop Relat Res 2015;473:226-232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Gumucio JP, Korn MA, Saripalli AL, et al. Aging-associated exacerbation in fatty degeneration and infiltration after rotator cuff tear. J Shoulder Elbow Surg 2014;23:99-108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Gumucio JP, Davis ME, Bradley JR, et al. Rotator cuff tear reduces muscle fiber specific force production and induces macrophage accumulation and autophagy. J Orthop Res 2012;30:1963-1970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Mendias CL, Marcin JE, Calerdon DR, Faulkner JA. Contractile properties of EDL and soleus muscles of myostatin-deficient mice. J Appl Physiol (1985) 2006;101:898-905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc 2008;3:1101-1108. [DOI] [PubMed] [Google Scholar]
- 16. Cohen DB, Kawamura S, Ehteshami JR, Rodeo SA. Indomethacin and celecoxib impair rotator cuff tendon-to-bone healing. Am J Sports Med 2006;34:362-369. [DOI] [PubMed] [Google Scholar]
- 17. Yu C, Chen Y, Cline GW, et al. Mechanism by which fatty acids inhibit insulin activation of insulin receptor substrate-1 (IRS-1)-associated phosphatidylinositol 3-kinase activity in muscle. J Biol Chem 2002;277:50230-50236. [DOI] [PubMed] [Google Scholar]
- 18. Liu L, Zhang Y, Chen N, et al. Upregulation of myocellular DGAT1 augments triglyceride synthesis in skeletal muscle and protects against fat-induced insulin resistance. J Clin Invest 2007;117:1679-1689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Wang H, Sreenivasan U, Hu H, et al. Perilipin 5, a lipid droplet-associated protein, provides physical and metabolic linkage to mitochondria. J Lipid Res 2011;52:2159-2168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Sandri M. Signaling in muscle atrophy and hypertrophy. Physiology (Bethesda) 2008;23:160-170. [DOI] [PubMed] [Google Scholar]
- 21. Liu X, Joshi SK, Samagh SP, et al. Evaluation of Akt/mTOR activity in muscle atrophy after rotator cuff tears in a rat model. J Orthop Res 2012;30:1440-1446. [DOI] [PubMed] [Google Scholar]
- 22. Joshi SK, Kim HT, Feeley BT, Liu X. Differential ubiquitin-proteasome and autophagy signaling following rotator cuff tears and suprascapular nerve injury. J Orthop Res 2014;32:138-144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Mazure NM, Pouysségur J. Hypoxia-induced autophagy: cell death or cell survival? Curr Opin Cell Biol 2010;22:177-180. [DOI] [PubMed] [Google Scholar]
- 24. Soslowsky LJ, Carpenter JE, DeBano CM, et al. Development and use of an animal model for investigations on rotator cuff disease. J Shoulder Elbow Surg 1996;5:383-392. [DOI] [PubMed] [Google Scholar]
- 25. Liu X, Manzano G, Kim HT, Feeley BT. A rat model of massive rotator cuff tears. J Orthop Res 2011;29:588-595. [DOI] [PubMed] [Google Scholar]
- 26. Bottinelli R, Canepari M, Pellegrino MA, Reggiani C. Force-velocity properties of human skeletal muscle fibres: myosin heavy chain isoform and temperature dependence. J Physiol 1996;495:573-586. [DOI] [PMC free article] [PubMed] [Google Scholar]