

Abstract
Data on the distribution of free-living amoebae is still lacking especially in Southeast Asian region. The aquatic environment revealed a high occurrence of free-living amoebae (FLA) due to its suitable condition and availability of food source, which subsequently causes infection to humans. A total of 94 water samples consisted of both treated and untreated from Laos (31), Myanmar (42), and Singapore (21) were investigated for the presence of pathogenic FLA. Each water sample was filtered and cultured onto non-nutrient agar seeded with live suspension of Escherichia coli and incubated at room temperature. Morphological identification was conducted for both trophozoites and cysts via microscopic stains (Giemsa and immunofluorescence). The presence of Naegleria-like structures was the most frequently encountered in both treated and untreated water samples, followed by Acanthamoeba-like and Vermamoeba-like features. To identify the pathogenic isolates, species-specific primer sets were applied for molecular identification of Acanthamoeba, Naegleria, and Vermamoeba. The pathogenic species of Acanthamoeba lenticulata and A. triangularis were detected from untreated water samples, while Vermamoeba vermiformis was found in both treated and untreated water samples. Our results suggested that poor water quality as well as inadequate maintenance and treatment might be the cause of this alarming problem since chlorine disinfection is ineffective in eradicating these amoebas in treated water samples. Regular monitoring and examination of water qualities are necessary in order to control the growth, hence, further preventing the widespread of FLA infections among the public.
Introduction
Free-living amoebae (FLA) belonging to the genera Acanthamoeba, Balamuthia, Naegleria, Sappinia, and Vermamoeba (= Hartmannella) are potentially pathogenic to humans [1,2]. They are also known as amphizoic amoeba due to the ability to exist within a host or in the environment as ‘free-living’. They are able to survive and proliferate in the environment independently and can be found in various natural and man-made aquatic environments, both fresh and marine, including lakes, ponds, swimming pools, and even treated water supplies [3–5]. The amoeba cysts are highly resistant to harsh conditions due to the process of encystment [6,7].
Many species under the genus of Acanthamoeba are reported to be the causative agent of keratitis in healthy individuals, often among contact lens wearers. In opportunistic infections, Acanthamoeba species can cause pneumonitis, fatal granulomatous encephalitis and skin infections [8]. To date, 20 different genotypes (T1-T20) of Acanthamoeba have been described [9]. Meanwhile, Naegleria is the only FLA that has the advantage of exhibiting a flagellate stage to ease its movement by swimming in water. Moreover, more than 40 species of Naegleria have been identified, only N. fowleri is found to be the causative agent of primary amoebic meningoencephalitis (PAM), a lethal brain infection [10]. Approximately, 440 of PAM cases have been reported worldwide until the year 2008 [11], with exposure of healthy individuals to warm (water temperatures of 25 to 44°C), untreated or poorly disinfected water systems. In addition, Vermamoeba (= Hartmannella) vermiformis seems to be the potential causative agent of human keratitis [12] and it can serves as a host to pathogenic bacteria, Legionella pneumophila [13,14].
In Southeast Asia, the occurrence of FLA were reported in Malaysia [15,16], Thailand [17], Vietnam [18] and the Philippines [19]. Although the reported cases are extremely rare, the diagnosis of FLA must never be overlooked as it is able to cause serious and fatal diseases. Keratitis infection caused by Acanthamoeba species was reported among contact lens wearers in Malaysia [20] and Singapore [21]. Meanwhile, N. fowleri, was reported to be the etiological agent for 12 cases of PAM in Thailand [22,23]. In addition, the first case of Balamuthia mandrillaris causing meningoencephalitis was reported from a Thai male patient after falling into a swamp [24].
The knowledge of pathogenic FLA emergence has gained much interest throughout the world due to its possible health implications. However, inadequate studies of FLA attributed to the lack of prevalence information across the Southeast Asian region. Therefore, we undertook an investigation to detect the occurrence and distribution of the FLA, together with the identification based on culture, staining, and molecular assay of each isolate. This information should be useful for early detection of potential infestation of pathogenic FLA in various water sources and further evaluating the risks of human contact with FLA. In fact, humans can be exposed to FLA via accidental splash/squirt of contaminated water to the face or through an open wound, hence, necessitating this study in bridging the gaps of awareness on FLA.
Materials and methods
Ethics statement/Permission approval
This study was carried out in three selected Southeast Asian countries namely; Lao PDR, Myanmar and Singapore during 2013 to 2015. Invitation letters were approved from the relevant institutions namely University of Health Sciences, Vientiane, Lao PDR, Advancing Life and Regenerating Motherland (ALARM), Yangon, Myanmar, and National University of Singapore, Singapore. The collaborators from each country had been consulted at the time of water samples collection.
Sample collection
A total of 94 samples of either treated or untreated water were collected at various locations in Vientiane, Laos (31), Yangon, Myanmar (42) and Singapore (21) (Figs 1 and 2). From each sampling point, four samples were collected in 50 mL sterile centrifuge tubes. The tubes were submerged beneath the surface of untreated water, while treated water obtained from pipes was allowed to flow into the tubes. All of the water samples were transported to the laboratory and processed within 4 hours (hrs) after sampling.
Fig 1. Sampling locations in Laos, Myanmar, and Singapore.

Fig 2. Flowchart of the overall water analysis.

Physicochemical analysis of water quality
Physical parameters (YSI 556 Multiprobe System, USA) of the water samples such as turbidity (NTU), temperature (°C), total dissolved solids (mg/L), salinity (PSU), and dissolved oxygen (mg/L) content were measured at the sampling sites. Baseline chemical parameters (e.g., ammonia, chlorine, nitrite, nitrate, and fluoride) were measured in situ using colorimeter (Hach DR/890 Portable Colorimeter, USA) and recorded as mean values of the overall sites from each country.
Isolation of free-living amoebae
Prior to cultivation, the samples were centrifuged at 2000 rpm for 15 minutes (mins). One or two drops of sediment samples were spread onto the non-nutrient agar plates coated with a layer of Escherichia coli (NNA-E. coli). The plates were then incubated at room temperature (25–28°C) for 1 week.
Morphological observation and identification
The first line plates were observed daily (up to 7 days) using an inverted microscope. The morphological appearances of the amoebic isolates were identified according to Page’s taxonomy keys [25,26]. The trophozoite and cyst stages of the amoebae were flushed from the culture plates and were subjected for direct screening, Giemsa and FITC-DAPI staining. For each microscopy observation, 25 μL of amoebic isolates were placed on a glass slide. Both direct screening and Giemsa staining were examined using light microscope (Olympus BX51, Japan). Giemsa stain was prepared by submerging the methanol-fixed slide into a mixture of 20 mL of buffered distilled water (pH 7.1), followed by 40 drops of Giemsa stain. For FITC-DAPI staining, each sample was stained with 50 μL of FITC-MAb of Giardia/Cryptosporidium (CeLLabs, Brookvale, Australia) and incubated in a humidified chamber for 37°C for 30 mins, followed by washing step using PBS solution (pH 7.2). The sample was further stained with 50 μL of DAPI solution (Louis, Missouri, USA) for 2 mins before the addition of 50 μL of distilled water for 1 min. Finally, a 20 μL aliquot of mounting medium was placed onto the slide, covered with coverslip, and left air-dried prior to examination under epifluorescence microscope (Olympus BX51, Tokyo, Japan).
Thermotolerance assay
Thermotolerance assay was carried out for the selected FLA isolates (V. vermiformis and Acanthamoeba spp.) that had been confirmed through microscopy examination. Sub-culturing was performed as described above to obtain axenic culture of the amoeba. The samples were introduced to certain temperatures (34°C, 37°C, 40°C and 45°C) for a period of 24 hrs, respectively [27]. The isolates were then observed microscopically to confirm its viability.
DNA extraction
The trophozoites stage of each amoebic isolate were harvested from the plates with 5 mL of cold Page’s Amoeba Saline (PAS) and transferred into 1.5 mL tubes. DNA was further extracted using a commercial QIAamp DNA Blood Mini Kit (Qiagen, Hilden, France) following the manufacturer’s procedures and stored at -20°C until further analysis.
PCR amplification
For Acanthamoeba species, amplification was carried out by targeting the 18S region, with a PCR mix containing 10X DNA polymerase (Thermo Scientific, Lithuania, USA), 25 mM of magnesium chloride (MgCl2) (Thermo Scientific, Lithuania, USA), 10 mM of deoxynucleotide triphosphate (dNTP) mix (Thermo Scientific, Lithuania, USA) and 1 U/μL of Taq DNA polymerase (Thermo Scientific, Lithuania, USA) with 200 nmoles of each primer: JDP1 (5`-GGCCCAGATCGTTACCGTGAA–3`) and JDP2 (5`- TCTCACAAGCTGCTAGGGAGTCA– 3`) [28], with 5 ng of DNA templates. The reaction was performed at 94°C for 5 mins, followed by 40 cycles at 94°C for 1 min, 60°C for 1 min, 72°C for 1 min, and an extension at 72°C for 5 mins [28].
N. fowleri was detected by amplification of the ITS region (ITS1, 5.8S, and ITS2) using species-specific primers: the forward primer NFITSFW (5’-TGAAAACCTTTTTTCCATTTACA-3’) and the reverse primer NFITSRV (5’-AATAAAAGATTGACCATTTGAAA-3’) [29]. Amplifications were performed in a PCR mix containing 10X DNA polymerase buffer (Thermo Scientific, Lithuania, USA), 25 mM of magnesium chloride (MgCl2) (Thermo Scientific, Lithuania, USA), 10 mM of deoxynucleotide triphosphate (dNTP) mix (Thermo Scientific, Lithuania), 200 nmoles of each primers, and 1 U/μL of Taq DNA polymerase (Thermo Scientific, Lithuania, USA), with 5 ng of DNA templates. The PCR temperature profiles consisted of 94°C for 6 mins, followed by 30 cycles at 94°C for 1 min, 55°C for 1.5 mins, 72°C for 2 mins and the elongation step at 72°C for 10 mins [29].
The PCR reaction of H. vermiformis was performed in a 20 μL of Pre-mix (Thermo Scientific, Lithuania, USA) with 200 nmoles primers: the forward primer NA1 (5’-GCT CCA ATA GCG TAT ATT AA-3’) and the reverse primer NA2 (5’-AGA AAG AGC TAT CAA TCT GT-3’) [30] with 5 ng of DNA templates. The PCR cycling condition included the denaturation at 94°C for 1 min, followed by 35 repetition cycles at 94°C for 35 seconds, 56°C for 45 seconds, 72°C for 1 min, and the final elongation at 72°C for 5 mins [30].
Amplifications were conducted in a thermal cycler (BioRad, Hercules, USA) with the amount of 50–100 ng DNA templates in a final volume of 25 μL. PCR amplicon was run in a 1.5% agarose gel in Tris-Acetate-EDTA (TAE) buffer (Thermo Scientific, Lithuania, USA) at 100 V for 45 mins. A 100-bp DNA ladder was used to compare the expected amplicon sizes in the gel. Post-staining method was carried out using ethidium bromide (Amresco, Ohio, USA) and visualized under UV-transilluminator.
Nucleotide sequencing and phylogenetic analysis
Positive samples of PCR were purified using QIAamp PCR purification kit (Qiagen, Hilden, France) and sent for sequencing with the respective forward and reverse primers. A homology search was conducted for each sequence results with the FLA species deposited in the GenBank using Basic Local Alignment Search Tool (BLAST) software provided by the National Centre for Biotechnology Information (NCBI). The similarity between the species were compared by performing maximum-likelihood into phylogenetic tree using MEGA version 6 software, followed by Kimura 2-parameter algorithm with bootstrap analysis of 1000 replicates. In addition, V. vermiformis from ice-cube sample in our study was compared to a similar isolate (i.e. SS1 and SS2 strains) discovered from snow sample in Spain [31].
Results
Table 1 summarizes the physicochemical analysis of water quality parameters, presented as calculated means ± standard deviation (SD), with 95% confidence intervals (CIs). Overall results showed untreated water samples from Myanmar had the highest readings of turbidity (65.21±14.85 NTU; CI: 29.53), TDS (121.30±94.20 mg/L; CI: 35.90), chlorine (0.28±0.39 mg/L; CI: 0.18). The presence of Acanthamoeba and Vermamoeba were detected in water samples with high TDS and turbidity. In Myanmar, Vermamoeba was the only pathogenic FLA found in treated water samples with high nitrate concentration (2.13±6.03 mg/L; CI: 3.34). In Laos, Acanthamoeba was detected in untreated water samples with high level of nitrite (0.38±0.20 mg/L; CI: 0.13) and Vermamoeba was also detected in treated water samples with high level of ammonia (2.05±5.36 mg/L; CI: 3.10).
Table 1. The analysis of physicochemical water quality variables from treated and untreated samples in Laos, Myanmar, and Singapore.
| Country | Type of water | Mean, standard deviation (SD), and confidence intervals (CI) | Physicochemical water quality variables | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Physical | Chemical | |||||||||
| Turbidity | TDSd | Salinity | DOe | Chlorine | Nitrate | Nitrite | Ammonia | |||
| (NTU)c | (mg/l) | (PSU) | (mg/l) | (mg/l) | (mg/l) | (mg/l) | (mg/l) | |||
| Laos | Treateda | Mean | 4.05 | 50.75 | 0.05 | 3.11 | 0.19 | 0.19 | 0.10 | 2.05g |
| SD | 3.71 | 43.30 | 0.04 | 3.25 | 0.32 | 0.14 | 0.15 | 5.36 | ||
| CI (95%) | 2.70 | 63.70 | 0.03 | 2.82 | 0.26 | 0.15 | 0.11 | 3.10 | ||
| Untreatedb | Mean | 20.55i | 57.88i | 0.05 | 2.65 | 0.16 | 0.32 | 0.38f | 0.55 | |
| SD | 15.75 | 68.45 | 0.02 | 2.52 | 0.15 | 0.41 | 0.20 | 0.51 | ||
| CI (95%) | 8.06 | 28.62 | 0.15 | 0.75 | 0.05 | 0.14 | 0.13 | 0.17 | ||
| Myanmar | Treateda | Mean | 1.34 | 85.30 | 0.25 | 5.42 | 0.22 | 2.13h | 0.05 | 0.74 |
| SD | 1.99 | 117.80 | 0.54 | 6.81 | 0.75 | 6.03 | 0.03 | 1.90 | ||
| CI (95%) | 0.85 | 50.75 | 0.39 | 2.46 | 0.31 | 3.34 | 0.04 | 1.42 | ||
| Untreatedb | Mean | 65.21i | 121.30i | 1.39 | 2.55 | 0.28 | 1.29 | 0.35 | 0.19 | |
| SD | 14.85 | 94.20 | 3.88 | 1.85 | 0.39 | 0.93 | 1.14 | 0.25 | ||
| CI (95%) | 29.53 | 35.90 | 0.95 | 0.36 | 0.18 | 0.15 | 0.27 | 0.08 | ||
| Singapore | Treateda | Mean | 1.01 | 15.13 | 0.03 | 0.52 | 0.25 | 0.20 | 0.01 | 0.13 |
| SD | 0.54 | 25.15 | 0.01 | 0.92 | 0.18 | 0.18 | 0.01 | 0.15 | ||
| CI (95%) | 0.55 | 16.11 | 0.02 | 0.54 | 0.25 | 0.14 | 0.00 | 0.16 | ||
| Untreatedb | Mean | 5.48 | 30.25 | 0.05 | 1.64 | 0.22 | 0.19 | 0.20 | 0.42 | |
| SD | 2.35 | 21.95 | 0.03 | 1.82 | 0.45 | 0.24 | 0.32 | 0.31 | ||
| CI (95%) | 1.88 | 48.87 | 0.04 | 0.83 | 0.20 | 0.17 | 0.13 | 0.18 | ||
aTreated water includes drinking water, water dispenser, mineral water, tap water, and swimming pools
bUntreated water includes rain water, springs, wells, recreational lake, rivers, waterfalls, canals/channels and effluent water
cNTU is nephelometric turbidity unit
dTDS is total dissolved solids
eDO is dissolved oxygen
fAcanthamoeba was detected in untreated water samples with high level of nitrite
hVermamoeba was detected in treated water samples with high concentration of nitrate
iAcanthamoeba and Vermamoeba were detected in untreated water samples with high level of TDS and turbidity
Initially, positive amoebic-like cells exist as mixed isolates with other organisms. The trophozoite and cyst stages were normally seen after 2–3 days and at least 5–6 days of cultivation, respectively. Both stages were photographed in Fig 3. All isolates were grown at room temperature. The morphology of all isolates can be clearly differentiated based on its motile stage of trophozoite/flagella. The Acanthamoeba-like cell exhibited irregular shape with distinct projection of pseudopodia/acanthopodia in multidirectional movement. The Vermamoeba-like trophozoite showed a predominant cylindrical form with monopodia and moving in one direction. Meanwhile, for Naegleria-like cell, the flagellate stage was observed on the watery surface of the medium agar (not shown).
Fig 3. Morphological observations of trophozoite and cyst.
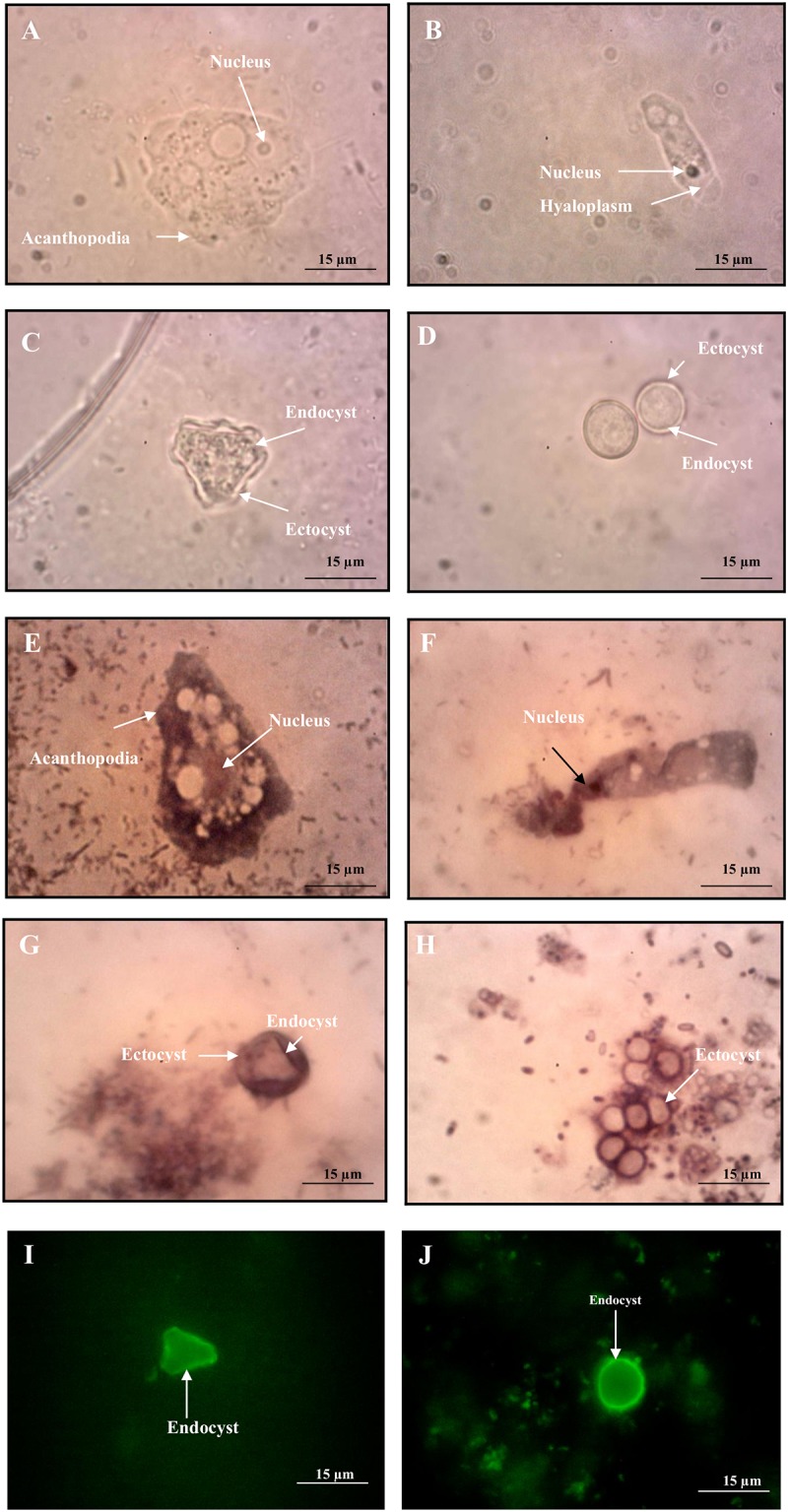
(A,E) Acanthamoeba trophozoite, (C,G) Acanthamoeba cyst, (B,F) Hartmannella trophozoite, (D,H) Hartmannella cyst under light microscope (LiM) and (I) Acanthamoeba cyst, (J) Hartmannella cyst under epifluorescencce microscope (EpM). (A, LiM X 400; E, LiM X 400) Acanthamoeba trophozoites showing typical eruptive pseudopodia/lobopodia with no stain (A) and Giemsa stain (E) (C, LiM X 400; G, LiM X 400) A single Acanthamoeba cyst showing smooth ectocyst and endocyst with no stain (C) and Giemsa stain (G) (B, LiM X 400; F, LiM X 400) Vermamoeba trophozoite with no stain (B) and Giemsa stain (F) (D, LiM X 400; H, LiM X 400) Rounded form of Vermamoeba cyst with no stain (D) and Giemsa stain (H) (I, EpM X 400; J, EpM X 400) Triangular shape of Acanthamoeba cyst (I) and rounded form of Vermamoeba cyst (J) with immunofluorescence stain.
Two staining methods (i.e. Giemsa and immunofluorescence) were used for the morphological identification of pathogenic amoebae and were compared with the non-stained slides as a control. In control slides, the nucleus and cytoplasm of the trophozoite were clearly distinguished, whereas two distinct layers of the cyst were observed for both Acanthamoeba and Vermamoeba isolates (Fig 3A–3D). For Giemsa, the trophozoite and cyst of the parasites were stained purple and showed a good contrast with the background. The eruptive pseudopodia of Acanthamoeba and the cylindrical form of Vermamoeba trophozoites were seen but unclear colour contrast between nucleus and cytoplasm was encountered (Fig 3E and 3F). Nevertheless, both outer and inner layers of the cysts were successfully observed as dark purple in colour (Fig 3G and 3H). The USEPA method 1623 was implemented to stain the amoebic cysts, similarly to Cryptosporidium and Giardia (oo)cysts. The inner layer was stained green, whereas the outer layer was appeared unstained against the dark background (Fig 3I and 3J). From the observation of thermotolerance assay, the trophozoites did not transformed into cyst stage for all of the tested temperatures.
The Vermamoeba-like trophozoite was detected in 1 of the 11 treated samples (9.1%) and 2 out of 31 untreated (6.5%) from Myanmar. In Laos, it was detected in 1 out of 9 treated samples (11.1%). Vermamoeba sp. was not detected in the samples collected in Singapore (Table 2). Naegleria-like flagella was the most frequently encountered in both treated and untreated water samples (Table 2). From Myanmar samples, it was found in all 11 treated water samples (100%) and 28 out of 31 untreated samples (90.3%). Samples from Laos showed the presence of flagella in 7 out of 9 treated samples (77.8%) and 18 of the 22 untreated samples (81.8%). Meanwhile, flagella were observed in 8 out of 15 untreated samples (53.3%) from Singapore. The Acanthamoeba isolate was observed only in untreated water samples (Table 2); 3 out of 31 samples (9.7%) from Myanmar, 1 out of 22 samples (4.5%) from Laos, and 1 of the 15 samples (6.7%) from Singapore.
Table 2. The occurrence of pathogenic FLA via microscopic examination and PCR in Laos, Myanmar, and Singapore.
| Country of origin | Type of water | No. of samples | Free-living amoeba (FLA) | |||||
|---|---|---|---|---|---|---|---|---|
| Vermamoeba sp. | Naegleria sp. | Acanthamoeba sp. | ||||||
| M (n) | PCR (n) | M (n) | PCR (n) | M (n) | PCR (n) | |||
| Laos | Treateda | 9 | 1 | V. vermiformis (1) | 7 | NDc | NDc | NDc |
| Untreatedb | 22 | NDc | NDc | 18 | NDc | 1 | A. lenticulata (1) | |
| Myanmar | Treateda | 11 | 2 | V. vermiformis (2) | 11 | NDc | NDc | NDc |
| Untreatedb | 31 | 2 | V. vermiformis (2) | 28 | NDc | 3 | A. lencticulata (2); A. triangularis (1) | |
| Singapore | Treateda | 6 | NDc | NDc | NDc | NDc | NDc | NDc |
| Untreatedb | 15 | NDc | NDc | 8 | NDc | 1 | NDc | |
| Total | 94 | |||||||
aTreated water includes drinking water, water dispenser, mineral water, tap water, and swimming pools
bUntreated water includes rain water, springs, wells, recreational lake, rivers, waterfalls, canals/channels and effluent water
cND = not detected; M = Microscopy; n = Number of samples; PCR = Polymerase chain reaction; V = Vermamoeba; A = Acanthamoeba
After PCR amplification, none of the 94 isolates showed amplicon bands of N. fowleri using the ITS species-specific primer. For Acanthamoeba typing, the DF3 region of 18S rRNA was amplified, producing a 450bp amplicon. A. triangularis was detected from one isolate (KX232518) from Myanmar and formed a clade with both A. triangularis (AF346662) and A. polyphaga (AF019061) in T4 group. Another two isolates (KX232517, KX232519) from Myanmar and one isolate (KX232520) from Laos were identified as A. lenticulata. They were grouped as T5 and formed a clade with A. lenticulata (U94739). Balamuthia mandrillaris (AF477022) that doesn’t contain DF3 region showed distantly related with the Acanthamoeba groups (Table 3, Fig 4). A 800 bp PCR product was obtained for all of the Vermamoeba isolates (KX856370, KX856371, KX856372, KX856373, KX856374) and sequence analysis revealed that the amoebae had a high homology of 99% to V. vermiformis (KC188996). All other V. vermiformis strains, including 5 environmental sequences from present study, clearly showed a clade with each other under the group of Vermamoebidae. At the family level, E. exundans formed a distinct group of Echinamoebidae against the Vermamoebidae, within the order of Echinamoebida (Table 3, Fig 5). V. vermiformis pairwise distance between ice cube isolate and snow strain from Spain showed 0.5% intraspecific variation. BLAST results revealed the similarity at 99% and 98% of ice cube isolate-SS1 and ice cube isolate-SS2 strains, respectively (data not shown).
Table 3. Pathogenic FLA isolated from selected Southeast Asian countries.
| Pathogenic FLA | Code | Accession number | Country of origin | Source of water |
|---|---|---|---|---|
| A. lenticulata | M.ut.1 | KX232517 | Myanmar | Well water |
| A. lenticulata | M.ut.2 | KX232519 | Myanmar | Recreational lake |
| A. lenticulata | L.ut.1 | KX232520 | Laos | Mekong up 2 |
| A. triangularis | M.ut.3 | KX232518 | Myanmar | Fish pond 2 |
| V. vermiformis | M.t.1 | KX856371 | Myanmar | Ice cube |
| V. vermiformis | M.t.2 | KX856372 | Myanmar | Swimming pool 2 |
| V. vermiformis | M.ut.1 | KX856373 | Myanmar | Well water |
| V. vermiformis | M.ut.3 | KX856374 | Myanmar | Fish pond 2 |
| V. vermiformis | L.t.1 | KX856370 | Laos | Swimming pool adult |
V = Vermamoeba; A = Acanthamoeba; FLA = Free-living amoeba
Fig 4. Phylogenetic analysis of pathogenic free-living Acanthamoeba spp. isolated from various types of water sources in selected Southeast Asian countries.

Fig 5. Phylogenetic analysis of pathogenic free-living Vermamoeba vermiformis isolated from various types of water sources in selected Southeast Asian countries.

Discussion and conclusions
Investigations of free-living amoebae have been dramatically increased because of their important role within the ecosystems and their ability to cause serious infections among humans. With the ever growing number of FLA cases, various studies had been carried out throughout the world, especially from environmental waters, in assisting to decrease the potential source of contamination. Several studies conducted in Peninsular Malaysia [15,16] and the Philippines [19] showed occurrence of FLA in environmental water samples even they were proven to be non-pathogenic using PCR assay. To our knowledge, the present study is the first comprehensive report of the occurrence of potentially pathogenic FLA in both treated and untreated water samples from selected Southeast Asian countries.
Here, we investigated the occurrence of FLA both from raw and treated water sources from three (3) selected Southeast Asian countries and demonstrated the distribution of the potential pathogenic species by both microscopy and molecular approaches. Following PCR and sequencing, only 2 families of potentially pathogenic amoeba belongs to the genera of Acanthamoeba (Acanthamoebidae) and Vermamoeba (Vermamoebidae) were found. Microscopically, Myanmar reported the highest occurrence of FLA (92%, 39/42) followed by Laos (87%, 27/31), and Singapore (43%, 9/21). The rate of FLA in Laos and Myanmar were surprisingly found higher in comparison to a previous study conducted in this region such as Thailand (45.2%, 43/95) [17], though these countries share much similarity in geographical distribution. Moreover, this figure is remarkably higher than other countries namely; Japan (49.5%, 47/95) [17], USA (43.4%, 143/330) and Iran (35%, 42/120), but lower than Bulgaria (93.9%, 31/33) [32–34].
The presence of both Acanthamoeba and Vermamoeba were revealed with high values for both turbidity and TDS. High turbidity showed that the water contained other soluble matters that can provide nutrition and encourage growth rate for the amoeba. The high reading of turbidity demonstrated that these sediments supported the microbial growth and in return, becoming the source of food for the amoeba [35]. This finding also supported by a study conducted in the Philippines’s water sources with the occurrence of Acanthamoeba and Naegleria that were rich in TDS [19]. This scenario will lead to reduction of water quality which reflects the nutrient availability that could help the parasites to revive in a suitable environment. The result also showed the ability of A. triangularis to uphold high concentration of nitrite (> 0.5 ppm) and ammonia (> 0.5 ppm) [36]. The high content of both nitrite and ammonia assist in the growth of Gram-negative bacteria that may become the food source for A. triangularis. In addition, nitrite was also found to be a good indicator for the presence of other parasites, namely, Cryptosporidium and Giardia [19, 37].
Another study was conducted in France, showing a wide biodiversity of FLA in water treatment plants that used river water as its main source [38]. A similar study was also carried out in Sarawak ofEast Malaysia that revealed the presence of FLA in various processing sites in treatment plants [39]. Relatively, similar treatment processes that include chlorination and filtration are practiced in Myanmar and Laos. Given this, the treated water in both aforementioned countries was contaminated with V. vermiformis. This may be due to the ability of V. vermiformis cyst to resist the harsh condition of physical and chemical treatments that are used in the treatment plants.
Cultivation of FLA obtained from water samples was carried out to impede excessive growth of unwanted contaminants such as bacteria and fungi. Hence, this technique was chosen as it is able to grow large quantity of FLA within a short period of time. FLA is able to grow at room temperature (i.e. 25°C), as higher temperature may trigger the overgrowth of bacteria instead of amoeba. The high ratio of bacteria to amoeba (i.e. 10 > 1) that contained in the uncultured water can suppress the growth of Acanthamoeba [40]. The cultivation method was performed on the NNA (non-nutrient agar) with non-mucoid bacteria (i.e. Escherichia coli) as the food source. In addition, bacteria with mucoid were not preferable as it may delay the phagocytosis by amoeba that may leads to bacterial overgrowth. Sub-cultivation was also performed to assist in removing debris that may contain living/dead bacterial cells, trapped inorganic particles, and organic fibers [41]. Furthermore, the contaminants may obstruct the detection of FLA, especially those obtained from environmental water samples. Staining of the stages of all pathogenic FLA revealed a better observation of the features (i.e. shape) and cellular organelles. The smears of the samples were firstly fixed using methanol to prevent the trophozoites and cysts to become distorted or shrunk. Giemsa, which is commonly used for blood parasites, has produced good contrast against the background for both trophozoites and cysts in the present study. Nevertheless, the space between the endocyst and ectocyst were also stained blue and can cause confusion in determining or confirming the distinct features of both layers. Moreover, trophozoite possessed a better contrast due to its larger size in comparison to cysts. The findings from this study showed that Giemsa stain can be used, especially for routine screening to identify the developmental stages of FLA in general, and trophozoites detection in particular, for clinical epidemiology and public health purposes.
In addition, fluorescence method is used in FLA identification based on its concept of conjugation of protein-antigen and proved to be useful because of the failure of Giemsa stain intake by the cysts. Only endocyst was stained with bright apple-green colour and able to differentiate the number of arms and polygonal shapes for detailed morphological classification of pathogenic Acanthamoeba. Hence, it is revealed that the endocyst of FLA consisted of cellulose, a similar material that covers the (oo)cysts of Cryptosporidium and Giardia [42] that enables simultaneous detection in detecting waterborne parasites containing FLA, Cryptosporidium, and Giardia.
Overall, observation of the trophozoite stage of FLA by microscopy has permitted the differentiation at genus level. The trophozoites of Naegleria were smaller in size as compared to those of Acanthamoeba and Vermamoeba and were observed to move faster in a unidirectional manner. Vermamoeba trophozoites with a transient cylindrical form possessed similar pattern of movement as Naegleria. Acanthamoeba showed multiple shapes of trophozoites and moving by the projection of lobopodia/acanthopodia in multidirectional. It is known that the classification of FLA based on morphological criteria is insufficient, while their identification is not problematic using different PCR assays, including conventional PCR.
The diagnosis of infection and identification of pathogenic FLA remains unsatisfactory, time-consuming, expensive, laborious, and prone to ethical issues when mouse pathogenicity test was taken into consideration. Due to the advancements in molecular detection, conventional PCR has been developed and seem to be reliable method for routine screening of FLA in environmental samples. One-step based PCR, paired with specific primer by targeting the region of ITS and 18S rDNA [27] is sufficient enough to differentiate between and within species of various organisms. Furthermore, the primer sets used in the present study showed high specificity and capable to amplify a product from potentially pathogenic FLA genotypes, but not with other closely related genera of amoeba. For example, the JDP1-JDP2 primers had been proven to produce an amplicon even from a single trophozoite of Acanthamoeba [27].
The internal transcribed spacers (ITS) region has been chosen to study the differences within the genus of Naegleria [43]. The ITS analysis has been used in studying heterogeneity in several organisms such as Cryptosporidium parvum, trichomonadid protozoa, and Valkamphia [42,44]. The species-specific primer was used to detect the presence of pathogenic N. fowleri [45]. Nevertheless, no positive amplification of N. fowleri was obtained against the Naegleria-like isolates. This might be due to the fact that N. fowleri is a thermophilic organism that proliferates at an ambient temperature (as high as 45°C) [46].
On the other hand, four of the environmental Acanthamoeba-like isolates were successfully amplified using the species-specific primer of JDP1 and JDP2, producing approximately 460 bp amplicons of potentially pathogenic A. triangularis and A. lenticulata. The sequence of 18S rDNA from Acanthamoeba isolates led into 13 different lineages containing either single or complexes of species. Based on the classification, both A. triangularis and A. lenticulata had been classified as type T4 and T5, respectively, and often been associated with human systemic infection and keratitis. Many previous studies similarly reported that T4 is the most prevalent genotype among both clinical specimens and environment samples. The presence of both isolates in our samples probably reflects their better adaptation to various growth conditions relative to isolates from other genotypes.
Morphology of Vermamoeba vermiformis isolates from the present study was similar to previously reported species under the microscope [47]. The typical trophozoites of Vermamoeba displayed worm-like cell body, stable anterior hyaline cap, and possess a tendency branch when they changed a direction. The 18S rRNA gene sequences of all reported V. vermiformis isolates in present study showed extremely high similarity with more than 96% identical to AF426157 and KC188996. Further, phylogenetic analysis of 18S rRNA gene sequences in Amoebozoa revealed that Hartmannellidae and Vermamoebidae are paraphyletic and monophyletic group, respectively.
Findings conducted throughout the world had reported that the Vermamoeba species is considered pathogenic that is also tolerant towards high temperature [48]. The thermotolerance behaviour is one of the factors that contributed to the virulence characteristic that able to cause diseases and leads to death. Interestingly, our study revealed positive isolate from cold condition of ice cube that are similar to the result carried out in Spain [31]. Thus, it was purported that V. vermiformis has the ability to survive in various conditions, besides exhibiting thermotolerant properties.
The presence of these parasites is rather alarming due to its ability in causing parasitic infections. Site observation in Myanmar and Laos revealed that fishing activities are prominent in the rivers and ponds. Consumption of these contaminated fishes (raw or under-cooked) with potentially pathogenic FLA could cause a fatal disease to humans. A previous study revealed that freshwater fishes contained several Hartmannella vermiformis strains found from its organs [49], thus, there is a possibility that fish can be a host for this parasite. However, FLA transmissions via food are still questionable and more studies in the future need to be carried out to further proven the presence of amoeba within host tissues.
For the past decade, keratitis cases had increased dramatically parallel with the popularity of contact lens usage, hence, resulting in more reports of clinically confirmed cases in Southeast Asia. Acanthamoeba species was revealed to be the main cause of keratitis infection in raw water in Thailand [50,51] and Vietnam [52], in regards with water-related activities (i.e. swimming and diving) and accidental water-splashing. In the present study, A. triangularis was found in fish pond sample obtained in Myanmar. A previous study has confirmed the role of A. triangularis in causing human keratitis [53]. Usually, Acanthamoeba spp. can be isolated from environmental water that is rich with sediments and other particulate matters [54]. In addition, our study confirmed the presence of A. lenticulata in water sources such as Mekong River, well water, as well as recreational lake in Myanmar and Laos. This amoeba has also been reported causing granulomatous amoebic encephalitis (GAE) in immunocompromised patients [55].
The occurrence of FLA in treated water cannot be neglected due to the increasing reports of keratitis in Malaysia [56], Indonesia [57], and the Philippines [58]. Keratitis had always been associated with the infection caused by Acanthamoeba, but Hartmannella (= Vermamoeba) too, is able to cause similar symptoms of watery eyes, redness and blurred vision [59,60]. In the present study, V. vermiformis was isolated mostly from treated water of swimming pools from Myanmar and Laos. This situation revealed that V. vermiformis is able to adapt in chlorinated water and showed the ability to survive in the ecology. Although limited epidemiological data available, chlorinated swimming pool can possibly be a new niche for FLA to survive. The alarming phenomenon is considered a major concern, especially to contact lens wearers as they are susceptible to keratitis infection if lens are not appropriately handled especially before and after entering the pool. In addition, the pipe that connects water into the swimming pools may contain biofilm that support the colonization of FLA [61].
From this finding, the isolation of FLA revealed a better understanding on the distribution and prevalence of both pathogenic Acanthamoeba and Vermamoeba in Myanmar and Laos. The high occurrence of amoebae in Myanmar must not be neglected by the local authorities to ensure good water quality is supplied to the consumers. Proper precaution measures in ensuring water quality must be carried out to assist in regular monitoring on any possibilities of the emerging amoebae. This is because of the wide occurrences of FLA is not only limited to one species at any particular water source or area. Meanwhile, water that is commonly used by public, such as swimming pool, must undergo efficient and appropriate treatment processes. The usage of alternative disinfectants such as Baquacil, chlorinated cyanurate and chlorine dioxide can be added into the water to control the growth of FLA [62,63]. Raw water that is used for recreational purposes (i.e. hotspring, coastal) must also be monitored closely as the untreated water may contain high number of FLA, thus, can transmit infections to the public [64,65]. In addition, the infection caused by FLA is still misdiagnosed or under-reported, particularly in this region. Thus, future investigations of the occurrence as well as distribution of FLA in treated and untreated water with a larger sample size need to be taken in serious consideration. The findings from the investigations will be reported to relevant authorities to prevent further widespread of water contamination with potentially pathogenic species of these free-living parasites.
Acknowledgments
The authors would like to thank Ms. Thulasi Kumar and Ms. Subashini Onichandran from Malaysia, technical staffs from Lao PDR; Dr. Aye Aye Win and Ms. Lin Myat Myat Aung from Myanmar, as well as Mr. Meng-Hwee Lim from Singapore, who helped us with the water sampling and laboratory works. We would also like to thank Dr. Kruawan Chotelersak from Srinakharinwirot University, Thailand, and Dr Romano Ngui from Malaysia for phylogenetic analysis, Dr Hany Sady from Yemen for thermotolerance assay and Ms. Alicia JC Choo from Malaysia for graphic improvement and data retrieved from external sources. Last but not least, we would also like to convey our deepest gratitude to Prof. Dr. Jacob Lorenzo-Morales from University of La Laguna, Spain for providing us the sequences of Vermamoeba vermiformis strains obtained from snow sample. This work was presented in part at the 1st International Conference on Pollutant Toxic Ions and Molecules (PTIM2015) on 2nd– 4th November, 2015 in Caparica, Portugal.
Data Availability
All relevant data are within the paper.
Funding Statement
University of Malaya High Impact Research Grant (UM.C/625/1/HIR/MoHE/MED/23 and UM. 0000041/HIR.C3) and University of Malaya Research Grants (UMRG 544/14HTM and UMRG 362/15AFR), received by VN and TTC.
References
- 1.Visvesvara GS, Moura H, Schuster FL. Pathogenic and opportunistic free living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immun Med Microbiol. 2007;50: 1–26. [DOI] [PubMed] [Google Scholar]
- 2.Qvarnstrom Y, da Silva AJ, Schuster FL, Gelman BB, Visvesvara GS. Molecular confirmation of Sappinia pedata as a causative agent of amoebic encephalitis. J Infect Dis. 2009;199: 1139–1142. 10.1086/597473 [DOI] [PubMed] [Google Scholar]
- 3.Schuster FL, Visvesvara GS. Free-living amoebae as opportunistic and non-opportunistic pathogens of humans and animals. Int J Parasitol. 2004;34: 1001–1027. 10.1016/j.ijpara.2004.06.004 [DOI] [PubMed] [Google Scholar]
- 4.Khan NA. Acanthamoeba: biology and increasing importance in human health. FEMS Microbiol Rev. 2006;30: 564–595. 10.1111/j.1574-6976.2006.00023.x [DOI] [PubMed] [Google Scholar]
- 5.Teixera LH, Rocha S, Pinto RMF, Caseiro MM, da Costa SOP. Prevalence of potentially pathogenic pathogenic free-living amoebae from Acanthamoeba and Naegleria genera in non-hospital, public, interval environments from the city of Santos, Brazil. Braz J of Infect Dis. 2009;13: 395–397. [DOI] [PubMed] [Google Scholar]
- 6.Greub G, Raoult D. Biocides currently used for bronchoscope decontamination are poorly effective against free-living amoebae. Infect Cont Hosp Ep. 2003;24: 784–786. [DOI] [PubMed] [Google Scholar]
- 7.Marciano-Cabral F, Cabral G. Acanthamoeba spp. as agents of disease in humans. Clin Microbiol Rev. 2003;16: 273–307. 10.1128/CMR.16.2.273-307.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Marciano-Cabral F, Puffenbarger R, Cabral GA. The increasing importance of Acanthamoeba infections. J Eukaryot Microbiol. 2000;47: 29–36. [DOI] [PubMed] [Google Scholar]
- 9.Corsaro D, Walochnik J, Kohsler M, Rott MB. Acanthamoeba misidentification and multiple labels: redefining genotypes T16, T19, and T20 and proposal for Acanthamoeba micheli sp. nov. (genotype T19). Parasitol Res. 2015;114: 2481–2490. 10.1007/s00436-015-4445-8 [DOI] [PubMed] [Google Scholar]
- 10.De Jonckheere JF. A century of research on the amoeboflagellate genus Naegleria. Acta Protozool. 2002;47: 309–342. [Google Scholar]
- 11.Kaushal V, Chhina DK, Ram S, Singh G, Kaushal RK, Kumar R. Primary Amoebic Meningoencephalitis due to Naegleria fowleri. J Assoc Phy Ind. 2008;56: 459–62. [PubMed] [Google Scholar]
- 12.Aitken D, Hay J, Kinnear FB, Kirkness CM, Lee WR, Seal DV. Amoebic keratitis in a wearer of disposable contact lenses due to a mixed Vahlkampfia and Hartmannella infection. Ophthalmology. 1996;103: 485–494. [DOI] [PubMed] [Google Scholar]
- 13.Brieland JK, Fantone JC, Remick DG, LeGendre M, McClain M, Engleberg NC. The role of Legionella pneumophila infected Hartmannella vermiformis as an infectious particle in a murine model of Legionnaires’ disease. Infect Immun. 1997;65: 5330–5333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Greub G, Raoult D. Microorganisms resistant to free-living amoeba. Clin Microbiol. 2004;17: 413–433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Init I, Lau YL, Arine FA, Foead AI, Neilson RS, Nissapatorn V. Detection of free-living amoebae, Acanthamoeba and Naegleria, in swimming pools, Malaysia. Trop Biomed. 2010;27: 566–577. [PubMed] [Google Scholar]
- 16.Ithoi I, Ahmad AF, Nissapatorn V, Lau YL, Mahmud R, Joon WM. Detection of Naegleria species in environmental samples from Peninsular Malaysia. Plos One. 2011;6: 324–327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nacapunchai D, Kino H, Ruangsitticha C, Sriwichai P, Ishih A, Terada M. A brief survey of free-living amebae in Thailand and Hamamatsu District Japan. Southeast Asian J Trop Med Public Health. 2001;32: 179–182. [PubMed] [Google Scholar]
- 18.Herriman R. Vietnam reports second ever ‘brain-eating amoeba’ death. 2012. http://www.examiner.com/article/vietnam-reports-second-ever-brain-eating-amoeba-death.
- 19.Onichandran S, Kumar T, Salibay CC, Dungca JZ, Tabo HAL, Tabo N, et al. Waterborne parasites: a current status from the Philippines. Parasit Vectors. 2014;7: 244 10.1186/1756-3305-7-244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ghani MKA, Annuar FH, Ahmad N, Abdullah NR, Hay J, Seal D. A case of waterborne contact lens associated Acanthamoeba keratitis from Malaysia: Successful treatment with Chlorhexidine and Propamidine. Int Med J. 2000;7: 63–65. [Google Scholar]
- 21.Por YM, Mehta JS, Chua JLL, Koh TH, Khor WB, Fong AC, et al. Acanthamoeba keratitis associated with contact lens wear in Singapore. Am J Ophthalmol. 2009;148: 1879–1891. [DOI] [PubMed] [Google Scholar]
- 22.Bunjongpuk S. Primary amoebic meningoencephalitis (PAM). Thai J Paed. 2000;39: 38–49. [Google Scholar]
- 23.Sithinamsuwan P, Sangruchi T, Chiewvit P, Poungvarin N. Free-living amoeba infections of the central nervous system in Thailand. Report of two patients. Int Med J Thai. 2000;17: 350–360. [Google Scholar]
- 24.Intalapaporn P, Suankratay C, Shuangshoti S, Phantumchinda K, Keelawat S, Wilde H. Balamuthia mandrillaris meningoencaphalitis: the first case in Southeast Asia. Am J Trop Med Hyg. 2004;70: 666–669. [PubMed] [Google Scholar]
- 25.Page FC. Taxonomic criteria for limax amoebae, with descriptions of 3 new species of Hartmannella and 3 of Vahlkampfia. J Protozool. 1967;14: 499–521. [DOI] [PubMed] [Google Scholar]
- 26.Page FC. A new key to freshwater and soil gymnamoebae. Freshwater Biol Assoc, Ambleside; 1988. UK [Google Scholar]
- 27.Qvarnstrom Y, Nerad TA, Visvesvara GS. Characterization of a new pathogenic Acanthamoeba species, A. byersi n. sp., isolated from a human with fatal amoebic encephalitis. J Eukaryot Microbiol. 2013;60(6): 626–633. 10.1111/jeu.12069 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Schroeder JM, Booton GC, Hay J, Niszl IA, Seal DV, Markus MB, et al. Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification of Acanthamoeba from humans with keratitis and from sewage sludge. J Clin Microbiol. 2001;39: 1903–1911. 10.1128/JCM.39.5.1903-1911.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Pelandakis M, Serre S, Pernin P. Analysis of the 5.8S rRNA gene and internal transcribed spacers in Naegleria spp. and in N. fowleri. J Eukaryot Microbiol. 2000;47: 116–121. [DOI] [PubMed] [Google Scholar]
- 30.Lasjardi Z, Niyyati M, Haghighi A, Shahabi S, Biderouni FT, Taghipour N, et al. Potentially pathogenic free-living amoebae isolated from hospital wards with immunodeficient patients in Tehran, Iran. Parasitol Res. 2011;109: 575–580. 10.1007/s00436-011-2288-5 [DOI] [PubMed] [Google Scholar]
- 31.Reyes-Batlle M, Niyyati M, Martin-Navarro CM, Lopez-Arencibia A, Valladares B, Martinez-Carretero E, et al. Unusual Vermamoeba vermiformis strain isolated from snow in Mount Teide, Tenerife, Canary Islands, Spain. Novelty in Biomedicine. 2015;4: 189–192. [Google Scholar]
- 32.Ettinger MR, Webb SR, Harris SA, McIninch SP, Garman G, Brown BL. Distribution of free-living amoebae in James River,Virginia, USA. Parasitol Res. 2003;89: 6–15. 10.1007/s00436-002-0707-3 [DOI] [PubMed] [Google Scholar]
- 33.Sh Ghadar-ghadr, Solhjoo K, Norouz-nejad MJ, Rohi R, Zia-Jahromi S. Isolation and identification of free living amoeba (Naegleria and Acanthamoeba) in Shiraz water resources by morphological criteria. J Jahrom Uni Med Sci. 2012;10: 26–33. [Google Scholar]
- 34.Tsvetkova N, Schild M, Panaiotov S, Kurdova-Mintcheva R, Gottstein B, Walochnik J, et al. The identification of free-living environmental isolates of amoebae from Bulgaria. Parasitol Res. 2004;92: 405–413. 10.1007/s00436-003-1052-x [DOI] [PubMed] [Google Scholar]
- 35.Valster RM, Wullings BA, van den Berg R, van der Kooji D. Relationships between free-living protozoa, cultivable Legionella spp., and water quality characteristics in three drinking water supplies in the Carribean. Appl Environ Microbiol. 2011;77: 7321–7328. 10.1128/AEM.05575-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Patil PN, Sawant DV, Deshmukh RN. Physico-chemical parameters for testing of water—A review. Int J Environ Sci. 2012;3: 1194–1207. [Google Scholar]
- 37.Natividad FF, Buerano CC, Lago CB, de Guzman BB, Seraspe EB, Samentar LP, et al. Prevalence rates of Giardia and Cryptosporidium among diarrheic patients in the Philippines. Southeast Asian J Trop Med. 2008;39: 991–999. [PubMed] [Google Scholar]
- 38.Thomas V, Loret JF, Jousset M, Greub G. Biodiversity of amoebae and amoebae-resisting bacteria in a drinking water treatment plant. Environ Microbiol. 2008;10: 2728–2745. 10.1111/j.1462-2920.2008.01693.x [DOI] [PubMed] [Google Scholar]
- 39.Richard RL, Ithoi I, Abd Majid MA, Wan Sulaiman WY, Tan TC, Nissapatorn V, Lim YAL. Monitoring of waterborne parasites in two drinking water treatment plants: a study in Sarawak, Malaysia. Int J Environ Res Public Health. 2016;13(7): 641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Schuster FL. Cultivation of pathogenic and opportunistic free-living amoebas. Clin Microbiol Rev. 2002;15: 342–354. 10.1128/CMR.15.3.342-354.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Eikelboom DH. Characteristics of activated sludge flocs In: Process control of activated sludge plants by microscopic investigation. United Kingdom: IWA Publishing; 2000. pp.29–41. [Google Scholar]
- 42.Brian JH. Microscopy of 4 Pathogenic Enteric Protozoan Parasites: A Review. Lab Med. 2008;39: 231–238. [Google Scholar]
- 43.Pelandakis M, Pernin P. Use of multiplex PCR and PCR restriction enzyme analysis for detection and exploration of the variability in the free-living amoeba Naegleria in the environment. Appl Environ Microbiol. 2002;68: 2061–2065. 10.1128/AEM.68.4.2061-2065.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Gartstecki T, Brown S, De Jonckheere JF. Description of Vahlkamphia signyensis sp. (Heterolobosea), based on morphological, ultrastructural and molecular characteristics. Eur J Protistol. 2005;41: 119–127. [Google Scholar]
- 45.De Jonckheere JF. Molecular definition and the ubiquity of species in the genus Naegleria. Protistol. 2004;155: 89–103. [DOI] [PubMed] [Google Scholar]
- 46.Izumiyama S, Yagita K, Furushima-Shimogawara R, Asakura T, Karasudani T, Endo T. Occurrence and distribution of Naegleria species in thermal waters in Japan. J Eukaryot Microbiol. 2003;50: 514–515. [DOI] [PubMed] [Google Scholar]
- 47.Smirnov AV, Brown S. Guide to the methods of study and identification of soil gymnamoebae. Protistol. 2004;3: 148–190. [Google Scholar]
- 48.Solgi R, Niyyati M, Haghighi A, Nazemalhosseini Mojarad E. Occurrence of thermotolerant Hartmannella vermiformis and Naegleria spp. in hot springs of Ardebil Province, Northwest Iran. Iranian J Parasitol. 2012;7(2): 47–52. [PMC free article] [PubMed] [Google Scholar]
- 49.Dyakoya I, Pindova Z, Fiala I, Dvorakova H, Machackova B. Fish-isolated of Hartmannella vermiformis Page, 1967: morphology, phylogeny and molecular diagnosis of the species in tissue lesions. Folia Parasitologica. 2005;52: 295–303. [PubMed] [Google Scholar]
- 50.Jongwutiwes S, Pariyakanok L, Charoenkorn M, Yagita K, Endo T. Heterogeneity in cyst morphology within isolates of Acanthamoeba from keratitis patients in Thailand. Trop Med Int Heal. 2000;5: 335–340. [DOI] [PubMed] [Google Scholar]
- 51.Siripanth C. Amphizoic amoebae: pathogenic free-living protozoa; review of the literature and review of cases in Thailand. J Med Assoc Thai. 2005;88: 701–707. [PubMed] [Google Scholar]
- 52.Phu NH, Mai NTH, Nghia HDT, Chau TTH, Loc PP, Thai IH, et al. Fatal consequences of freshwater pearl diving. The Lancet. 2013;381: 176. [DOI] [PubMed] [Google Scholar]
- 53.Xuan YH, Chung BS, Hong YC, Kong HH, Hahn TW, Chung DI. Keratitis by Acanthamoeba triangularis: Report of cases and characterization of isolates. Korean J Parasitol. 2008;46: 157–164. 10.3347/kjp.2008.46.3.157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kyle DE, Noblet GP. Vertical distribution of potentially pathogenic free-living amoebae in freshwater lakes. J Protozool. 1985;33: 422–434. [DOI] [PubMed] [Google Scholar]
- 55.Berete S, Combes A, de Jonkheere JF, Datry A, Varnous S, Martinez V, et al. Fatal disseminated Acanthamoeba lenticulata infection in a heart transplant patient. Emerg Infect Dis. 2007;13: 736–738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mohamed Kamel AG, Faridah H, Yusof S, Norazah A, Nakisah MA. A case of trauma related Acanthamoeba keratitis. Trop Biomed. 2004;21: 135–8. [PubMed] [Google Scholar]
- 57.Putri DE, Edwar L, Susiyanti M. Acanthamoeba keratitis in a non-contact lens wearer: A challenge in diagnosis and management. J Case Rep. 2014;4: 419–423. [Google Scholar]
- 58.Buerano CC, Trinidad AD, Fajardo LSN, Cua IY, Baclig MO, Natividad FF. Isolation of Acanthamoeba genotype T4 from a non-contact lens wearer from the Philippines. Trop Med Health. 2014;4: 145–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Abedkhojasteh H, Niyyati M, Rahimi F, Hei-Dari M, Farnia S, Rezaeian M. First report of Hartmannella keratitis in a cosmetic soft contact lens wearer in Iran. Iran J Parasitol. 2013;8: 481–485. [PMC free article] [PubMed] [Google Scholar]
- 60.Lorenzo-Morales J, Martinez-Carretero E, Batista N, Alvarez-Marin J, Bahaya Y, Walochnik J, et al. Early diagnosis of amoebic keratitis due to a mixed infection with Acanthamoeba and Hartmannella. Parasitol Res. 2007;102: 167–169. 10.1007/s00436-007-0754-x [DOI] [PubMed] [Google Scholar]
- 61.Kilvington S, Gray T, Dart J, Morlet N, Beeching JR, Frazer DG, et al. Acanthamoeba keratitis: The role of domestic tap water contamination in the United Kingdom. Invest Ophth Vis Sci. 2004;45: 165–169. [DOI] [PubMed] [Google Scholar]
- 62.Roslee NF, Abd Ghani MK, Nordin A, Suboh Y, Ab Rahim N. Pemencilan Acanthamoeba spp. daripada persekitaran akuatik. Jurnal Sains Kesihatan. 2010;8: 15–19 [Google Scholar]
- 63.Marciano-Cabral F. Biology of Naegleria spp. Microbiol Rev. 1988;52: 114–133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Szenasi Z, Endo T, Yagita K, Nagy E. Isolation, identification and increasing importance of free living amoebae causing human disease. J Med Microbiol. 1998;47: 5–16. 10.1099/00222615-47-1-5 [DOI] [PubMed] [Google Scholar]
- 65.Ma P, Visvesvara GS, Martinez AJ, Theodore FH, Daggett PM, Sawyer TK. Naegleria and Acanthamoeba infections. Rev Infect Dis. 1990;12: 490–512. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All relevant data are within the paper.