

Abstract
Diabetogenic single nucleotide polymorphisms (SNPs) have recently been associated with multiple myeloma (MM) risk but their impact on overall survival (OS) of MM patients has not been analysed yet. In order to investigate the impact of 58 GWAS-identified variants for type 2 diabetes (T2D) on OS of patients with MM, we analysed genotyping data of 936 MM patients collected by the International Multiple Myeloma rESEarch (IMMENSE) consortium and an independent set of 700 MM patients recruited by the University Clinic of Heidelberg. A meta-analysis of the cox regression results of the two sets showed that rs7501939 located in the HNF1B gene negatively impacted OS (HRRec= 1.44, 95% CI = 1.18–1.76, P = 0.0001). The meta-analysis also showed a noteworthy gender-specific association of the SLC30A8rs13266634 SNP with OS. The presence of each additional copy of the minor allele at rs13266634 was associated with poor OS in men whereas no association was seen in women (HRMen-Add = 1.32, 95% CI 1.13–1.54, P = 0.0003). In conclusion, these data suggest that the HNF1Brs7501939 SNP confers poor OS in patients with MM and that a SNP in SLC30A8 affect OS in men.
Keywords: multiple myeloma, diabetes, genetic variants, survival
INTRODUCTION
Multiple myeloma is an incurable and heterogeneous plasma cell neoplasm that affects about 6.3 per 100.000 people per year worldwide (i.e., 25.850 new cases only in 2015) and represents 1.6% of all cancers and 2% of all cancer deaths [1]. In spite of the widespread use of proteasome inhibitors and immunomodulatory drugs, which has dramatically improved the life expectancy of MM patients over the last few decades [2, 3], MM survival still remains poor with a 5-year survival of 46.6% (SEER Cancer Statistics Review, http://seer.cancer.gov/csr/1975_2012/).
Epidemiological and observational studies have consistently identified several factors that affect MM patient survival such as age at diagnosis [4, 5], stage at diagnosis (coded by either the Durie-Salmon staging system (DSS) [6] or the International Staging System (ISS)) [7], Eastern Cooperative Oncology Group (ECOG) performance status [8], renal failure [9, 10], high plasma cell proliferative rate [11, 12], high lactate deshydrogenase (LDH) levels [13] and chromosomal abnormalities [14–18]. Increasing evidences point towards a positive correlation of pre-existing type 2 diabetes (T2D) with MM risk [19] but also with the appearance of severe clinical complications [20–23] and patient survival [24, 25]. In this regard, Chiu et al. (2006) reported that high level of postload glucose was associated with increased risk of mortality in hematological malignancies [24] whereas Chou et al. (2012) reported that MM patients with pre-existing T2D have 50% higher all-cause mortality compared with non-diabetic patients [25]. These observations might be explained, at least in part, by the stimulatory effects of T2D-associated hyperglycaemia, insulin resistance and resulting hyperinsulinemia on MM cell growth [26, 27] but also by the deregulation of tumour-suppressor genes linked to T2D (such as CDKN2A-2B, KCNQ1, HNF1B) [28–30] that might lead to uncontrolled cell proliferation, cell differentiation and disease progression and, consequently, to shorter survival periods. In support of this notion, CDKN2A-2B genes have been found to be frequently hypermethylated in MM [31–33] whereas loss of expression of KCNQ1 has been associated with poor overall survival in cancer patients [34]. Furthermore, emerging evidences also suggest that the activation of certain T2D-related genes (such as NOTCH2) may induce MM cell migration from the infiltrated site to different bone marrow districts [35] and promote osteoclast formation [36], which is a process intimately related to proliferation and long-term survival of MM cells [37].
Although germline variants may influence the susceptibility of MM [38–43] and survival [39, 44–46], the knowledge regarding the role of diabetogenic variants in modulating the risk of MM and survival remains scarce. We have recently reported that diabetogenic variants influence MM risk [47] and recent genome-wide association studies (GWAS) have also suggested the involvement of genetic variants within the MTHFD1L, AKAP12 and FOPNL loci in determining MM patient survival [39, 44, 45] but also an indirect implication of diabetogenic genes such as TCF7L2 [45]. Johnson et al. (2016) reported in their GWAS a strong association of rs12374648, which maps to a binding site for the transcription factor TCF7L2, with MM overall survival (OS) and proposed a functional mechanism of this variant to modulate the synthesis of purines and the regulation of cell cycle [45].
Based on these findings, we explored for the first time the relationship between diabetogenic variants and OS of MM patients in a study developed in the context of the International Multiple Myeloma rESEarch (IMMENSE) consortium. We attempted to confirm our findings by analysing GWAS data on an independent set of German MM patients (Heidelberg cohort) [45].
RESULTS
The demographic and clinical characteristics of the MM patients included in the IMMENSE (n = 939) and Heidelberg (n = 700) cohorts are listed in Table 1. The median age at diagnosis was similar in both populations (59.73 ± 10.08 vs. 55.85 ± 8.33) but the male/female ratio was higher in the Heidelberg cohort (1.36 vs. 1.06). Durie-Salmon stage was available for IMMENSE and Heidelberg cohorts and included patients at stages I, II and III (11.83%, 23.54% and 64.63% vs. 0.8%, 12.4% and 86.8%, respectively).
Table 1. Clinical characteristics of IMMEnSE and Heidelberg cohorts.
| IMMENSE population | |||
|---|---|---|---|
| Country of origin | MM patients (n = 936) | ||
| Gender M/F (Total) | Mean Age (± STD) | Median Age (Range) | |
| Italy | 69/69 (138) | 61.31 ± 9.47 | 51.0 (35–86) |
| Poland | 145/163 (308) | 62.48 ± 10.50 | 52.0 (34–86) |
| Spain | 49/55 (104) | 62.44 ± 11.45 | 66.0 (22–88) |
| France | 42/33 (75) | 55.80 ± 9.04 | 41.0 (34–75) |
| Portugal | 14/22 (36) | 65.22 ± 9.54 | 35.0 (45–80) |
| Denmark | 163/112 (275) | 55.18 ± 7.32 | 51.0 (29–69) |
| Demographic variables | |||
| Age (years, average ± SD) | 59.73 ± 10.08 | ||
| Sex ratio (male/female) | 1.06 (482/454) | ||
| Overall survival (months) | 99.69 [92.98,106.39] | ||
| Number of deaths | 323 | ||
| Median follow-up time (months) | 100 (52–111) | ||
| Disease stage (Durie-Salmon)* | |||
| Stage I | 93 (11.83) | ||
| Stage II | 185 (23.54) | ||
| Stage III | 508 (64.63) | ||
| Heidelberg population | |||
|---|---|---|---|
| MM patients (n = 700) | |||
| Country of origin | Gender M/F (Total) | Mean Age (± STD) | Median Age (Range) |
| Germany (HD3 trial) | 56/42 (98) | 55.42 ± 7.47 | 56.5 (38–65) |
| Germany (HD4 trial) | 170/121 (291) | 55.02 ± 7.30 | 57.0 (27–65) |
| Germany (non-trial) | 178/133 (311) | 56.76 ± 9.37 | 57.2 (24–73) |
| Demographic variables | |||
| Age (years, average ± SD) | 55.85 ± 8.33 | ||
| Sex ratio (male/female) | 1.36 (404/296) | ||
| Overall survival (months)∂ | 91.2 [85.5,105] | ||
| Number of deaths | 326 | ||
| Median follow-up time (months) | 84.3 (80–88) | ||
| Disease stage (Durie-Salmon) | |||
| Stage I | 5 (0.8) | ||
| Stage II | 82 (12.4) | ||
| Stage III | 575 (86.8) | ||
Abbreviations: IMMENSE, International Multiple Myeloma rESEarch; MM, Multiple Myeloma; SD, standard deviation.
Durie-Salmon data was not available for 150 MM patients.
Median overall survival after diagnosis (IMMENSE) or the 1st autotransplant (Heidelberg cohort) (KM estimators).
All SNPs tested showed genotype frequencies consistent with the HWE (P > 0.001) and the observed allele frequency for all selected SNPs was in accordance with Hapmap data. When we evaluated the effect of selected polymorphisms on MM OS in the IMMENSE population, we found that 6 SNPs showed a noteworthy association with OS. The most relevant effect was observed for the HNF1Brs7501939 SNP that was associated with poor OS when recessive and log-additive models of inheritance were assumed (HRREC = 1.49, 95% CI 1.11–2.00, P = 0.008 and HRADD = 1.34 95% CI 1.13–1.59, P = 0.001, respectively; Table 2). Patients harbouring the non-diabetogenic HNF1Brs7501939T/T genotype showed a median survival time (MST) significantly shorter than those carrying the C allele (MSTT/T = 81.91 months vs. MSTC/C+C/T = 101.42 months; Figure 1A). This result was confirmed with the 700 MM patients recruited from the University Clinic of Heidelberg (HRREC = 1.40, 95% CI 1.06–1.84 and MSTT/T = 74.4 months vs. MSTC/C+C/T = 97.2 months; Table 3 and Figure 1B). The result of the meta-analysis for this SNP remained significant after correction for multiple testing (HRMeta-Rec = 1.44, 95% CI 1.18–1.76, P = 0.0001, I2 = 0.0%, PHet = 0.74; Table 3). According to publicly available eQTL data for human peripheral blood mononuclear cells, the risk allele (T) was associated with higher HNF1B mRNA expression levels (Z score = 3.31, P = 9.23·10–4 and FDR = 0.23). However, we could not validate this finding using eQTL data on plasma cells from 665 German MM patients (P = 0.60; Supplementary Table S1). Nonetheless, according to Haploreg and ENCODE annotation data, the rs7501939 SNP resides near of a poised promoter in many cell lines including a lymphoblastoid and human stem cell lines (GM12878 and H1-HSCs) that might be rapidly activated upon specific stimuli. In addition, this SNP was predicted to change binding motifs for 2 regulatory transcription factors (CEBPB and p300) and mapped among enhancer histone marks in primary naïve and memory forms of cytotoxic T cells (CD8+) and helper T cells (CD4+) from peripheral blood.
Table 2. Association of T2D–related variants and overall survival (OS) of MM patients.
| Variant_dbSNP | Gene | OVERALL (N = 936) | MEN (N = 482) | WOMEN (N= 454) | ||||
|---|---|---|---|---|---|---|---|---|
| OR (95% CI)a | Pvalue | OR (95% CI)b | Pvalue | OR (95% CI)b | Pvalue | PInteraction | ||
| rs2641348 | ADAM30 | 0.94 (0.69–1.28) | 0.69 | 0.99 (0.67–1.48) | 0.97 | 0.87 (0.53–1.41) | 0.56 | 0.70 |
| rs4607103† | ADAMTS9 | 1.23 (0.76–2.00) | 0.40 | 0.83 (0.42–1.63) | 0.58 | 2.53 (1.26–5.07) | 0.009 | 0.024 |
| rs11708067 | ADCY5 | 0.87 (0.68–1.11) | 0.25 | 1.02 (0.75–1.39) | 0.89 | 0.63 (0.42–0.95) | 0.028 | 0.08 |
| rs10885122 | ADRA2A | 1.03 (0.79–1.33) | 0.83 | 1.01 (0.72–1.40) | 0.97 | 1.07 (0.71–1.62) | 0.73 | 0.89 |
| rs1552224 | ARAPI, CENTD2 | 0.88 (0.68–1.15) | 0.35 | 0.90 (0.64–1.26) | 0.55 | 0.83 (0.54–1.25) | 0.37 | 0.77 |
| rs10490072 | BCL11A | 1.05 (0.83–1.32) | 0.70 | 1.22 (0.90–1.65) | 0.19 | 0.85 (0.59–1.23) | 0.39 | 0.11 |
| rs12779790 | CDC123, CAMK1D | 0.86 (0.67–1.11) | 0.24 | 1.06 (0.77–1.46) | 0.72 | 0.64 (0.43–0.97) | 0.035 | 0.06 |
| rs7754840 | CDKAL1 | 1.14 (0.90–1.44) | 0.27 | 1.22 (0.90–1.65) | 0.20 | 1.06 (0.74–1.53) | 0.75 | 0.52 |
| rs564398† | CDKN2A–2B | 0.64 (0.42–0.98) | 0.042 | 0.62 (0.36–1.07) | 0.084 | 0.71 (0.34–1.47) | 0.36 | 0.88 |
| rs10811661 | CDKN2A–2B | 0.93 (0.72–1.19) | 0.55 | 0.93 (0.67–1.30) | 0.68 | 0.92 (0.63–1.35) | 0.67 | 0.92 |
| rs2383208 | CDKN2A–2B | 0.94 (0.73–1.21) | 0.64 | 0.92 (0.66–1.29) | 0.64 | 0.95 (0.65–1.41) | 0.81 | 0.87 |
| rs4240702 | COL5A1 | 0.88 (0.68–1.15) | 0.35 | 0.85 (0.60–1.21) | 0.36 | 0.95 (0.63–1.44) | 0.82 | 0.65 |
| rs11605924 | CRY2 | 0.91 (0.70–1.18) | 0.47 | 0.81 (0.58–1.14) | 0.23 | 1.09 (0.71–1.69) | 0.68 | 0.29 |
| rs1153188 | DCD | 1.18 (0.94–1.48) | 0.16 | 1.25 (0.92–1.69) | 0.15 | 1.09 (0.76–1.57) | 0.63 | 0.58 |
| rs1113132 | EXT2 | 1.02 (0.81–1.28) | 0.86 | 0.96 (0.71–1.30) | 0.79 | 1.08 (0.75–1.56) | 0.66 | 0.57 |
| rs174550 | FADS1 | 1.00 (0.79–1.26) | 1.00 | 0.97 (0.72–1.30) | 0.82 | 1.05 (0.73–1.51) | 0.80 | 0.71 |
| rs11071657 | FAM148B | 0.86 (0.68–1.09) | 0.22 | 0.96 (0.70–1.32) | 0.81 | 0.77 (0.53–1.12) | 0.17 | 0.44 |
| rs17044137 | FLJ39370 | 1.06 (0.84–1.33) | 0.64 | 1.07 (0.79–1.44) | 0.67 | 1.03 (0.71–1.50) | 0.87 | 0.92 |
| rs8050136 | FTO | 0.91 (0.70–1.18) | 0.46 | 0.81 (0.58–1.13) | 0.21 | 1.10 (0.72–1.67) | 0.66 | 0.23 |
| rs560887† | G6PC2 | 1.11 (0.74–1.66) | 0.61 | 0.78 (0.44–1.39) | 0.40 | 1.76 (1.00–3.09) | 0.050 | 0.045 |
| rs1799884 | GCK | 0.99 (0.76–1.28) | 0.92 | 1.10 (0.80–1.52) | 0.57 | 0.81 (0.52–1.26) | 0.36 | 0.39 |
| rs1260326† | GCKR | 1.36 (1.01–1.82) | 0.043 | 1.24 (0.84–1.83) | 0.28 | 1.53 (0.97–2.42) | 0.068 | 0.47 |
| rs1111875 | HHEX | 1.00 (0.78–1.27) | 0.97 | 0.92 (0.67–1.26) | 0.61 | 1.19 (0.80–1.77) | 0.38 | 0.33 |
| rs7957197 | HNF1A (TCF1) | 1.17 (0.92–1.48) | 0.20 | 1.18 (0.87–1.59) | 0.30 | 1.16 (0.80–1.70) | 0.43 | 0.89 |
| rs7501939† | HNF1B (TCF2) | 1.49 (1.11–2.00) | 0.008 | 1.49 (1.03–2.17) | 0.036 | 1.49 (0.91–2.45) | 0.11 | 0.99 |
| rs35767 | IGF1 | 0.87 (0.67–1.12) | 0.27 | 0.78 (0.56–1.10) | 0.16 | 1.00 (0.68–1.48) | 1.00 | 0.40 |
| rs4402960 | IGF2BP2 | 0.90 (0.71–1.13) | 0.36 | 0.78 (0.58–1.06) | 0.11 | 1.10 (0.76–1.60) | 0.60 | 0.16 |
| rs20541† | IL13 | 1.42 (0.89–2.27) | 0.14 | 0.93 (0.45–1.90) | 0.84 | 2.38 (1.24–4.57) | 0.009 | 0.07 |
| rs2943641 | IRS1 | 1.13 (0.89–1.43) | 0.33 | 1.01 (0.75–1.38) | 0.93 | 1.38 (0.94–2.02) | 0.10 | 0.30 |
| rs864745 | JAZF1 | 0.90 (0.71–1.15) | 0.40 | 1.01 (0.73–1.39) | 0.98 | 0.75 (0.52–1.09) | 0.14 | 0.22 |
| rs5215 | KCNJ11 | 1.09 (0.86–1.37) | 0.49 | 1.39 (1.02–1.90) | 0.038 | 0.79 (0.55–1.13) | 0.20 | 0.022 |
| rs5219 | KCNJ11 | 1.11 (0.87–1.43) | 0.39 | 1.37 (0.98–1.91) | 0.063 | 0.88 (0.60–1.28) | 0.50 | 0.12 |
| rs2237897 | KCNQ1 | 1.25 (0.88–1.77) | 0.21 | 1.28 (0.81–2.01) | 0.28 | 1.22 (0.71–2.10) | 0.48 | 0.83 |
| rs2074196 | KCNQ1 | 1.57 (1.03–2.40) | 0.036 | 1.83 (1.01–3.32) | 0.047 | 1.42 (0.77–2.59) | 0.26 | 0.48 |
| rs2237892 | KCNQ1 | 1.38 (0.97–1.97) | 0.070 | 1.53 (0.97–2.41) | 0.065 | 1.24 (0.70–2.18) | 0.46 | 0.53 |
| rs2237895 | KCNQ1 | 1.06 (0.82–1.36) | 0.66 | 1.30 (0.94–1.80) | 0.11 | 0.74 (0.50–1.11) | 0.15 | 0.033 |
| rs231362 | KCNQ1OT1 | 1.15 (0.88–1.51) | 0.31 | 1.04 (0.73–1.47) | 0.84 | 1.34 (0.87–2.07) | 0.19 | 0.33 |
| rs1041981 | LTA | 0.86 (0.68–1.09) | 0.21 | 0.76 (0.56–1.04) | 0.088 | 1.04 (0.71–1.51) | 0.85 | 0.25 |
| rs7944584† | MADD | 0.68 (0.46–1.01) | 0.058 | 0.55 (0.32–0.95) | 0.031 | 0.83 (0.46–1.52) | 0.55 | 0.27 |
| rs12970134 | MCR4 | 0.89 (0.70–1.12) | 0.31 | 0.86 (0.64–1.16) | 0.31 | 0.94 (0.65–1.35) | 0.74 | 0.71 |
| rs1387153 | MTNR1B | 0.89 (0.71–1.12) | 0.32 | 1.01 (0.75–1.37) | 0.93 | 0.71 (0.49–1.02) | 0.063 | 0.16 |
| rs10923931 | NOTCH2 | 0.98 (0.73–1.31) | 0.88 | 1.05 (0.72–1.53) | 0.81 | 0.88 (0.55–1.41) | 0.59 | 0.58 |
| rs6698181 | PKN2 | 1.17 (0.92–1.48) | 0.20 | 1.28 (0.94–1.75) | 0.12 | 1.00 (0.69–1.46) | 0.98 | 0.33 |
| rs1801282 | PPARG | 0.84 (0.65–1.10) | 0.21 | 0.66 (0.46–0.96) | 0.030 | 1.12 (0.76–1.65) | 0.56 | 0.09 |
| rs8042680† | PRC1 | 0.91 (0.64–1.29) | 0.60 | 1.13 (0.74–1.73) | 0.56 | 0.56 (0.29–1.07) | 0.081 | 0.08 |
| rs340874 | PROX1 | 1.03 (0.78–1.36) | 0.83 | 0.75 (0.52–1.07) | 0.11 | 1.60 (1.02–2.50) | 0.041 | 0.016 |
| rs7593730 | RBMS1 | 0.92 (0.73–1.16) | 0.48 | 0.77 (0.56–1.05) | 0.10 | 1.20 (0.83–1.75) | 0.33 | 0.07 |
| rs1531343 | RPSAP52, HMGA2 | 1.11 (0.84–1.46) | 0.45 | 1.40 (1.00–1.97) | 0.053 | 0.76 (0.46–1.23) | 0.26 | 0.07 |
| rs11920090 | SLC2A2 | 0.88 (0.67–1.14) | 0.34 | 0.87 (0.62–1.22) | 0.41 | 0.91 (0.59–1.40) | 0.67 | 0.91 |
| rs13266634∂ | SLC30A8 | 1.24 (1.05–1.47) | 0.011 | 1.42 (1.14–1.77) | 0.002 | 1.01 (0.77–1.33) | 0.94 | 0.057 |
| rs7903146 | TCF7L2 | 0.83 (0.66–1.05) | 0.12 | 0.72 (0.53–0.97) | 0.030 | 1.04 (0.72–1.50) | 0.82 | 0.13 |
| rs12255372 | TCF7L2 | 0.87 (0.69–1.09) | 0.23 | 0.73 (0.54–0.98) | 0.039 | 1.14 (0.79–1.65) | 0.49 | 0.08 |
| rs7578597 | THADA | 1.18 (0.88–1.58) | 0.27 | 0.97 (0.67–1.41) | 0.88 | 1.59 (0.99–2.55) | 0.054 | 0.12 |
| rs896854† | TP53INP1 | 0.76 (0.58–1.00) | 0.050 | 0.74 (0.52–1.06) | 0.10 | 0.77 (0.50–1.19) | 0.24 | 0.81 |
| rs7961581 | TSPAN8, LGR5 | 1.09 (0.87–1.37) | 0.47 | 1.08 (0.80–1.47) | 0.62 | 1.14 (0.79–1.63) | 0.49 | 0.77 |
| rs9472138 | VEGFA | 1.03 (0.82–1.30) | 0.80 | 0.93 (0.69–1.26) | 0.63 | 1.19 (0.83–1.71) | 0.33 | 0.32 |
| rs10010131 | WFS1 | 1.00 (0.78–1.28) | 0.99 | 1.15 (0.81–1.61) | 0.44 | 0.85 (0.59–1.23) | 0.38 | 0.28 |
Abbreviations: SNP, single nucleotide polymorphism; OR, odds ratio; CI, confidence interval; n/s, not specified.
Estimates were adjusted for age, sex, country of origin and Durie–Salmon stage.P < 0.05 in bold.
Estimates calculated according to a dominant model of inheritance.
Estimates calculated according to a dominant model of inheritance and adjusted for age, region and Durie–Salmon stage.
Estimates calculated according to a recessive model of inheritance.
Estimates calculated according to an additive model of inheritance.
Figure 1. Kaplan-Meier plots for the HNF1Brs7501939 SNP in the IMMENSE (A) and Heidelberg (B) populations.
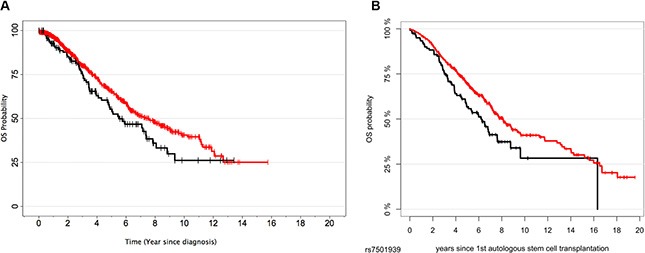
Table 3. Meta–analysis for the association of T2D–related variants and overall survival (OS) of MM patients.
| IMMENSE (N = 936) | GWAS (N = 700) | META-ANALYSIS (N = 1636) | |||||
|---|---|---|---|---|---|---|---|
| Variant_dbSNP | Gene | OR (95% CI)a | Pvalue | HR (95% CI)b | Pvalue | HR (95% CI)c | Pvalue |
| rs2641348 | ADAM30 | 0.94 (0.69–1.28) | 0.69 | 1.12 (0.86–1.45) | 0.41 | 1.04 (0.85–1.27) | 0.69 |
| rs4607103† | ADAMTS9 | 1.23 (0.76–2.00) | 0.40 | 0.99 (0.63–1.55)* | 0.98 | 1.10 (0.79–1.52)* | 0.59 |
| rs11708067 | ADCY5 | 0.87 (0.68–1.11) | 0.25 | 0.89 (0.71–1.11)* | 0.31 | 0.88 (0.75–1.04)* | 0.13 |
| rs10885122 | ADRA2A | 1.03 (0.79–1.33) | 0.83 | 0.85 (0.65–1.12) | 0.24 | 0.94 (0.78–1.13) | 0.52 |
| rs1552224 | ARAPI, CENTD2 | 0.88 (0.68–1.15) | 0.35 | 1.12 (0.89–1.42) | 0.34 | 1.00 (0.79–1.27) | 1.00 |
| rs10490072 | BCL11A | 1.05 (0.83–1.32) | 0.70 | 1.30 (1.04–1.62)* | 0.019 | 1.17 (0.95–1.44)* | 0.14 |
| rs12779790 | CDC123, CAMK1D | 0.86 (0.67–1.11) | 0.24 | ND | ND | ND | ND |
| rs7754840 | CDKAL1 | 1.14 (0.90–1.44) | 0.27 | 1.15 (0.92–1.43) | 0.22 | 1.14 (0.98–1.35) | 0.10 |
| rs564398† | CDKN2A–2B | 0.64 (0.42–0.98) | 0.042 | 1.17 (0.88–1.55) | 0.29 | 0.88 (0.49–1.59) | 0.68 |
| rs10811661 | CDKN2A–2B | 0.93 (0.72–1.19) | 0.55 | ND | ND | ND | ND |
| rs2383208 | CDKN2A–2B | 0.94 (0.73–1.21) | 0.64 | ND | ND | ND | ND |
| rs4240702 | COL5A1 | 0.88 (0.68–1.15) | 0.35 | 1.12 (0.87–1.44) | 0.39 | 1.00 (0.79–1.26) | 0.97 |
| rs11605924 | CRY2 | 0.91 (0.70–1.18) | 0.47 | 1.05 (0.82–1.35)* | 0.70 | 0.98 (0.82–1.18)* | 0.83 |
| rs1153188 | DCD | 1.18 (0.94–1.48) | 0.16 | 0.85 (0.68–1.07)* | 0.16 | 1.00 (0.73–1.38)* | 0.99 |
| rs1113132 | EXT2 | 1.02 (0.81–1.28) | 0.86 | 1.01 (0.81–1.26) | 0.93 | 1.02 (0.87–1.19) | 0.86 |
| rs174550 | FADS1 | 1.00 (0.79–1.26) | 1.00 | 1.10 (0.88–1.37) | 0.39 | 1.05 (0.90–1.24) | 0.54 |
| rs11071657 | FAM148B | 0.86 (0.68–1.09) | 0.22 | 1.01 (0.81–1.27) | 0.91 | 0.94 (0.80–1.10) | 0.42 |
| rs17044137 | FLJ39370 | 1.06 (0.84–1.33) | 0.64 | 1.04 (0.83–1.31)* | 0.71 | 1.05 (0.89–1.23)* | 0.56 |
| rs8050136 | FTO | 0.91 (0.70–1.18) | 0.46 | 1.02 (0.81–1.29) | 0.84 | 0.97 (0.82–1.15) | 0.73 |
| rs560887† | G6PC2 | 1.11 (0.74–1.66) | 0.61 | 0.91 (0.61–1.35) | 0.63 | 1.01 (0.77–1.33) | 0.94 |
| rs1799884 | GCK | 0.99 (0.76–1.28) | 0.92 | 1.04 (0.83–1.31)* | 0.74 | 1.02 (0.86–1.21)* | 0.84 |
| rs1260326† | GCKR | 1.36 (1.01–1.82) | 0.043 | 1.10 (0.83–1.47) | 0.51 | 1.22 (0.99–1.50) | 0.061 |
| rs1111875 | HHEX | 1.00 (0.78–1.27) | 0.97 | 1.00 (0.79–1.25)* | 0.97 | 1.00 (0.85–1.18)* | 1.00 |
| rs7957197 | HNF1A (TCF1) | 1.17 (0.92–1.48) | 0.20 | 1.07 (0.85–1.35)* | 0.56 | 1.12 (0.95–1.32)* | 0.19 |
| rs7501939† | HNF1B (TCF2) | 1.49 (1.11–2.00) | 0.008 | 1.40 (1.06–1.84) | 0.016 | 1.44 (1.18–1.76) | 0.0001 |
| rs35767 | IGF1 | 0.87 (0.67–1.12) | 0.27 | 1.08 (0.85–1.37) | 0.53 | 0.98 (0.79–1.20) | 0.81 |
| rs4402960 | IGF2BP2 | 0.90 (0.71–1.13) | 0.36 | 0.90 (0.72–1.12) | 0.34 | 0.90 (0.77–1.06) | 0.20 |
| rs20541† | IL13 | 1.42 (0.89–2.27) | 0.14 | 0.82 (0.48–1.41) | 0.47 | 1.10 (0.64–1.88) | 0.73 |
| rs2943641 | IRS1 | 1.13 (0.89–1.43) | 0.33 | 1.08 (0.86–1.36) | 0.49 | 1.10 (0.94–1.30) | 0.24 |
| rs864745 | JAZF1 | 0.90 (0.71–1.15) | 0.40 | 1.01 (0.79–1.30)* | 0.94 | 0.95 (0.80–1.13)* | 0.58 |
| rs5215 | KCNJ11 | 1.09 (0.86–1.37) | 0.49 | 0.98 (0.78–1.23) | 0.85 | 1.03 (0.88–1.22) | 0.70 |
| rs5219 | KCNJ11 | 1.11 (0.87–1.43) | 0.39 | 0.98 (0.78–1.23) | 0.85 | 1.04 (0.88–1.23) | 0.67 |
| rs2237897 | KCNQ1 | 1.25 (0.88–1.77) | 0.21 | ND | ND | ND | ND |
| rs2074196 | KCNQ1 | 1.57 (1.03–2.40) | 0.036 | ND | ND | ND | ND |
| rs2237892 | KCNQ1 | 1.38 (0.97–1.97) | 0.070 | 1.09 (0.80–1.49) | 0.59 | 1.21 (0.96–1.53) | 0.11 |
| rs2237895 | KCNQ1 | 1.06 (0.82–1.36) | 0.66 | 0.94 (0.75–1.19) | 0.62 | 0.99 (0.84–1.18) | 0.93 |
| rs231362 | KCNQ1OT1 | 1.15 (0.88–1.51) | 0.31 | 1.04 (0.80–1.35) | 0.76 | 1.09 (0.91–1.32) | 0.36 |
| rs1041981 | LTA | 0.86 (0.68–1.09) | 0.21 | 0.95 (0.76–1.18) | 0.64 | 0.91 (0.77–1.07) | 0.24 |
| rs7944584† | MADD | 0.68 (0.46–1.01) | 0.058 | 0.83 (0.56–1.24)* | 0.37 | 0.75 (0.57–0.99)* | 0.044 |
| rs12970134 | MCR4 | 0.89 (0.70–1.12) | 0.31 | 1.11 (0.89–1.39) | 0.34 | 1.00 (0.80–1.24) | 0.98 |
| rs1387153 | MTNR1B | 0.89 (0.71–1.12) | 0.32 | 1.12 (0.90–1.30) | 0.30 | 1.01 (0.81–1.26) | 0.94 |
| rs10923931 | NOTCH2 | 0.98 (0.73–1.31) | 0.88 | 1.10 (0.85–1.44)* | 0.48 | 1.04 (0.86–1.27)* | 0.66 |
| rs6698181 | PKN2 | 1.17 (0.92–1.48) | 0.20 | 0.97 (0.78–1.21) | 0.77 | 1.06 (0.88–1.27) | 0.54 |
| rs1801282 | PPARG | 0.84 (0.65–1.10) | 0.21 | 0.90 (0.70–1.16)* | 0.41 | 0.87 (0.73–1.05)* | 0.14 |
| rs8042680† | PRC1 | 0.91 (0.64–1.29) | 0.60 | 1.05 (0.75–1.47) | 0.77 | 0.98 (0.77–1.25) | 0.87 |
| rs340874 | PROX1 | 1.03 (0.78–1.36) | 0.83 | 0.81 (0.64–1.03)* | 0.08 | 0.90 (0.71–1.14)* | 0.40 |
| rs7593730 | RBMS1 | 0.92 (0.73–1.16) | 0.48 | 1.09 (0.87–1.36)* | 0.44 | 1.00 (0.85–1.19)* | 0.96 |
| rs1531343 | RPSAP52, HMGA2 | 1.11 (0.84–1.46) | 0.45 | 0.88 (0.66–1.17)* | 0.39 | 0.99 (0.79–1.25)* | 0.94 |
| rs11920090 | SLC2A2 | 0.88 (0.67–1.14) | 0.34 | 0.73 (0.56–0.95) | 0.022 | 0.80 (0.66–0.97) | 0.020 |
| rs13266634∂ | SLC30A8 | 1.24 (1.05–1.47) | 0.011 | 1.20 (1.02–1.41) | 0.025 | 1.22 (1.09–1.37) | 0.001 |
| rs7903146 | TCF7L2 | 0.83 (0.66–1.05) | 0.12 | 0.84 (0.67–1.05) | 0.12 | 0.84 (0.71–0.98) | 0.028 |
| rs12255372 | TCF7L2 | 0.87 (0.69–1.09) | 0.23 | 0.83 (0.67–1.04) | 0.10 | 0.85 (0.73–1.00) | 0.043 |
| rs7578597 | THADA | 1.18 (0.88–1.58) | 0.27 | 0.95 (0.73–1.24) | 0.72 | 1.05 (0.85–1.30) | 0.66 |
| rs896854† | TP53INP1 | 0.76 (0.58–1.00) | 0.050 | 0.97 (0.75–1.25)* | 0.82 | 0.86 (0.68–1.10)* | 0.23 |
| rs7961581 | TSPAN8, LGR5 | 1.09 (0.87–1.37) | 0.47 | 0.92 (0.74–1.14) | 0.44 | 1.00 (0.85–1.18) | 0.98 |
| rs9472138 | VEGFA | 1.03 (0.82–1.30) | 0.80 | 1.10 (0.88–1.37) | 0.39 | 1.07 (0.91–1.25) | 0.43 |
| rs10010131 | WFS1 | 1.00 (0.78–1.28) | 0.99 | 1.07 (0.86–1.35) | 0.54 | 1.04 (0.88–1.23) | 0.66 |
Abbreviations: SNP, single nucleotide polymorphism; HR, hazard ratio; CI, confidence interval. ND, not determined.
Estimates were adjusted for age, sex, country of origin and Durie–Salmon stage. P < 0.05 in facebold.
Estimates calculated according to a dominant model of inheritance and adjusted for age, gender, region and Durie–Salmon stage.
Estimates calculated according to a dominant model of inheritance and adjusted for age, gender and clinical trial.
Meta–analyses were performed assuming a random effect model.
Estimates calculated according to a recessive model of inheritance.
Estimates calculated according to an additive model of inheritance.
Estimates based on imputed genotypes.
We also observed significant associations at P < 0.05 for SNPs within CDKN2A-2B, GCKR, KCNQ1 and SLC30A8 genes with OS in the IMMENSE population. Thus, patients carrying the KCNQ1rs2074196T and SLC30A8rs13266634T alleles or the GCKRrs1260326T/T genotype had an increased risk of death whereas subjects bearing the CDKN2A-2Brs564398C/C genotype showed longer OS (Table 2 and Supplementary Tables S2–S4). The association of the SLC30A8rs13266634T allele with OS was confirmed in the Heidelberg population and the meta-analysis showed that the presence of each additional copy of the SLC30A8rs13266634T allele was associated with poor OS (HRMeta-Add = 1.22, 95% CI 1.09-1.37; Table 3). Although the association of the SLC2A2rs11920090 SNP with OS was not significant in the IMMENSE population, we observed a significant association of this variant with MM survival in the Heidelberg population that remained significant in the pooled analysis. Patients harbouring the SLC2A2rs11920090T allele showed a better survival compared with those carrying the A/A genotype (HRMeta-Dom = 0.80, 95% CI 0.66-0.97; Table 3). The meta-analysis also showed a weak association of the TCF7L2rs7903146T allele with better survival that was neither significant in the IMMENSE population nor in the Heidelberg cohort (HRMeta-Dom = 0.84, 95% CI 0.71–0.98). Based on Haploreg data, the missense rs13266634 SNP was predicted to change binding motifs for transcription factors implicated in tumorigenesis (AP1 and PAX5) and mapped on enhancer histone marks in several human embrionic stem cell lines. In addition, this polymorphism affects binding to 5 proteins implicated in cancer development (CCNT2, GATA2, TAL1, KAP1 and CTCF). On the other hand, the rs11920090 and rs7903146 SNPs mapped among enhancer and promoter histone marks in bone marrow- and/or adipose-derived mesenchymal stem cells. In addition, the rs7903146 SNP was predicted to alter the binding site of 7 transcription factors. However, despite the consistency and the potential interest of these findings, none of the associations of the SLC2A2, SLC30A8, and TCF7L2 SNPs with OS remained significant after correction for multiple testing and, therefore, require further confirmation. Given the lack of genetic information in the GWAS conducted in the Heidelberg population for SNPs within MADD and KCNQ1 genes, we imputed genotypes to test whether the preliminary associations observed in the IMMENSE population could be validated. Although there was no imputed data available for the KCNQ1 SNP, the meta-analysis of IMMENSE data with imputed genotypes of MADD variants in the Heidelberg cohort suggested a link between this locus and MM survival (HRMeta-Rec = 0.75, 95% CI 0.57–0.99; Table 3).
Based on the evidences that point toward the existence of gender-associated differences in survival for patients with MM [48], we decided to carry out a gender-stratified analysis. This analysis revealed gender-specific associations for SNPs within or near the ADAMTS9, KCNJ11, PROX1 and SLC30A8 genes with OS. We found that men carrying the KCNJ11rs5215C or SLC30A8rs13266634T alleles had poorer OS compared with those harbouring the wild type genotype whereas an opposite but not significant effect was seen in women (PInteraction= 0.022 and PInteraction = 0.057, respectively; Table 2 and Supplementary Table S2–S4). We also observed that women carrying the PROX1rs340874G allele or the ADAMTS9rs4607103T/T genotype experienced a poorer survival with an opposite but not significant effect in men (PInteraction = 0.016 and PInteraction = 0.024). In order to confirm these gender-specific associations, we performed a meta-analysis with available GWAS data of the Heidelberg population. Although there was a partial overlapping of SNPs between both populations that limited our ability to validate some potentially interesting gender-associated effects on OS, we could confirm the strong association of the SLC30A8rs13266634 SNP with OS in men that could not be detected in women (per-allele HRMen = 1.32, 95% CI 1.13-1.54; Supplementary Table S5). This gender-specific association remained significant at the experiment-wide significance threshold. On the other hand, although it was not statistically significant in the analysis of the IMMENSE population, the pooled analysis also showed that men carrying the BCL11Ars10490072C allele had a poorer OS compared with those carrying the wild type genotype whereas no effect was seen in women (HRMen = 1.37, 95% CI 1.10-1.70). Finally, we observed in the pooled analysis that women bearing the PRC1rs8042680A allele or men carrying the PROX1rs340874G allele or the MADDrs7944584T/T genotype showed a significantly better OS when compared with those patients carrying the corresponding wild type allele or genotype (HRWomen = 0.62, 95% CI 0.39–0.98; HRMen = 0.74, 95% CI 0.59–0.94 and HRMen = 0.59, 95% CI 0.39–0.87; Supplementary Table S5). The regulatory characteristics of the rs10490072, rs8042680 and rs7944584 SNPs were changes in transcription binding motifs for transcription factors involved in tumorigenesis and T- and B-cell malignancies. The rs10490072 changed sites for HNF4B, Pou2f2 and Pou5f1 whereas the rs8042680 altered sites for GR, PAX5 and TAL1. The rs7944584 was found to modify regulatory motifs for AP1, AP4, IRF and KAP1. Finally, the rs340874 mapped among promoter and enhancer histone marks in primary naïve and memory forms of helper T cells (CD4+) and regulatory T cells from peripheral blood.
DISCUSSION
Previous population-based studies have demonstrated the impact of GWAS-identified variants for T2D on cancer susceptibility [47, 49–52] and patient survival [53]. However, despite these important research advances, there is still a noticeable lack of information regarding the role of T2D-related variants in modulating patient survival especially in hematological malignancies. In this scenario, we decided to investigate for the first time to our knowledge the relationship between 58 genetic variants associated with T2D identified by GWAS and OS of MM patients.
The analysis of the IMMENSE consortium data revealed a significant association of the intronic HNF1Brs7501939 SNP with poor OS. We successfully replicated this association in a large and independent population recruited by the University Clinic of Heidelberg. However, although a positive correlation between this variant and eQTL data on PBMCs has been reported [54], we failed to find correlation between the risk allele and HNF1B mRNA expression levels on plasma cells from a large cohort of MM patients. This suggested that the effect of this variant on overall survival is not mediated by changes in transcriptional activity of the gene. Nonetheless, given that HFN1B contains multiple independent SNPs or haplotypes that have been associated with HNF1B mRNA expression [55, 56] and methylation [57] levels but also with the risk of developing several types of cancer [55–57], it seems to be reasonable to consider the possibility that other SNPs within this locus and showing a stronger association with OS could explain better the link between the HNF1B and clinical outcome. However, when we analysed imputed common SNPs from the GWAS conducted in the Heidelberg cohort, we could not find any stronger association signals with OS in the region, which suggested that the HNF1Brs7501939 SNP or perhaps a rare SNP in LD with it might be responsible of the observed effect. Future fine-mapping studies encompassing common but also rare variants within or near the HNF1B locus are needed to elucidate whether a rare variant or haplotype might account for the observed effect.
HNF1B contains 9 exons and expands over 58 kb on chromosome 17p21 [58, 59]. It encodes for a transcription factor that has been associated with multiple clinical features including early-onset of T2D [56]. In line with this, it has been also suggested that HNF1B may induce impaired glucose tolerance and attenuated insulin sensitivity in a miRNA-dependent manner [60], which might lead to an enhanced insulin secretion and the activation of the IGF1 pathway, an important factor mediating myeloma cell growth, proliferation and cell maturation [27, 59, 61]. Alternatively, it has been postulated that HNF1B is able to influence cancer cell survival by promoting the activation of NFkB pathway or through the inhibition of mitochondria-associated apoptotic signals [62]. In support of the tumorogenic effect of HFN1B, it has also been reported that it may act as an oncogene [63] and that the HNF1B gene is amplified in 23% of all cancers and in about 5% of all haematological malignancies (http://broadinstitute.org/tumorscape). On the contrary, it has also been reported that HFN1B may act as tumour suppressor gene [56] and that its expression may largely vary depending on the target tissue. Whereas HNF1B has been found to be overexpressed in ovarian clear cell carcinomas [58] and prostate [64] or endometrial [65] cancers and its silencing induces apoptosis of cancer cells [58], it has been found to be down-regulated in serous epithelial ovarian cancer [57] and colorectal, gastric and pancreatic cancer cell lines [66]. In addition, it has been reported that the down-regulation of HFN1B gene is associated with progression in hepatocarcinoma [67] and poor prognosis in renal [68] and prostate [69] cancers. Considering all the above but also the fact that the association of the HNF1Brs7501939 SNP with OS was driven by a non-diabetogenic (T) allele that does not affect HNF1B mRNA expression, we hypothesize that the effect of this variant to contribute to tumour progression in MM might be mediated by a non-insulin-dependent mechanism. There was a reasonable amount of regulatory data for the HNF1Brs7501939 SNP that supported evidence of the active role of the HNF1B locus. However, whether elevated HNF1B levels lead to tumour transformation and disease progression is not yet understood and functional studies to examine whether HNF1B variants influence cancer prognosis are lacking.
Another interesting finding of this study was the association of the BCL11A, MADD, PRC1, PROX1, SCL30A8, SLC2A2 and TCF7L2 SNPs with OS. We found an overall association of the SLC2A2rs11920090T and TCF7L2rs7903146T alleles with better OS whereas the association of the SLC30A8rs13266634T, BCL11Ars10490072C, PRC1rs8042680A and PROX1rs340874G alleles or the MADDrs7944584T/T genotype with OS was restricted to male or female genders. Despite the potential interest of the associations observed for these SNPs with OS, only the association of the SLC30A8rs13266634 SNP with poor OS in men reached significance at experiment-wide significance threshold. This result suggested a key role of the SLC30A8 locus in the modulation of overall survival. However, given the consistency of the overall or gender-specific associations observed for BCL11A, MADD, PRC1, PROX1, SLC2A2 and TCF7L2 SNPs with OS across the populations tested and considering that gender-specific genetic alterations might influence MM survival [48], we suggest that these variants might also exert a modest effect to modulate patient survival.
SCL30A8 gene encodes a zinc transporter involved in the control of insulin processing and secretion [70]. Although no previous studies have reported a link between this locus and MM, there are evidences that suggest that zinc transporters might contribute to cancinogenesis [71, 72] through a gender-dependent mechanism [73]. The association of the coding SLC30A8rs13266634 SNP with OS was due to a non-diabetogenic allele suggesting that, rather than modulating glucose homeostasis and insulin secretion [74], the effect attributed to the SLC30A8 locus on MM survival might be driven by a direct effect of Zinc in biological processes such as DNA and RNA stabilization [75], binding of protooncogenes to DNA [75–77] and the activation of IGF1 [26, 27, 61] or telomerase [78]. The SLC30A8rs13266634C allele has been consistently associated with decreased rates of Zinc transport activitiy and reduced intragranular Zinc levels [79]. However, eQTL data on plasma cells from MM patients did not reveal correlation between this variant and SLC30A8 mRNA levels suggesting that, rather than regulating gene expression, the T allele affect transporter activity in an allele-dose-dependent manner causing increased Zinc concentration and thereby promoting unlimited proliferation of MM cells, disease progression and poor survival. In addition, regulatory data suggest that the SLC30A8 locus might play a role in survival through the modulation of specific transcription factors implicated in tumour promotion and dissemination.
As for the HNF1B and SLC30A8 SNPs, the association of the BCL11Ars10490072 and MADDrs7944584 SNPs with OS was determined by non-diabetogenic alleles. BCL11A functions as a myeloid and B-cell proto-oncogen and has been associated with the development of B-cell malignancies [80, 81] whereas MADD encodes for a MAP-kinase activating cell domain involved in the control of physiological cell death through TNF- and caspase-dependent apoptosis [82]. In contrast to these associations, the association of the TCF7L2rs7903146,SLC2A2rs11920090 PRC1rs8042680, and PROX1rs340874 SNPs with OS was driven by diabetogenic alleles, which suggested that the effect of these variants on OS might be explained by their regulatory effect on insulin secretion and, consequently, on cell proliferation and tumour cell growth. Whereas SLC2A2 encodes a highly efficient glucose transport that is expressed in pancreatic cells and regulates insulin secretion by modulating entry of glucose into the pancreatic cell [83], TCF7L2, PRC1 and PROX1 are proteins that have been involved in β-cell survival and function [84] and in glucose and nonesterified fatty acids or branched-chain amino acids metabolism in liver [85, 86]. Despite these interesting results and the regulatory data observed for all these SNPs, the lack of information regarding T2D status among MM patients did not allow us to ensure that the observed effect of the TCF7L2, SLC2A2, PRC1, and PROX1 SNPs on OS could not be due to a different distribution of diabetic patients when grouping by genotype or gender.
This study had both strengths and limitations. Strengths include the use of relative large discovery and replication populations that allowed us to validate the most interesting associations. Limitations include lack of information regarding the classical genetic prognostic factors (chromosomal abnormalities, etc.), T2D status and a relatively small statistical power to detect modest associations with OS, especially when gender-stratified analysis were performed. Another limitation was the partial overlapping of genetic information between studies and the use of imputed genotypes that did not allow to perform a reliable validation of the association observed for genetic variants within ADAMTS9, BCL11A, KCNQ1, MADD and PROX1 genes with overall survival.
In conclusion, this study reports the first evidence of an association between the HNF1Brs7501939 SNP and OS for MM and suggests that the HNF1B locus might, likely through a non-insulin-dependent mechanism, play an important role in modulating MM prognosis. Likewise, this study shows a strong association of the SLC30A8rs13266634 SNP with poor OS in men that might, at least in part, account for gender differences in OS. Additional studies using larger and well-characterized populations are needed to further replicate these findings but also those involving the BCL11A, MADD, PRC1, PROX1, SLC2A2 and TCF7L2 loci on OS.
MATERIALS AND METHODS
Patients, clinical data collection and survival endpoint definition
A total of 1420 Caucasian MM patients were ascertained through the IMMEnSE consortium. Full details of this consortium have been published elsewhere [87]. In brief, inclusion criteria were newly diagnosed MM with Salmon & Durie stage I, II and III, age 18–90 years inclusive and Caucasian origin. DNA was purified from blood specimens using the QIAamp DNA Blood Mini Kit (Qiagen) and clinicophathological characteristics including age, gender, country of origin and disease stage (Durie-Salmon) were retrospectively gathered from medical records in each participant institution (Table 1). Diagnosis of patients with symptomatic MM was carried out by hematologists according to the International Myeloma Working Group (IMWG) criteria [88, 89]. All patients within the IMMENSE consortium for whom survival information was available were included in the study (936 MM cases, 454 women and 482 men) (Table 1). All participants gave their written informed consent to participate in the study.
SNP selection and genotyping
Fifty-eight variants were selected based on the GWAS for T2D [84, 90–126] and were genotyped in the IMMEnSE consortium population (Table 4). We considered only SNPs that were replicated in large and independent populations or which came up in several GWAS or their meta-analyses. Additional criteria were potential functionality and linkage disequilibrium (LD) between the reported SNPs. The genotyping of the selected polymorphisms was carried out at GENYO (Centre for Genomics and Oncological Research: Pfizer/University of Granada/Andalusian Regional Government, Granada, Spain) using KASPar® assays (LGC Genomics, Hoddesdon, UK) according to manufacturer's instructions. For internal quality control, 5% of samples were randomly selected and included as duplicates. Concordance between the original and the duplicate samples for the 58 SNPs was ≥ 99.0%. Call rates for all SNPs were ≥ 90.0% with the exception of the WFS1rs734312 SNP that was excluded from further analyses.
Table 4. Selected type-2 diabetes-related SNPs.
| Gene name | dbSNP rs# | Nucleotide substitution | Reference allele IMMENSE | GWAS-identified risk allele for T2D | Location/Aa substitution | References |
|---|---|---|---|---|---|---|
| ADAM30 | rs2641348 | T/C | T | C | L359P | [103, 124] |
| ADAMTS9 | rs4607103 | T/C1 | C | C | Near gene | [84, 104, 124] |
| ADCY5 | rs11708067 | T/C2 | T | T | Intronic | [96, 111] |
| ADRA2A | rs10885122 | G/T | G | G | Near ADRA2A | [96] |
| ARAPI, CENTD2 | rs1552224 | G/T | T | T | Near gene | [105, 120] |
| BCL11A | rs10490072 | C/T | T | T | Near gene | [124] |
| CDC123, CAMK1D | rs12779790 | A/G3 | A | G | Near gene | [84, 104, 124] |
| CDKAL1 | rs7754840 | C/G | G | C | Intronic | [95, 97, 113] |
| CDKN2A-2B | rs564398 | T/C4 | T | T | Near gene | [84, 95, 104, 113, 116, 122, 124] |
| CDKN2A-2B | rs10811661 | T/C | T | T | Near gene | |
| CDKN2A-2B | rs2383208 | A/G | A | A | Near gene | |
| COL5A1 | rs4240702 | C/T | C | n/s | Intronic | [91] |
| CRY2 | rs11605924 | A/C | C | A | Intronic | [96] |
| DCD | rs1153188 | A/T | A | A | Near gene | [124] |
| EXT2 | rs1113132 | C/G | C | C | Intronic | [97, 114] |
| FADS1 | rs174550 | C/T | T | T | Intronic | [96] |
| FAM148B | rs11071657 | A/G | A | A | Near gene | [93, 96] |
| FLJ39370 | rs17044137 | A/T | T | A | Near gene | [95] |
| FTO | rs8050136 | A/C | C | A | Intronic | [104, 125, 126] |
| G6PC2 | rs560887 | G/A | G | G | Intronic | [91, 92, 94, 96, 107] |
| GCK | rs1799884 | G/A | G | A | Near gene | [91, 92, 94, 96, 107] |
| GCKR | rs1260326 | C/T5 | C | T | L445P | [91, 96, 111] |
| HHEX | rs1111875 | G/A | G | G | Near gene | [95, 97, 104, 113, 114, 125, 126] |
| HMGA2 | rs1531343 | C/G | G | C | Near gene | [105, 120] |
| HNF1A (TCF1) | rs7957197 | A/T | T | T | Intronic | [105, 120] |
| HNF1B (TCF2) | rs7501939 | C/T6 | C | C | Intronic | [101, 110] |
| IGF1 | rs35767 | C/T | C | C | Near gene | [96, 106] |
| IGF2BP2 | rs4402960 | G/T | G | T | Intronic | [84, 95, 97, 104, 113, 125, 126] |
| IL13 | rs20541 | C/T7 | C | T | R144Q | [95] |
| IRS1 | rs2943641 | C/T | C | C | Near gene | [109, 117, 120] |
| JAZF1 | rs864745 | A/G | A | A | Intronic | [84, 124] |
| KCNJ11 | rs5215 | T/C8 | T | C | V337I | [95, 98, 104, 113, 121, 125, 126] |
| KCNJ11 | rs5219 | C/T | C | T | K23E | |
| KCNQ1 | rs2237897 | C/T | T | C | Intronic | [118, 119, 122, 123] |
| KCNQ1 | rs2074196 | G/T9 | G | G | Intronic | |
| KCNQ1 | rs2237892 | C/T | C | C | Intronic | |
| KCNQ1 | rs2237895 | A/C | A | C | Intronic | |
| KCNQ1OT1 | rs231362 | G/A | G | G | Intronic | [105, 118, 120] |
| LTA | rs1041981 | A/C | A | A | T60N | [102] |
| MADD | rs7944584 | A/T10 | A | A | Intronic | [96] |
| MCR4 | rs12970134 | A/G | G | A | Near gene | [93] |
| MTNR1B | rs1387153 | C/T | C | T | Near gene | [91, 107, 120] |
| NOTCH2 | rs10923931 | G/T | G | T | Intronic | [104, 124] |
| PKN2 | rs6698181 | C/T | C | T | Intergenic | [95] |
| PPARG | rs1801282 | C/G11 | C | C | P12A | [90, 95, 104, 113, 121, 124–126] |
| PRC1 | rs8042680 | A/C | C | A | Intronic | [105, 120] |
| PROX1 | rs340874 | A/G12 | A | G | Promoter | [96] |
| RBMS1 | rs7593730 | C/T | C | T | Intronic | [108] |
| SLC2A2 | rs11920090 | A/T | A | T | Intronic | [96] |
| SLC30A8 | rs13266634 | C/T13 | C | C | R325W | [84, 90, 95–97, 104, 113, 114, 125, 126] |
| TCF7L2 | rs7903146 | C/T14 | C | T | Intronic | [95–97, 99, 104, 11–115, 125, 126 |
| TCF7L2 | rs12255372 | G/T15 | G | T | Intronic | |
| THADA | rs7578597 | T/C | T | T | T1187A | [124] |
| TP53INP1 | rs896854 | A/G | G | G | Intronic | [105, 120] |
| TSPAN8 | rs7961581 | C/T | T | C | Near gene | [100] |
| VEGFA | rs9472138 | C/T | C | T | Near gene | [124] |
| WFS1 | rs734312 | A/G | A | n/s | H611R | [110] |
| WFS1 | rs10010131 | A/G | G | G | Intronic | [110] |
n/s, not specified; Aa, Aminoacid; GWAS, genome-wide association studies; OS, overall survival.
References are listed in Supplementary Material. Effect allele in bold and underlined.
T/T genotype was associated with poor OS in women with an opposite but not significant effect in men.
C allele was associated with better OS in women with no effect in men.
G allele was associated with better OS in women with no effect in men.
C/C genotype was associated with better OS. No gender-specific effect was observed.
T/T genotype was associated with poor OS. No gender-specific effect was observed.
T allele was associated with poor OS. No gender-specific effect was observed.
T/T genotype was associated with poor OS in women with an opposite but not significant effect in men.
C allele was associated with poor OS in men with an opposite but not significant effect in women.
T allele was associated with poor OS. No gender-specific effect was observed.
T/T genotype was associated with better OS in men with no significant effect in women.
G allele was associated with better OS in men with an opposite but not significant effect in women.
G allele was associated with poor OS in women with an opposite but not significant effect in men.
The presence of each additional copy of the T allele was associated with poor OS in men with no effect in women (additive effect).
T allele was associated with better OS in men with no effect in women.
T allele was associated with better OS in men with an opposite but not significant effect in women.
Replication
For replication purposes, seven hundred MM patients (296 women and 404 men) were provided by the University Clinic of Heidelberg (Germany). This cohort consists of 98 GMMG-HD3 trial patients, 291 GMMG-HD4 trial patients and 311 patients transplanted in Heidelberg but not enrolled in clinical trials (Table 1). Ethical approval for these patients and written informed consent of trial patients was also obtained. Clinical and survival data were prospectively collected for trial patients on case report forms and retrospectively gathered from medical records for none-trial patients. Genetic information of 53 SNPs (36 genotyped SNPs and 17 imputed SNPs) was extracted from the GWAS conducted in the Heidelberg cohort. After imputation, no information was available for 5 SNPs.
In silico functional analysis
Haploreg (http://www.broadinstitute.org/mammals/haploreg/haploreg.php) and ENCODE annotation data (https://genome.ucsc.edu/ENCODE/) were used to predict the functional role of potentially interesting SNPs.
eQTL analysis
We also assessed whether selected SNPs correlated with mRNA expression levels in a public eQTL browser for peripheral blood mononuclear cells (http://genenetwork.nl/bloodeqtlbrowser/) [54]. Expression quantitative trait loci (eQTL) data on malignant plasma cells of 658 patients from the University Clinic of Heidelberg (Germany) were also available for this study. Detailed information on sample collection and clinico-pathological characteristics of MM patients as well as technical details of gene expression analysis have been published elsewhere [127].
Statistical analysis
We used chi-square tests to assess Hardy–Weinberg Equilibrium (HWE) for each SNP among IMMEnSE patients. The primary outcome was OS and the endpoint was defined as death from any cause. Survival time was calculated as the time from MM diagnosis (discovery population) or the first stem cell transplantation (replication population) until the occurrence of the study endpoint, censoring at the date of death or the last observed follow-up time. Association with OS defined as hazard ratio (HR) was calculated for each SNP using Cox regression multivariate analysis adjusted for age, gender, country of origin and Durie-Salmon stage (IMMENSE cohort) or for age, gender and clinical trial (Heidelberg cohort). Association estimates were calculated according to dominant, recessive and log-additive models of inheritance with the major allele as reference for regression analyses (Table 4). We also performed gene-gender interaction analyses to determine whether the association between SNPs and MM OS was of similar magnitude in men and women. Survival function was displayed using the Kaplan-Meier method [128] and survival differences across genotypic groups were analysed using the log-rank test.
In order to account for multiple comparisons, we used the Meff/MeffLi method [129], which calculates the effective number of independent genetic markers analysed (N = 54) on the basis of the spectral decomposition (SpD) of matrices of pairwise LD between SNPs (http://neurogenetics.qimrberghofer.edu.au/SNPSpDlite). In addition, we also considered the number of genetic inheritance models tested (dominant, recessive and log-additive). This resulted in a study-wide significance threshold of 0.00031 ([0.05/54]/3) to keep type I error rate at 5%.
Finally, in order to confirm significant associations, a meta-analysis combining genetic data obtained in the IMMENSE population with those extracted from the GWAS conducted in the Heidelberg cohort was also performed following dominant, recessive and additive models of inheritance. The I2 statistic was used to assess heterogeneity between both studies and the pooled HR was computed using the random-effect model (assuming that between-study variation might depend on chance or random variation and an individual study effect). Random-effects models are more conservative than fixed-effects models and give rise to wider confidence intervals (CI), which ensures the reliability of the results even though the data come from studies with a relatively different design. All statistics were calculated using SPSS (v.20) and STATA (v.12) for MAC.
SUPPLEMENTARY TABLES
Footnotes
CONFLICTS OF INTEREST
All authors have nothing to disclose.
GRANT SUPPORT
This work was supported by grants from the FIBAO foundation (Granada, Spain), from the CRIS foundation against cancer, from the Cancer Network of Excellence (RD12/10 Red de Cáncer), from the Instituto de Salud Carlos III (Madrid, Spain; PI12/02688) and from the Dietmar Hopp Foundation and the German Ministry of Education and Science (BMBF: CLIOMMICS [01ZX1309]).
REFERENCES
- 1.Alexander DD, Mink PJ, Adami HO, Cole P, Mandel JS, Oken MM, Trichopoulos D. Multiple myeloma: a review of the epidemiologic literature. Int J Cancer. 2007;120:40–61. doi: 10.1002/ijc.22718. [DOI] [PubMed] [Google Scholar]
- 2.Rios-Tamayo R, Sanchez MJ, Puerta JM, Sainz J, Chang DY, Rodriguez T, Lopez P, de Pablos JM, Navarro P, de Veas JL, Romero A, Garrido P, Moratalla L, et al. Trends in survival of multiple myeloma: a thirty-year population-based study in a single institution. Cancer Epidemiol. 2015;39:693–699. doi: 10.1016/j.canep.2015.08.002. [DOI] [PubMed] [Google Scholar]
- 3.Kumar SK, Rajkumar SV, Dispenzieri A, Lacy MQ, Hayman SR, Buadi FK, Zeldenrust SR, Dingli D, Russell SJ, Lust JA, Greipp PR, Kyle RA, Gertz MA. Improved survival in multiple myeloma and the impact of novel therapies. Blood. 2008;111:2516–2520. doi: 10.1182/blood-2007-10-116129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Avet-Loiseau H, Attal M, Campion L, Caillot D, Hulin C, Marit G, Stoppa AM, Voillat L, Wetterwald M, Pegourie B, Voog E, Tiab M, Banos A, et al. Long-term analysis of the IFM 99 trials for myeloma: cytogenetic abnormalities [t(4;14), del(17p), 1q gains] play a major role in defining long-term survival. J Clin Oncol. 2012;30:1949–1952. doi: 10.1200/JCO.2011.36.5726. [DOI] [PubMed] [Google Scholar]
- 5.Ludwig H, Bolejack V, Crowley J, Blade J, Miguel JS, Kyle RA, Rajkumar SV, Shimizu K, Turesson I, Westin J, Sonneveld P, Cavo M, Boccadoro M, et al. Survival and years of life lost in different age cohorts of patients with multiple myeloma. J Clin Oncol. 2010;28:1599–1605. doi: 10.1200/JCO.2009.25.2114. [DOI] [PubMed] [Google Scholar]
- 6.Durie BG, Salmon SE. A clinical staging system for multiple myeloma. Correlation of measured myeloma cell mass with presenting clinical features, response to treatment, and survival. Cancer. 1975;36:842–854. doi: 10.1002/1097-0142(197509)36:3<842::aid-cncr2820360303>3.0.co;2-u. [DOI] [PubMed] [Google Scholar]
- 7.Greipp PR, San Miguel J, Durie BG, Crowley JJ, Barlogie B, Blade J, Boccadoro M, Child JA, Avet-Loiseau H, Kyle RA, Lahuerta JJ, Ludwig H, Morgan G, et al. International staging system for multiple myeloma. J Clin Oncol. 2005;23:3412–3420. doi: 10.1200/JCO.2005.04.242. [DOI] [PubMed] [Google Scholar]
- 8.Oken MM, Creech RH, Tormey DC, Horton J, Davis TE, McFadden ET, Carbone PP. Toxicity and response criteria of the Eastern Cooperative Oncology Group. Am J Clin Oncol. 1982;5:649–655. [PubMed] [Google Scholar]
- 9.Augustson BM, Begum G, Dunn JA, Barth NJ, Davies F, Morgan G, Behrens J, Smith A, Child JA, Drayson MT. Early mortality after diagnosis of multiple myeloma: analysis of patients entered onto the United kingdom Medical Research Council trials between 1980 and 2002—Medical Research Council Adult Leukaemia Working Party. J Clin Oncol. 2005;23:9219–9226. doi: 10.1200/JCO.2005.03.2086. [DOI] [PubMed] [Google Scholar]
- 10.Knudsen LM, Hjorth M, Hippe E. Renal failure in multiple myeloma: reversibility and impact on the prognosis. Nordic Myeloma Study Group. Eur J Haematol. 2000;65:175–181. doi: 10.1034/j.1600-0609.2000.90221.x. [DOI] [PubMed] [Google Scholar]
- 11.Greipp PR, Witzig TE, Gonchoroff NJ, Habermann TM, Katzmann JA, O’Fallon WM, Kyle RA. Immunofluorescence labeling indices in myeloma and related monoclonal gammopathies. Mayo Clin Proc. 1987;62:969–977. doi: 10.1016/s0025-6196(12)65066-6. [DOI] [PubMed] [Google Scholar]
- 12.Witzig TE, Dhodapkar MV, Kyle RA, Greipp PR. Quantitation of circulating peripheral blood plasma cells and their relationship to disease activity in patients with multiple myeloma. Cancer. 1993;72:108–113. doi: 10.1002/1097-0142(19930701)72:1<108::aid-cncr2820720121>3.0.co;2-t. [DOI] [PubMed] [Google Scholar]
- 13.Chim CS, Sim J, Tam S, Tse E, Lie AK, Kwong YL. LDH is an adverse prognostic factor independent of ISS in transplant-eligible myeloma patients receiving bortezomib-based induction regimens. Eur J Haematol. 2015;94:330–335. doi: 10.1111/ejh.12434. [DOI] [PubMed] [Google Scholar]
- 14.Avet-Loiseau H. Role of genetics in prognostication in myeloma. Best Pract Res Clin Haematol. 2007;20:625–635. doi: 10.1016/j.beha.2007.08.005. [DOI] [PubMed] [Google Scholar]
- 15.Fonseca R, Bergsagel PL, Drach J, Shaughnessy J, Gutierrez N, Stewart AK, Morgan G, Van Ness B, Chesi M, Minvielle S, Neri A, Barlogie B, Kuehl WM, et al. International Myeloma Working Group molecular classification of multiple myeloma: spotlight review. Leukemia. 2009;23:2210–2221. doi: 10.1038/leu.2009.174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Munshi NC, Anderson KC, Bergsagel PL, Shaughnessy J, Palumbo A, Durie B, Fonseca R, Stewart AK, Harousseau JL, Dimopoulos M, Jagannath S, Hajek R, Sezer O, et al. Consensus recommendations for risk stratification in multiple myeloma: report of the International Myeloma Workshop Consensus Panel 2. Blood. 2011;117:4696–4700. doi: 10.1182/blood-2010-10-300970. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Boyd KD, Ross FM, Chiecchio L, Dagrada GP, Konn ZJ, Tapper WJ, Walker BA, Wardell CP, Gregory WM, Szubert AJ, Bell SE, Child JA, Jackson GH, et al. A novel prognostic model in myeloma based on co-segregating adverse FISH lesions and the ISS: analysis of patients treated in the MRC Myeloma IX trial. Leukemia. 2012;26:349–355. doi: 10.1038/leu.2011.204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Fonseca R, Blood E, Rue M, Harrington D, Oken MM, Kyle RA, Dewald GW, Van Ness B, Van Wier SA, Henderson KJ, Bailey RJ, Greipp PR. Clinical and biologic implications of recurrent genomic aberrations in myeloma. Blood. 2003;101:4569–4575. doi: 10.1182/blood-2002-10-3017. [DOI] [PubMed] [Google Scholar]
- 19.Castillo JJ, Mull N, Reagan JL, Nemr S, Mitri J. Increased incidence of non-Hodgkin lymphoma, leukemia, and myeloma in patients with diabetes mellitus type 2: a meta-analysis of observational studies. Blood. 2012;119:4845–4850. doi: 10.1182/blood-2011-06-362830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Badros A, Goloubeva O, Dalal JS, Can I, Thompson J, Rapoport AP, Heyman M, Akpek G, Fenton RG. Neurotoxicity of bortezomib therapy in multiple myeloma: a single-center experience and review of the literature. Cancer. 2007;110:1042–1049. doi: 10.1002/cncr.22921. [DOI] [PubMed] [Google Scholar]
- 21.Libourel EJ, Sonneveld P, van der Holt B, de Maat MP, Leebeek FW. High incidence of arterial thrombosis in young patients treated for multiple myeloma: results of a prospective cohort study. Blood. 2010;116:22–26. doi: 10.1182/blood-2009-12-257519. [DOI] [PubMed] [Google Scholar]
- 22.Palumbo A, Rajkumar SV, Dimopoulos MA, Richardson PG, San Miguel J, Barlogie B, Harousseau J, Zonder JA, Cavo M, Zangari M, Attal M, Belch A, Knop S, et al. Prevention of thalidomide- and lenalidomide-associated thrombosis in myeloma. Leukemia. 2008;22:414–423. doi: 10.1038/sj.leu.2405062. [DOI] [PubMed] [Google Scholar]
- 23.Snowden JA, Ahmedzai SH, Ashcroft J, D'sa S, Littlewood T, Low E, Lucraft H, Maclean R, Feyler S, Pratt G, Bird JM, Haemato-oncology Task Force of British Committee for Standards in H, Forum UKM Guidelines for supportive care in multiple myeloma 2011. Br J Haematol. 2011;154:76–103. doi: 10.1111/j.1365-2141.2011.08574.x. [DOI] [PubMed] [Google Scholar]
- 24.Chiu BC, Gapstur SM, Greenland P, Wang R, Dyer A. Body mass index, abnormal glucose metabolism, and mortality from hematopoietic cancer. Cancer Epidemiol Biomarkers Prev. 2006;15:2348–2354. doi: 10.1158/1055-9965.EPI-06-0007. [DOI] [PubMed] [Google Scholar]
- 25.Chou YS, Yang CF, Chen HS, Yang SH, Yu YB, Hong YC, Liu CY, Gau JP, Liu JH, Chen PM, Chiou TJ, Tzeng CH, Hsiao LT. Pre-existing diabetes mellitus in patients with multiple myeloma. Eur J Haematol. 2012;89:320–327. doi: 10.1111/j.1600-0609.2012.01828.x. [DOI] [PubMed] [Google Scholar]
- 26.Grimberg A. Mechanisms by which IGF-I may promote cancer. Cancer Biol Ther. 2003;2:630–635. [PMC free article] [PubMed] [Google Scholar]
- 27.Sprynski AC, Hose D, Kassambara A, Vincent L, Jourdan M, Rossi JF, Goldschmidt H, Klein B. Insulin is a potent myeloma cell growth factor through insulin/IGF-1 hybrid receptor activation. Leukemia. 2010;24:1940–1950. doi: 10.1038/leu.2010.192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kim WY, Sharpless NE. The regulation of INK4/ARF in cancer and aging. Cell. 2006;127:265–275. doi: 10.1016/j.cell.2006.10.003. [DOI] [PubMed] [Google Scholar]
- 29.Koh J, Enders GH, Dynlacht BD, Harlow E. Tumour-derived p16 alleles encoding proteins defective in cell-cycle inhibition. Nature. 1995;375:506–510. doi: 10.1038/375506a0. [DOI] [PubMed] [Google Scholar]
- 30.Than BL, Goos JA, Sarver AL, O'sullivan MG, Rod A, Starr TK, Fijneman RJ, Meijer GA, Zhao L, Zhang Y, Largaespada DA, Scott PM, Cormier RT. The role of KCNQ1 in mouse and human gastrointestinal cancers. Oncogene. 2013:3861–3868. doi: 10.1038/onc.2013.350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Chim CS, Kwong YL, Liang R. Gene hypermethylation in multiple myeloma: lessons from a cancer pathway approach. Clin Lymphoma Myeloma. 2008;8:331–339. doi: 10.3816/CLM.2008.n.048. [DOI] [PubMed] [Google Scholar]
- 32.Ng MH, Chung YF, Lo KW, Wickham NW, Lee JC, Huang DP. Frequent hypermethylation of p16 and p15 genes in multiple myeloma. Blood. 1997;89:2500–2506. [PubMed] [Google Scholar]
- 33.Mateos MV, Garcia-Sanz R, Lopez-Perez R, Moro MJ, Ocio E, Hernandez J, Megido M, Caballero MD, Fernandez-Calvo J, Barez A, Almeida J, Orfao A, Gonzalez M, San Miguel JF. Methylation is an inactivating mechanism of the p16 gene in multiple myeloma associated with high plasma cell proliferation and short survival. Br J Haematol. 2002;118:1034–1040. doi: 10.1046/j.1365-2141.2002.03749.x. [DOI] [PubMed] [Google Scholar]
- 34.Than BL, Goos JA, Sarver AL, O'sullivan MG, Rod A, Starr TK, Fijneman RJ, Meijer GA, Zhao L, Zhang Y, Largaespada DA, Scott PM, Cormier RT. The role of KCNQ1 in mouse and human gastrointestinal cancers. Oncogene. 2014;33:3861–3868. doi: 10.1038/onc.2013.350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mirandola L, Apicella L, Colombo M, Yu Y, Berta DG, Platonova N, Lazzari E, Lancellotti M, Bulfamante G, Cobos E, Chiriva-Internati M, Chiaramonte R. Anti-Notch treatment prevents multiple myeloma cells localization to the bone marrow via the chemokine system CXCR4/SDF-1. Leukemia. 2013;27:1558–1566. doi: 10.1038/leu.2013.27. [DOI] [PubMed] [Google Scholar]
- 36.Colombo M, Galletti S, Garavelli S, Platonova N, Paoli A, Basile A, Taiana E, Neri A, Chiaramonte R. Notch signaling deregulation in multiple myeloma: A rational molecular target. Oncotarget. 2015;6:26826–26840. doi: 10.18632/oncotarget.5025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.van Stralen E, van de Wetering M, Agnelli L, Neri A, Clevers HC, Bast BJ. Identification of primary MAFB target genes in multiple myeloma. Exp Hematol. 2009;37:78–86. doi: 10.1016/j.exphem.2008.08.006. [DOI] [PubMed] [Google Scholar]
- 38.Chubb D, Weinhold N, Broderick P, Chen B, Johnson DC, Forsti A, Vijayakrishnan J, Migliorini G, Dobbins SE, Holroyd A, Hose D, Walker BA, Davies FE, et al. Common variation at 3q26.2, 6p21.33, 17p11.2 and 22q13.1 influences multiple myeloma risk. Nat Genet. 2013;45:1221–1225. doi: 10.1038/ng.2733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Weinhold N, Johnson DC, Chubb D, Chen B, Forsti A, Hosking FJ, Broderick P, Ma YP, Dobbins SE, Hose D, Walker BA, Davies FE, Kaiser MF, et al. The CCND1 c. 870G>A polymorphism is a risk factor for t(11;14)(q13;q32) multiple myeloma. Nat Genet. 2013;45:522–525. doi: 10.1038/ng.2583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Martino A, Campa D, Jamroziak K, Reis RM, Sainz J, Buda G, Garcia-Sanz R, Lesueur F, Marques H, Moreno V, Jurado M, Rios R, Szemraj-Rogucka Z, et al. Impact of polymorphic variation at 7p15.3, 3p22.1 and 2p23.3 loci on risk of multiple myeloma. Br J Haematol. 2012;158:805–809. doi: 10.1111/j.1365-2141.2012.09244.x. [DOI] [PubMed] [Google Scholar]
- 41.Campa D, Martino A, Sainz J, Buda G, Jamroziak K, Weinhold N, Vieira Reis RM, Garcia-Sanz R, Jurado M, Rios R, Szemraj-Rogucka Z, Marques H, Lesueur F, et al. Comprehensive investigation of genetic variation in the 8q24 region and multiple myeloma risk in the IMMEnSE consortium. Br J Haematol. 2012;157:331–338. doi: 10.1111/j.1365-2141.2012.09047.x. [DOI] [PubMed] [Google Scholar]
- 42.Martino A, Campa D, Buda G, Sainz J, Garcia-Sanz R, Jamroziak K, Reis RM, Weinhold N, Jurado M, Rios R, Szemraj-Rogucka Z, Marques H, Szemraj J, et al. Polymorphisms in xenobiotic transporters ABCB1, ABCG2, ABCC2, ABCC1, ABCC3 and multiple myeloma risk: a case-control study in the context of the International Multiple Myeloma rESEarch (IMMEnSE) consortium. Leukemia. 2012;26:1419–1422. doi: 10.1038/leu.2011.352. [DOI] [PubMed] [Google Scholar]
- 43.Campa D, Martino A, Varkonyi J, Lesueur F, Jamroziak K, Landi S, Jurczyszyn A, Marques H, Andersen V, Jurado M, Brenner H, Petrini M, Vogel U, et al. Risk of multiple myeloma is associated with polymorphisms within telomerase genes and telomere length. Int J Cancer. 2015;136:E351–358. doi: 10.1002/ijc.29101. [DOI] [PubMed] [Google Scholar]
- 44.Ziv E, Dean E, Hu D, Martino A, Serie D, Curtin K, Campa D, Aftab B, Bracci P, Buda G, Zhao Y, Caswell-Jin J, Diasio R, et al. Genome-wide association study identifies variants at 16p13 associated with survival in multiple myeloma patients. Nat Commun. 2015;6:7539. doi: 10.1038/ncomms8539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Johnson DC, Weinhold N, Mitchell JS, Chen B, Kaiser M, Begum DB, Hillengass J, Bertsch U, Gregory WA, Cairns D, Jackson GH, Forsti A, Nickel J, et al. Genome-wide association study identifies variation at 6q25. 1 associated with survival in multiple myeloma. Nat Commun. 2016;7:10290. doi: 10.1038/ncomms10290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Pang H, Hauser M, Minvielle S. Pathway-based identification of SNPs predictive of survival. Eur J Hum Genet. 2011;19:704–709. doi: 10.1038/ejhg.2011.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Rios R, Lupianez CB, Campa D, Martino A, Martinez-Lopez J, Martinez-Bueno M, Varkonyi J, Garcia-Sanz R, Jamroziak K, Dumontet C, Cayuela AJ, Wetek M, Landi S, et al. Type 2 diabetes-related variants influence the risk of developing multiple myeloma: results from the IMMEnSE consortium. Endocr Relat Cancer. 2015;22:545–559. doi: 10.1530/ERC-15-0029. [DOI] [PubMed] [Google Scholar]
- 48.Boyd KD, Ross FM, Chiecchio L, Dagrada G, Konn ZJ, Tapper WJ, Walker BA, Wardell CP, Gregory WM, Szubert AJ, Davies FE, Morgan GJ. Gender disparities in the tumor genetics and clinical outcome of multiple myeloma. Cancer Epidemiol Biomarkers Prev. 2011;20:1703–1707. doi: 10.1158/1055-9965.EPI-11-0157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ma RC, So WY, Tam CH, Luk AO, Ho JS, Wang Y, Lam VK, Lee HM, Kong AP, Tong PC, Xu G, Chow CC, Ng MC, et al. Genetic variants for type 2 diabetes and new-onset cancer in Chinese with type 2 diabetes. Diabetes Res Clin Pract. 2014;103:328–337. doi: 10.1016/j.diabres.2013.12.016. [DOI] [PubMed] [Google Scholar]
- 50.Cheng I, Caberto CP, Lum-Jones A, Seifried A, Wilkens LR, Schumacher FR, Monroe KR, Lim U, Tiirikainen M, Kolonel LN, Henderson BE, Stram DO, Haiman CA, Le Marchand L. Type 2 diabetes risk variants and colorectal cancer risk: the Multiethnic Cohort and PAGE studies. Gut. 2011;60:1703–1711. doi: 10.1136/gut.2011.237727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Machiela MJ, Lindstrom S, Allen NE, Haiman CA, Albanes D, Barricarte A, Berndt SI, Bueno-de-Mesquita HB, Chanock S, Gaziano JM, Gapstur SM, Giovannucci E, Henderson BE, et al. Association of type 2 diabetes susceptibility variants with advanced prostate cancer risk in the Breast and Prostate Cancer Cohort Consortium. Am J Epidemiol. 2012;176:1121–1129. doi: 10.1093/aje/kws191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Pierce BL, Austin MA, Ahsan H. Association study of type 2 diabetes genetic susceptibility variants and risk of pancreatic cancer: an analysis of PanScan-I data. Cancer Causes Control. 2011;22:877–883. doi: 10.1007/s10552-011-9760-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Bao PP, Zhao ZG, Gao YT, Zheng Y, Zhang B, Cai H, Zheng W, Shu XO, Lu W. Association of type 2 diabetes genetic variants with breast cancer survival among Chinese women. PLoS One. 2015;10:e0117419. doi: 10.1371/journal.pone.0117419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Westra HJ, Peters MJ, Esko T, Yaghootkar H, Schurmann C, Kettunen J, Christiansen MW, Fairfax BP, Schramm K, Powell JE, Zhernakova A, Zhernakova DV, Veldink JH, et al. Systematic identification of trans eQTLs as putative drivers of known disease associations. Nat Genet. 2013;45:1238–1243. doi: 10.1038/ng.2756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Harries LW, Perry JR, McCullagh P, Crundwell M. Alterations in LMTK2, MSMB and HNF1B gene expression are associated with the development of prostate cancer. BMC cancer. 2010;10:315. doi: 10.1186/1471-2407-10-315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Painter JN, O’Mara TA, Batra J, Cheng T, Lose FA, Dennis J, Michailidou K, Tyrer JP, Ahmed S, Ferguson K, Healey CS, Kaufmann S, Hillman KM, et al. Fine-mapping of the HNF1B multicancer locus identifies candidate variants that mediate endometrial cancer risk. Hum Mol Genet. 2015;24:1478–1492. doi: 10.1093/hmg/ddu552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Shen H, Fridley BL, Song H, Lawrenson K, Cunningham JM, Ramus SJ, Cicek MS, Tyrer J, Stram D, Larson MC, Kobel M, Consortium P, Ziogas A, et al. Epigenetic analysis leads to identification of HNF1B as a subtype-specific susceptibility gene for ovarian cancer. Nat Commun. 2013;4:1628. doi: 10.1038/ncomms2629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Tsuchiya A, Sakamoto M, Yasuda J, Chuma M, Ohta T, Ohki M, Yasugi T, Taketani Y, Hirohashi S. Expression profiling in ovarian clear cell carcinoma: identification of hepatocyte nuclear factor-1 beta as a molecular marker and a possible molecular target for therapy of ovarian clear cell carcinoma. Am J Pathol. 2003;163:2503–2512. doi: 10.1016/s0002-9440(10)63605-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Ferlin M, Noraz N, Hertogh C, Brochier J, Taylor N, Klein B. Insulin-like growth factor induces the survival and proliferation of myeloma cells through an interleukin-6-independent transduction pathway. Br J Haematol. 2000;111:626–634. doi: 10.1046/j.1365-2141.2000.02364.x. [DOI] [PubMed] [Google Scholar]
- 60.Roth HP, Kirchgessner M. Influence of alimentary zinc deficiency on the concentration of growth hormone (GH), insulin-like growth factor I (IGF-I) and insulin in the serum of force-fed rats. Horm Metab Res. 1994;26:404–408. doi: 10.1055/s-2007-1001718. [DOI] [PubMed] [Google Scholar]
- 61.Bommert K, Bargou RC, Stuhmer T. Signalling and survival pathways in multiple myeloma. Eur J Cancer. 2006;42:1574–1580. doi: 10.1016/j.ejca.2005.12.026. [DOI] [PubMed] [Google Scholar]
- 62.Suzuki E, Kajita S, Takahashi H, Matsumoto T, Tsuruta T, Saegusa M. Transcriptional upregulation of HNF-1beta by NF-kappaB in ovarian clear cell carcinoma modulates susceptibility to apoptosis through alteration in bcl-2 expression. Lab Invest. 2015;95:962–972. doi: 10.1038/labinvest.2015.73. [DOI] [PubMed] [Google Scholar]
- 63.Shao DD, Tsherniak A, Gopal S, Weir BA, Tamayo P, Stransky N, Schumacher SE, Zack TI, Beroukhim R, Garraway LA, Margolin AA, Root DE, Hahn WC, Mesirov JP. ATARiS: computational quantification of gene suppression phenotypes from multisample RNAi screens. Genome Res. 2013;23:665–678. doi: 10.1101/gr.143586.112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Sun J, Zheng SL, Wiklund F, Isaacs SD, Purcell LD, Gao Z, Hsu FC, Kim ST, Liu W, Zhu Y, Stattin P, Adami HO, Wiley KE, et al. Evidence for two independent prostate cancer risk-associated loci in the HNF1B gene at 17q12. Nat Genet. 2008;40:1153–1155. doi: 10.1038/ng.214. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Spurdle AB, Thompson DJ, Ahmed S, Ferguson K, Healey CS, O’Mara T, Walker LC, Montgomery SB, Dermitzakis ET, Australian National Endometrial Cancer Study G. Fahey P, Montgomery GW, Webb PM, et al. Genome-wide association study identifies a common variant associated with risk of endometrial cancer. Nat Genet. 2011;43:451–454. doi: 10.1038/ng.812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Terasawa K, Toyota M, Sagae S, Ogi K, Suzuki H, Sonoda T, Akino K, Maruyama R, Nishikawa N, Imai K, Shinomura Y, Saito T, Tokino T. Epigenetic inactivation of TCF2 in ovarian cancer and various cancer cell lines. Br J Cancer. 2006;94:914–921. doi: 10.1038/sj.bjc.6602984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Lazarevich NL, Cheremnova OA, Varga EV, Ovchinnikov DA, Kudrjavtseva EI, Morozova OV, Fleishman DI, Engelhardt NV, Duncan SA. Progression of HCC in mice is associated with a downregulation in the expression of hepatocyte nuclear factors. Hepatology. 2004;39:1038–1047. doi: 10.1002/hep.20155. [DOI] [PubMed] [Google Scholar]
- 68.Buchner A, Castro M, Hennig A, Popp T, Assmann G, Stief CG, Zimmermann W. Downregulation of HNF-1B in renal cell carcinoma is associated with tumor progression and poor prognosis. Urology. 2010;76:507–e506–511. doi: 10.1016/j.urology.2010.03.042. [DOI] [PubMed] [Google Scholar]
- 69.Glinsky GV, Glinskii AB, Stephenson AJ, Hoffman RM, Gerald WL. Gene expression profiling predicts clinical outcome of prostate cancer. J Clin Invest. 2004;113:913–923. doi: 10.1172/JCI20032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Chimienti F, Devergnas S, Pattou F, Schuit F, Garcia-Cuenca R, Vandewalle B, Kerr-Conte J, Van Lommel L, Grunwald D, Favier A, Seve M. In vivo expression and functional characterization of the zinc transporter ZnT8 in glucose-induced insulin secretion. J Cell Sci. 2006;119:4199–4206. doi: 10.1242/jcs.03164. [DOI] [PubMed] [Google Scholar]
- 71.Zowczak M, Iskra M, Torlinski L, Cofta S. Analysis of serum copper and zinc concentrations in cancer patients. Biol Trace Elem Res. 2001;82:1–8. doi: 10.1385/BTER:82:1-3:001. [DOI] [PubMed] [Google Scholar]
- 72.Leitzmann MF, Stampfer MJ, Wu K, Colditz GA, Willett WC, Giovannucci EL. Zinc supplement use and risk of prostate cancer. J Natl Cancer Inst. 2003;95:1004–1007. doi: 10.1093/jnci/95.13.1004. [DOI] [PubMed] [Google Scholar]
- 73.Pound LD, Sarkar SA, Ustione A, Dadi PK, Shadoan MK, Lee CE, Walters JA, Shiota M, McGuinness OP, Jacobson DA, Piston DW, Hutton JC, Powell DR, O’Brien RM. The physiological effects of deleting the mouse SLC30A8 gene encoding zinc transporter-8 are influenced by gender and genetic background. PLoS One. 2012;7:e40972. doi: 10.1371/journal.pone.0040972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Nicolson TJ, Bellomo EA, Wijesekara N, Loder MK, Baldwin JM, Gyulkhandanyan AV, Koshkin V, Tarasov AI, Carzaniga R, Kronenberger K, Taneja TK, da Silva Xavier G, Libert S, et al. Insulin storage and glucose homeostasis in mice null for the granule zinc transporter ZnT8 and studies of the type 2 diabetes-associated variants. Diabetes. 2009;58:2070–2083. doi: 10.2337/db09-0551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Franz MC, Anderle P, Burzle M, Suzuki Y, Freeman MR, Hediger MA, Kovacs G. Zinc transporters in prostate cancer. Mol Aspects Med. 2013;34:735–741. doi: 10.1016/j.mam.2012.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Cho Y, Gorina S, Jeffrey PD, Pavletich NP. Crystal structure of a p53 tumor suppressor-DNA complex: understanding tumorigenic mutations. Science. 1994;265:346–355. doi: 10.1126/science.8023157. [DOI] [PubMed] [Google Scholar]
- 77.Verhaegh GW, Parat MO, Richard MJ, Hainaut P. Modulation of p53 protein conformation and DNA-binding activity by intracellular chelation of zinc. Mol Carcinog. 1998;21:205–214. doi: 10.1002/(sici)1098-2744(199803)21:3<205::aid-mc8>3.0.co;2-k. [DOI] [PubMed] [Google Scholar]
- 78.Nemoto K, Kondo Y, Himeno S, Suzuki Y, Hara S, Akimoto M, Imura N. Modulation of telomerase activity by zinc in human prostatic and renal cancer cells. Biochem Pharmacol. 2000;59:401–405. doi: 10.1016/s0006-2952(99)00334-2. [DOI] [PubMed] [Google Scholar]
- 79.Rutter GA, Chimienti F. SLC30A8 mutations in type 2 diabetes. Diabetologia. 2015;58:31–36. doi: 10.1007/s00125-014-3405-7. [DOI] [PubMed] [Google Scholar]
- 80.Satterwhite E, Sonoki T, Willis TG, Harder L, Nowak R, Arriola EL, Liu H, Price HP, Gesk S, Steinemann D, Schlegelberger B, Oscier DG, Siebert R, et al. The BCL11 gene family: involvement of BCL11A in lymphoid malignancies. Blood. 2001;98:3413–3420. doi: 10.1182/blood.v98.12.3413. [DOI] [PubMed] [Google Scholar]
- 81.Weniger MA, Pulford K, Gesk S, Ehrlich S, Banham AH, Lyne L, Martin-Subero JI, Siebert R, Dyer MJ, Moller P, Barth TF. Gains of the proto-oncogene BCL11A and nuclear accumulation of BCL11A(XL) protein are frequent in primary mediastinal B-cell lymphoma. Leukemia. 2006;20:1880–1882. doi: 10.1038/sj.leu.2404324. [DOI] [PubMed] [Google Scholar]
- 82.Bi W, Wei Y, Wu J, Sun G, Guo Y, Zhang Q, Dong L. MADD promotes the survival of human lung adenocarcinoma cells by inhibiting apoptosis. Oncol Rep. 2013;29:1533–1539. doi: 10.3892/or.2013.2258. [DOI] [PubMed] [Google Scholar]
- 83.Mueckler M. Facilitative glucose transporters. Eur J Biochem. 1994;219:713–725. doi: 10.1111/j.1432-1033.1994.tb18550.x. [DOI] [PubMed] [Google Scholar]
- 84.Shu L, Sauter NS, Schulthess FT, Matveyenko AV, Oberholzer J, Maedler K. Transcription factor 7-like 2 regulates beta-cell survival and function in human pancreatic islets. Diabetes. 2008;57:645–653. doi: 10.2337/db07-0847. [DOI] [PubMed] [Google Scholar]
- 85.Kretowski A, Adamska E, Maliszewska K, Wawrusiewicz-Kurylonek N, Citko A, Goscik J, Bauer W, Wilk J, Golonko A, Waszczeniuk M, Lipinska D, Hryniewicka J, Niemira M, et al. The rs340874 PROX1 type 2 diabetes mellitus risk variant is associated with visceral fat accumulation and alterations in postprandial glucose and lipid metabolism. Genes Nutr. 2015;10:454. doi: 10.1007/s12263-015-0454-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Stancakova A, Civelek M, Saleem NK, Soininen P, Kangas AJ, Cederberg H, Paananen J, Pihlajamaki J, Bonnycastle LL, Morken MA, Boehnke M, Pajukanta P, Lusis AJ, et al. Hyperglycemia and a common variant of GCKR are associated with the levels of eight amino acids in 9,369 Finnish men. Diabetes. 2012;61:1895–1902. doi: 10.2337/db11-1378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Martino A, Sainz J, Buda G, Jamroziak K, Reis RM, Garcia-Sanz R, Jurado M, Rios R, Szemraj-Rogucka Z, Marques H, Lesueur F, Moreno V, Orciuolo E, et al. Genetics and molecular epidemiology of multiple myeloma: the rationale for the IMMEnSE consortium (review) Int J Oncol. 2012;40:625–638. doi: 10.3892/ijo.2011.1284. [DOI] [PubMed] [Google Scholar]
- 88.Rajkumar SV, Dimopoulos MA, Palumbo A, Blade J, Merlini G, Mateos MV, Kumar S, Hillengass J, Kastritis E, Richardson P, Landgren O, Paiva B, Dispenzieri A, et al. International Myeloma Working Group updated criteria for the diagnosis of multiple myeloma. Lancet Oncol. 2014;15:e538–548. doi: 10.1016/S1470-2045(14)70442-5. [DOI] [PubMed] [Google Scholar]
- 89.International Myeloma Working G Criteria for the classification of monoclonal gammopathies, multiple myeloma and related disorders: a report of the International Myeloma Working Group. Br J Haematol. 2003;121:749–757. [PubMed] [Google Scholar]
- 90.Altshuler D, Hirschhorn JN, Klannemark M, Lindgren CM, Vohl MC, Nemesh J, Lane CR, Schaffner SF, Bolk S, Brewer C, Tuomi T, Gaudet D, Hudson TJ, et al. The common PPARgamma Pro12Ala polymorphism is associated with decreased risk of type 2 diabetes. Nat Genet. 2000;26:76–80. doi: 10.1038/79216. [DOI] [PubMed] [Google Scholar]
- 91.Bouatia-Naji N, Bonnefond A, Cavalcanti-Proenca C, Sparso T, Holmkvist J, Marchand M, Delplanque J, Lobbens S, Rocheleau G, Durand E, De Graeve F, Chevre JC, Borch-Johnsen K, et al. A variant near MTNR1B is associated with increased fasting plasma glucose levels and type 2 diabetes risk. Nat Genet. 2009;41:89–94. doi: 10.1038/ng.277. [DOI] [PubMed] [Google Scholar]
- 92.Bouatia-Naji N, Rocheleau G, Van Lommel L, Lemaire K, Schuit F, Cavalcanti-Proenca C, Marchand M, Hartikainen AL, Sovio U, De Graeve F, Rung J, Vaxillaire M, Tichet J, et al. A polymorphism within the G6PC2 gene is associated with fasting plasma glucose levels. Science. 2008;320:1085–1088. doi: 10.1126/science.1156849. [DOI] [PubMed] [Google Scholar]
- 93.Chambers JC, Elliott P, Zabaneh D, Zhang W, Li Y, Froguel P, Balding D, Scott J, Kooner JS. Common genetic variation near MC4R is associated with waist circumference and insulin resistance. Nat Genet. 2008;40:716–718. doi: 10.1038/ng.156. [DOI] [PubMed] [Google Scholar]
- 94.Chen WM, Erdos MR, Jackson AU, Saxena R, Sanna S, Silver KD, Timpson NJ, Hansen T, Orru M, Grazia Piras M, Bonnycastle LL, Willer CJ, Lyssenko V, et al. Variations in the G6PC2/ABCB11 genomic region are associated with fasting glucose levels. J Clin Invest. 2008;118:2620–2628. doi: 10.1172/JCI34566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Diabetes Genetics Initiative of Broad Institute of H. Mit LU, Novartis Institutes of BioMedical R. Saxena R, Voight BF, Lyssenko V, Burtt NP, de Bakker PI, Chen H, Roix JJ, Kathiresan S, Hirschhorn JN, Daly MJ, et al. Genome-wide association analysis identifies loci for type 2 diabetes and triglyceride levels. Science. 2007;316:1331–1336. doi: 10.1126/science.1142358. [DOI] [PubMed] [Google Scholar]
- 96.Dupuis J, Langenberg C, Prokopenko I, Saxena R, Soranzo N, Jackson AU, Wheeler E, Glazer NL, Bouatia-Naji N, Gloyn AL, Lindgren CM, Magi R, Morris AP, et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat Genet. 2010;42:105–116. doi: 10.1038/ng.520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Florez JC, Manning AK, Dupuis J, McAteer J, Irenze K, Gianniny L, Mirel DB, Fox CS, Cupples LA, Meigs JB. A 100K genome-wide association scan for diabetes and related traits in the Framingham Heart Study: replication and integration with other genome-wide datasets. Diabetes. 2007;56:3063–3074. doi: 10.2337/db07-0451. [DOI] [PubMed] [Google Scholar]
- 98.Gloyn AL, Weedon MN, Owen KR, Turner MJ, Knight BA, Hitman G, Walker M, Levy JC, Sampson M, Halford S, McCarthy MI, Hattersley AT, Frayling TM. Large-scale association studies of variants in genes encoding the pancreatic beta-cell KATP channel subunits Kir6. 2 (KCNJ11) and SUR1 (ABCC8) confirm that the KCNJ11 E23K variant is associated with type 2 diabetes. Diabetes. 2003;52:568–572. doi: 10.2337/diabetes.52.2.568. [DOI] [PubMed] [Google Scholar]
- 99.Grant SF, Thorleifsson G, Reynisdottir I, Benediktsson R, Manolescu A, Sainz J, Helgason A, Stefansson H, Emilsson V, Helgadottir A, Styrkarsdottir U, Magnusson KP, Walters GB, et al. Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes. Nat Genet. 2006;38:320–323. doi: 10.1038/ng1732. [DOI] [PubMed] [Google Scholar]
- 100.Grarup N, Andersen G, Krarup NT, Albrechtsen A, Schmitz O, Jorgensen T, Borch-Johnsen K, Hansen T, Pedersen O. Association testing of novel type 2 diabetes risk alleles in the JAZF1, CDC123/CAMK1D, TSPAN8, THADA, ADAMTS9, and NOTCH2 loci with insulin release, insulin sensitivity, and obesity in a population-based sample of 4,516 glucose-tolerant middle-aged Danes. Diabetes. 2008;57:2534–2540. doi: 10.2337/db08-0436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Gudmundsson J, Sulem P, Steinthorsdottir V, Bergthorsson JT, Thorleifsson G, Manolescu A, Rafnar T, Gudbjartsson D, Agnarsson BA, Baker A, Sigurdsson A, Benediktsdottir KR, Jakobsdottir M, et al. Two variants on chromosome 17 confer prostate cancer risk, and the one in TCF2 protects against type 2 diabetes. Nat Genet. 2007;39:977–983. doi: 10.1038/ng2062. [DOI] [PubMed] [Google Scholar]
- 102.Hamid YH, Urhammer SA, Glumer C, Borch-Johnsen K, Jorgensen T, Hansen T, Pedersen O. The common T60N polymorphism of the lymphotoxin-alpha gene is associated with type 2 diabetes and other phenotypes of the metabolic syndrome. Diabetologia. 2005;48:445–451. doi: 10.1007/s00125-004-1659-1. [DOI] [PubMed] [Google Scholar]
- 103.Lyssenko V, Nagorny CL, Erdos MR, Wierup N, Jonsson A, Spegel P, Bugliani M, Saxena R, Fex M, Pulizzi N, Isomaa B, Tuomi T, Nilsson P, et al. Common variant in MTNR1B associated with increased risk of type 2 diabetes and impaired early insulin secretion. Nat Genet. 2009;41:82–88. doi: 10.1038/ng.288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Mohlke KL, Boehnke M, Abecasis GR. Metabolic and cardiovascular traits: an abundance of recently identified common genetic variants. Hum Mol Genet. 2008;17:R102–108. doi: 10.1093/hmg/ddn275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Nielsen T, Sparso T, Grarup N, Jorgensen T, Pisinger C, Witte DR, Diabetes Genetics R, Meta-analysis C, Hansen T, Pedersen O. Type 2 diabetes risk allele near CENTD2 is associated with decreased glucose-stimulated insulin release. Diabetologia. 2011;54:1052–1056. doi: 10.1007/s00125-011-2054-3. [DOI] [PubMed] [Google Scholar]
- 106.Pechlivanis S, Wagner K, Chang-Claude J, Hoffmeister M, Brenner H, Forsti A. Polymorphisms in the insulin like growth factor 1 and IGF binding protein 3 genes and risk of colorectal cancer. Cancer Detect Prev. 2007;31:408–416. doi: 10.1016/j.cdp.2007.10.001. [DOI] [PubMed] [Google Scholar]
- 107.Prokopenko I, Langenberg C, Florez JC, Saxena R, Soranzo N, Thorleifsson G, Loos RJ, Manning AK, Jackson AU, Aulchenko Y, Potter SC, Erdos MR, Sanna S, et al. Variants in MTNR1B influence fasting glucose levels. Nat Genet. 2009;41:77–81. doi: 10.1038/ng.290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Qi L, Cornelis MC, Kraft P, Stanya KJ, Linda Kao WH, Pankow JS, Dupuis J, Florez JC, Fox CS, Pare G, Sun Q, Girman CJ, Laurie CC, et al. Genetic variants at 2q24 are associated with susceptibility to type 2 diabetes. Hum Mol Genet. 2010;19:2706–2715. doi: 10.1093/hmg/ddq156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Rung J, Cauchi S, Albrechtsen A, Shen L, Rocheleau G, Cavalcanti-Proenca C, Bacot F, Balkau B, Belisle A, Borch-Johnsen K, Charpentier G, Dina C, Durand E, et al. Genetic variant near IRS1 is associated with type 2 diabetes, insulin resistance and hyperinsulinemia. Nat Genet. 2009;41:1110–1115. doi: 10.1038/ng.443. [DOI] [PubMed] [Google Scholar]
- 110.Sandhu MS, Weedon MN, Fawcett KA, Wasson J, Debenham SL, Daly A, Lango H, Frayling TM, Neumann RJ, Sherva R, Blech I, Pharoah PD, Palmer CN, et al. Common variants in WFS1 confer risk of type 2 diabetes. Nat Genet. 2007;39:951–953. doi: 10.1038/ng2067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Saxena R, Hivert MF, Langenberg C, Tanaka T, Pankow JS, Vollenweider P, Lyssenko V, Bouatia-Naji N, Dupuis J, Jackson AU, Kao WH, Li M, Glazer NL, et al. Genetic variation in GIPR influences the glucose and insulin responses to an oral glucose challenge. Nat Genet. 2010;42:142–148. doi: 10.1038/ng.521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Scott LJ, Bonnycastle LL, Willer CJ, Sprau AG, Jackson AU, Narisu N, Duren WL, Chines PS, Stringham HM, Erdos MR, Valle TT, Tuomilehto J, Bergman RN, et al. Association of transcription factor 7-like 2 (TCF7L2) variants with type 2 diabetes in a Finnish sample. Diabetes. 2006;55:2649–2653. doi: 10.2337/db06-0341. [DOI] [PubMed] [Google Scholar]
- 113.Scott LJ, Mohlke KL, Bonnycastle LL, Willer CJ, Li Y, Duren WL, Erdos MR, Stringham HM, Chines PS, Jackson AU, Prokunina-Olsson L, Ding CJ, Swift AJ, et al. A genome-wide association study of type 2 diabetes in Finns detects multiple susceptibility variants. Science. 2007;316:1341–1345. doi: 10.1126/science.1142382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Sladek R, Rocheleau G, Rung J, Dina C, Shen L, Serre D, Boutin P, Vincent D, Belisle A, Hadjadj S, Balkau B, Heude B, Charpentier G, et al. A genome-wide association study identifies novel risk loci for type 2 diabetes. Nature. 2007;445:881–885. doi: 10.1038/nature05616. [DOI] [PubMed] [Google Scholar]
- 115.Steinthorsdottir V, Thorleifsson G, Reynisdottir I, Benediktsson R, Jonsdottir T, Walters GB, Styrkarsdottir U, Gretarsdottir S, Emilsson V, Ghosh S, Baker A, Snorradottir S, Bjarnason H, et al. A variant in CDKAL1 influences insulin response and risk of type 2 diabetes. Nat Genet. 2007;39:770–775. doi: 10.1038/ng2043. [DOI] [PubMed] [Google Scholar]
- 116.Takeuchi F, Serizawa M, Yamamoto K, Fujisawa T, Nakashima E, Ohnaka K, Ikegami H, Sugiyama T, Katsuya T, Miyagishi M, Nakashima N, Nawata H, Nakamura J, et al. Confirmation of multiple risk Loci and genetic impacts by a genome-wide association study of type 2 diabetes in the Japanese population. Diabetes. 2009;58:1690–1699. doi: 10.2337/db08-1494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Tang Y, Han X, Sun X, Lv C, Zhang X, Guo W, Ren Q, Luo Y, Zhang X, Zhou X, Ji L. Association study of a common variant near IRS1 with type 2 diabetes mellitus in Chinese Han population. Endocrine. 2013;43:84–91. doi: 10.1007/s12020-012-9693-0. [DOI] [PubMed] [Google Scholar]
- 118.Tsai FJ, Yang CF, Chen CC, Chuang LM, Lu CH, Chang CT, Wang TY, Chen RH, Shiu CF, Liu YM, Chang CC, Chen P, Chen CH, et al. A genome-wide association study identifies susceptibility variants for type 2 diabetes in Han Chinese. PLoS Genet. 2010;6:e1000847. doi: 10.1371/journal.pgen.1000847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Unoki H, Takahashi A, Kawaguchi T, Hara K, Horikoshi M, Andersen G, Ng DP, Holmkvist J, Borch-Johnsen K, Jorgensen T, Sandbaek A, Lauritzen T, Hansen T, et al. SNPs in KCNQ1 are associated with susceptibility to type 2 diabetes in East Asian and European populations. Nat Genet. 2008;40:1098–1102. doi: 10.1038/ng.208. [DOI] [PubMed] [Google Scholar]
- 120.Voight BF, Scott LJ, Steinthorsdottir V, Morris AP, Dina C, Welch RP, Zeggini E, Huth C, Aulchenko YS, Thorleifsson G, McCulloch LJ, Ferreira T, Grallert H, et al. Twelve type 2 diabetes susceptibility loci identified through large-scale association analysis. Nat Genet. 2010;42:579–589. doi: 10.1038/ng.609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Willer CJ, Bonnycastle LL, Conneely KN, Duren WL, Jackson AU, Scott LJ, Narisu N, Chines PS, Skol A, Stringham HM, Petrie J, Erdos MR, Swift AJ, et al. Screening of 134 single nucleotide polymorphisms (SNPs) previously associated with type 2 diabetes replicates association with 12 SNPs in nine genes. Diabetes. 2007;56:256–264. doi: 10.2337/db06-0461. [DOI] [PubMed] [Google Scholar]
- 122.Yamauchi T, Hara K, Maeda S, Yasuda K, Takahashi A, Horikoshi M, Nakamura M, Fujita H, Grarup N, Cauchi S, Ng DP, Ma RC, Tsunoda T, et al. A genome-wide association study in the Japanese population identifies susceptibility loci for type 2 diabetes at UBE2E2 and C2CD4A-C2CD4B. Nat Genet. 2010;42:864–868. doi: 10.1038/ng.660. [DOI] [PubMed] [Google Scholar]
- 123.Yasuda K, Miyake K, Horikawa Y, Hara K, Osawa H, Furuta H, Hirota Y, Mori H, Jonsson A, Sato Y, Yamagata K, Hinokio Y, Wang HY, et al. Variants in KCNQ1 are associated with susceptibility to type 2 diabetes mellitus. Nat Genet. 2008;40:1092–1097. doi: 10.1038/ng.207. [DOI] [PubMed] [Google Scholar]
- 124.Zeggini E, Scott LJ, Saxena R, Voight BF, Marchini JL, Hu T, de Bakker PI, Abecasis GR, Almgren P, Andersen G, Ardlie K, Bostrom KB, Bergman RN, et al. Meta-analysis of genome-wide association data and large-scale replication identifies additional susceptibility loci for type 2 diabetes. Nat Genet. 2008;40:638–645. doi: 10.1038/ng.120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Zeggini E, Weedon MN, Lindgren CM, Frayling TM, Elliott KS, Lango H, Timpson NJ, Perry JR, Rayner NW, Freathy RM, Barrett JC, Shields B, Morris AP, et al. Replication of genome-wide association signals in UK samples reveals risk loci for type 2 diabetes. Science. 2007;316:1336–1341. doi: 10.1126/science.1142364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Wellcome Trust Case Control C Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature. 2007;447:661–678. doi: 10.1038/nature05911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Weinhold N, Meissner T, Johnson DC, Seckinger A, Moreaux J, Forsti A, Chen B, Nickel J, Chubb D, Rawstron AC, Doughty C, Dahir NB, Begum DB, et al. The 7p15. 3 (rs4487645) association for multiple myeloma shows strong allele-specific regulation of the MYC-interacting gene CDCA7L in malignant plasma cells. Haematologica. 2015;100:e110–113. doi: 10.3324/haematol.2014.118786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Kaplan ELaMP. Nonparametric Estimation from Incomplete Observations: Taylor & Francis, Ltd. on behalf of the American Statistical Association) 1958 [Google Scholar]
- 129.Nyholt DR. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am J Hum Genet. 2004;74:765–769. doi: 10.1086/383251. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.