

Abstract
Background:
It is well known that the naturally occurring modified triterpenes in plants have a wide diversity of chemical structures and biological functions. The lupane-, oleanane-, and ursane-type triterpenes are the three major members of natural triterpenes with a wide range of biological properties. A systematic approach is necessary to review their structures and biological activities according to the backbones and the different substituents.
Objective:
Thirty lupane-(L1-7), oleanane-(O1-14), and ursane-type (U1-9) triterpenes with structural diversity were examined to evaluate their cytotoxic activities against two cancer cell lines, human hepatocellular carcinoma (HepG2) and AGS cells.
Materials and Methods:
They were isolated from Hedera helix, Juglans sinensis, and Pulsatilla koreana using a series of column chromatography methods and were treated to evaluate their cytotoxic activities against HepG2 and AGS human gastric adenocarcinoma cell. Further, two triterpenes showing the most potent activities were subjected to the apoptotic screening assay using flow cytometry.
Results:
The polar groups, such as an oxo group at C-1, a free hydroxyl at C-2, C-3, or C-23, and a carboxylic moiety at C-28, as well as the type of backbone, explicitly increased the cytotoxic activity on two cancer cells. O5 and U5 showed significantly the potent cytotoxic activity in comparison to other glycosidic triterpenes. In annexin-V/propidium iodide (PI) staining assay, the percentage of late apoptosis (annexin-V+/PI+) 12 and 24 h after treatment with O5 and U5 at 25 μM increased from 14.5% to 93.1% and from 46.4% to 49.1%, respectively, in AGS cells. The cytotoxicity induced by O5 showed a significant difference compared to U5 for 12 and 24 h.
Conclusion:
In the study, we can suggest the potent moieties which influence their cytotoxic activities against two cancer cells. The polar groups at C-1, C-2, C-3, C-23, and C-28 and the linkage of sugar moieties influenced the different cytotoxic activities.
SUMMARY
Thirty naturally occurring oleanane-, ursane-, and lupane-type triterpenes were isolated from Hedera helix, Juglans sinensis, and Pulsatilla koreana
An oxo, a free hydroxyl, a carboxylic moiety, and the types of aglycone influenced the cell cytotoxicity
Corosolic acid and α-hederin showed the most potent cytotoxicity via apoptosis.
Keywords: Flow cytometry, lupane, oleanane, triterpene, ursane
INTRODUCTION
Triterpenes are a member of terpenes, which are structurally diverse class of natural compounds derived from the combination of C5 isoprene unitsand have been abundantly found in the plant kingdom.[1] They mainly exist in the cyclized form of squalene (30 carbons) which are acyclic hydrocarbons formed by two units of farnesyl diphosphate.[2] Triterpenes are precursors of steroids in both plants and animals which have a wide range of pharmacological properties, such as anti-inflammatory, anti-angiogenesis, anti-oxidative, pro-apoptotic, and re-differentiation effects.[3] Especially, lupane-, oleanane-, and ursane-type triterpenes showed the cytotoxicity against various cancer cell lines and have been considered as promising anti-cancer agents.[3,4] Their cytotoxic activity on cancer cells is significantly affected by the type of basic backbone and the present number and position of derivatives, such as alkyl, hydroxyl, carboxyl, and amino acids.[5,6] Among these, triterpene glycosides (i.e., triterpenoidal saponins) occurring by the combination of sugar residues to basic backbone also have significant cytotoxic effects on cancer cells.[7,8]
To date, it has been mainly demonstrated the structure–activity relationship between synthesized triterpene derivatives on cancer cells.[9,10,11] We evaluated the cytotoxic activities of naturally occurring lupane-, oleanane-, and ursane-type triterpenes isolated from Juglans sinensis Dode (Juglandaceae),[12,13] Pulsatilla koreana Nakai (Ranunculaceae),[14] and Hedera helix L. (Araliaceae) against human hepatocellular carcinoma (HepG2) and AGS human gastric adenocarcinoma cell. We tried to suggest structurally some key determinants in the structure of triterpene derivatives on the cytotoxicity against cancer cell linesand also explore their preliminary mechanisms through inhibiting cell cycle and inducing apoptosis.
MATERIALS AND METHODS
General
Compounds L1-7, O1-14, and U1-9 were isolated from J. sinensis Dode (Juglandaceae) (L1, O1-4, O10-14, and U1-9),[12,13] P. koreana Nakai (Ranunculaceae) (L2-7, O6, and O9),[14] and H. helix L. (O5, O7, and O8) using repeated column chromatography and high-performance liquid chromatography. The structures of these compounds were unequivocally determined by diverse spectroscopic analyses, such as 1D, 2D nuclear magnetic resonance (NMR) experiments, mass spectrometry analyses, as well as by comparison with the literature for the known compounds. The 1H and 13C NMR 1H-1H correlation spectroscopy, Heteronuclear single quantum coherence spectroscopy (HSQC), and heteronuclear multiple bond correlation spectra were recorded on a Bruker AMX 400 (Bruker BioSpin GmbH, Karlsruhe, Germany) or 500 (Bruker BioSpin GmbH, Karlsruhe, Germany) spectrometer in pyridine-d5. High- and low-resolution FABMS (Fast Atom Bombardment Mass Spectrometry) were obtained on a JEOL JMS-AX505WA. Detection of DNA cycle and apoptosis on hepatic stellate cells-T6 cells was conducted by flow cytometry (BD Biosciences, FACSCalibur, Franklin Lakes, NJ, USA).
Cell lines
HepG2 and AGS (human gastric adenoma carcinoma cells) cell lines were purchased from ATCC (Manassas, VA, USA). HepG2 and AGS cells were maintained in Dulbecco's modified eagle medium and Roswell Park Memorial Institute 1640 medium, respectively, supplemented with 10% (v/v) fetal bovine serum, penicillin (100 U/mL), and streptomycin (100 μg/mL). They were incubated in a humidified atmosphere of 5% CO2 gas at 37°C.
Cytotoxicity assay
The cytotoxicity of L1-7, O1-14, and U1-9 against two cancer cell lines was evaluated by modified 3-(4,5-dimethylthiazol-2yl)-2,5- diphenyltetrazolium bromide (MTT) assay.[15] All samples were dissolved in dimethyl sulfoxide (final concentration, 0.1%) and diluted in distilled water. Cells were seeded in 96-well plates at a density of 3 × 104 cells/well for the cytotoxicity assay. After 24 h incubation, serum-free medium was changed and two cell lines were treated with vehicle or L1-7, O1-14, and U1-9 at the concentration of 100 μM for 24 or 48 h and incubated with 2 mg/mL of MTT for 2 h. Reduction of MTT to formazan was assessed in an ELISA plate reader at 540 nm. Doxorubicin (Sigma-Aldrich, St. Louis, MO, USA) was used as a positive control. Cell viability rate was calculated as the percentage of MTT absorption as follows: % survival = (mean experimental absorbance/mean control absorbance × 100). Data were expressed as the mean of three independent experiments.
Flow cytometry for analyzing apoptosis and caspase-3 activity
AGS cells were seeded in 6-well plates at a density of 6 × 105 cells/well for measuring the DNA cycle, apoptosis, and caspase-3 using flow cytometry. After 24 h, cells were treated with O5 and U5 at 10 and 25 μM, respectively. After 12 or 24 h, cells were trypsinized, washed twice with PBS buffer, and centrifuged at room temperature. For evaluating the DNA cycle, cell pellets were suspended in ice-cold 70% ethanol for fixation of the stage of cycle at 4°C. After overnight, cells were centrifuged and resuspended in 500 μL of PI/RNASE staining buffer (BD Pharmingen, Franklin Lakes, NJ, USA), incubated at room temperature for 30 min, and analyzed. For measuring apoptosis using a fluorescein isothiocyanate (FITC)-Annexin V apoptosis detection kit (BD Pharmingen, Franklin Lakes, NJ, USA), cells were stained with annexin V and PI, subsequently, according to the manufacturer's manual, and measured by flow cytometry. The alteration of caspase-3 activity by O5 and U5 was carried out according to the manufacturer's manual (FITC Active Caspase-3 kit, BD Pharmingen, Franklin Lakes, NJ, USA). All experiments were performed by flow cytometry (BD Biosciences, FACSCalibur, Franklin Lakes, NJ, USA).
Statistical analysis
The evaluation of statistical significance was determined by one-way ANOVA test, with P < 0.001 and P < 0.01 considered to be statistically significant.
RESULTS AND DISCUSSION
Cytotoxic activities of 30 triterpenes against HepG2 and AGS cell lines
We evaluated the cytotoxicity of 30 triterpene derivatives including lupane-(L1-7), oleanane-(O1-14), and ursane-type (U1-9) against HepG2 and AGS cell lines by MTT cell viability assay in 24 and 48 h.[15] All triterpenes were isolated from J. sinensis Dode (Juglandaceae) (L1, O1-4, O10-14, and U1-9),[12,13] P. koreana Nakai (Ranunculaceae) (L2-7, O6, and O9),[14] and H. helix L. (O5O7and O8). Their names and structures are listed and provided in Table 1 and Figure 1, respectively.
Table 1.
The list of triterpenes L1.L7, O1.O14 and U1.U9

Figure 1.

(a-f) Structures of L1-L7, O1-O14, and U1-U9
Relationship of structures and activities of 30 triterpenes
The cytotoxic activity of L1-7, O1-14, and U1-9 at the concentration of 100 μM against HepG2 and AGS cells for 24 and 48 h was plotted in Figure 2. In lupane-type triterpenes (L1-7), L2 including an O-linkage of two glycosides at C-3, a carboxylic group at C-17, and a hydroxyl group at C-23 showed the most significant cytotoxic activity than other lupane-type triterpenes against two cancer cell lines. The substituents of sugar moieties at C-28 unambiguously decreased the cytotoxicity against cancer cells (L2 vs. L3-7). In addition, L1 with a methyl group instead of a carboxylic group at C-17 did not show the potent inhibitory activity compared to L2. These results supported that a free carboxylic acid instead of a methyl or sugar group at C-17 was essential for cytotoxic activity.[16] Similar cytotoxic results were also observed when oleanane-type triterpenes (O1-14) were treated on two cancer cells. The compounds with sugar moieties at C-28 (O3, O4, O7-9, and O12) showed no potent cytotoxic activities compared to those with a free carboxylic acid (O1, O2, O5, O10, O11, and O13). Interestingly, introduction of sugar moiety at C-3 did not influence the loss of cytotoxic activity (O5). The presence of a hydroxyl moiety at C-23 could significantly increase cell growth inhibition (O10 vs. O11). In terms of ursane-type triterpenes, the cytotoxic results of HepG2 and AGS cell lines treated with U1-9 generally showed more potent inhibition than lupane- and oleanane-type triterpenes on cell proliferation. In addition, some modifications, such as hydroxylation at C-2 (U3 vs. U5 and U4 vs. U6) or C-22 (U7 vs. U8), oxidation from hydroxyl to oxo at C-3 (U4 vs. U9), and glycosylation at C-28 (U4 vs. U7), did not significantly change the cell viability. However, the configuration of hydroxyl group at C-3 had an important influence on the anti-tumor activity. U1 and U2 which have α-hydroxyl group at C-3 showed no cytotoxic activity compared to others which have β-hydroxyl group (U3-9). On the basis of the above results, it was suggested that a free hydroxyl group at C-2, C-3, and C-23, and a free carboxylic group at C-28 were essential for the cell growth inhibition, and the addition of sugar moiety at C-3 or C-28 reduced the cytotoxic effect as the length of glycosides was increased. In comparison between oleanane- and ursane-type triterpenes having same modifications, ursane-type triterpenes had slightly more potent cytotoxicity than oleanane-type triterpenes (U3 vs. O1, U6 vs. O2, and U7 vs. O3) as reported in the literature.[17,18,19,20]
Figure 2.
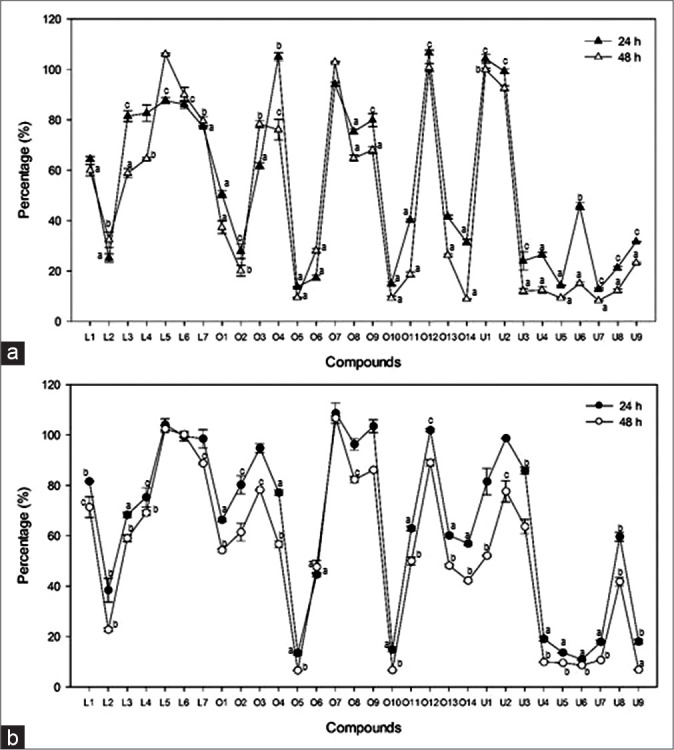
The cytotoxic activity of L1-7, O1-14, and U1-9 against HepG2 and AGS cell lines. Cytotoxic effect of L1-7, O1-14, and U1-9 were evaluated against AGS (a) and HepG2 (b) cells. The cells were treated with samples at the concentration of 100 μM for 24 or 48 h as indicated in the materials and methods. Each point represents a mean ± standard deviation of 3 experiments. Doxorubicin was used as a positive control and showed IC 50 for AGS and 3.7% (24 h) and 1.5% (48 h) for HepG2, respectively. a: P < 0.001, b: P < 0.05, and c: P < 0.01 compared with control
Apoptotic activities of O5 and U5 against HepG2 and AGS cell lines
We investigated the changes in progress of apoptosis induced by O5 (IC50 7.6 μM in AGS cells and 15.2 μM in HepG2 cells) and U5 (IC50 11.3 μM in AGS cells and 24.5 μM in HepG2 cells), which showed the most significant inhibitory activity against two cancer cells using the flow cytometry [Figure 3]. While the structure of U5 includes all the factors, such as ursane-type aglycone, a free hydroxyl group at C-2, C-3, and C-23, and a free carboxylic group at C-28, which essentially influence the cell viability against cancer cells in the cytotoxic assay, O5 as oleanane-type has not only a hydroxyl moiety at C-23 and a free carboxylic acid at C-28, but also two sugars at C-3. O5 and U5 at concentrations of 10 and 25 μM were treated in AGS cells for 12 and 24 h. The annexin-V/PI double staining results described that two compounds, O5 and U5, showed different aspects on the stage of apoptosis in AGS cells. When treated with 10 and 25 μM for 12 h incubation, U5 had much more potent cytotoxicity than O5 (Figure 3; b1 vs. d1 and c1 vs. e1). While O5 at 10 μM for 24 h did not induce the significant apoptosis compared to control group, the population of the apoptotic cells treated with O5 at 25 μM for 24 h in late apoptosis stage (annexin-V+/PI+) was dramatically increased to about 93.1% compared to U5 at the same concentration (49.1%). Our finding suggested that the delayed cytotoxicity of O5 might arise from the alteration of cell permeability by two glycosides at C-3 in structure. In the case of U5, the percentage of late apoptotic cells (annexin-V+/PI+) treated with 10 μM has doubled from 20.9% in 12 h to 43.6% in 24 h, while the number of late apoptotic cells induced by 25 μM showed no significant change as time passed (46.4% in 12 h and 49.1% in 24 h).
Figure 3.
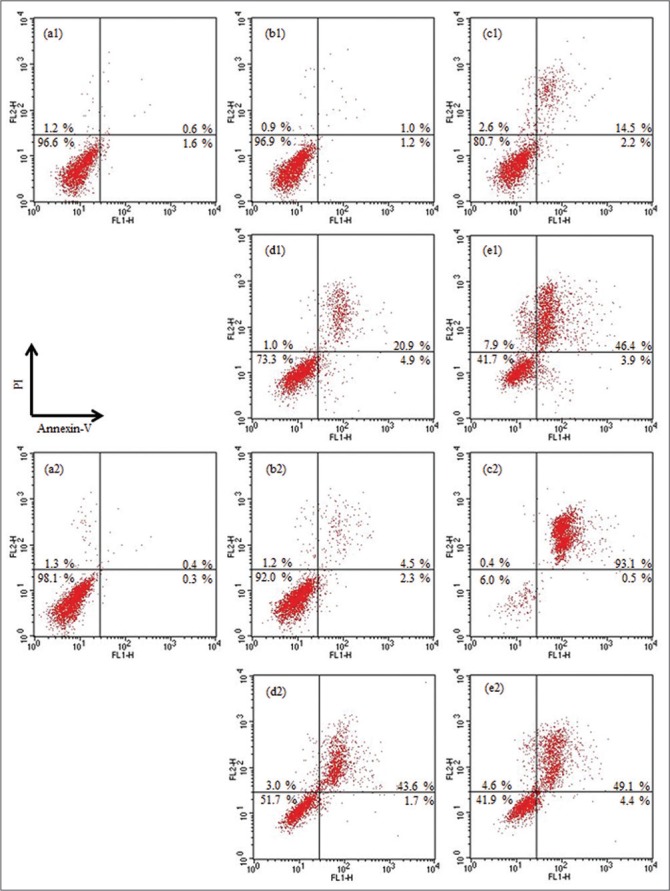
The flow cytometry analysis of the stage of O5 and U5-induced apoptosis in AGS cells. After 12 (a1-e1) or 24 h (a2-e2) incubated with O5 (b and c) and U5 (d and e) at the concentrations of 10 (b and d) and 25 (c and e) μM, respectively. (a1) and (a2) were the negative controls which were treated with the vehicle. Values are represented as percentage. Data are representative of three independent experiments
Changes of caspase-3 activity by O5 and U5 in AGS cell line
To further understand the mechanism of O5 and U5-induced apoptosis, we evaluated the caspase-3 activity that plays a pivotal role in the process of apoptosis in AGS cells.[21] O5 and U5 gradually increased the percentage of the caspase-3 activity in a concentration-dependent manner, when the cells were incubated with samples for 24 h [Figure 4]. The percentages of the activated caspase-3 treated with O5 and U5 compared to those of the control cells increased gradually with increasing the concentrations from 10 (O5; 41.1% and U5; 45.0%) to 25 μM (O5; 56.6% and U5; 55.2%). These results suggested that active caspase-3 played an important role in executing apoptosis by O5 and U5 in AGS cells.
Figure 4.

The flow cytometry analysis of O5 and U5-induced caspase-3 activity in AGS cells. The cells were incubated for 24 h with vehicle (control, white color), O5 or U5 at the indicated concentration (red color). Values are represented as percentage. Data are representative of three independent experiments
CONCLUSION
In the present study, we examined the cytotoxic activities of 30 lupane-, oleanane-, and ursane-type triterpenes against HepG2 and AGS cell lines. The types of backbones and simple structural modifications, such as hydroxylation and glycosylation, significantly influenced their biological activities. O5, an oleanolic acid derivative with hydroxyl group at C-23 and two sugar groups at C-3, and U5, an ursolic acid derivative with hydroxyl group at C-2, were the most potent apoptotic activities on the two cancer cell lines via caspase-3 regulation. Although further studies are needed to clarify the mechanism related with apoptosis by O5 and U5 on the inhibition of cancer cells proliferation, the structure–activity relationship of 30 triterpenes including O5 and U5 might give an idea for developing the therapeutic agents consisting of a triterpenes-concentrated preparation.
Financial support and sponsorship
This work was supported by the 2014 Research Grant from Kangwon National University (No. 120140653) and Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (No. NRF-2015R1C1A1A010553892).
Conflicts of interest
There are no conflicts of interest.
Acknowledgement
This work was supported by the 2014 Research Grant from Kangwon National University.
REFERENCES
- 1.Hill RA, Connolly JD. Triterpenoids. Nat Prod Rep. 2012;29:780–818. doi: 10.1039/c2np20027a. [DOI] [PubMed] [Google Scholar]
- 2.Dewick PD. Medicinal Natural Products-A Biosynthetic Approach. 3rd ed. Wiley: West Sussex; 2009. [Google Scholar]
- 3.Liu J. Oleanolic acid and ursolic acid: Research perspectives. J Ethnopharmacol. 2005;100:92–4. doi: 10.1016/j.jep.2005.05.024. [DOI] [PubMed] [Google Scholar]
- 4.Laszczyk MN. Pentacyclic triterpenes of the lupane, oleanane and ursane group as tools in cancer therapy. Planta Med. 2009;75:1549–60. doi: 10.1055/s-0029-1186102. [DOI] [PubMed] [Google Scholar]
- 5.Meng YQ, Liu D, Cai LL, Chen H, Cao B, Wang YZ. The synthesis of ursolic acid derivatives with cytotoxic activity and the investigation of their preliminary mechanism of action. Bioorg Med Chem. 2009;17:848–54. doi: 10.1016/j.bmc.2008.11.036. [DOI] [PubMed] [Google Scholar]
- 6.Ma CM, Cai SQ, Cui JR, Wang RQ, Tu PF, Hattori M, et al. The cytotoxic activity of ursolic acid derivatives. Eur J Med Chem. 2005;40:582–9. doi: 10.1016/j.ejmech.2005.01.001. [DOI] [PubMed] [Google Scholar]
- 7.Bachran C, Bachran S, Sutherland M, Bachran D, Fuchs H. Saponins in tumor therapy. Mini Rev Med Chem. 2008;8:575–84. doi: 10.2174/138955708784534445. [DOI] [PubMed] [Google Scholar]
- 8.Fuchs H, Bachran D, Panjideh H, Schellmann N, Weng A, Melzig MF, et al. Saponins as tool for improved targeted tumor therapies. Curr Drug Targets. 2009;10:140–51. doi: 10.2174/138945009787354584. [DOI] [PubMed] [Google Scholar]
- 9.Huang Z, Fu J, Liu L, Sun Y, Lai Y, Ji H, et al. Glycosylated diazeniumdiolate-based oleanolic acid derivatives: Synthesis, in vitro and in vivo biological evaluation as anti-HepG2 agents. Org Biomol Chem. 2012;10:3882–91. doi: 10.1039/c2ob25252j. [DOI] [PubMed] [Google Scholar]
- 10.Gao J, Li X, Gu G, Liu S, Cui M, Lou HX. Facile synthesis of triterpenoid saponins bearing ß-Glu/Gal-(1>3)-ß-GluA methyl ester and their cytotoxic activities. Bioorg Med Chem Lett. 2012;22:2396–400. doi: 10.1016/j.bmcl.2012.02.032. [DOI] [PubMed] [Google Scholar]
- 11.Chen L, Wu JB, Lei F, Qian S, Hai L, Wu Y. Synthesis and biological evaluation of oleanolic acid derivatives as antitumor agents. J Asian Nat Prod Res. 2012;14:355–63. doi: 10.1080/10286020.2011.654110. [DOI] [PubMed] [Google Scholar]
- 12.Yang H, Jeong EJ, Kim J, Sung SH, Kim YC. Antiproliferative triterpenes from the leaves and twigs of Juglans sinensis on HSC-T6 cells. J Nat Prod. 2011;74:751–6. doi: 10.1021/np1008202. [DOI] [PubMed] [Google Scholar]
- 13.Yang H, Cho HJ, Sim SH, Chung YK, Kim DD, Sung SH, et al. Cytotoxic terpenoids from Juglans sinensis leaves and twigs. Bioorg Med Chem Lett. 2012;22:2079–83. doi: 10.1016/j.bmcl.2012.01.010. [DOI] [PubMed] [Google Scholar]
- 14.Yang H, Cho YW, Kim SH, Kim YC, Sung SH. Triterpenoidal saponins of Pulsatilla koreana roots. Phytochemistry. 2010;71:1892–9. doi: 10.1016/j.phytochem.2010.07.013. [DOI] [PubMed] [Google Scholar]
- 15.Mosmann T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65:55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 16.Shao JW, Dai YC, Xue JP, Wang JC, Lin FP, Guo YH. In vitro and in vivo anticancer activity evaluation of ursolic acid derivatives. Eur J Med Chem. 2011;46:2652–61. doi: 10.1016/j.ejmech.2011.03.050. [DOI] [PubMed] [Google Scholar]
- 17.Li J, Guo WJ, Yang QY. Effects of ursolic acid and oleanolic acid on human colon carcinoma cell line HCT15. World J Gastroenterol. 2002;8:493–5. doi: 10.3748/wjg.v8.i3.493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yan SL, Huang CY, Wu ST, Yin MC. Oleanolic acid and ursolic acid induce apoptosis in four human liver cancer cell lines. Toxicol In Vitro. 2010;24:842–8. doi: 10.1016/j.tiv.2009.12.008. [DOI] [PubMed] [Google Scholar]
- 19.Shan JZ, Xuan YY, Ruan SQ, Sun M. Proliferation-inhibiting and apoptosis-inducing effects of ursolic acid and oleanolic acid on multi-drug resistance cancer cells in vitro. Chin J Integr Med. 2011;17:607–11. doi: 10.1007/s11655-011-0815-y. [DOI] [PubMed] [Google Scholar]
- 20.Lin CC, Huang CY, Mong MC, Chan CY, Yin MC. Antiangiogenic potential of three triterpenic acids in human liver cancer cells. J Agric Food Chem. 2011;59:755–62. doi: 10.1021/jf103904b. [DOI] [PubMed] [Google Scholar]
- 21.Pradelli LA, Bénéteau M, Ricci JE. Mitochondrial control of caspase-dependent and -independent cell death. Cell Mol Life Sci. 2010;67:1589–97. doi: 10.1007/s00018-010-0285-y. [DOI] [PMC free article] [PubMed] [Google Scholar]