

ABSTRACT
Libraries of defined mutants are valuable research tools but necessarily lack gene knockouts that are lethal under the conditions used in library construction. In this study, we augmented a Vibrio fischeri mutant library generated on a rich medium (LBS, which contains [per liter] 10 g of tryptone, 5 g of yeast extract, 20 g of NaCl, and 50 mM Tris [pH 7.5]) by selecting transposon insertion mutants on supplemented LBS and screening for those unable to grow on LBS. We isolated strains with insertions in alr, glr (murI), glmS, several heme biosynthesis genes, and ftsA, as well as a mutant disrupted 14 bp upstream of ftsQ. Mutants with insertions in ftsA or upstream of ftsQ were recovered by addition of Mg2+ to LBS, but their cell morphology and motility were affected. The ftsA mutant was more strongly affected and formed cells or chains of cells that appeared to wind back on themselves helically. Growth of mutants with insertions in glmS, alr, or glr was recovered with N-acetylglucosamine (NAG), d-alanine, or d-glutamate, respectively. We hypothesized that NAG, d-alanine, or d-glutamate might be available to V. fischeri in the Euprymna scolopes light organ; however, none of these mutants colonized the host effectively. In contrast, hemA and hemL mutants, which are auxotrophic for δ-aminolevulinate (ALA), colonized at wild-type levels, although mutants later in the heme biosynthetic pathway were severely impaired or unable to colonize. Our findings parallel observations that legume hosts provide Bradyrhizobium symbionts with ALA, but they contrast with virulence phenotypes of hemA mutants in some pathogens. The results further inform our understanding of the symbiotic light organ environment.
IMPORTANCE By supplementing a rich yeast-based medium, we were able to recover V. fischeri mutants with insertions in conditionally essential genes, and further characterization of these mutants provided new insights into this bacterium's symbiotic environment. Most notably, we show evidence that the squid host can provide V. fischeri with enough ALA to support its growth in the light organ, paralleling the finding that legumes provide Bradyrhizobium ALA in symbiotic nodules. Taken together, our results show how a simple method of augmenting already rich media can expand the reach and utility of defined mutant libraries.
KEYWORDS: Photobacterium, Aliivibrio, symbiosis, aminolevulinic acid, hemin, photobacteria
INTRODUCTION
Defined knockout mutant libraries are useful resources that have been generated to promote research in several bacterial experimental models and pathogens (1–10). In such studies, researchers have often listed essential genes, the knockouts of which were apparently lethal under the conditions of library construction and therefore not represented by mutants in the library. In order to recover insertion mutants collectively representing as many disrupted genes as possible, the bacteria targeted for mutation are generally grown on complex rich media, which makes many genes expendable. A popular choice for a growth medium has been LB (1–6, 9), which contains sodium chloride, tryptone as a rich source of peptides (10 g liter−1), and a complex array of vitamins and common metabolites from yeast extract (5 g liter−1). Despite its richness and complexity, LB needs further amendment to support growth of certain mutants, for example, those disrupted in the synthesis of certain bacterial cell wall components that are not found in yeast. Therefore, gaps in defined mutant libraries include some mutants that are missing because the respective gene is essential for growth on LB, although it could be nonessential in other media.
An effort is under way to generate a defined mutant library for Vibrio fischeri (11), a genetically tractable organism studied for its bioluminescence (12, 13), cell-cell signaling (14–16), interesting physiology (17, 18), biofilm formation (19–22), and light organ symbiosis with the Hawaiian bobtail squid Euprymna scolopes (23–26). Both this insertion mutant library, which is based on a mini-Tn5 transposon (27), and a recent complementary mariner-based InSeq study with V. fischeri (28) involve selection on LBS medium (29), which is a Tris-buffered high-NaCl derivative of LB (containing [per liter] 10 g of tryptone, 5 g of yeast extract, 20 g of NaCl, and 50 mM Tris [pH 7.5]). In this study, we sought to augment the suite of mutants being generated by screening for mutants able to grow on supplemented LBS (SLBS) but not on LBS. Moreover, we tested whether the host environment could support colonization by mutants that are unable to grow on LBS.
RESULTS AND DISCUSSION
To search for supplements that could reverse the lethality or strong negative effects of specific mutations during growth on LBS, we amended this medium with MgSO4, riboflavin, d-alanine, d-glutamate, diaminopimelic acid (DAP), N-acetylglucosamine (NAG), ribose, succinate, glycerol, biotin, NAD+, folate, thymidine, cytidine, uracil, and hemin. In addition, we added the iron chelator 2,2-bipyridyl, which stimulates the uptake of hemin by V. fischeri (30). The choices of supplements were based on precedent, genome annotations for V. fischeri, and conjecture. Although some of these compounds are present in LBS, we considered the possibility that larger amounts might be necessary to support growth of some mutants. For example, thymidine and riboflavin may be present in yeast extract, but supplementing LB with larger amounts of these compounds is necessary for robust growth of certain mutants of other bacteria (31, 32).
We isolated 25 mini-Tn5 transposon insertion mutants that grew on SLBS but not on LBS. The chromosomal location of the transposon insertion and the supplement requirement for each mutant are summarized in Table 1. We used the DOOR Database of Prokaryotic Operons (33) to assess which transposon insertions might occur in larger multigene transcripts, and these results are also listed in Table 1. Some of the mutants, for example, hemC mutants KP1 and SV1, are likely siblings, because they have insertions in identical locations and were isolated from the same mating to introduce the transposon. On the other hand, although three of the hemG mutants carry transposon insertions in the same location, only mutants AN2 and SV13 (and not EP1) were isolated from the same mating. In all, our screen yielded at least 22 independent mutants of ES114, with insertions corresponding to 13 different genes. Each of these genes was previously designated “putative essential” for growth on LBS in a recent InSeq analysis (28), although it should be noted that when assigning genes as putative essential, Brooks et al. (28) considered the possibility that insertions toward the 3′ end of genes might not eliminate function, whereas some of our insertion mutations may lead to partial gene function, as discussed below.
TABLE 1.
Mutants isolated in this study
| Strain(s) | Disrupted genea | Insertion after bp | Gene length (bp) | Putative functionb | Component that best restores growthc | Putative operon structured |
|---|---|---|---|---|---|---|
| RP5 | ftsA (VF_2197) | 894 | 1,263 | Cell division | Mg2+ or Ca2+ | ftsQAZ-lpxC (VF_2199–VF_2195) |
| SV11 | ftsQ (VF_2199) | −14 | 522 | Cell division | Mg2+ or Ca2+ | Tn-ftsQAZ-lpxC (VF_2199–VF_2195) |
| NL81 | glmS (VF_2372) | 686 | 1,830 | Glucosamine-6P synthesis | NAG or glucosamine | Monocistronic |
| AN4, DD1, RM1 | alr (VF_0302) | 246, 465, 134 | 1,080 | Alanine racemase | d-Ala | dnaB-alr-hyp (VF_0301–VF_0303) |
| AN3, SV8, SV14 | glr (murI) (VF_2433) | 154, 431, 431 | 786 | Glutamate racemase | d-Glu | Monocistronic |
| RP4 | hemA (VF_0767) | 894 | 1,257 | Glutamyl-tRNA reductase | Hemin or ALA | hemA-prfA-prmC-hyp-ychA-kdsA (VF_0767–VF_0772) |
| NL67, RP1 | hemL (VF_2135) | 475, 342 | 1,296 | ALA synthase | Hemin or ALA | Monocistronic |
| DD2, NP18, NP19 | hemB (VF_0053) | 880, 872, 322 | 1,017 | ALA dehydratase | Hemin | Monocistronic |
| KP1, SV1 | hemC (VF_0066) | 346, 346 | 936 | PBG deaminase | Hemin | hemCDXY (VF_0066–VF_0063) |
| DD3 | hemD (VF_0065) | 189 | 762 | Uroporphyrinogen III synthase | Hemin | hemCDXY (VF_0066–VF_0063) |
| AN6 | hemE (VF_2402) | 596 | 1,068 | Uroporphyrinogen III decarboxylase | Hemin | hemE-yhgI (VF_2402–VF_2401) |
| AN2, EP1, NL66, SV13 | hemG (VF_0028) | 245, 245, 238, 245 | 522 | Protoporphyrinogen oxidase | Hemin | yigZ-trkH-hemG (VF_0026–VF_0028) |
| AN5, RP3 | hemH (VF_0794) | 895, 329 | 969 | Ferrochelatase | Hemin | adk-hemH (VF_0793–VF_0794) |
The gene designation is followed by the ORF number (VF number) from the V. fischeri ES114 genome.
ALA, δ-aminolevulinic acid; PBG, porphobilinogen.
This column lists the component of SLBS that best restores growth in LBS, as well as other supplements that enable growth (see the text). The iron chelator 2,2-bipyridyl was required to stimulate uptake of hemin in these auxotrophs.
Operon composition predicted by the DOOR Database of Prokaryotic Operons (33). Underlined genes were categorized as “putative essential” (possibly due to polar effects) in an InSeq analysis (28). The disrupted gene is in bold, except in the case of SV11, where the Tn insertion is upstream of the operon (indicated by Tn-). “hyp” indicates “hypothetical” when no gene designation is provided by the genome annotation.
Five of the supplements in SLBS restored growth to particular mutants when added to LBS: MgSO4, NAG, d-alanine, d-glutamate, and hemin/bipyridyl. Representative mutants were characterized with respect to ability to colonize E. scolopes, and in some cases we further defined their auxotrophy and/or other defects.
Mutants rescued by magnesium.
Two mutants, RP5 and SV11, were rescued by adding MgSO4 to LBS (Table 1), and in each case MgCl2 also rescued growth, indicating that Mg2+ likely underlies this growth recovery. Because Mg2+ rescues the growth of certain V. fischeri mutants with multiple defects in lipid A biosynthesis (34), we speculate that Mg2+ bridging lipopolysaccharide (LPS) molecules structurally promotes outer membrane integrity and counteracts mutations or conditions that might compromise the outer membrane. Alternatively, dramatic regulatory effects of Mg2+ have been observed in V. fischeri (35–37), and Mg2+ may restore growth to mutants through a regulatory mechanism. Mutants RP5 and SV11 could also be rescued by supplementing LBS with 20 mM Ca2+, which has regulatory and structural effects distinct from but similar to those of Mg2+ (38, 39).
Unexpectedly, we recovered mutants with insertions in or near ftsA and ftsQ, which play a critical role in cell division in E. coli (40) and were previously designated essential in V. fischeri (28). Each of these mutants isolated in this study may retain partial function. The Tn insertion in SV11 is just upstream of ftsQ and does not disrupt the open reading frame (ORF) itself, while the insertion in ftsA leaves nearly the first three quarters of the protein intact (Table 1). It should also be noted that ftsQ (VF_2199) was originally misannotated as two genes (VF_2199 and VF_2198), which explains why its ORF number is two apart from that of ftsA (VF_2197) despite these being adjacent genes (41). The DOOR operon prediction database predicts that ftsQAZ is cotranscribed with lpxC; however, over 100 bp separate ftsZ from lpxC. We complemented mutant RP5 with ftsQAZ in trans on pJLS234, which restored growth on LBS without Mg2+, whereas the parent vector, pVSV105, did not (data not shown).
These two fts mutants were phenotypically similar to each other, with the ftsA mutant displaying more severe deviations from the wild type. Both grew detectably in LBS, but even with growth enhanced by Mg2+ supplementation, the mutants grew somewhat slower than the wild type. Growth of the mutants was likewise improved in SWTO medium (see Materials and Methods) with its marine salts, including Mg2+, although it still did not match that of the wild type. The fts mutants also had altered cell morphology (elongating and chaining) consistent with decreased or disrupted function of ftsQAZ (Fig. 1). The Tn-ftsQ mutant formed chains of cells (Fig. 1B) some reaching 100 μm or more in length (Fig. 1C). The ftsA::Tn mutant also formed chains (Fig. 1D and E), and these tended to become tangled into large aggregates of cells (Fig. 1F), some of which precipitated from culture. Interestingly, elongated cells or chains of cells often looped back and wound around themselves in a helical structure, a phenotype that was especially common in the ftsA::Tn mutant (Fig. 1D and E). A similar hairpin turn leading to a double helical structure was reported for an mreB mutant of the curved alphaproteobacterium Caulobacter crescentus (42). Wild-type V. fischeri forms slightly comma-shaped rods (43), but a spiral aspect to its cell shape has not been described, nor is it evident in our mutants except where cells wind around themselves. We speculate that the wild type may have a slight and nonrigid helical pitch to its comma-shaped cells, although it remains to be tested whether the fts mutations allow a natural cell shape to be seen or perhaps actually cause it in deviation from the wild type.
FIG 1.

Cell morphology of fts mutants. Phase-contrast images show wild-type strain ES114 (A), Tn-ftsQ mutant SV11 (B and C), and ftsA::Tn mutant RP5 (D, E, and F). Bars indicate size standards of 10 μm (A to E) and 100 μm (F). White arrows in panels D and E indicate where mutant cells or strings of cells have looped back on themselves. Cells were grown to mid-log phase in SWTO at 28°C with shaking at 200 rpm.
The Tn-ftsQ mutant was severely attenuated in colonizing the host (Fig. 2), and the ftsA::Tn mutant was difficult to test because the cells clumped so severely in the inoculum (e.g., Fig. 1F). In either case, a colonization defect could simply reflect a lack of motility. Motility is a key colonization factor for V. fischeri (44, 45), and nonmotile mutants are unable to colonize E. scolopes. Although microscopic examination of these mutants revealed a few swimming cells, the strains were virtually nonmotile in swim plate motility assays (data not shown). It seems likely that the fts mutants can be exploited to learn more about V. fischeri cell architecture, but they seem unlikely to inform our understanding of the light organ environment.
FIG 2.

Colonization of squid by representative mini-Tn5 insertion mutants. Squid were inoculated in parallel with either a representative mutant or ES114, and the CFU in each of the squid was determined 48 h later. Colonization by ES114 ranged from 0.7 × 105 to 3.8 × 105 CFU/squid across experiments, and mutant CFU/squid data were therefore normalized to those for the wild type and presented as the percentage of the wild-type CFU/squid in the same experiment. Each symbol indicates results (average CFU/squid for multiple squid) of an independent experiment. Each mutant was tested on at least 7 and no more than 19 squid in each experiment. Open boxes indicate the limit of detection in experiments where no mutant colonies were recovered from inoculated squid. Boxes indicate that mutant colonization and wild-type colonization were significantly different (P < 0.01) in a Mann-Whitney test, and circles indicate that the values for the mutant and wild type were not significantly different (P > 0.01).
Mutants affected in biosynthesis of peptidoglycan precursors.
Multiple independent mutants were isolated with insertions in the alr and glr (murI) genes, encoding alanine and glutamate racemases, respectively, and one mutant had an insertion in glmS, which encodes glucosamine-6-phosphate synthase (Table 1). These enzymes are required to make d-Ala, d-Glu, and NAG, respectively, which are key components of peptidoglycan. In Gram-negative bacteria, these genes often are essential unless the medium is appropriately supplemented (46, 47). Accordingly, alr, glr (murI), and glmS mutants required the d-Ala, d-Glu, or NAG supplements from SLBS to grow in LBS. Growth of the glmS mutant could alternatively be restored with glucosamine rather than NAG.
Mutants RM1 (alr) and AN3 (glr) grew and swam similarly to the wild type when appropriately supplemented (data not shown), suggesting that they should be able to colonize E. scolopes if the host provides the corresponding d-amino acid. Interestingly, d-amino acids and d-Ala in particular have been found in tissues of marine animals (48, 49), leading us to speculate that at least the alr mutant might colonize squid. However, both RM1 and AN3 were severely attenuated in colonization (Fig. 2), so it appears likely either that d-Ala and d-Glu are not abundant osmolytes in this cephalopod or that they are restricted from the symbiotic environment of the light organ.
The glmS mutant was similarly unable to colonize E. scolopes (Fig. 2), although it grew at a rate similar to that of the wild type in LBS when provided 1 mM NAG or glucosamine (data not shown). This result is consistent with recently published studies suggesting that NAG becomes a significant carbon source for symbiotic V. fischeri well after initial infection (50, 51) and therefore would not be expected to be supplied by the host in sufficient amounts to support colonization in our experiments.
Mutants affected in heme biosynthesis.
Several mutants auxotrophic for hemin were isolated, and each of them contained insertions in hem genes involved in hemin (heme) biosynthesis (Table 1). V. fischeri appears to use the typical bacterial C5 biosynthetic pathway (i.e., the Beale pathway) for heme, with HemA and HemL responsible for making the first committed tetrapyrrole precursor, δ-aminolevulinate (ALA), HemB catalyzing conversion of ALA to porphobilinogen, and several enzymes, including HemC, HemD, HemE, HemF, HemN, HemG, and HemH, sequentially forming protoporphyrin IX and inserting iron to generate heme (52). Using a representative mutant for each locus that was disrupted (Table 1), we found no evidence that growth or motility was attenuated relative to those of the wild type in media with hemin and 2,2-bipyridyl (data not shown). However, mutants with insertions in hemB, hemC, hemD, hemE, hemG, and hemH were severely attenuated in host colonization (Fig. 2). Induction of transcriptional gfp reporters fused to the promoters of hemin-uptake genes was visualized during squid colonization, and a ΔhutD-hutZ mutant lacking this hemin uptake system was outcompeted by the wild type 3-fold over 4 days (30), indicating a symbiotic role for hemin uptake. However, the data reported here suggest that hemin uptake in the light organ is insufficient to support robust colonization if de novo synthesis is blocked. Given the Fur-dependent upregulation of hemin uptake in response to low-iron conditions (30), it seems likely that iron acquisition is the symbiotic function of hemin scavenging by V. fischeri in the squid light organ.
Interestingly, mutants with transposon insertions in hemA and hemL appeared to colonize E. scolopes as well as the wild type (Fig. 2). A mutant engineered with an in-frame deletion of hemA was similarly able to colonize the host light organ (Fig. 3A), although it displayed a 4-fold defect in competition with the wild type (Fig. 3B). The hemA and hemL genes encode enzymes for the first two dedicated steps in hemin biosynthesis, leading to production of ALA, and growth of the hemA and hemL mutants could be supported by supplementing the medium with ALA. We hypothesized that the hemA and hemL mutants are able to colonize the light organ using ALA obtained from the host; however, we considered the alternative possibility that their growth in the light organ is actually supported by hemin. This alternative seemed unlikely given that other hemin-dependent mutants (e.g., a hemB mutant) impaired in the heme biosynthetic pathway downstream of ALA cannot colonize the host (Fig. 2), but this observation could potentially be explained if accumulation of heme pathway intermediates from blocked de novo synthesis were more toxic to cells during growth in the host, or if hemin uptake was accompanied by breakdown and recycling to ALA in the host. We therefore generated and tested a ΔhemA mutant that also has a ΔhutD-hutZ (VF_1220 to VF_1228) deletion of the hemin uptake locus, and we found that it, too, can colonize the E. scolopes light organ (Fig. 3A). Thus, it seems more likely that ALA, and not hemin, is available from the host and supports colonization by the hemA and hemL mutants.
FIG 3.
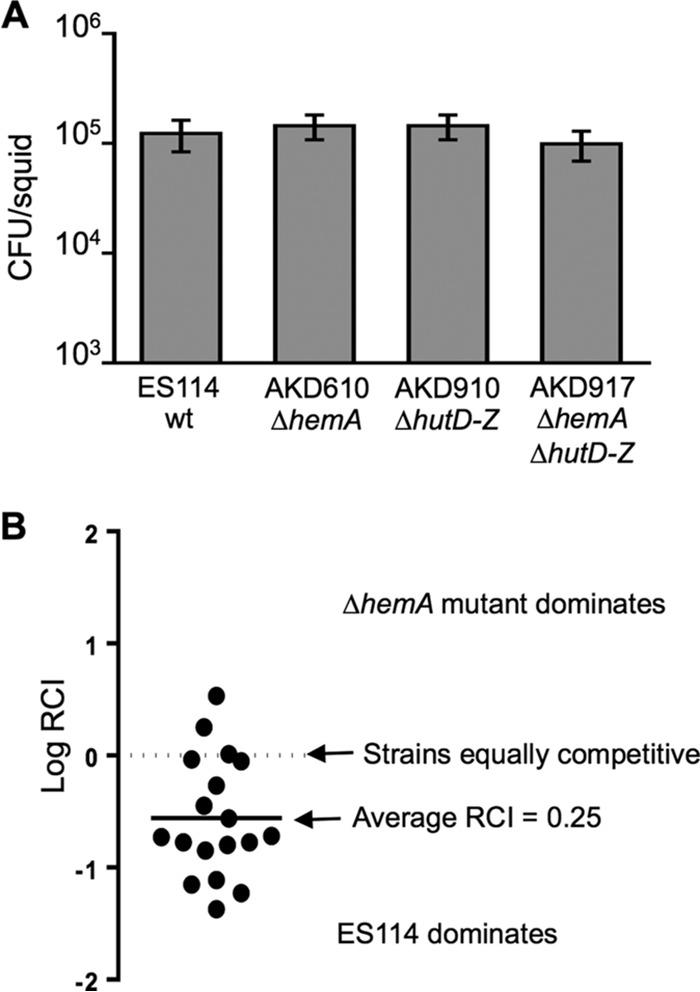
Effects of a ΔhemA mutation on squid colonization. (A) CFU per squid 72 h after inoculation with V. fischeri ES114 (wt), AKD610 (ΔhemA), AKD910 (ΔhutD-hutZ), or AKD917 (ΔhemA ΔhutD-hutZ). Bars indicate standard errors of the means for colonization levels in 20 animals (n = 20). A one-way analysis of variance (ANOVA) indicated that there was no statistical difference in colonization levels for the tested strains (P > 0.05). (B) Relative symbiotic competitiveness of ΔhemA mutant AKD610 43 h after inoculation in a mix with ES114. Each circle indicates the AKD610:ES114 ratio in an individual animal divided by the ratio of these strains in the inoculum. The average log RCI of −0.6 (RCI = 0.25) indicated a statistically significant competitive disadvantage for the mutant (P < 0.0001).
To put these results into context, we tested how much ALA is required to restore growth to cells in culture (Fig. 4). We saw a dose-dependent response to a range of ALA concentrations, from 50 μg ml−1 of ALA, which restored nearly wild-type growth, to 1 μg ml−1 of ALA, which had no detectable effect (Fig. 4). Intermediate growth recovery was observed with 5 or 10 μg ml−1 of ALA. When the ΔhemA mutant was cocultured with the wild type for 20 generations in LBS containing 50 μg ml−1 of ALA, the relative competitiveness index (RCI) was between 0.97 and 0.98 per generation, whereas the RCI during competitive colonization of the host (Fig. 3B), similarly normalized to a per-generation metric by estimating 20 to 30 generations in the host, was between 0.93 and 0.95. A recent survey of ALA in foods indicated that the amount of ALA in squid is near or below the lower end of the range we tested, although the squid species and tissue were not reported (53). Given the amounts of ALA in various animal tissues, it seems neither obvious nor impossible that ALA should be available in sufficient quantity to support growth in the E. scolopes light organ.
FIG 4.

Concentration-dependent recovery of ΔhemA mutant growth by ALA. Wild-type strain ES114 and the ΔhemA mutant AKD610 were grown in LBS supplemented with ALA as indicated. Bars indicate standard errors (n = 3). For clarity, cultures of ES114 with added ALA were not graphed, but addition of ALA to ES114 did not affect its growth significantly.
Our results are remarkably reminiscent of the Bradyrhizobium-legume symbiosis, where the symbionts' hemA is dispensable, hemB is not, and ALA provided by the host is taken up by the colonizing bacteria (54–56). Such availability of ALA does not seem to be universal in host-associated bacteria. For example, a hemA mutant of Vibrio cholerae was significantly (10-fold) attenuated in colonizing the infant mouse gut (57), and a Staphylococcus aureus hemA mutant was similarly reduced in virulence and colonization of hearts and livers in a mouse model (58). In the case of V. fischeri, symbionts in the light organ are provisioned with many amino acids and peptides from the host (59), which may prime cells for uptake of ALA via peptide transport systems (60). The observation that a ΔhemA mutant is slightly outcompeted by the wild type in mixed inoculations could indicate that de novo synthesis of ALA allows somewhat faster growth than uptake, that the supply of ALA is limiting for the mutant in the light organ, or that the availability of ALA is not spatially and temporally uniform in the different microenvironments of this symbiotic tissue.
Conclusions of this mutagenic screen.
In conclusion, by simply amending a rich medium with extra ingredients, we identified new and interesting mutants that have provided insights that would be missed in the defined mutant library generated on LBS. The same approach could likewise extend the gains made from defined mutant libraries of other bacteria, although the specific additives that prove useful may differ. It should be noted that our screen is not entirely saturated, and we know at least two additional mutants could be isolated by this approach. Specifically, a hemin-dependent hemN mutant was isolated but was removed from the study when it was found to contain integrated vector sequences outside the transposon, and a Mg2+-dependent gspE mutant disrupted in the type II general secretion apparatus (61) was inadvertently discarded from our freezer stocks. However, as we have now isolated independent mutants in some of the same loci multiple times (Table 1), the screen is nearing saturation. Future screening for mutants with mutations of conditionally essential genes is more likely to reveal new insights if done under different conditions, for example, screening for mutants that can grow anaerobically but not aerobically. In any case, this study has yielded new insights and provided strains that will facilitate further studies of Vibrio fischeri's cell biology and symbiotic lifestyle.
MATERIALS AND METHODS
Bacteria and media.
V. fischeri strain ES114, which was isolated from an E. scolopes light organ (62), was the wild-type strain used in this study. Plasmids were transformed into Escherichia coli strain DH5α (63) or DH5αλpir (64), with the latter used for plasmids with the R6K replication origin. Plasmids were transferred to V. fischeri via triparental mating using E. coli strain CC118λpir (65) carrying the conjugative helper plasmid pEVS104 as previously described (66).
E. coli was grown at 37°C in LB medium (67) or brain heart infusion (BHI) medium. V. fischeri was grown at 28°C (unless otherwise stated). The media used for V. fischeri include SWTO (68) and ASWT (30), both of which contain yeast extract (3 g liter−1), tryptone (5 g liter−1), and glycerol (3 ml liter−1) and have 700 ml liter−1 of Instant Ocean mixed to 36 ppt (Aquarium Systems, Mentor, OH) in place of seawater in the recipe Boettcher and Ruby used for SWT (62), with SWTO also including extra NaCl (∼4.4 g liter−1) so that its osmolarity approximated that of seawater. Most culturing was performed in LBS, which contains (per liter) 10 g of tryptone, 5 g of yeast extract, 20 g of NaCl, and 50 mM Tris (pH 7.5) (29), or SLBS, which consisted of LBS with added riboflavin (400 μg ml−1), d-alanine (400 μg ml−1), d-glutamate (400 μg ml−1), diaminopimelic acid (DAP; 100 μg ml−1), N-acetylglucosamine (NAG; 1 mM), ribose (2 mM), succinate (5 mM), glycerol (5 mM), hemin (20 μg ml−1), 2,2-bipyridyl (16 μg ml−1), biotin (1 μg ml−1), NAD (NAD+; 25 μg ml−1), folate (15 μg ml−1), thymidine (50 μg ml−1), cytidine (50 μg ml−1), uracil (50 μg ml−1), and MgSO4 (20 mM). These supplements, as well as 50 μg ml−1 of δ-aminolevulinate (ALA), 20 mM MgCl2, or 1 mM glucosamine, were also added to LBS or SWTO individually at the concentrations indicated above to test their ability to support growth of particular mutants.
Solid media were prepared with 15 mg ml−1 of agar for plating. Selection of E. coli was accomplished using chloramphenicol (CAM) or kanamycin (KAN) in LB at final concentrations of 20 and 40 μg ml−1, respectively, or using erythromycin (ERM) in BHI at a final concentration of 150 μg ml−1. Selection of V. fischeri was accomplished using CAM, KAN, and ERM in LBS at final concentrations of 2, 100, and 5 μg ml−1, respectively.
Transposon mutagenesis.
We used pEVS170 to deliver a mini-Tn5 transposon that includes the R6K origin of replication, two bidirectional rho-independent transcriptional terminators, and a gene encoding resistance to ERM (27). Mutants were generated as previously described (27), except that mutants were selected by plating onto SLBS (rather than LBS) containing ERM. Plates were incubated overnight at ∼25°C, individual colonies were patched onto both LBS and SLBS containing ERM, and these patch plates were incubated overnight at ∼25°C. Patches that grew on SLBS, but not on LBS, were examined further. To confirm that the ERM-resistant clones contained transposon insertions and not the entire pEVS170 plasmid, mutants were screened for KAN resistance, which is encoded outside the transposon on pEVS170, and KAN-resistant mutants were disregarded. Transposon insertion sites were determined by cloning insertions from genomic DNA and sequencing across the genome-transposon junctions as previously described (27, 35). For this cloning, DNA ligase and restriction enzymes were acquired from New England BioLabs (Beverly, MA), the GenElute plasmid miniprep kit (Sigma-Aldrich, Inc., St. Louis, MO) was used to purify plasmids, and the DNA Clean and Concentrator-5 kit (Zymo Research, Orange, CA) was used to purify DNA from digestion and ligation reactions. Sequencing was performed at the University of Michigan DNA Sequencing Core Facility.
Targeted mutant construction.
We generated a targeted ΔhemA allele, with an in-frame deletion of the gene formed by placing an NheI site between the second codon and the stop codon. Sequence upstream of hemA and including the first two codons was PCR amplified using primers 5′-ATGTCAGGCACGGTGGAAACCT-3′ (hemAmut1F) and 5′-ACAGCTAGCTGACATTCGGCAAACGGACCAAT-3′ (hemAmut1R), and the amplicon was directly blunt-end cloned into SmaI-digestd pEVS122 (64). Sequence from the hemA stop codon and extending downstream of hemA was PCR amplified using primers 5′-ACAGCTAGCTAAGCGTAGCGTTCGCTACCAAC-3′ (hemAmut2F) and 5′-AGCGGTTTGCGGTTCAAAACGAACAT (hemAmut2R), and the amplicon was directly cloned into pCRBlunt-II-TOPO (Invitrogen, Carlsbad, CA). The plasmids with these sequences flanking hemA were linearized with NheI and fused to generate pAKD610, which contains the hemA deletion allele. The ΔhemA allele on pAKD610 was exchanged into ES114 and the hemin uptake-defective ΔhutD-hutZ (VF_1220-VF_1228) mutant AKD910 (30) to generate mutants AKD610 and AKD917, respectively. The cloned regions in pAKD610 were confirmed by sequencing, and allelic-exchange mutants were confirmed by PCR.
Complemention with ftsQAZ.
The ftsQAZ locus and a small fragment of the downstream lpxC gene were PCR amplified from ES114 genomic template using primers 5′-CGCAAGGCGCTGGAGATGTTGG-3′ (JLSftsQF2) and 5′-GGATCCGCAGGGAAATCAACTGG-3′ (JLSftsZR2), and the resulting amplicon was cloned into pCRBlunt-II-TOPO using the Zero Blunt Topo PCR cloning kit (Invitrogen), generating pJLS233. A portion of the insert containing only ftsQAZ was then PCR amplified from pJLS233 using primers 5′-GTAAAACGACGGCCAG-3′ (M13F) and 5′-TCAGGGTACCGATGATTAATTAGTCAGCTTGGCGT-3′ (JLSftsZR; the underlined sequence is a KpnI site), the amplicon was digested with SphI and KpnI, and the resulting fragment was ligated into SphI- and KpnI-digested pVSV105, which is a stable shuttle vector derived from a plasmid native to V. fischeri (64, 69). The resulting plasmid, pJLS234, and its pVSV105 parent were transferred into V. fischeri in conjugative matings performed on LBS plates supplemented with 20 mM MgCl2.
Microscopy.
Bacteria were viewed by phase-contrast microscopy using a Nikon (Melville, NY) Eclipse E600 microscope. Images were captured with a Nikon Coolpix 5000 camera and edited using the recolor, contrast, brighten, and sharpen features in PowerPoint for Mac 2011 (Microsoft, Redmond, WA). Scale was determined by comparison to an image taken of a micrometer slide.
Dose-dependent reliance of hemA mutant on ALA.
To assay restoration of growth to the hemA mutant with ALA, three independent cultures each of ES114 and AKD610 were grown overnight in LBS medium and in LBS medium containing 50 μg/ml of ALA, respectively. A 100-μl aliquot of culture was pelleted, the supernatant was removed, and the pellets were resuspended in 1 ml of filter-sterilized Instant Ocean. Two-microliter volumes of the resuspended pellets were used to inoculate 200 μl of LBS medium containing 50, 10, 5, 1, or 0 μg ml−1 of ALA in a 96-well plate. The plate was incubated with shaking (double orbital shaking 282 cpm [3 mm]) in a Synergy H1 microplate reader (BioTek, Winooski, VT), with absorbance readings (600 nm) taken approximately every 15 min. The experiment was repeated three times, with results of one representative experiment shown in Fig. 4.
Motility assays.
Semisolid media for assessing swimming motility was prepared by adding 2.5 mg ml−1 of agar to ASWT medium, amended as necessary depending on requirements of the mutants. Overnight cultures were diluted and spotted onto motility plates, and the diameters of areas visibly covered by swimming cells were measured periodically to determine a swimming rate. The motile but ERM resistance-marked ES114 derivative AKD100 (30) and the nonmotile flaJ::aph mutant DM131 (70) were used as positive and negative motility controls, respectively.
Colonization of E. scolopes.
Aposymbiotic E. scolopes hatchlings were inoculated with V. fischeri using previously described procedures (71, 72). Briefly, to prepare inocula, cultures were grown to an optical density at 595 nm (OD595) between 0.4 and 0.7 in 5 ml of ASWT containing the required supplement in 50-ml conical tubes at 28°C without shaking. Cultures were diluted to ∼3,000 CFU ml−1 in Instant Ocean, and hatchling squid were added to 5-ml aliquots of this inoculum individually in 20-ml vials. After 12 to 14 h, hatchlings were rinsed into V. fischeri-free Instant Ocean. Although inocula ranged from 1,500 to 5,000 CFU ml−1, within each experiment squid were exposed to similar concentrations of mutant or wild-type cells in the respective inocula. Infected squid were homogenized at the desired time postinoculation and then serially diluted and plated onto LBS containing the supplement required by the mutant being tested. Plates were incubated overnight at 28°C, and the colonies were counted to calculate CFU per squid. For competition experiments, the mutant and wild-type strains were mixed close to a 1:1 ratio, and the relative competitiveness index (RCI) was calculated as the ratio of mutant to wild type in each animal divided by the ratio of these strains in the inoculum. The results reported either are from multiple experiments or are representative of multiple independent colonization assays.
ACKNOWLEDGMENTS
We thank Cheryl Whistler for helpful discussions, as well as Anisha Nathu, Satyam Veean, Neil Patel, Kishen Patel, Hank Kimbrough, Tyler Shadid, Rickey Patel, Edwina Perschke, and Rosa Muñoz for technical assistance.
A.N.S. was supported by funds awarded by the Department of Defense, Air Force Office of Scientific Research, National Defense Science and Engineering Graduate (NDSEG) Fellowship, 32 CFR 168a. The National Science Foundation supported this research under grants DBI-0755182, IOS-0841480, IOS-1121106, and IOS-1557964 to E.V.S. and MCB-1050687 to A.K.D.
REFERENCES
- 1.Baba T, Ara T, Hasegawa M, Takai Y, Okumura Y, Baba M, Datsenko KA, Tomita M, Wanner BL, Mori H. 2006. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol 2:2006.0008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Cameron DE, Urbach JM, Mekalanos JJ. 2008. A defined transposon mutant library and its use in identifying motility genes in Vibrio cholerae. Proc Natl Acad Sci U S A 105:8736–8741. doi: 10.1073/pnas.0803281105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Gallagher LA, Ramage E, Patrapuvich R, Weiss E, Brittnacher M, Manoil C. 2013. Sequence-defined transposon mutant library of Burkholderia thailandensis. mBio 4:e00604-13. doi: 10.1128/mBio.00604-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Jacobs MA, Alwood A, Thaipisuttikul I, Spencer D, Haugen E, Ernst S, Will O, Kaul R, Raymond C, Levy R, Chun-Rong L, Guenthner D, Bovee D, Olson MV, Manoil C. 2003. Comprehensive transposon mutant library of Pseudomonas aeruginosa. Proc Natl Acad Sci U S A 100:14339–14344. doi: 10.1073/pnas.2036282100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kang Y, Durfee T, Glasner JD, Qiu Y, Frisch D, Winterberg KM, Blattner FR. 2004. Systematic mutagenesis of the Escherichia coli genome. J Bacteriol 186:4921–4930. doi: 10.1128/JB.186.15.4921-4930.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kobayashi K, Ehrlich SD, Albertini A, Amati G, Andersen KK, Arnaud M, Asai K, Ashikaga S, Aymerich S, Bessieres P, Boland F, Brignell SC, Bron S, Bunai K, Chapuis J, Christiansen LC, Danchin A, Debarbouille M, Dervyn E, Deuerling E, Devine K, Devine SK, Dreesen O, Errington J, Fillinger S, Foster SJ, Fujita Y, Galizzi A, Gardan R, Eschevins C, Fukushima T, Haga K, Harwood CR, Hecker M, Hosoya D, Hullo MF, Kakeshita H, Karamata D, Kasahara Y, Kawamura F, Koga K, Koski P, Kuwana R, Imamura D, Ishimaru M, Ishikawa S, Ishio I, Le Coq D, Masson A, Mauel C, et al. 2003. Essential Bacillus subtilis genes. Proc Natl Acad Sci U S A 100:4678–4683. doi: 10.1073/pnas.0730515100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kraemer PS, Mitchell A, Pelletier MR, Gallagher LA, Wasnick M, Rohmer L, Brittnacher MJ, Manoil C, Skerett SJ, Salama NR. 2009. Genome-wide screen in Francisella novicida for genes required for pulmonary and systemic infection in mice. Infect Immun 77:232–244. doi: 10.1128/IAI.00978-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.McAdam RA, Quan S, Smith DA, Bardarov S, Betts JC, Cook FC, Hooker EU, Lewis AP, Woollard P, Everett MJ, Lukey PT, Bancroft GJ, Jacobs WR Jr, Duncan K. 2002. Characterization of a Mycobacterium tuberculosis H37Rv transposon library reveals insertions in 351 ORFs and mutants with altered virulence. Microbiology 148:2975–2986. doi: 10.1099/00221287-148-10-2975. [DOI] [PubMed] [Google Scholar]
- 9.Porwollik S, Santiviago CA, Cheng P, Long F, Desai P, Fredlund J, Srikumar S, Silva CA, Chu W, Chen X, Canals R, Reynolds MM, Bogomolnaya L, Shields C, Cui P, Guo J, Zheng Y, Endicott-Yazdani T, Yang HJ, Maple A, Ragoza Y, Blondel CJ, Valenzuela C, Andrews-Polymenis H, McClelland M. 2014. Defined single-gene and multi-gene deletion mutant collections in Salmonella enterica sv Typhimurium. PLoS One 9:e99820. doi: 10.1371/journal.pone.0099820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Widhelm TJ, Yajjala VK, Endres JL, Fey PD, Bayles KW. 2014. Methods to generate a sequence-defined transposon mutant library in Staphylococcus epidermidis strain 1457. Methods Mol Biol 1106:135–142. doi: 10.1007/978-1-62703-736-5_12. [DOI] [PubMed] [Google Scholar]
- 11.Stabb EV, Visick KL. 2013. Vibrio fischeri: a bioluminescent light-organ symbiont of the bobtail squid Euprymna scolopes, p 497–532. In Rosenberg E, DeLong EF, Stackebrandt E, Lory S, Thompson F (ed), The prokaryotes, 4th ed Springer-Verlag, Berlin, Germany. [Google Scholar]
- 12.Hastings JW, Nealson KH. 1977. Bacterial bioluminescence. Annu Rev Microbiol 31:549–595. doi: 10.1146/annurev.mi.31.100177.003001. [DOI] [PubMed] [Google Scholar]
- 13.Stabb EV. 2005. Shedding light on the bioluminescence “paradox.” ASM News 71:223–229. [Google Scholar]
- 14.Dunn AK, Stabb EV. 2007. Beyond quorum sensing: the complexities of prokaryotic parliamentary procedures. Anal Bioanal Chem 387:391–398. doi: 10.1007/s00216-006-0730-9. [DOI] [PubMed] [Google Scholar]
- 15.Miyashiro T, Ruby EG. 2012. Shedding light on bioluminescence regulation in Vibrio fischeri. Mol Microbiol 84:795–806. doi: 10.1111/j.1365-2958.2012.08065.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Stabb EV, Schaefer A, Bose JL, Ruby EG. 2008. Quorum signaling and symbiosis in the marine luminous bacterium Vibrio fischeri, p 233–250. In Winans SC, Bassler BL (ed), Chemical communication among bacteria. ASM Press, Washington, DC. [Google Scholar]
- 17.Dunn AK. 2012. Vibrio fischeri metabolism: symbiosis and beyond. Adv Microb Physiol 61:37–68. doi: 10.1016/B978-0-12-394423-8.00002-0. [DOI] [PubMed] [Google Scholar]
- 18.Dunn AK, Karr EA, Wang Y, Batton AR, Ruby EG, Stabb EV. 2010. The alternative oxidase (AOX) gene in Vibrio fischeri is controlled by NsrR and upregulated in response to nitric oxide. Mol Microbiol 77:44–55. doi: 10.1111/j.1365-2958.2010.07194.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Brooks JF II, Mandel MJ. 2016. The histidine kinase BinK is a negative regulator of biofilm formation and squid colonization. J Bacteriol 198:2596–2607. doi: 10.1128/JB.00037-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Norsworthy AN, Visick KL. 2015. Signaling between two interacting sensor kinases promotes biofilms and colonization by a bacterial symbiont. Mol Microbiol 96:233–248. doi: 10.1111/mmi.12932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Ray VA, Driks A, Visick KL. 2015. Identification of a novel matrix protein that promotes biofilm maturation in Vibrio fischeri. J Bacteriol 197:518–528. doi: 10.1128/JB.02292-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Visick KL, Quirke KP, McEwen SM. 2013. Arabinose induces pellicle formation by Vibrio fischeri. Appl Environ Microbiol 79:2069–2080. doi: 10.1128/AEM.03526-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.McFall-Ngai MJ. 2014. The importance of microbes in animal development: lessons from the squid-vibrio symbiosis. Annu Rev Microbiol 68:177–194. doi: 10.1146/annurev-micro-091313-103654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Norsworthy AN, Visick KL. 2013. Gimme shelter: how Vibrio fischeri successfully navigates an animal's multiple environments. Front Microbiol 4:356. doi: 10.3389/fmicb.2013.00356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Schwartzman JA, Ruby EG. 2016. A conserved chemical dialog of mutualism: lessons from squid and vibrio. Microbes Infect 18:1–10. doi: 10.1016/j.micinf.2015.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Stabb EV. 2006. The Vibrio fischeri-Euprymna scolopes light organ symbiosis, p 204–218. In Thompson FL, Austin B, Swings J (ed), The biology of vibrios. ASM Press, Washington, DC. [Google Scholar]
- 27.Lyell NL, Dunn AK, Bose JL, Vescovi SL, Stabb EV. 2008. Effective mutagenesis of Vibrio fischeri by using hyperactive mini-Tn5 derivatives. Appl Environ Microbiol 74:7059–7063. doi: 10.1128/AEM.01330-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Brooks JF II, Gyllborg MC, Cronin DC, Quillin SJ, Mallama CA, Foxall R, Whistler C, Goodman AL, Mandel MJ. 2014. Global discovery of colonization determinants in the squid symbiont Vibrio fischeri. Proc Natl Acad Sci U S A 111:17284–17289. doi: 10.1073/pnas.1415957111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Stabb EV, Reich KA, Ruby EG. 2001. Vibrio fischeri genes hvnA and hvnB encode secreted NAD+-glycohydrolases. J Bacteriol 183:309–317. doi: 10.1128/JB.183.1.309-317.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Septer AN, Wang Y, Ruby EG, Stabb EV, Dunn AK. 2011. The heme-uptake gene cluster in Vibrio fischeri is regulated by Fur and contributes to symbiotic colonization. Environ Microbiol 13:2855–2864. doi: 10.1111/j.1462-2920.2011.02558.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hemberger S, Pedrolli DB, Stolz J, Vogl C, Lehmann M, Mack M. 2011. RibM from Streptomyces davawensis is a riboflavin/roseoflavin transporter and may be useful for the optimization of riboflavin production strains. BMC Biotechnol 11:119. doi: 10.1186/1472-6750-11-119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Valle E, Ledon T, Cedre B, Campos J, Valmaseda T, Rodriguez B, Garcia L, Marrero K, Benitez J, Rodriguez S, Fando R. 2000. Construction and characterization of a nonproliferative El Tor cholera vaccine candidate derived from strain 638. Infect Immun 68:6411–6418. doi: 10.1128/IAI.68.11.6411-6418.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mao F, Dam P, Chou J, Olman V, Xu Y. 2009. DOOR: a database for prokaryotic operons. Nucleic Acids Res 37:D459–D463. doi: 10.1093/nar/gkn757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Adin DM, Phillips NJ, Gibson BW, Apicella MA, Ruby EG, McFall-Ngai MJ, Hall DB, Stabb EV. 2008. Characterization of htrB and msbB mutants of the light organ symbiont Vibrio fischeri. Appl Environ Microbiol 74:633–644. doi: 10.1128/AEM.02138-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lyell NL, Dunn AK, Bose JL, Stabb EV. 2010. Bright mutants of Vibrio fischeri ES114 reveal conditions and regulators that control bioluminescence and expression of the lux operon. J Bacteriol 192:5103–5114. doi: 10.1128/JB.00524-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.O'Shea TM, DeLoney-Marino CR, Shibata S, Aizawa S-I, Wolfe AJ, Visick KL. 2005. Magnesium promotes flagellation of Vibrio fischeri. J Bacteriol 187:2058–2065. doi: 10.1128/JB.187.6.2058-2065.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.O'Shea TM, Klein AH, Geszvain K, Wolfe AJ, Visick KL. 2006. Diguanylate cyclases control magnesium-dependent motility of Vibrio fischeri. J Bacteriol 188:8196–8205. doi: 10.1128/JB.00728-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Coughlin RT, Tonsager S, McGroarty EJ. 1983. Quantitation of metal cations bound to membranes and extracted lipopolysaccharide of Escherichia coli. Biochemistry 22:2002–2007. doi: 10.1021/bi00277a041. [DOI] [PubMed] [Google Scholar]
- 39.García Véscovi E, Soncini FC, Groisman EA. 1996. Mg2+ as an extracellular signal: environmental regulation of Salmonella virulence. Cell 84:165–174. doi: 10.1016/S0092-8674(00)81003-X. [DOI] [PubMed] [Google Scholar]
- 40.Shih YL, Rothfield L. 2006. The bacterial cytoskeleton. Microbiol Mol Biol Rev 70:729–754. doi: 10.1128/MMBR.00017-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Mandel MJ, Stabb EV, Ruby EG. 2008. Comparative genomics-based investigation of resequencing targets in Vibrio fischeri: focus on point miscalls and artefactual expansions. BMC Genomics 9:138. doi: 10.1186/1471-2164-9-138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Charbon G, Cabeen MT, Jacobs-Wagner C. 2009. Bacterial intermediate filaments: in vivo assembly, organization, and dynamics of crescentin. Genes Dev 23:1131–1144. doi: 10.1101/gad.1795509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Allen RD, Baumann P. 1971. Structure and arrangement of flagella in species of the genus Beneckea and Photobacterium fischeri. J Bacteriol 107:295–302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Graf J, Dunlap PV, Ruby EG. 1994. Effect of transposon-induced motility mutations on colonization of the host light organ by Vibrio fischeri. J Bacteriol 176:6986–6991. doi: 10.1128/jb.176.22.6986-6991.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Millikan DS, Ruby EG. 2002. Alterations in Vibrio fischeri motility correlate with a delay in symbiosis initiation and are associated with additional symbiotic colonization defects. Appl Environ Microbiol 68:2519–2528. doi: 10.1128/AEM.68.5.2519-2528.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gerdes SY, Scholle MD, Campbell JW, Balazsi G, Ravasz E, Daugherty MD, Somera AL, Kyrpides NC, Anderson I, Gelfand MS, Bhattacharya A, Kapatral V, D'Souza M, Baev MV, Grechkin Y, Mseeh F, Fonstein MY, Overbeek R, Barabasi AL, Oltvai ZN, Osterman AL. 2003. Experimental determination and system level analysis of essential genes in Escherichia coli MG1655. J Bacteriol 185:5673–5684. doi: 10.1128/JB.185.19.5673-5684.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Langridge GC, Phan MD, Turner DJ, Perkins TT, Parts L, Haase J, Charles I, Maskell DJ, Peters SE, Dougan G, Wain J, Parkhill J, Turner AK. 2009. Simultaneous assay of every Salmonella Typhi gene using one million transposon mutants. Genome Res 19:2308–2316. doi: 10.1101/gr.097097.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Abe H, Yoshikawa N, Sarower MG, Okada S. 2005. Physiological function and metabolism of free D-alanine in aquatic animals. Biol Pharm Bull 28:1571–1577. doi: 10.1248/bpb.28.1571. [DOI] [PubMed] [Google Scholar]
- 49.Felbeck H, Wiley S. 1987. Free D-amino acids in the tissues of marine bivalves. Biol Bull 173:252–259. doi: 10.2307/1541877. [DOI] [PubMed] [Google Scholar]
- 50.Pan M, Schwartzman JA, Dunn AK, Lu Z, Ruby EG. 2015. A single host-derived glycan impacts key regulatory nodes of symbiont metabolism in a coevolved mutualism. mBio 6:e00811-15. doi: 10.1128/mBio.00811-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sun Y, Verma SC, Bogale H, Miyashiro T. 2015. NagC represses N-acetyl-glucosamine utilization genes in Vibrio fischeri within the light organ of Euprymna scolopes. Front Microbiol 6:741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Beale SI. 2007. Biosynthesis of hemes. EcoSal Plus 2007 doi: 10.1128/ecosalplus.3.6.3.11. [DOI] [PubMed] [Google Scholar]
- 53.Rodriguez BL, Curb JD, Davis J, Shintani T, Perez MH, Apau-Ludlum N, Johnson C, Harrigan RC. 2012. Use of the dietary supplement 5-aminiolevulinic acid (5-ALA) and its relationship with glucose levels and hemoglobin A1C among individuals with prediabetes. Clin Transl Sci 5:314–320. doi: 10.1111/j.1752-8062.2012.00421.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Chauhan S, O'Brian MR. 1993. Bradyrhizobium japonicum δ-aminolevulinic acid dehydratase is essential for symbiosis with soybean and contains a novel metal-binding domain. J Bacteriol 175:7222–7227. doi: 10.1128/jb.175.22.7222-7227.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.McGinnis SD, O'Brian MR. 1995. The rhizobial hemA gene is required for symbiosis in species with deficient δ-aminolevulinic acid uptake activity. Plant Physiol 108:1547–1552. doi: 10.1104/pp.108.4.1547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sangwan I, O'Brian MR. 1991. Evidence for an inter-organismic heme biosynthetic pathway in symbiotic soybean root nodules. Science 251:1220–1222. doi: 10.1126/science.251.4998.1220. [DOI] [PubMed] [Google Scholar]
- 57.Ravichandran M, Ali SA, Rashid NH, Kurunathan S, Yean CY, Ting LC, Bakar AS, Lalitha P, Zainuddin ZF. 2006. Construction and evaluation of a O139 Vibrio cholerae vaccine candidate based on a hemA gene mutation. Vaccine 24:3750–3761. doi: 10.1016/j.vaccine.2005.07.016. [DOI] [PubMed] [Google Scholar]
- 58.Hammer ND, Reniere ML, Cassat JE, Zhang Y, Hirsch AO, Indriati Hood M, Skaar EP. 2013. Two heme-dependent terminal oxidases power Staphylococcus aureus organ-specific colonization of the vertebrate host. mBio 4:e00241-13. doi: 10.1128/mBio.00241-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Graf J, Ruby EG. 1998. Host-derived amino acids support the proliferation of symbiotic bacteria. Proc Natl Acad Sci U S A 95:1818–1822. doi: 10.1073/pnas.95.4.1818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Verkamp E, Backman VM, Bjornsson JM, Soll D, Eggertsson G. 1993. The periplasmic dipeptide permease system transports 5-aminolevulinic acid in Escherichia coli. J Bacteriol 175:1452–1456. doi: 10.1128/jb.175.5.1452-1456.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Dalbey RE, Kuhn A. 2012. Protein traffic in Gram-negative bacteria—how exported and secreted proteins find their way. FEMS Microbiol Rev 36:1023–1045. doi: 10.1111/j.1574-6976.2012.00327.x. [DOI] [PubMed] [Google Scholar]
- 62.Boettcher KJ, Ruby EG. 1990. Depressed light emission by symbiotic Vibrio fischeri of the sepiolid squid Euprymna scolopes. J Bacteriol 172:3701–3706. doi: 10.1128/jb.172.7.3701-3706.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hanahan D. 1983. Studies on transformation of Escherichia coli with plasmids. J Mol Biol 166:557–580. doi: 10.1016/S0022-2836(83)80284-8. [DOI] [PubMed] [Google Scholar]
- 64.Dunn AK, Martin MO, Stabb EV. 2005. Characterization of pES213, a small mobilizable plasmid from Vibrio fischeri. Plasmid 54:114–134. doi: 10.1016/j.plasmid.2005.01.003. [DOI] [PubMed] [Google Scholar]
- 65.Herrero M, De Lorenzo V, Timmis KN. 1990. Transposon vectors containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in Gram-negative bacteria. J Bacteriol 172:6557–6567. doi: 10.1128/jb.172.11.6557-6567.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Stabb EV, Ruby EG. 2002. RP4-based plasmids for conjugation between Escherichia coli and members of the Vibrionaceae. Methods Enzymol 358:413–426. doi: 10.1016/S0076-6879(02)58106-4. [DOI] [PubMed] [Google Scholar]
- 67.Miller JH. 1992. A short course in bacterial genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. [Google Scholar]
- 68.Bose JL, Kim U, Bartkowski W, Gunsalus RP, Overley AM, Lyell NL, Visick KL, Stabb EV. 2007. Bioluminescence in Vibrio fischeri is controlled by the redox-responsive regulator ArcA. Mol Microbiol 65:538–553. doi: 10.1111/j.1365-2958.2007.05809.x. [DOI] [PubMed] [Google Scholar]
- 69.Dunn AK, Millikan DS, Adin DM, Bose JL, Stabb EV. 2006. New rfp- and pES213-derived tools for analyzing symbiotic Vibrio fischeri reveal patterns of infection and lux expression in situ. Appl Environ Microbiol 72:802–810. doi: 10.1128/AEM.72.1.802-810.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Adin DM, Engle JT, Goldman WE, McFall-Ngai MJ, Stabb EV. 2009. Mutations in ampG and lytic transglycosylase genes affect the net release of peptidoglycan monomers from Vibrio fischeri. J Bacteriol 191:2012–2022. doi: 10.1128/JB.01547-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ruby EG, Asato LM. 1993. Growth and flagellation of Vibrio fischeri during initiation of the sepiolid squid light organ symbiosis. Arch Microbiol 159:160–167. doi: 10.1007/BF00250277. [DOI] [PubMed] [Google Scholar]
- 72.Stabb EV, Ruby EG. 2003. Contribution of pilA to competitive colonization of the squid Euprymna scolopes by Vibrio fischeri. Appl Environ Microbiol 69:820–826. doi: 10.1128/AEM.69.2.820-826.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]