

Abstract
Nocturnal dung beetles (Scarabaeus satyrus) are currently the only animals that have been demonstrated to use the Milky Way for reliable orientation. In this study, we tested the capacity of S. satyrus to orient under a range of artificial celestial cues, and compared the properties of these cues with images of the Milky Way simulated for a beetle's visual system. We find that the mechanism that permits accurate stellar orientation under the Milky Way is based on an intensity comparison between different regions of the Milky Way. We determined the beetles' contrast sensitivity for this task in behavioural experiments in the laboratory, and found that the resulting threshold of 13% is sufficient to detect the contrast between the southern and northern arms of the Milky Way under natural conditions. This mechanism should be effective under extremely dim conditions and on nights when the Milky Way forms a near symmetrical band that crosses the zenith. These findings are discussed in the context of studies of stellar orientation in migratory birds and itinerant seals.
This article is part of the themed issue ‘Vision in dim light’.
Keywords: star, sky compass, straight-line orientation, vision
1. Introduction
For many animal species, a sense of direction is vital to survival and reproduction. While landmarks and other terrestrial cues can prove impermanent and unreliable over the course of evolutionary history, celestial cues, such as the sun, the moon, skylight and the stars, are both stable and predictable over millennia. It is perhaps no wonder that numerous species rely on celestial cues as a frame of reference for accurate orientation [1]. While solar orientation has been characterized in a range of both migratory (e.g. Lepidoptera [2,3], birds [4], teleost fishes [5]) and non-migratory (e.g. desert ants [6,7], bees [8,9], toads [10]) species, the mechanisms by which animals orient using the stars [11–18] remain less well understood. Most human cultures have developed methods to use the stars for navigation [19], and it has been shown that migratory birds [12,13,15] and perhaps also seals [16] navigate using star patterns. Stellar orientation has also been implied in the cricket frog Acris gryllus [11] and the yellow underwing moth Noctua pronuba [14], although the strategy by which this is achieved is less clear.
The nocturnal dung beetle Scarabaeus satyrus performs a robust orientation behaviour on moonless nights when only the Milky Way is visible [17,18] and represents an ideal species for the study of stellar orientation strategies in an insect. On arriving at a fresh dung pile, these ball-rolling dung beetles sculpt a dung ball, which they then roll in a straight line away from the dung pile and competing dung beetles, until, at a sufficient distance, the ball can be buried and consumed undisturbed. Although larger ‘brood balls’ are constructed by breeding dung beetles (males, females or pairs), both sexes feed by constructing, rolling and consuming dung balls throughout their period of activity. In order to maintain this straight-line travel, S. satyrus uses the celestial cues available, which may include the moon [20], the lunar polarization pattern [21] or the Milky Way [17,18]. While it is thought that special visual adaptations are needed to interpret the lunar polarization pattern and the Milky Way when orienting [18], it is not known what strategy these beetles employ to determine an accurate geographical heading using the Milky Way. The Milky Way may prove particularly challenging to interpret as an orientation cue. On occasions where it forms a symmetric band that passes close to the zenith, its symmetric ambiguity could lead to 180° orientation errors if no difference between the two halves of the sky can be discerned. Nevertheless, S. satyrus is well oriented on nights when the Milky Way passes close to the zenith [17,18].
In this study, we investigated how S. satyrus cope with the symmetric ambiguity in stellar cues, and probed which strategies they may use to reduce ambiguity in stellar orientation. Previous investigations of stellar orientation suggest that other animal species learn to recognize star patterns as orientation cues. Migrating birds have been shown to orient relative to the starry sky's centre of rotation [13,15], which they recognize via the pattern of stars within that sky region [13]. The harbour seal Phoca vitulina can learn to swim towards a ‘lodestar’, which they are thought to identify via a learnt star pattern [16]. We tested whether S. satyrus also uses a pattern-matching strategy for stellar orientation, or if they use another strategy, such as fixating the brightest point [18] or matching a memorized intensity gradient [22]. We propose that, while a pattern-matching strategy would involve recording the relative positions of multiple bright sky regions when facing in the intended direction, a stored representation of an intensity gradient would contain only information about the direction of the arc bisecting the celestial hemisphere within which the maximum difference in intensity between the two halves of that arc was observed. This would be distinct from a strategy of fixating the brightest point, which would require only information about the position of the brightest point relative to the intended direction. The different experimental conditions were compared with estimates of the whole-sky radiance pattern obtained from calibrated photographs of the Milky Way recorded at the field site in South Africa, to estimate how the strategies identified here might predict performance under natural conditions.
2. Material and methods
(a). Sky imaging
To characterize the celestial light available for accurate orientation on a clear moonless night, we determined the radiance distribution in the night sky from calibrated digital photographs. Photographs were taken at the game farm ‘Stonehenge’ (26°23′55.82″ S, 24°19′37.38″ E), in South Africa, using a digital camera (D810: Nikon Corp., Shinjuku, Japan) fitted with a fisheye lens (Sigma 8 mm F3.5 EX DG: Sigma Corp., Kawasaki, Japan), which, when pointed straight upwards, imaged the whole celestial hemisphere. The camera was calibrated in order to correct for different exposure times, ISO speed settings and apertures, as well as for the effect of vignetting (i.e. lower exposure at the image edges) to provide absolute radiance measurements for each pixel [23]. In order to reduce the influence of bright lights at the horizon (e.g. airglow, albedo, light pollution from distant settlements) on our analysis, and because these areas are often blocked from the beetles' view by vegetation under natural conditions, we blacked out any parts of the image below 20° of elevation. Details of the accuracy of measurements of spectral photon radiance via this method are reported in the electronic supplementary material, ‘Accuracy of the imaging method’.
Calibrated and cropped images were filtered in custom-made software, written in Matlab 2015b (MathWorks, Natick, USA), to approximate blurring by compound eye optics. As the acceptance functions of photoreceptors in the dorsal eye of dung beetles are not known, the median interommatidial angle of 2° (estimated from anatomical data in S. satyrus) was used as an estimate of maximum potential acuity. Information theory predicts that with interommatidial angles of 2°, ideal sampling would be achieved with an acceptance angle of 4° [24], but many insects are known to have much wider acceptance functions in their dorsal eye, and especially at later stages of neural processing (e.g. [25]). We, therefore, applied a number of Gaussian filters ranging in half-width (full width at half maximum; FWHM) from 2° to 16°. Filter kernels were accurately calculated to take into account the distortion of the photographs by the camera lens. The resulting images are shown in figure 1a.
Figure 1.

An analysis of the celestial light available for orientation on a moonless night. (a) Linear images (pixel values proportional to absolute spectral photon radiance measured in each channel) of the sky in November 2015, oriented so that magnetic north is to the left and west at the top. Areas below 20° elevation were removed, and images were filtered with a Gaussian filter of 2°, 4°, 8° and 16° half-width. (b) Spectral photon radiance along the Milky Way in November 2015 (red line) and February 2016 (blue line). The maximum at southern elevations of approximately 30° and 45°, in November and February, respectively, corresponds to the ‘bright spot’ consisting of the constellation Crux and the Southern Pleiades. (c) Michelson contrast between points of the same elevation in the southern versus the northern arm of the Milky Way. Colours as in (b).
To define the position of the band of the Milky Way, we fitted a quadratic function through the east–west intensity maxima (as the Milky Way was almost north–south in all of our images) of the most highly filtered image, and applied this positional function to all images (electronic supplementary material, figure S1). The contrast between the southern and the northern arms of the Milky Way was calculated as Michelson contrast, i.e. 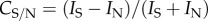 , where IS and IN are the radiance of points in the southern and northern arm, respectively. Contrasts between the Milky Way and the background were similarly calculated as, for example,
, where IS and IN are the radiance of points in the southern and northern arm, respectively. Contrasts between the Milky Way and the background were similarly calculated as, for example, 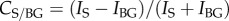 , where IBG is the mean radiance of the background. For the determination of background intensity, we calculated the mean of all parts of the image that were (i) more than 30° above the horizon and (ii) more than 150 pixels (corresponding to approx. 30°) away from the fitted band of the Milky Way. These exclusions removed most bright areas at the horizon and around the Milky Way, resulting in a slightly conservative estimate of the Milky Way's contrast against the background (electronic supplementary material, figures S2–S3).
, where IBG is the mean radiance of the background. For the determination of background intensity, we calculated the mean of all parts of the image that were (i) more than 30° above the horizon and (ii) more than 150 pixels (corresponding to approx. 30°) away from the fitted band of the Milky Way. These exclusions removed most bright areas at the horizon and around the Milky Way, resulting in a slightly conservative estimate of the Milky Way's contrast against the background (electronic supplementary material, figures S2–S3).
(b). Beetle handling
To determine orientation precision under different celestial cues, individual beetles were placed with their front legs touching a dung ball in the centre of a flat, 1 m diameter circular arena, marked at 5° increments from 0° to 355° along its perimeter. Each beetle was permitted to climb onto its ball, perform an ‘orientation dance’ and then roll its ball towards the edge of the arena. On reaching the edge, the stimulus was changed and the beetle was placed back at the centre of the arena and allowed to roll to the edge again, thus completing a second trial. For each trial, the exit heading was recorded and the difference between these headings was calculated to give the ‘change in heading’ angle. The change in stimulus orientation between the two trials (e.g. 90° clockwise or anticlockwise) was then subtracted from this change in heading to give a measure of ‘orientation error’ relative to the stimulus' position. The number of beetles tested in each condition ranged between 20 and 40. Since all beetles exhibited normal foraging behaviour during trials, the sexes of individual beetles were not identified. All trials were recorded using an IR-sensitive camcorder (HDR-CX730: Sony, Tokyo, Japan) and beetles were marked with a strip of high-gain reflective tape (Scotchlite™ 7610: 3M Company, Maplewood, USA) to maximize visibility in the recorded videos. Experiments were carried out at Lund University, Sweden, at the game farm ‘Stonehenge’, north-west of Vryburg, North-West Province, South Africa and at the Nicama Lodge (24°49'40.85″ S, 27°59'32.60″ E), Bela-Bela, Limpopo, South Africa.
(c). Symmetric cues
To determine if a symmetric pattern of skylight, such as that presented by the main band of the Milky Way across the zenith, is sufficient for accurate orientation, the ability of nocturnal dung beetles to orient to two lights 180° apart in azimuth was tested. Under the ‘equal intensity’ condition (figure 2), two green (λmax = 525 nm) LED arrays (LZ1-10G100: LedEngin Inc., San Jose, USA), acting as light spots, were displayed simultaneously, either at the north and south or east and west positions at an elevation of 45°. These light spots were displayed at a distance of 2 m from the centre of a 1 m diameter arena. To simulate nocturnal conditions, these LEDs were dimmed using four layers of 1.2ND filter (Lee Filters 299: Andover, UK) and one layer of 1/8 white diffusion filter (Lee Filters 252) to a photon flux of 1.25 × 107 photons cm−2 s−1 (see electronic supplementary material, Intensity Measurements). For this experiment, the stimulus was not moved between trials, and orientation error was therefore calculated from the difference in headings between two trials with identical stimuli. These data were compared with orientation errors recorded with no light spots illuminated (no lights) and with an ‘intensity difference’ condition, in which one of these light spots was dimmed using a further layer of 1.2ND filter and both light spots were closer (650 mm as opposed to 2 m) to the arena centre, resulting in intensities of 1.99 × 108 photons cm−2 s−1 and 1.45 × 107 photons cm−2 s−1, respectively (a Michelson contrast of 86.5%). All trials were carried out at Lund University, between 7 and 13 May 2015, with the exception of the ‘intensity difference’ experiment, which was carried out at the ‘Stonehenge’ field site in South Africa on 2 February 2015. Trials at Lund University were carried out between 13.00 and 18.00 for beetles that had habituated to a 12.00–00.00 dark period, while trials in South Africa were carried out between 20.00 and 00.00 during the typical activity period for S. satyrus [18].
Figure 2.
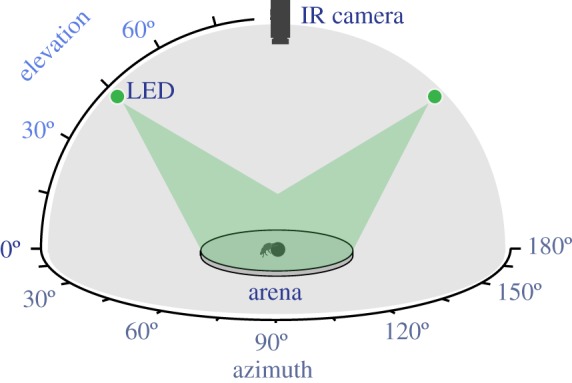
The arrangement used to create symmetric cues. The ‘equal intensity’ symmetric stimulus consisted of two light spots 180° apart. In the ‘intensity difference’ symmetric stimulus one of the two light spots was dimmed by one order of magnitude. In the ‘no lights’ condition no light spots were illuminated, but all other aspects of the arrangement were identical to the other conditions. (Online version in colour.)
(d). Significance of star patterns
In order to determine whether an overhead intensity pattern would allow for accurate orientation of the kind observed under the Milky Way [17,18], patterns containing directional information were displayed using a set of 16 ‘cool white’ LEDs (DDW-UJ2-TU1-1: Roithner LaserTechnik GmbH, Vienna, Austria), arranged in two perpendicular rows of 8, arching over the test arena (figure 3a). One band went from the north-east of the arena to the south-west, and the other from the north-west to the south-east. The light from each LED was projected down a 100 mm tube with a 25 mm aperture, onto one layer of 1.2ND filter (Lee Filters 299) followed by four layers of 1/16 white diffusion filter (Lee Filters 452) to produce a light spot (figure 3b). This arrangement was intended to limit fluctuations in brightness with viewing angle when observed from the arena's surface. The light spots were arranged in each band such that they were 1600 mm from the arena centre and positioned at elevations of 30°, 45°, 60° and 75° on each side (half) of the band. The absolute irradiance of a single light spot in this arrangement was estimated (see electronic supplementary material, ‘Intensity Measurements’) to be 8.23 × 107 photons cm−2 s−1 when observed from the arena centre.
Figure 3.

Arrangement used to create overhead intensity patterns. (a) Above view showing light spots positioned in two perpendicular bands. Between trials the band's orientation was changed by 90°. (b) A surface-mounted LED projected white light onto a series of filters, creating a diffuse light spot. (c) The ‘high elevation’ pattern, in which there was a difference between the two sides of the overhead intensity pattern at 45° and 60°. (d) The ‘low elevation’ pattern, in which this difference was at 30° and 45°. (e) The ‘one side lit’ condition, in which only the four light spots on the northern side of the band were illuminated. (f) The ‘no lights’ condition. (g) For all contrast levels, except for 100% (one side lit) and 0%, all eight light spots were lit, but the four on the southern side were dimmer than those on the northern side. (h) The 0% contrast stimulus, in which all eight light spots were equally bright. (Online version in colour.)
Pattern stimuli were designed such that the two sides of the band contained the same number of illuminated light spots, but in different configurations with unambiguous directional information. In the ‘high elevation’ condition, the light spots illuminated in the north–south direction along the band were at 30°, 45°, 75°, 75°, 60° and 30° elevations (figure 3c), such that the directional cue lay in whether there was a light spot at 45° or 60° on a given side. In the ‘low elevation’ condition (figure 3d), only the light spots at 45°, 75°, 75° and 30° elevations were illuminated, and hence the directional cue available was the presence of a light spot at 30° or 45°. A further ‘one side lit’ condition (figure 3e), in which only the light spots at 30°, 45°, 60° and 75° on the northern side of the band were lit, was used to determine the maximum level of directedness achievable under this type of stimulus. This ‘one side lit’ stimulus (with four light spots illuminated) produced similar ambient lighting conditions to the ‘low elevation’ pattern. During these experiments, each beetle viewed the pattern first in one orientation, either NE–SW or NW–SE, and then at 90° to this in the following trial. Change in heading angle between the two trials, relative to this 90° change in stimulus position, was recorded as a measure of ‘orientation error’. A ‘no lights’ condition in which all light spots were turned off (figure 3f) was also tested to control for the presence of alternative directional cues. Experiments were carried out at Nicama Lodge in South Africa, between 20.00 and 00.00, on 15–18 February 2016.
(e). Significance of contrast
In order to determine the level of contrast required by S. satyrus for accurate orientation under an otherwise symmetric band of light, such as the Milky Way, we performed a series of experiments testing orientation under a symmetric band of light spots containing a brightness difference (figure 3g); the light spots at 30°, 45°, 60° and 75° on the northern side of the band were brighter than their counterparts on the opposite side. As before, the band was displayed in the first trial in either a NE–SW or NW–SE orientation, and turned by 90° for the second trial (figure 3a). A condition in which there was no difference between the two sides (‘0% contrast’; figure 3h) was also tested to determine a typical response to a ‘symmetric’ band stimulus and to control for the presence of any other directional cues. The absolute irradiance of a single light spot in this arrangement was varied across the range 5.55 × 107− 8.29 × 108 photons cm−2 s−1, when observed from the arena centre, through changes in current for contrasts between 0% and 20.6% and via removal of the 1.2ND filter from light spots on the band's northern side for the 87.5% contrast condition. Experiments were carried out at Nicama Lodge in South Africa, between 20.00 and 00.00, on 15–18 February 2016, with the exception of the 9.5% contrast condition, which was tested at Lund University between 13.00 and 18.00 (see §2c), on 26–27 March 2016.
(f). Statistics
The V test [26] was used to determine whether the pattern of orientation error was centred on zero. All directional statistical tests were carried out using the package ‘circular’ [27] in R v. 3.3.0 [28]. Levels of orientation were compared using absolute orientation error [22,29]. Since this is a bounded measure (in the range 0–180°) that does not necessarily follow a normal distribution, a Mann–Whitney rank-sum test was used to determine if median absolute orientation error at each contrast level was significantly lower than in the absence of directional cues (the ‘no illumination’ condition), or than 90°: the average for either a uniform distribution (in the case of disoriented behaviour) or a bimodal distribution (in the case of a trend in both 0° and 180° errors driven by stimulus ambiguity) of absolute orientation errors. The Mann–Whitney test was thus used to test for ‘extent of disorientation’ and to identify conditions that reduced ‘disorientation’ for a significant proportion of individuals.
To determine the lower contrast threshold for orientation under the ‘contrasting band’ stimulus, a contrast-sensitivity curve was fitted to the absolute orientation error data across all contrast levels, including the ‘no difference’ condition as 0% contrast and the ‘one side lit’ condition as 100% contrast (assuming a radiance of approximately zero from the unlit region of the stimulus band). This curve was fitted with a presumed upper asymptote at 90°, the average absolute orientation error of a uniform or bimodal distribution. The lower asymptote was determined via an optimization routine using the ‘glm.WH’ function in the package ‘psyphy’ [30] in R 3.3.0 [28]. The threshold was calculated from the inflection point of the fitted curve, the point halfway between the lower and upper asymptotes.
3. Results and discussion
(a). Sky imaging
In order to determine what orientation cues might be present in the starry sky as viewed by dung beetles, calibrated photographs were used to map the radiance distribution in the night sky. These calibrated images of spectral radiance in the moonless sky show a distinctive Milky Way main band that bisects the sky, crossing the zenith (figure 1a). Depending on the half-width of the filter applied to these images, different features of the Milky Way were available, the ‘bright spot’ being clearly discernible for images filtered with half-widths of 2° and 4°, but less so for images filtered with half-widths of 8° and 16° (figure 1a). Images recorded in November 2015 and February 2016 showed a similar intensity profile across the main band of the Milky Way (figure 1b), one difference being the position of the bright spot in the south, which fell at approximately 30° in the November image and 45° in the February image.
In order to investigate asymmetries that might be used as orientation cues, Michelson contrast was calculated between points in the (brighter) southern and (dimmer) northern regions of the Milky Way band at the same elevation. These opposing points could be compared by an orienting beetle while rolling its dung ball or during the orientation dance [31]. Excluding elevations near to the horizon and the 20° cut-off point, maximum south–north contrasts between points at the same elevation on the Milky Way's main band fell at the bright spot (figure 1c; with the exception of the 16° filtered image from February), with maximum contrasts here ranging between 12.6% and 41.1% depending on the extent of image filtering. These images indicate that the Milky Way's main band forms a distinctive feature of the moonless night sky that, while near symmetric in configuration when crossing the zenith (electronic supplementary material, figure S3), presents an asymmetric intensity profile that could be used as an orientation cue across a range of likely visual acuity levels.
(b). Symmetric cues
On moonless starlit nights in South Africa the Milky Way's main band regularly crosses the zenith (figure 1a), resulting in a near symmetrical pattern of celestial intensity that nocturnal dung beetles need to interpret to achieve oriented behaviour. The results of previous studies [17,18] show that, even under these particularly challenging conditions, S. satyrus is capable of maintaining its heading with low orientation error. One way that they could achieve this is by taking a snapshot of the light pattern across the whole sky as they are facing their chosen direction, and then matching this pattern after any interruption to their rolling path. For example, it has been demonstrated that the diurnal ball-rolling dung beetle Scarabaeus lamarcki stores a snapshot of the positions of celestial cues during its orientation dance, and accesses this information, following an interruption of rolling behaviour, when permitted to continue rolling [31]. If used, this strategy would result in a bimodal distribution of orientation errors (clustered around 0° and 180°) when the beetles are presented with a fully symmetrical light pattern. This type of error distribution has been observed in the southern cricket frog A. gryllus attempting to orient under the Milky Way [11] and dung beetles presented with a polarizing filter [21,22,32]. In this study, beetles in the ‘equal intensity’ condition were not significantly unimodally oriented (V statistic = 0.059, p = 0.343, μ ± s.d = 71.2 ± 105.8°, n = 24; figure 4a). However, in contrast with the predictions of a stellar pattern-matching strategy, orientation error also did not follow a bimodal distribution (V statistic = −0.042, p = 0.614, μ ± s.d = 127.9 and 307.9 ± 53.8°, n = 24). In addition, absolute orientation error (figure 4b) was not significantly lower than that in the ‘no lights’ condition (Mann–Whitney rank-sum test, W = 461.5, p = 0.675, nequal intensity = 24, nno lights = 36, median absolute orientation error: ŷequal intensity = 77.5), indicating that the presence of these two equally bright lights did not improve heading fidelity. This finding supports that of el Jundi and colleagues [33] in diurnal dung beetles (under much brighter conditions). Nevertheless, in the ‘intensity difference’ condition in the present study, the beetles were well oriented (V statistic = 0.610, p < 0.001, μ ± s.d = 355.8 ± 56.9°, n = 20), with significantly lower absolute orientation error than for the ‘no lights’ condition (Mann–Whitney rank-sum test, W = 197, p = 0.003, ŷintensity difference = 35.0) (figure 4a,b). These results indicate that, at starlight illumination levels, S. satyrus were not capable of using a symmetric stimulus with two points of maximum brightness for accurate orientation, and suggest that S. satyrus' stellar orientation strategy requires some intensity difference within a symmetric configuration to function.
Figure 4.

A comparison of directedness in the absence of light stimuli (no lights), under two equally bright light spots 180° apart (equal intensity), and for the same arrangement with an order of magnitude difference between the two light spots (intensity difference). Orientation errors recorded by Smolka et al. [18] for beetles with a full view of the Milky Way are shown for reference. (a) Orientation errors for each individual under each condition (points), shown with mean vectors (red arrow) and mean direction ± s.d. (red arc), where V tests showed significant orientation. (b) Comparison of the same data in the form of absolute orientation error, such that errors in the clockwise and anticlockwise direction are both shown as positive. Points show the difference in heading between two trials performed by the same individual under the same conditions, and the bar and box show the median and quartile bounds, respectively. When there was an intensity difference between the two light spots, absolute orientation error was significantly lower than when no light spots were displayed, but not when the two light spots were equal in intensity.
None of the pattern-based strategies proposed to date for stellar orientation (e.g. [13,16]) would predict a uniform distribution of errors for an animal observing two points of maximum brightness offset by 180°. Given stable conditions, the recording and matching of salient stimulus features should produce a bimodal distribution of errors. A beetle attempting to match a memorized stellar pattern, centred on a lodestar [16] or the sky's centre of rotation [13,15], would orient well when the observed configuration was, by chance, fixated in its original position and produce 180° errors when the reversed configuration appeared to match the recorded pattern through its symmetric ambiguity. It has previously been suggested that dung beetles may attempt to fixate the brightest point in the celestial hemisphere [18]. This strategy, while less computationally intensive than matching a pattern, would still lead the beetle to fixate either light spot with an equal probability, again resulting in bimodal errors. However, the diurnal species S. lamarcki can also orient using an intensity gradient [22], when observed in combination with solar position or polarized skylight cues. Under the ‘equal intensity’ condition, given that the intensity difference across the axis joining the two light spots was not greater than that in any other axis bisecting the celestial hemisphere, the direction of a matched intensity gradient would not have been a reliable orientation cue. An intensity-gradient matching strategy could, therefore, produce the uniform distribution of errors observed here.
There are a number of factors, however, that may have also contributed to the uniform, rather than bimodal, distribution of errors observed in the presence of the two equally bright light spots. As the light spots were at a fixed distance from the centre of the arena, as opposed to celestial cues, which are situated at an indiscernibly large distance, the relative position, size and intensity of these cues as observed by the beetle change as the beetle moves across the arena. Even so, both the low orientation error in the ‘intensity difference’ condition (figure 4) and recent work on diurnal dung beetles [31,33] show that, when there is an intensity difference or a spectral difference between two lights 180° apart in azimuth, ball-rolling beetles are well oriented in spite of such travel-induced changes in configuration. Travel-induced changes in the apparent size and brightness of each light spot, on the other hand, may pose greater challenges. These changes could occur numerous times during a single trial, producing an effectively uniform distribution of errors. Again, these challenges are also faced by dung beetles orienting under an artificial gradient in intensity or spectrum, and therefore, whatever the cause of this disorientation is, it must be related to the stimulus' symmetric configuration specifically.
In contrast with these results, dung beetles [21,22,32] and other polarization-sensitive animal species (e.g. [34,35]) presented with a single angle of polarization as an orientation cue do produce a bimodal distribution of orientation errors. This presumably results from the fact that a retinal match to the polarization pattern observed in the first trial can be obtained by facing in either direction, since photoreceptor responses follow a 180° periodicity as a function of polarization angle [36]. It may be the case that the dung beetle's compass system allows for symmetric ambiguity in the recognition of polarization cues, but not for patterns, brightest points or broad-field gradients in intensity.
(c). Significance of star patterns
Although the results of the ‘equal intensity’ condition indicated that a generic pattern-recognition strategy is not used by S. satyrus for stellar orientation, this experiment did not exclude the possibility that a Milky Way-like pattern, organized in a band, can be memorized and used for stellar orientation. We propose that such a strategy would involve recording a snapshot of the relative positions of multiple bright sky regions (i.e. bright regions within the band) and using this as a frame of reference for the intended heading. The visual acuity of nocturnal dung beetles [37,38] is probably not sufficient to resolve the spaces between bright stars in the Milky Way band, so the type of patterns that would be available to a dung beetle observing the Milky Way are likely to consist of only broad-scale features. These could be, for example the dark clouds formed by the ‘great rift’ adjacent to the constellation Cygnus and the ‘coalsack’ within the constellation Crux, the ‘large Magellanic cloud’ within the constellation Mensa, and the bright spot between the NCG 3532 open cluster, the constellation Crux and the Southern Pleiades (figure 1a). The last of these was identified by Smolka et al. [18] as a potential fixation point. The ‘high elevation’ and ‘low elevation’ patterns (figure 3c,d) used in this study were designed to provide configurational directional information, while minimising the intensity difference between the two ‘sides’ of the band, which could otherwise be used by the beetles to identify an intensity gradient. When attempting to orient under either of these patterns, orientation error was, however, not significantly clustered around 0° (table 1a). By contrast, the ‘one side lit’ stimulus (figure 3e) produced significantly oriented behaviour (figure 5a; table 1a). For this high-contrast stimulus, absolute orientation error was also significantly lower than either the ‘no lights’ condition (figure 5b; table 2a) or the 90° theoretical disoriented average (table 2b), indicating significantly enhanced orientation accuracy, but this was not the case for either the ‘high elevation’ or ‘low elevation’ pattern stimuli (figure 5b; table 2).
Table 1.
The results of V tests performed on the relative orientation change between two trials for each condition, that is the difference in heading between two trials, in which the same stimulus was displayed rotated by 90° relative to its position in the first trial, minus this 90° rotation (so that this quantity represents the difference in heading relative to the stimulus' position). (a) Shows V tests of the clustering of these orientation errors around 0°, the minimum achievable error. P-Values of less than 0.05 indicate significant orientation relative to the stimulus' position. Means are the direction of the resultant vector of all observations and standard deviations are the natural logarithm of that resultant vector's length divided by observation number, multiplied by −2 and square rooted 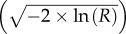 [39]. (Bold: oriented at 0°. ′p<0.10, *p<0.05, **p<0.01, ***p<0.001.) (b) Shows V tests of the orientation errors multiplied by 2 [26] so as to test for orientation to the 0–180° axis. Means and standard deviations are half of their equivalent measure for the same doubled observations. (Bold: oriented at 0° and 180° but not at 0°. ′p<0.10, *p<0.05, **p<0.01, ***p<0.001.
[39]. (Bold: oriented at 0°. ′p<0.10, *p<0.05, **p<0.01, ***p<0.001.) (b) Shows V tests of the orientation errors multiplied by 2 [26] so as to test for orientation to the 0–180° axis. Means and standard deviations are half of their equivalent measure for the same doubled observations. (Bold: oriented at 0° and 180° but not at 0°. ′p<0.10, *p<0.05, **p<0.01, ***p<0.001.
| (a) | ||||
|---|---|---|---|---|
| condition | V (μ = 0°) | p-value | μ ± s.d. | n |
| high elevation pattern | −0.121 | 0.860 | 210.6 ± 113.5° | 40 |
| low elevation pattern | 0.097 | 0.196 | 301.1 ± 104.7° | 39 |
| one side lit, 100% contrast | 0.458 | <0.001*** | 341.3 ± 69.1° | 30 |
| 87.5% contrast | 0.349 | 0.003** | 13.8 ± 82.0° | 30 |
| 20.6% contrast | 0.348 | 0.001** | 354.7 ± 83.1° | 40 |
| 9.5% contrast | 0.104 | 0.257 | 334.7 ± 119.1° | 20 |
| 3.5% contrast | 0.178 | 0.056′ | 335.3 ± 103.4° | 40 |
| 2.8% contrast | −0.015 | 0.552 | 125.3 ± 155.3° | 40 |
| 2.2% contrast | 0.013 | 0.454 | 298.3 ± 153.7° | 40 |
| 1.6% contrast | 0.067 | 0.277 | 292.4 ± 106.6° | 39 |
| 0% contrast | 0.057 | 0.307 | 50.3 ± 126.2° | 40 |
| (b) | ||||
|---|---|---|---|---|
| condition | V (μ=0° and 180°) | p-value | μ and (μ+180) ± s.d. | n |
| high elevation pattern | 0.167 | 0.065′ | 165.6 and 345.6° ± 51.9° | 40 |
| low elevation pattern | 0.095 | 0.201 | 174.0 and 354.0 ± 61.9° | 39 |
| one side lit, 100% contrast | 0.255 | 0.024* | 166.5 and 346.5 ± 45.3° | 30 |
| 87.5% contrast | 0.381 | 0.001** | 177.9 and 357.9 ± 39.7° | 30 |
| 20.6% contrast | 0.018 | 0.436 | 139.1 and 319.1 ± 58.1° | 40 |
| 9.5% contrast | 0.161 | 0.155 | 24.4 and 204.4 ± 48.1° | 20 |
| 3.5% contrast | 0.223 | 0.023* | 2.4 and 182.4 ± 49.6° | 40 |
| 2.8% contrast | 0.376 | <0.001*** | 8.5 and 188.5 ± 39.2° | 40 |
| 2.2% contrast | 0.163 | 0.072' | 1.1 and 181.1 ± 54.5° | 40 |
| 1.6% contrast | 0.110 | 0.167 | 11.4 and 191.4 ± 59.1° | 39 |
| 0% contrast | 0.249 | 0.013* | 11.5 and 191.5 ± 46.4° | 40 |
Figure 5.

Directedness under different band-like patterns. In the ‘high elevation’ pattern light spots were illuminated at 30°, 45°, 75°, 75°, 60° and 30° elevations along the band's north–south axis, while for the ‘low elevation’ pattern only light spots at 45°, 75°, 75° and 30° were illuminated. For the ‘one side lit’ pattern four light spots were again illuminated, this time at 30°, 45°, 60° and 75° on the northern side only. In the ‘no lights’ condition no light spots were illuminated within the set-up. Neither of the ‘high elevation’ nor ‘low elevation’ patterns, which were designed to produce no intensity difference between the two ‘sides’ of the band, contributed significantly to orientation, whereas under the ‘one side lit’ condition the beetles were well oriented. Orientation errors recorded by Smolka et al. [18] for beetles with a full view of the Milky Way shown for reference. (a) Orientation errors for each individual under each condition (points), shown with mean vectors (red arrow) and mean direction ± s.d. (red arc), where V tests showed significant orientation (table 1). (b) Absolute orientation errors. Asterisks denote median error significantly lower than in the ‘no lights’ condition (table 2a). (Online version in colour.)
Table 2.
The results of Mann–Whitney rank-sum tests performed on the absolute orientation error between two trials for each condition, that is the difference in heading between two trials, minus the change in stimulus position, for which values greater than 180° have been converted to their smallest angle from 0°. (a) Shows tests for a true median absolute orientation error lower than that in the ‘no lights’ condition, where orientation error is assumed to be at maximum since no directional cues were available. p-Values of less than 0.05 indicate significantly better orientation than in the absence of any cues. (Bold: true ŷ < no lights. ′p<0.10, *p<0.05, **p<0.01, ***p<0.001.) (b) Shows tests for a true median absolute orientation error lower than 90°, the theoretical average for both a uniform distribution (in the absence of directional cues) and a bimodal distribution (trend towards 180° errors). p-Values of less than 0.05 indicate significantly lower orientation error than the theoretical maximum for these experiments. (Bold: true ŷ < 90°. ′p<0.10, *p<0.05, **p<0.01, ***p<0.001.)
| (a) | ||||
|---|---|---|---|---|
| condition | W (ŷ < no lights) | p-value | ŷ | n |
| high elevation pattern | 423.5 | 0.647 | 105.0 | 40 |
| low elevation pattern | 355.5 | 0.293 | 70.0 | 39 |
| one side lit, 100% contrast | 176.5 | 0.007** | 35.0 | 30 |
| 87.5% contrast | 203.0 | 0.028* | 37.5 | 30 |
| 20.6% contrast | 281.5 | 0.032* | 50.0 | 40 |
| 9.5% contrast | 363.0 | 0.266 | 70.0 | 20 |
| 3.5% contrast | 321.5 | 0.110 | 62.5 | 40 |
| 2.8% contrast | 412.5 | 0.581 | 80.0 | 40 |
| 2.2% contrast | 366.0 | 0.299 | 87.5 | 40 |
| 1.6% contrast | 344.0 | 0.233 | 80.0 | 39 |
| 0% contrast | 345.0 | 0.196 | 100.0 | 40 |
| (b) | ||||
|---|---|---|---|---|
| condition | W (ŷ < 90°) | p-value | ŷ | n |
| no lights | 114.0 | 0.639 | 107.5 | 20 |
| high elevation pattern | 463.5 | 0.647 | 105.0 | 40 |
| low elevation pattern | 312.5 | 0.849 | 70.0 | 39 |
| one side lit, 100% contrast | 80.5 | 0.002** | 35.0 | 30 |
| 87.5% contrast | 120.5 | 0.011* | 37.5 | 30 |
| 20.6% contrast | 165.0 | 0.002** | 50.0 | 40 |
| 9.5% contrast | 81.5 | 0.300 | 70.0 | 20 |
| 3.5% contrast | 264.5 | 0.040* | 62.5 | 40 |
| 2.8% contrast | 466.5 | 0.778 | 80.0 | 40 |
| 2.2% contrast | 347.5 | 0.279 | 87.5 | 40 |
| 1.6% contrast | 326.5 | 0.264 | 80.0 | 39 |
| 0% contrast | 363.0 | 0.266 | 100.0 | 40 |
Since these two patterns were designed to provide only configurational directional information, it is likely that it is again the lack of a sufficient intensity gradient that prevented the beetles from being able to orient. Given that the beetles were well oriented under the ‘one side lit’ condition, it seems unlikely that it was either the absolute intensity of the stimuli, or the shape of the band stimulus itself, that prevented the beetles from orienting. Poor motivation as a result of either factor might also be ruled out, as beetles were capable of rolling their dung balls to the edge of the arena in the absence of any light stimuli (the ‘no lights’ condition) within the same arrangement.
One limitation of these tests is the travel-induced change in light spot position, relative to the beetle, discussed above (see §3b). The largest change in position would occur for the LEDs at 75° elevation, which could change elevation by up to 7° for a beetle travelling from the arena centre to its edge. Since minimum elevation differences between two adjacent LEDs were 15° for the ‘high elevation’ pattern and 30° for the ‘low elevation’ pattern, it might be predicted that these movements would not distort the pattern's configuration beyond recognition for the purposes of orientation. Changes in the distances between the light spots and the beetle's eye, however, the greatest of which would have ranged between ±32 and 220 mm, may well have had disorienting effects of the type suggested for the two equal intensity light spots (see §3b). While we cannot rule out some role for pattern recognition in stellar orientation under more stable natural conditions, these results do suggest that the matching of a remembered configuration is not as robust for this species, at starlight illumination levels, as fixating an intensity difference between two halves of the sky.
(d). Significance of contrast
Taken together, the results of the Symmetric cues (§3b) and Significance of star patterns (§3c) experiments indicated that S. satyrus requires some level of intensity difference across the sky in order to achieve robust orientation under starlight irradiances. This may suggest that intensity ‘gradient’ information is integrated into the compass cues used by S. satyrus, as is the case for related diurnal species S. lamarcki, which can use an intensity gradient in combination with solar position and polarized skylight cues [22]. Since no equivalent cues were available for these experiments or on the moonless nights used in previous studies [17,18], it is likely that a starlight intensity gradient can be used in isolation by S. satyrus. In the context of a starlit sky, an intensity gradient might be identified as the direction followed by the arc (bisecting the sky) along which the maximum intensity difference between the two halves of the sky is observed by the beetle, with all other information about the pattern of brightness across the sky being discarded in the stored representation. In order to determine whether this strategy would be effective under natural levels of intensity difference, the level of contrast between the two halves of a symmetric band was varied across the 0–100% range so that the threshold for accurate orientation could be compared with that recorded via photographic measurements of the Milky Way main band at zenith (figure 1).
Orientation error relative to stimulus position was significantly clustered at 0° for Michelson contrast levels between the two sides of the band of 100% (see §3c above), 87.5% and 20.6% (figure 6a; table 1a). This was not the case for trials conducted under 9.5%, 3.5%, 2.8%, 2.2% and 1.6% contrast (table 1a). This increase in orientation ability as a function of contrast between the two sides was also clear when comparing absolute orientation error (figure 6b). Absolute error was significantly lower for 100–20.6% contrast than in the ‘no lights’ condition (table 2a). For 3.5% contrast, median orientation error was significantly lower than 90°, perhaps suggesting that at least some of the individuals tested were able to use this intensity difference to orient. Nonetheless, for trials at 9.5% contrast and between 2.8 and 0% contrast median absolute orientation errors were not significantly lower than 90° (table 2b). For the 0% contrast condition, in which both sides of the band were of equal intensity, again orientation error was not significantly clustered at 0° (table 1a), but followed a bimodal distribution around the 0–180° axis (table 1b); the same was also true for 2.8% and 3.5% contrast (table 1). The fitted contrast-sensitivity curve (figure 6c) indicated a threshold for orientation using a contrasting symmetric band at 13.0% contrast. At this point, a beetle's capacity to minimize orientation error would be half of its level at 100% contrast. This contrast level is lower than that identified between the northern and southern halves of the Milky Way band at all levels of filtering (figure 1c), with the exception of the 16° filter half-width image, for which maximum contrast was approximately equal to 13%.
Figure 6.

A comparison of directedness under different levels of Michelson contrast between the two halves of an otherwise symmetric band. Significant unimodal orientation (mean orientation error = 0°) was lost at between 20.6% and 9.5% contrast, but, with the exception of the 3.5% and 2.8% contrast conditions, this error did not become significantly bimodally oriented until contrast was less than 1.6%. Median absolute orientation error increased as a function of decreasing contrast, reaching a peak at around 90° for contrasts of less than 3.5% and for the ‘no lights’ condition. These results indicate that sufficient contrast is vital for oriented behaviour under a zenithal band of light. Orientation errors recorded by Smolka et al. [18] for beetles with a full view of the Milky Way shown for reference. (a) Orientation errors for each individual under each condition (points), shown with mean vectors (red arrow) and mean direction ± s.d. (red arc), where V tests showed significant orientation (table 1). (b) Absolute orientation errors. Asterisks denote significantly lower than in the ‘no lights’ condition (table 2a). (c) The relationship between contrast between the two halves of an otherwise symmetric band and absolute orientation error. Points show absolute orientation error plotted against contrast, on a log10 (contrast + 1%) scale. Point size is scaled to show the number of observations at each error angle for each level. The red line shows a generalized linear model fitted to the relationship between absolute orientation error and level of Michelson contrast using maximum-likelihood estimation, via a probit link that has been modified to accept a fixed estimate of upper asymptote [30], at 90°, and an estimate of lower asymptote obtained via an optimisation algorithm (minimum ≈ 59.2°). The inflection point for this curve, the point halfway between the upper and lower asymptote at which the curve's change in slope reaches zero, was selected as an estimate of the threshold level of contrast for orientation under a symmetric band.
The relationship between orientation performance and contrast between two halves of a celestial band presented here suggests that, for a beetle observing the Milky Way's main band as it passes near to the zenith, above-threshold contrasts in broad-field celestial cues should be sufficient to maintain a heading with some degree of accuracy. Since the Milky Way is made up of numerous different star clusters of different brightnesses (figure 1b), the Milky Way's intensity gradient is less predictable than the solar intensity gradient, which is always at its brightest closest to the sun's position. Although not necessarily divided by regular steps in intensity, any given view of the Milky Way band, crossing the zenith or otherwise, will have a brighter and darker half.
As an alternative to using an intensity ‘gradient’, it might also be the case that S. satyrus attempts to fixate the brightest point in the sky, and that the estimated position for this brightest point falls reliably somewhere within the brightest side of a two-level contrasting band. Under an equal-intensity band, with the addition of visual noise, the brightest point detected at any one time could fall anywhere within the band. Hence, estimates of brightest point direction would follow a bimodal distribution. As for the ‘equal intensity’ condition, the direction of an intensity gradient fitted to this light band would be less stable. The tendency towards bimodal orientation for the 0% contrast condition and some, but not all, of the below-threshold contrast conditions might support a ‘brightest point’-over an ‘intensity gradient’-based strategy. It may be the case that the two light spots presented in the ‘equal intensity’ condition were insufficiently broad-field, or indeed insufficiently bright on their own, to be incorporated into this strategy, causing the beetles to become disoriented. Future work may demonstrate that some form of switch or reliability assessment alters the stellar orientation strategy, with the addition of more spatial detail or greater light levels, as in the results presented here. Nevertheless, a ‘brightest point’ strategy fits poorly with both those low contrast conditions in which there was no significant trend towards bimodal orientation (figure 6), and the results of the §3b and Significance of star patterns experiments. For the purposes of this study, it may not be necessary to distinguish between the ‘brightest point’ and ‘intensity gradient’ strategies definitively with regards to Milky Way orientation. Both mechanisms represent a more generalized approach to stellar orientation that is distinct from the pattern-recognition strategies proposed previously in seals [16] and migratory birds [12,13,15].
We propose that these beetles record the azimuth of either the brightest point or strongest intensity gradient observed during the orientation dance, and attempt to match the stored representation to the observed sky as closely as possible. Although it is possible that the celestial compass system of S. satyrus could include a matched filter for either the Milky Way bright spot or the intensity gradient across the Milky Way main band, this might prove unreliable given the potential for variation in the position and visible features of the Milky Way at different times of night and throughout the year. It may, therefore, be more plausible that a generalized system is used to identify the brightest point or direction of an intensity gradient within a particular view of the Milky Way, and that this information is recorded and compared with the current view of the sky while rolling.
(e). Stellar orientation strategies
Of the animals thought to use the stars as an indicator of geographic heading, all those identified to date have been proposed to use some form of pattern matching, either to find a reliably positioned ‘lodestar’ or to fixate the sky's centre of rotation. Given the results of the pattern orientation experiments in this study, it seems unlikely that S. satyrus can use either of these proposed strategies for Milky Way orientation. The types of orientation behaviour performed by migrating birds and itinerant seals do, however, occur over different time-scales from the necessarily-brief ball-rolling behaviour of S. satyrus. The pattern of stars that makes up the Milky Way rotates by only a very small angle over the few minutes that it takes a dung beetle to roll its ball away from the dung pile and begin to bury it. For travel over longer distances, birds and seals, and perhaps also some species of insect that are nocturnal migrants or central place foragers, require strategies that are more robust to gradual celestial rotation.
The band stimulus used in this study (figure 3) is no doubt an imperfect replication of the Milky Way main band as viewed by S. satyrus. It may well be the case, as has recently been shown for lunar orientation [21] and for diurnal species [21,31], that nocturnal dung beetles are able to combine multiple sources of directional information to achieve a high degree of accuracy under starlight conditions. The minimum absolute orientation error for the contrast-sensitivity curve fitted in this study lies at 59.3° (figure 6c), which is considerably greater than typical orientation errors under the Milky Way itself (figure 6a,b). Nonetheless, it may be suggested that contrast between the two halves of a Milky Way band represents a primary orientation cue. Once this intensity gradient has been identified, other forms of directional information could then be added, when available, to improve heading fidelity. This strategy appears well matched to the ocular morphology of S. satyrus and other nocturnal dung beetle species, which are thought to sacrifice spatial acuity to increase absolute sensitivity [37,38]. This trade-off may originate from an evolutionary process in which patterning and other fine-scale details in the retinal image of the celestial hemisphere were sacrificed in favour of broad-field cues, which were sufficiently stable for the purposes of ball-rolling dung beetles. Smolka et al. [18] noted that orienting S. satyrus only outperform S. lamarcki, a diurnal ball-rolling dung beetle species, under shaded full moon and starlight-only conditions. This could be because the dim-light adaptations in the eyes and brain of S. satyrus are mainly effective in the detection of the lunar polarization pattern and the Milky Way, both of which are broad-field cues. The results presented here also suggest that S. satyrus is well adapted for using broad-field cues, and that these adaptations bestow them with sufficient absolute sensitivity and contrast sensitivity under dim-light conditions to achieve accurate orientation using an otherwise symmetrically ambiguous Milky Way band.
4. Conclusion
While other animal species that use the stars as an orientation cue are thought to rely on memorized star patterns, nocturnal dung beetle S. satyrus could use broad-field intensity differences across the sky as their primary stellar orientation cue. This strategy is likely to be both robust to small changes in atmospheric conditions and flexible across different star patterns visible at different times of year. It may also be well suited to S. satyrus’ visual system, which probably sacrifices some visual acuity for the absolute sensitivity necessary for reliable detection of broad-field starlight cues.
In order to define the stellar orientation strategy of S. satyrus more completely, future studies should attempt to manipulate the perceived direction of the intensity gradient, and the position of the brightest point, in the Milky Way main band by obscuring specific regions and adding artificial light stimuli. In combination with versions of the experiments performed as part of this study, these techniques may be developed to form an assay for different stellar orientation strategies.
Supplementary Material
Supplementary Material
Acknowledgements
The authors thank Peter Olsson, Anna Stöckl, Emily Baird, Esa-Ville Immonen and Eric Warrant for their helpful discussions of the work and data. We also thank James Harrison, and Ted and Winnie Harvey for their help in the field.
Ethics
All experiments were performed in accordance with Swedish and South African guidelines for animal experiments.
Data accessibility
Tables of all exit headings are available as the electronic supplementary material. All orientation error data reported here, and the exit headings from which they were calculated, are available as the electronic supplementary material.
Authors' contributions
J.J.F., B.e.J. and M.D. designed the behavioural experiments. J.J.F., B.e.J., L.K., M.J.B. and M.D. conducted experiments. J.J.F. analysed the behavioural data. J.S. and D.-E.N. designed the sky imaging protocol, J.S. analysed the sky imaging data. J.J.F. drafted the manuscript. J.J.F., B.e.J., J.S., L.K., D.-E.N., M.J.B. and M.D. revised the manuscript.
Competing interests
The authors declare no competing interests.
Funding
Funding was provided by the Wallenberg Foundation, Carl Trygger's Foundation for Scientific Research (CTS 14:95 and 15:108), the Royal Physiographic Society of Lund, the Lars-Hiertas Minne Foundation (FO2015-0540), the Swedish Research Council and the South African National Research Foundation.
References
- 1.Gould JL, Gould CG. 2012. Nature's compass—the mystery of animal navigation. Oxford, UK: Princeton University Press. [Google Scholar]
- 2.Baker RR. 1968. Sun orientation during migration in some British butterflies. Proc. R. Entomol. Soc. A 143, 89–95. ( 10.1111/j.1365-3032.1968.tb01033.x) [DOI] [Google Scholar]
- 3.Mouritsen H, Frost BJ. 2002. Virtual migration in tethered flying monarch butterflies reveals their orientation mechanisms. Proc. Natl Acad. Sci. USA 99, 10 162–10 166. ( 10.1073/pnas.152137299) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Schmidt-Koenig K. 1990. The sun compass. Experientia 46, 338–342. ( 10.1007/BF01952166) [DOI] [Google Scholar]
- 5.Schwassmann HO, Hasler AD. 1964. The role of the sun's altitude in sun orientation of fish. Physiol. Zool. 37, 163–178. ( 10.1086/physzool.37.2.30152329) [DOI] [Google Scholar]
- 6.Santschi F. 1923. Messor et autres fourmis paléarctiques. Rev. Suisse Zool. 30, 317–336. [Google Scholar]
- 7.Müller M, Wehner R. 1988. Path integration in desert ants, Cataglyphis fortis. Proc. Natl Acad. Sci. USA 85, 5287–5290. ( 10.1073/pnas.85.14.5287) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.von Frisch K. 1974. Decoding the language of the bee. Science 185, 663–668. ( 10.1126/science.185.4152.663) [DOI] [PubMed] [Google Scholar]
- 9.Evangelista C, Kraft P, Dacke M, Labhart T, Srinivasan MV. 2014. Honeybee navigation: critically examining the role of the polarization compass. Phil. Trans. R. Soc. B 369, 20130037 ( 10.1098/rstb.2013.0037) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Landreth HF, Ferguson DE. 1968. The Sun Compass of Fowler's Toad, Bufo woodhousei fowleri. Behaviour 30, 27–43. ( 10.1163/156853968X00162) [DOI] [Google Scholar]
- 11.Ferguson DE, Landreth HF, Turnipseed MR. 1965. Astronomical orientation of the southern cricket frog, Acris gryllus. Copeia 1965, 58–66. ( 10.2307/1441240) [DOI] [Google Scholar]
- 12.Emlen S. 1967. Migratory orientation in the Indigo Bunting, Passerina cyanea. Part I: evidence for use of celestial cues. Auk 84, 309–342. ( 10.2307/4083084) [DOI] [Google Scholar]
- 13.Emlen S. 1967. Migratory orientation in the Indigo Bunting, Passerina cyanea. Part II: mechanism of celestial orientation. Auk 84, 463–489. ( 10.2307/4083330) [DOI] [Google Scholar]
- 14.Sotthibandhu S, Baker RR. 1979. Celestial orientation by the large yellow underwing moth, Noctua pronuba L. Anim. Behav. 27, 786–800. ( 10.1016/0003-3472(79)90015-0) [DOI] [Google Scholar]
- 15.Mouritsen H, Larsen ON. 2001. Migrating songbirds tested in computer-controlled Emlen funnels use stellar cues for a time-independent compass. J. Exp. Biol. 204, 3855–3865. [DOI] [PubMed] [Google Scholar]
- 16.Mauck B, Gläser N, Schlosser W, Dehnhardt G. 2008. Harbour seals (Phoca vitulina) can steer by the stars. Anim. Cogn. 11, 715–718. ( 10.1007/s10071-008-0156-1) [DOI] [PubMed] [Google Scholar]
- 17.Dacke M, Baird E, Byrne M, Scholtz CH, Warrant EJ. 2013. Dung beetles use the Milky Way for orientation. Curr. Biol. 23, 298–300. ( 10.1016/j.cub.2012.12.034) [DOI] [PubMed] [Google Scholar]
- 18.Smolka J, Baird E, el Jundi B, Reber T, Byrne MJ, Dacke M. 2016. Night sky orientation with diurnal and nocturnal eyes: dim-light adaptations are critical when the moon is out of sight. Anim. Behav. 111, 127–146. ( 10.1016/j.anbehav.2015.10.005) [DOI] [Google Scholar]
- 19.Feinberg R. 1988. Polynesian seafaring and navigation. Kent, OH: The Kent State University Press. [Google Scholar]
- 20.Dacke M, Byrne MJ, Baird E, Scholtz CH, Warrant EJ. 2011. How dim is dim? Precision of the celestial compass in moonlight and sunlight. Phil. Trans. R. Soc. B 366, 697–702. ( 10.1098/rstb.2010.0191) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.el Jundi B, Warrant EJ, Byrne MJ, Khaldy L, Baird E, Smolka J, Dacke M. 2015. Neural coding underlying the cue preference for celestial orientation. Proc. Natl Acad. Sci. USA 112, 11 395–11 400. ( 10.1073/pnas.1501272112) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.el Jundi B, Smolka J, Baird E, Byrne MJ, Dacke M. 2014. Diurnal dung beetles use the intensity gradient and the polarization pattern of the sky for orientation. J. Exp. Biol. 217, 2422–2429 ( 10.1242/jeb.101154) [DOI] [PubMed] [Google Scholar]
- 23.Smolka J, Nilsson D-E. In preparation. The visual environment plot standard—a new tool for characterising light in habitats.
- 24.Land MF. 1981. Optics and vision in invertebrates. In Handbook of sensory physiology VII/6B (ed. Autrum H.), pp. 471–592. Berlin, Germany: Springer. [Google Scholar]
- 25.Labhart T. 1980. Specialized photoreceptors at the dorsal rim of the honeybee's compound eye: polarizational and angular sensitivity. J. Comp. Physiol. A 141, 19–30. ( 10.1007/BF00611874) [DOI] [Google Scholar]
- 26.Batschelet E. 1981. Circular statistics in biology. London, UK: Academic Press. [Google Scholar]
- 27.Lund U, Agostinelli C. 2014. Package ‘circular’. version 0.4-7. See https://CRAN.R-project.org/package=circular.
- 28.R Development Core Team. 2016. R: a language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. [Google Scholar]
- 29.Dacke M, el Jundi B, Smolka J, Byrne M, Baird E. 2014. The role of the sun in the celestial compass of dung beetles. Phil. Trans. R. Soc. B 369, 20130036 ( 10.1098/rstb.2013.0036) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Knoblauch K. 2014. psyphy: functions for analyzing psychophysical data in R. version 0.1-9. See https://CRAN.R-project.org/package=psyphy.
- 31.el Jundi B, Foster JJ, Khaldy L, Byrne MJ, Dacke M, Baird E. 2016. A snapshot-based mechanism for celestial orientation. Curr. Biol. 26, 1456–1462. ( 10.1016/j.cub.2016.03.030) [DOI] [PubMed] [Google Scholar]
- 32.Dacke M, Nilsson D-E, Scholtz CH, Byrne M, Warrant EJ. 2003. Insect orientation to polarized moonlight. Nature 424, 33 ( 10.1038/424033a) [DOI] [PubMed] [Google Scholar]
- 33.el Jundi B, Foster JJ, Byrne MJ, Baird E, Dacke M. 2015. Spectral information as an orientation cue in dung beetles. Biol. Lett. 11, 20150656 ( 10.1098/rsbl.2015.0656) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Jander R. 1957. Die optische Richtungsorientierung der Roten Waldameise (Formica rufa L.). J. Comp. Physiol. 40, 162–238. ( 10.1007/BF00297947) [DOI] [Google Scholar]
- 35.Novales Flamarique I, Browman HI. 2000. Wavelength-dependent polarization orientation in Daphnia. J. Comp. Physiol. A 186, 1073–1087. ( 10.1007/s003590000162) [DOI] [PubMed] [Google Scholar]
- 36.Bernard GD, Wehner R. 1977. Functional similarities between polarization vision and color vision. Vis. Res. 17, 1019–1028. ( 10.1016/0042-6989(77)90005-0) [DOI] [PubMed] [Google Scholar]
- 37.Caveney S, McIntyre P. 1981. Design of graded-index lenses in the superposition eyes of scarab beetles. Phil. Trans. R. Soc. B 294, 589–632. ( 10.1098/rstb.1981.0119) [DOI] [Google Scholar]
- 38.Warrant EJ, McIntyre PD. 1990. Limitations to resolution in superposition eyes. J. Comp. Physiol. A 167, 785–803. ( 10.1007/BF00189768) [DOI] [Google Scholar]
- 39.Mardia KV. 1972. Statistics of directional data. sec. 26.5, p. 617 London, UK: Academic Press. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Tables of all exit headings are available as the electronic supplementary material. All orientation error data reported here, and the exit headings from which they were calculated, are available as the electronic supplementary material.