

Abstract
Advancing the production efficiency and profitability of aquaculture is dependent upon the ability to utilize a diverse array of genetic resources. The ultimate goals of aquaculture genomics, genetics and breeding research are to enhance aquaculture production efficiency, sustainability, product quality, and profitability in support of the commercial sector and for the benefit of consumers. In order to achieve these goals, it is important to understand the genomic structure and organization of aquaculture species, and their genomic and phenomic variations, as well as the genetic basis of traits and their interrelationships. In addition, it is also important to understand the mechanisms of regulation and evolutionary conservation at the levels of genome, transcriptome, proteome, epigenome, and systems biology. With genomic information and information between the genomes and phenomes, technologies for marker/causal mutation-assisted selection, genome selection, and genome editing can be developed for applications in aquaculture. A set of genomic tools and resources must be made available including reference genome sequences and their annotations (including coding and non-coding regulatory elements), genome-wide polymorphic markers, efficient genotyping platforms, high-density and high-resolution linkage maps, and transcriptome resources including non-coding transcripts. Genomic and genetic control of important performance and production traits, such as disease resistance, feed conversion efficiency, growth rate, processing yield, behaviour, reproductive characteristics, and tolerance to environmental stressors like low dissolved oxygen, high or low water temperature and salinity, must be understood. QTL need to be identified, validated across strains, lines and populations, and their mechanisms of control understood. Causal gene(s) need to be identified. Genetic and epigenetic regulation of important aquaculture traits need to be determined, and technologies for marker-assisted selection, causal gene/mutation-assisted selection, genome selection, and genome editing using CRISPR and other technologies must be developed, demonstrated with applicability, and application to aquaculture industries.
Major progress has been made in aquaculture genomics for dozens of fish and shellfish species including the development of genetic linkage maps, physical maps, microarrays, single nucleotide polymorphism (SNP) arrays, transcriptome databases and various stages of genome reference sequences. This paper provides a general review of the current status, challenges and future research needs of aquaculture genomics, genetics, and breeding, with a focus on major aquaculture species in the United States: catfish, rainbow trout, Atlantic salmon, tilapia, striped bass, oysters, and shrimp. While the overall research priorities and the practical goals are similar across various aquaculture species, the current status in each species should dictate the next priority areas within the species. This paper is an output of the USDA Workshop for Aquaculture Genomics, Genetics, and Breeding held in late March 2016 in Auburn, Alabama, with participants from all parts of the United States.
Keywords: Aquaculture, Genetic resources, Genome, Transcriptome, QTL, RNA-Seq, SNP, Fish, Shellfish
Background
The major goals of research programs having components related to aquaculture genomics, genetics and breeding are to enhance aquaculture production efficiency, sustainability, product quality and profitability in support of the commercial sector and for the benefit of U.S. consumers. Progress towards achieving these goals includes genetic improvement of production, performance and animal welfare/fitness traits, and this progress is predicated upon the access and utilization of an array of genetic resources within each species group. To this end, various genetic stock enhancement approaches are currently being studied by the aquaculture research community, and major progress has been made since the start of aquaculture genomics research 20 years ago [1]. Such progress includes advances in traditional selection, intraspecific crossbreeding, interspecific hybridization, genome-enabled selection (e.g., marker/causal mutation-assisted selection and/or genomic selection), polypoidy, sex reversal and breeding, xenogenesis, gene transfer, and genome editing. Some of the most important traits studied for genetic improvement in U.S. aquaculture species include disease resistance, feed conversion efficiency, growth rate, behaviour, processing yield, reproductive characteristics and tolerance to environmental stressors like low dissolved oxygen, high or low water temperature and salinity, body composition, and flesh quality. Traditionally, genetic improvement in the commercial aquaculture sector relied on phenotypes and pedigree information, but recently leading international breeding companies have begun to implement genome technologies into their breeding programs for some of the species where advanced genomic resources and tools are available (e.g., [2–4]).
Genomic information provides powerful tools to enhance physiological research, the results of which may be used for optimization of husbandry practices, feeding and feed formulations, breeding technologies, or non-genetic selection or screening (e.g., epigenetics, proteomics, and metabolomics). Whole genome sequences, in various states of assembly, are now available for many aquaculture species, enabling the identification of genomic variations such as insertions/deletions, single nucleotide polymorphisms (SNPs), copy number variations, and differentially methylated regions. However, this information is only useful when used to predict phenotypes that have a positive impact on production or product quality. For this reason, genetic mapping, quantitative trait loci (QTL) analysis, genome-wide association studies (GWAS), expression profiling, and bioinformatic analysis can be used to identify genotypic variants associated with particular phenotypic traits, which can then be exploited in breeding programs. For some aquaculture species we have reached the point where genome-based technologies such as marker-assisted and whole genome selection can be applied to enhance aquaculture traits and research is beginning to shift toward understanding functional polymorphisms and the gene regulatory networks underlying commercially important traits. A more complete understanding of the gene networks underlying growth, reproduction, and disease resistance will provide the knowledge-base for developing more robust and productive genetic stocks for the aquaculture industry.
The degree to which genome enabled technologies and genomic information have been or can be applied in genetic improvement programs varies across aquaculture species. Private sector investment in research and development for the implementation of new technologies is dependent on unique industry structure (e.g., overall size of the industry, size of individual companies) and the level of vertical integration. In addition, the approach used for germplasm improvement and status of existing breeding programs dictates whether and which genome enabled technologies are suitable for a given industry. Industries with centralized breeding, such as rainbow trout and salmon, have greater potential to benefit from new technologies compared to industries where breeding activities are widely distributed. Finally, the current demand for species-specific genomic tools (such as high through-put genotyping assays) among the diverse aquaculture industry sectors is low, rendering them commercially unaffordable. This forces some industries interested in genetic improvement to rely on the public sector for resources that enable application of state-of-the-art genomic technologies.
Here we review the development of genomic tools and application of genome enabled technologies for the genetic improvement of aquaculture species. Specifically, we review the status of genome mapping and sequencing, identify gaps in our current knowledge, and highlight the need to implement new technologies in aquaculture. We then propose a set of priorities for future research in aquaculture genomics, genetics, and breeding.
Whole genome sequencing and assembly
The genomes of several major aquaculture species in the United States, especially those under the USDA National Research Support Project 8 (NRSP-8), have been sequenced or are being sequenced (Table 1), including catfish [5], Atlantic salmon [6], rainbow trout [7], tilapia [8], striped bass (Reading, personal communication), Pacific oyster [9], eastern oyster (Gomez-Chiarri, personal communication), and Pacific white shrimp (Xiang, personal communication) as well as yellow perch and bluegill sunfish (Wang, personal communication). These accomplishments were achieved through support of USDA, NOAA, and other U.S. funding agencies. National Institute of Food and Agriculture (NIFA) AFRI programs, especially the Animal Genomics, Genetics and Breeding program, were central to the historical achievements of generating the reference genome sequences for these fish and shellfish species. Strong international collaboration was also important for the achievements. For instance, the genome project for the Pacific oyster was led by scientists from China and the U.S. [9]; the Atlantic salmon project was led by scientists from Norway, Canada, and Chile [6]; the rainbow trout project was led by scientists from France and currently is a collaborative international effort primarily between the U.S. and Norway; the genome project for Pacific white shrimp is led by Chinese scientists (Xiang, personal communication), and a reference genome for the original specific pathogen-free (SPF) broodstocks developed by the U.S. Marine Shrimp Farming Program in Oahu, HI is being generated (Alcivar-Warren, personal communication).
Table 1.
Some examples of whole genome sequencing of aquatic and aquaculture species
| Species | References |
|---|---|
| Ictalurus punctatus (Channel catfish) | Liu et al. 2016 [5] |
| Ictalurus furcatus (Blue catfish) | Waldbieser and Liu, unpublished data |
| Oncorhynchus mykiss (Rainbow trout) | Berthelot et al. 2014 [7] |
| Salmo salar (Atlantic salmon) | Lien et al. 2016 [6] |
| Oreochromis niloticus (Nile tilapia) | Brawand et al. 2014 [8] |
| Crassostrea virginica (Eastern oyster) | Gomez-Chiarri et al. 2015 [15] & personal communication |
| Crassostrea gigas (Pacific oyster) | Zhang et al. 2012 [9] |
| Penaeus/Litopenaeus vannamei (Pacific white shrimp) | Xiang, 2016, personal communication |
| Penaeus monodon (Giant tiger prawn) | Warren, personal communication |
| Atlantic Cod | Star et al. 2011 [142] |
| Bluegill sunfish | Wang, personal communication |
| California yellowtail | Severin, Purcell, Hyde, personal communication |
| Cavefish | McGaugh et al. 2014 [143] |
| Coelacanth | Amemiya et al. 2013 [144] |
| Common carp | Xu et al. 2014b [145] |
| Indian catfish | Das, personal communication |
| Japanse flounder | Chen, Yellow Sea Fisheries Institute, China, personal communication |
| Grass carp | Wang et al. 2015 [146] |
| Lamprey | Smith et al. 2013 [147] |
| Medaka | Kasahara et al. 2007 [148] |
| Pacific abalone | Severin, Purcell, Hyde, personal communication |
| Pearl oyster | Du, personal communication |
| Platyfish | Schartl et al. 2013 [149] |
| Rohu carp | Das, personal communication |
| Sea bass | Tine et al. 2014 [150] |
| Scallops | Bao, Ocean University of China, personal communication |
| Sea cucumber | Xiang, Chinese Academy of Sciences, China, personal communication |
| Shark | Venkatesh et al. 2014 [151] |
| Sole | Chen et al. 2014 [152] |
| Stickleback | Jones et al. 2012 [153] |
| Striped bass | Reading, 2016, personal communication |
| Tetraodon | Jaillon et al. 2004 [154] |
| Turbot | Figueras et al. 2016 [155] |
| White bass | Reading, 2016, personal communication |
| Yellow croaker | Wu et al. 2014 [156] |
| Yellow perch | Wang, personal communication |
| Zebrafish | Howe et al. 2013 [157] |
Bold data are the species initially included in the NRSP-8 Project (Alcivar-Warren et al. 1997)
Various technologies have been used for the generation of the whole genome sequence of aquaculture species. However, the Illumina and PacBio platforms have contributed the most to the progress of aquaculture genome sequencing. Illumina sequencing generates accurate, but short reads at a relatively low cost, while PacBio sequencing generates longer, but less accurate reads at a higher cost. The proportion of sequences generated using these two platforms varies depending on the species and the status of the sequencing technology when the genome sequencing projects were initiated. A significant decrease in the cost of PacBio sequencing has generally led to increased use of this technology and the enhancement of contig/scaffold lengths in sequence assemblies.
Like the selection of sequencing technologies, various sequencing templates were used for the generation of the whole genome sequence assemblies in aquaculture species. These included mixtures of outbred individuals, single diploid males or females, individuals from inbred lines, and completely homozygous doubled haploids. The use of more homozygous templates greatly simplifies the computation of genome assemblies, which are complicated by the high levels of heterozygosity and sequence polymorphism characteristic of several aquaculture species [6, 7, 10]. The choice of sequencing templates has largely been dictated by the availability of the preferred homozygous templates. For instance, doubled haploids, produced through gynogenesis or androgenesis, are the preferred sequencing template for most teleost sequencing projects. However, the generation of doubled haploids is not generally feasible in many shellfish species, due to the unequal first cleavage that is sensitive to manipulation [11]. For some species, multiple individuals must be used because the DNA extracted from a single individual is not sufficient for the sequencing process.
While the mechanics of generating a large number of sequence reads is no longer difficult, calculation of a high quality of sequence assembly remains a challenging task. Four specific metrics are generally used to evaluate the quality of whole genome sequence assemblies including 1) Contiguity, as reflected in contig numbers and distribution of contig sizes; 2) Connectivity, as reflected in the number of scaffolds and distribution of scaffold sizes; 3) Completeness, as reflected in the total size of the genome assemblies and the percentage of coverage of the whole genome; and 4) Accuracy, as validated by at least one additional methodology such as genetic linkage mapping, physical mapping, or optical mapping. In addition, the integration of the whole genome sequences with genetic linkage maps is important for genetic studies.
The quality of current whole genome sequences as measured by these four metrics, varies among species. Quality measurements of the whole genome sequence assemblies of the aquaculture species are summarized in Table 2. In general, sequence assemblies of fish species are of higher quality than those for shellfish species. This is in part because the genomes of the shellfish species are highly heterozygous and contain a high level of repetitive elements. For instance, the oysters are among the most polymorphic animals; SNP density was estimated at 1.22 SNPs per 100 bp for the Pacific oyster [9] and either 1.85 SNPs [12] or 4.2 SNPs per 100 bp [10] at population levels for the eastern oyster. Moreover, repetitive elements account for over 80% of the shrimp genome (Xiang, personal communications).
Table 2.
Status of whole genome sequencing and assembly of major aquaculture species in the United States, listed in the order of scaffold N50 sizes
| Species | Contig N50 [141] | Scaffold N50 (Mb) | Scaffolds | % on chromosome | Sequencing platform | Total size (Mb) | References |
|---|---|---|---|---|---|---|---|
| Catfish | 77.2 | 7.73 | 9974 | 97.2 | Illumina, PacBio | 783 | Liu et al. 2016 [5] |
| 99.1 | Zeng et al. 2017 [13] | ||||||
| Atlantic Salmon | 57.6 | 2.97 | 843,055 | 75.4 | Sanger, Illumina, PacBio | 2970 | Lien et al. 2016 [6] |
| Tilapia | 29.3 | 2.80 | - | 70.9 | Illumina, PacBio | 928 | Brawand et al. 2014 [8] |
| 3090 | - | 86.9 | 1010 | Conte et al. 2016, PC | |||
| Eastern oyster | 1.59 | 2.50 | 849 | In progress | PacBio, Illumina | 819 | Wes Warren, PC |
| Rainbow trout | 7.7 | 0.38 | 54.0 | Illumina | 1900 | Brawand et al. 2014 [8] Palti and Gao, PC | |
| 13.9 | 1.72 | 82.0 | 2178 | ||||
| Zebrafish | 25.0 | 1.55 | 96.5 | Sanger, Illumina | 1410 | Howe et al. 2013 [157] | |
| California yellowtail | 139.3 | 1.49 | 4439 | - | Illumina | 685 | Andrew Severin, PC |
| PacBio | |||||||
| Pacific white shrimp (Litopenaeus vannamei) | 57.1 | 0.66 | 6007 | 71.6 | Illumina | 1779 | Jianhai Xiang, 2016, PC |
| PacBio | |||||||
| Pacific oyster | 19.4 | 0.4 | 11,969 | - | Illumina | 559 | Zhang et al. 2012b [9] |
| Striped bass | 20.9 | 0.03 | 35,010 | - | Illumina | 585 | Benjamin Reading, 2016, PC |
| PacBio | |||||||
| White bass (male/female) | In process | In process | 56,818/57,533 | - | Illumina | 644/643 | Benjamin Reading, 2016, PC |
| Pacific abalone | In process | In process | - | - | Illumina | 2000 | Severin, Purcell, Hyde, PC |
| Yellow perch (male/female) | In process | In process | - | - | Illumina | 1380/1240 | Haping Wang, PC |
Zebrafish is included as a reference. PC: personal communications
For species under the NRSP-8 program, reference genome sequence assemblies for catfish, tilapia, Atlantic salmon, and rainbow trout are of good quality. For catfish, 50% of the genome sequence is included in only 31 of the largest scaffolds; 90, 95, and 98% of the genome is included in 185, 314, and 594 scaffolds, respectively. The catfish reference genome sequence was assessed to be nearly complete as 99.7% of re-sequencing reads were mapped to the reference genome sequence. In addition, the number of complete genes included in the reference genome sequence is larger than that of any of the sequenced diploid fish species, including zebrafish [5]. The catfish reference genome sequence assembly was validated by genetic mapping. The positions of 253,744 genetically mapped SNPs were fully concordant with those on the reference genome sequence with four exceptions [13]. The vast majority of the reference genome sequence (99.1%) has been anchored to chromosomes [13].
The reference genome assembly of Atlantic salmon is also of high quality [6]. The genome was sequenced with Sanger and Illumina technologies. It is complete as 2.97 Gb reference genome sequences were assembled, with the unassembled sequences being just repetitive elements. The largest 9447 scaffolds accounted for 2.24 Gb of the 2.97 Gb genome sequence. This is a remarkable achievement considering the very complex nature of the genome. The Atlantic salmon genome is largely tetraploid due to a recent genome duplication. It also has a high repeat content (58–60%); the dispersed Tc1 transposons represented 12.89% of the genome [6]. Similarly, the assembly of the rainbow trout genome is of good quality [7]. Since the publication of the genome paper, the reference genome sequence of rainbow trout has been further improved. The contig N50 has increased from 7.7 Kb to 13.9 Kb, and the scaffold N50 has increased from 380 Kb to 1.72 Mb. More importantly, over 82% of the genome sequence has been mapped to chromosomes (Palti, personal communication).
The published tilapia genome sequence [8] was already of good quality, but the recent use of PacBio long sequencing technology allowed a new high quality assembly (Matthew Conte, personal communication). The contig L50 length reached 3.09 Mb, and 50% of the genome is included in the largest 93 contigs. Importantly, over 86.9% of the reference genome sequence is anchored to chromosomes, enhancing the utility of the reference genome sequence for genetic analyses. The whole genome sequences of striped bass, white bass, yellow perch, and bluegill sunfish are at the stage of draft assemblies.
A published genome sequence exists for the Pacific oyster, but the assembly is highly fragmented [9]. Efforts are ongoing to improve the genome assembly and contiguity, completeness, and accuracy are significantly better now (Zhang, personal communication). Linkage analyses were conducted to validate the genome sequence assembly [14]. The whole genome assembly of eastern oysters is at the draft sequence stage. Several strategies were employed to address challenges encountered in the assembly of the Pacific oyster genome. A single, highly inbred individual, produced through multiple generations of inbreeding and one generation of meiotic gynogenesis (Guo, personal communication) was used as a template. PacBio sequencing was used to provide 50x genome coverage in addition to Illumina sequencing ([15]; Gomez-Chiarri, personal communication). Initial statistics suggest this assembly is of much higher quality than that of the Pacific oyster. The draft assembly (Table 2) is now being validated using high-density linkage maps generated by the Guo laboratory.
The Pacific shrimp genome has been sequenced and assembled, but is not yet published. As shown in Table 2, the genome assembly is of high quality, with a contig N50 of 57.1 kb. The whole genome is included in 6007 scaffolds. Importantly, 71.6% of the genome sequence is anchored to chromosomes through linkage mapping (Xiang, personal communications). Of all the aquaculture genomes, the shrimp genome is perhaps the hardest to deal with because of the difficulty in isolating high molecular weight DNA due to enhanced DNase activity, the large chromosome number, and high levels of heterozygosity and repetitive elements. Physical mapping has been hindered by the lack of BAC libraries with very large inserts. The only BAC library of shrimp, pECBAC1, has an average insert size of approximately 101 kb [16].
For XY heterogametic species, often only the homogametic gender was used as sequencing template, and so information on the sex chromosomes is lacking. For instance, the catfish genome sequence was produced using a doubled haploid female produced through gynogenesis, and therefore the Y sex chromosome was not sequenced. Similarly, the Atlantic salmon genome was produced by using DNA template from a single double-haploid female produced by mitotic androgenesis. Therefore, the Y chromosome is not included in the reference genome. The rainbow trout genome was sequenced using a YY doubled haploid. While it provided Y chromosome information, the X chromosome was not covered in the reference genome sequence. Furthermore, sex determination in some fish and shellfish is complicated by having multifactorial sex determining mechanisms, including genetic sex determination (GSD), environmental sex determination (ESD) and their interactions. With WZ heterogametic species like some of tilapia species, sequencing a single representative of each gender may not be sufficient if there is a polygenic sex determination.
The first genome sequence is a historical milestone for any aquaculture species. However, in order to enable the utility of a reference sequence, additional work is required. For all aquaculture species, further refinement of the reference genome sequence, including improvements in contiguity, completion, and accuracy, as well as anchoring the reference genome sequence to chromosomes and obtaining sex chromosome sequences, is a priority (Table 3). Integration of genome sequence and linkage maps is also very important for genetic and breeding work, and can be accomplished relatively quickly. Sequencing of the Y or X chromosome is essential to study sex determining mechanisms, and sex-related traits, such as sexual dimorphism in growth or sexual size dimorphism (SSD). For instance, with tilapia and bluegill, males grow much faster and bigger than females. In contrast, females grow faster and bigger with yellow perch (Hanping Wang, personal communication). Such differences can be exploited as excellent natural models for the analysis of the genomic basis for sexual bimorphisms.
Table 3.
Examples of additional work to enhance the utility of the whole genome reference sequences of major aquaculture species in the United States
| Species | Contiguity, completion, and accuracy | Anchoring sequence to chromosomes | Sex chromosome sequencing |
|---|---|---|---|
| Catfish | + | + | Y chromosome need to be sequenced |
| Atlantic salmon | ++ | ++ | Y chromosome need to be sequenced |
| Tilapia | + | + | |
| Rainbow trout | ++ | +++ | |
| California yellowtail | ++ | +++++ | |
| Pacific oyster | +++ | +++ | |
| Striped bass | ++++ | +++++ | |
| White bass | ++++ | +++++ | |
| Eastern oyster | +++ | ++++ | |
| Shrimp | +++ | +++ | |
| Pacific abalone | +++ | +++++ |
+ indicate some additional work required, and additional “+” signs indicate the level of additional work required; additional “+” signs indicate larger amount of improvements are needed
Genomic variations, polymorphic markers, and genotyping platforms
Catalogues of genome variations and efficient genotyping platforms are essential to fully exploit whole genome sequences. One of the most useful by-products of whole genome sequencing is the development of thousands of DNA markers. In the first decade of aquaculture genome research, major effort was focused on developing polymorphic markers [17]. As whole genome sequencing projects were conducted, large numbers of polymorphic markers were identified. Whole genome sequencing with diploid sequencing templates allows identification of both microsatellites and SNPs. Analysis of SNPs between the two alleles of the sequenced individual also allow a rough assessment of the level of heterozygosity of the species.
In addition to whole genome sequencing, SNPs can be identified through genome re-sequencing or RNA-Seq projects. For instance, genome re-sequencing projects have identified more than 8.3 and 9.7 million putative SNPs in channel catfish [18] and Atlantic salmon [19] respectively. Large numbers of SNPs have been identified in most major aquaculture species, with those for the species under the NRSP-8 summarized in Table 4. SNP markers are a much-needed resource for genetic and genomic studies, the construction of high-density SNP arrays, and the development of high-density linkage maps. Validation and testing of these SNPs using SNP arrays will form the material basis for GWAS and whole genome-based selection.
Table 4.
Some examples of SNPs identified from the aquaculture species under NRSP-8
| Species | SNPs from genome sequencing | Numbers of SNPs | Method of identification | Reference |
|---|---|---|---|---|
| Catfish | None | 8.3 million | Genome re-sequencing, transcriptome sequencing | Sun et al. 2014 [18] |
| Liu et al. 2012 [158] | ||||
| Rainbow trout | None | 145,168 | RAD sequencing | Palti et al. 2014 [159] |
| 5052 | RNA-Seq | Christensen et al. 2013 [160], Al-Tobasei et al. 2016 [161] | ||
| 50,000 | RNA-Seq | Palti et al. 2015 [23] | ||
| 1.8 million | Genome re-sequencing | |||
| Atlantic salmon | None | 9.7 million | Genome re-sequencing | Yáñez et al. 2016 [19] |
| Tilapia | Yes | 3569 | Genome re-sequencing | Van Bers et al. 2012 [162] |
| Striped bass | Yes | - | RNA-Seq | Li et al. 2014 [163] |
| Pacific oyster | Yes | 3.8 million | Genome re-sequencing | Zhang et al., 2012 [9] |
| 4122 | RNA-Seq | Hedgecock et al. 2015 [14] | ||
| Pacific white shrimp | Yes | 96,040 | RNA-Seq | Yu et al. 2014 [164] |
Those SNPs identified from genome sequencing are not included here
A key advantage of SNP over microsatellite markers is the potential for rapid, low-cost genotyping. For many aquaculture species, the identification of large numbers of SNPs led to the development of efficient genotyping platforms. Available high-density SNP arrays for aquaculture species are listed in Table 5 and include the 15, 286, and 930 K Atlantic salmon arrays [6, 20, 21], the 250 and 690 K catfish arrays [13, 22], the 57 K rainbow trout array [23], and the 250 K common carp array [24]. The SNP arrays for each of the four aforementioned species have high marker densities and good genome coverage. SNP arrays need to be developed for tilapia, striped bass, oysters, and shrimp. As with genome assembly, the development of SNP arrays for some species (e.g., oysters and shrimp) is complicated by extremely high levels of polymorphism.
Table 5.
Development of high density SNP arrays in aquaculture species, PC: personal communications
| Species | SNP array technology | SNP array density | References |
|---|---|---|---|
| Atlantic salmon | Illumina iSelect technology | 15 K | Gidskehaug et al. 2011 [20] |
| Affymetrix Axiom technology | 286 K | Houston et al. 2014 [21] | |
| Affymetrix Axiom technology | 930 K | Lien et al. 2016 [6] | |
| Catfish | Affymetrix Axiom technology | 250 K | Liu et al. 2014 [22] |
| Affymetrix Axiom technology | 690 K | Zeng et al. 2017 [13] | |
| Common carp | Affymetrix Axiom technology | 250 K | Xu et al. 2014 [147] |
| Rainbow trout | Affymetrix Axiom technology | 57 K | Palti et al. 2015 [23] |
| Affymetrix Axiom technology | 50 K | Salem et al. PC |
Linkage mapping and physical mapping
Ultimately, genomic information must be translated into genetic terms to facilitate genetic enhancement in aquaculture. Genetic linkage maps derived from genetic analysis of recombination during meiosis are important for the assembly of chromosome-scale sequence scaffolds. Mapping of sequence-tagged genetic markers derived from the reference genome allows sequence contigs to be arranged in an order that corresponds to the linkage group or chromosome. In addition, linkage mapping is a good method for validating reference genome assemblies.
Linkage maps have been constructed for most of the major aquaculture species (Table 6). For the species under NRSP-8, high density maps exist for catfish, Atlantic salmon, rainbow trout, tilapia, oysters, and shrimp. The linkage maps for catfish and salmonids have the highest marker densities, with the latest catfish linkage map ordering 253,087 markers [13], and the Atlantic salmon linkage map ordering 565,887 markers [6]. The latest linkage maps for the Pacific and eastern oysters have 3367 and 4316 markers, respectively [25] (Guo, personal communication). A large proportion of the genome sequence has been anchored to linkage maps in catfish (99.1%), tilapia (86.9%), Atlantic salmon (75.4%), Pacific shrimp (71.6%), and rainbow trout (54%).
Table 6.
Examples of genetic linkage maps in aquaculture species, with the species under the NRSP-8 in bold
| Species | Number and type of markers | Mapping population | Unique map positions | References |
|---|---|---|---|---|
| Asian seabass | 790 microsatellites and SNPs | 93 fish from two families | 501 | Wang et al. 2011 [165] |
| Atlantic salmon | 5650 SNPs | 3297 fish from 143 families | 2894 in female genetic map, 1009 in male specific map | Lien et al. 2011 [166] |
| Brown trout | 288 microsatellites, 13 allozymes | 93 fish from 4 families | - | Gharbi et al. 2006 [167] |
| Catfish | 54,342 SNPs | 576 fish from three channel catfish families | 15,598 | Li et al. 2015 [168] |
| 26,239 SNPs | 288 interspecific backcross progenies | 12,776 | Liu et al. 2016 [169] | |
| 253,087 SNPs | 465 fish from four channel catfish families | 30,591 | Zeng et al. 2017 [13] | |
| Common carp | 28,194 SNPs | 108 fish from one yellow river carp family | 14,146 | Peng et al. 2016 [170] |
| Eastern oyster | 4607 SNPs | 112 progenies from one family | 4136 | Guo, personal communication |
| European seabass | 190 microsatellites, 176 AFLP, 2 SNP | 50 fish from one Venezia Fbis family | - | Chistiakov et al. 2008 [171] |
| Grass carp | 279 microsatellites and SNPs | 192 progenies from two families | 245 | Xia et al. 2010 [172] |
| Japanese flounder | 1268 microsatellites, 105 SNPs, 2 genes | 45 offspring from one family | 235 in male genetic map, 184 in female genetic map | Castaño-Sánchez et al. 2010 [173] |
| Pacific oyster | 1172 SNPs and microsatellites | 336 progenies from five families | 1172 unique markers mapped | Hedgecock et al. 2015 [14] |
| 424 in consensus linkage map | ||||
| Rainbow trout | 2226 microsatellites and SNPs | 120 individuals from two unrelated doubled haploid lines | 1366 in synthetic map | Guyomard et al. 2012 [174] |
| 47,939 SNPs | 5716 fish | 47,939 mapped to genome sequence scaffolds | Gonzalez-Pena et al. 2016 [26]; Palti personal communication | |
| Scallop | 3806 SNPs | 96 progenies from one Farrer’s scallop family | 2983 | Jiao et al. 2013 [175] |
| Sea bream | 321 microsatellites, ESTs, and SNPs | 50 individuals from one family | 229 | Tsigenopoulos et al. 2014 [176] |
| Giant tiger prawn | 3959 SNPs | 1024 offspring from seven black tiger shrimp family | - | Baranski et al. 2014 [177] |
| Pacific white shrimp | 429 AFLP, 22 microsatellites | F2 cross of slow and fast growth parents, 43 shrimp | - | Andriantahina et al. 2013 [178] |
| 6146 SNPs | 205 progenies from one Pacific white shrimp family | 4650 | Yu et al. 2015 [179] | |
| Tilapia | 525 microsatellites, 20 genes | 70 individuals from one family | 435 | Lee et al. 2005 [180] |
| 401 microsatellites | 95 individuals from two families | 352 | Liu et al. 2013 [181] | |
| Yellowtail | 217 microsatellites | 90 progenies from one family | 105 in female genetic map, 83 in male genetic map | Ohara et al. 2005 [182] |
| 1480 microsatellites and 601 SNPs | 94 offspring of one family | - | Aoki et al. 2015 [183] | |
| 6275 SNPs | 460 individuals from five wild families | - | Ozaki et al. 2016 [184] |
The major issue for linkage maps of aquaculture species is resolution. While the number of markers on the high density SNP arrays is large, map resolution has been limited by the size of the mapping populations. In most cases, the number of samples used for genetic mapping was not very large, leading to a high level of marker stacking. The exceptions are the Atlantic salmon and rainbow trout where over 2000 and 5000 individuals, respectively, were used for linkage analysis, leading to a very high resolution of the linkage map [6, 26]. While the high fecundity of fish and shellfish species makes it possible to generate large mapping families, the major limitation for high resolution linkage mapping is funding, as genotyping costs are directly proportional to the sample sizes in linkage analysis.
Physical maps have been constructed for only a few aquaculture species (Table 7) including Atlantic salmon [27], tilapia [28], catfish [29, 30], rainbow trout [31], common carp [32], Asian seabass [33], Pacific oyster [34] and scallop [35]. Over time, BAC-based physical mapping has been replaced in favour of next generation sequencing and optical mapping technologies [36]. The existing physical maps and related BAC resources, however, are still useful for validation of reference genome sequences.
Table 7.
Examples of physical maps constructed from aquaculture species
| Species with physical maps | References |
|---|---|
| Atlantic salmon | Ng et al. 2005 [27] |
| Tilapia | Katagiri et al. 2005 [28] |
| Channel catfish | Xu et al. 2007 [30] |
| Quiniou et al. 2007 [29] | |
| Pacific oyster | Gaffney, 2008 [34] |
| Rainbow trout | Palti et al. 2009 [31] |
| Common carp | Xu et al. 2011 [32] |
| Pacific white shrimp | Yu et al. 2015 [179] |
| Asian seabass | Xia et al. 2010 [33] |
| Scallop | Zhang et al. 2011 [35] |
Transcriptome resources
Proper annotation of the genome sequences presents a challenge that can be at least partially overcome with transcriptome information. Specifically, gene models and gene structures need to be supported by experimental data; exon-intron borders need to be defined; alternatively spliced and differentially polyadenylated transcripts need to be identified and their translated proteins verified; and expression and function of the genes need to be studied. In addition to protein-coding genes, non-coding RNAs need to be identified and mechanisms of their target interactions need to be understood.
Large numbers of expressed sequence tag (EST) resources exist for major aquaculture species. As summarized in Table 8, almost a half million ESTs were generated for Atlantic salmon, over 350,000 for channel catfish, and almost 290,000 for rainbow trout. These EST resources are useful for the assembly of full length transcripts for genome annotation; however, with the advent of low-cost next generation sequencing technologies, transcriptomes are now more efficiently characterized with RNA-Seq.
Table 8.
EST resources of selected aquaculture species (with >10,000 ESTs)
| Species | Number of ESTs |
|---|---|
| Danio rerio (zebrafish) | 1,488,275 |
| Ciona intestinalis | 1,205,674 |
| Xenopus laevis (African clawed frog) | 677,911 |
| Oryzias latipes (Japanese medaka) | 666,891 |
| Salmo salar (Atlantic salmon) | 498,245 |
| Ictalurus punctatus (channel catfish) | 354,516 |
| Oncorhynchus mykiss (rainbow trout) | 287,564 |
| Morone saxatilis (striped bass) | 230,151 |
| Crassostrea gigas | 206,388 |
| Litopenaeus vannamei | 161,248 |
| Ictalurus furcatus | 139,475 |
| Oreochromis niloticus (Nile tilapia) | 120,991 |
| Petromyzon marinus (sea lamprey) | 120,731 |
| Sparus aurata | 79,216 |
Zebrafish is included as a reference
Large RNA-Seq datasets have been generated by various institutions for important aquaculture species in the United States (https://www.ncbi.nlm.nih.gov/sra) to characterize differentially expressed genes in response to disease or stress in catfish [37–40], disease in salmon [41, 42] and to identify markers associated with growth, heat stress, and disease and tissue specificity in rainbow trout [43–46]. In striped bass, RNA-Seq studies focused on reproduction traits and egg quality [47–49], while in tilapia, they were conducted to identify genes responsive to alkalinity stress [50], salinity adaptation [51], and adaptation to low or high fat diets [52]. In yellow perch and bluegill, RNA sequencing of neo-males (perch), neo-females (bluegill), regular males and regular females is being conducted to investigate epigenomic modification of SSD and sex determination in fish (Wang, personal communication). RNA-Seq studies have also been conducted to characterize the Pacific oyster response to environmental stress (e.g., temperature, salinity, air exposure and heavy metals) [9, 53–55] and Ostreid herpesvirus [52]. In eastern oysters, RNA-Seq studies identified genes associated with osmoregulation [12], characterized the transcriptomic response to a bacterial pathogen [56], and revealed extensive expansion of gene families associated with innate immunity [15, 57]. In shrimp, genes associated with early development [58] and resistance to Taura syndrome virus (TSV) [59] have been identified via RNA-Seq analysis, and improved shrimp transcriptome were reported [60]. When coupled with genetic analysis such as bulk segregant analysis (e.g., [38, 43]), transcriptome analyses using RNA-Seq will enable the identification of candidate genes for important aquaculture traits.
Transcriptome resources also empower proteomics analysis [48, 61, 62]. Proteomics offers great promise for advancing our understanding of the functions of genes that underlie important production traits, however these methods rely on existing homologous protein-coding sequence databases, which remain incomplete for many non-model organisms, including important aquaculture species. Tandem mass spectrometry approaches in proteomics use these databases to identify protein fragments by mass spectrometry and thus require amino acid (or protein-coding nucleic acid) sequence information, optimally from the research organism under investigation. Thousands of different proteins have already been identified and measured with tandem mass spectrometry approaches to answer important questions about reproduction in striped bass and the closely related white perch, which serves as a research model [61–64]. A similar proteomic approach identified important proteins related to muscle atrophy in rainbow trout [65].
Non-coding transcripts, regulation of genome expression, and epigenomics
Despite their importance in regulating gene expression, non-coding transcripts are much less understood than protein-coding transcripts in aquaculture species. Limited work has been conducted in this relatively new area of research. Among aquaculture species, most of the work on non-coding RNAs was conducted in rainbow trout. A few studies were devoted to identification of microRNAs and long non-coding RNAs [66–71]. In a number of cases, microRNAs were found to be associated with performance traits. For instance, a large number of microRNAs were differentially expressed between sexually mature and immature fish; in association with egg quality and muscle growth and quality [72–74]. In addition, differential expression of long non-coding RNAs studied in three genetic lines of rainbow trout identified important long-coding RNAs in response to infection with Flavobacterium psychrophilum [75].
In Atlantic salmon, several studies were conducted to characterize the microRNA repertoire. In one study, Bekaert et al. [76] identified 888 microRNA genes. In another study, Andreassen et al. [77] identified a total 180 distinct mature microRNAs, and found that many microRNAs were conserved across species, and a few microRNAs were expressed in a tissue-specific fashion. In another study, Kure et al. [78] found that 18 microRNAs were differentially expressed upon exposure to acidic aluminium-rich water.
Research on non-coding RNAs in catfish, striped bass, tilapia, oysters, and shrimp is limited. For instance, residue microRNA profiling was reported in catfish [79–81], tilapia [82], oysters [83, 84], and shrimp [85, 86]. However, now with the high quality reference genome sequences, it is expected that large numbers of projects will be conducted with aquaculture species in this area. This aligns very well with the FAANG (Functional Annotation of Animal Genomes) Project. As the importance and detailed operational protocols are well discussed in the white paper published in Genome Biology [87], we will not repeat them here, but this will be an important area for future research with aquaculture species as well, especially those with a well assembled reference genome sequence.
Genome scale analysis of epigenetic regulation have been conducted with oysters [88–93], Atlantic salmon [94], rainbow trout [95, 96], and tilapia [97], yellow perch, bluegill (Wang, personal communication) and additional projects are being initiated in several other major aquaculture species. Apparently, this is an area of active research, and functional annotation of non-protein coding genome elements is an important area. Again, this aligns well with those objectives of the FAANG Project [87].
Performance traits, phenotypic variations, and QTL analysis
The practical purpose of aquaculture genomics and genetics studies is to reveal the genetic basis of performance and production traits, and to use such information for genetic enhancement programs. Domestication of most aquaculture species is still in the early stages, occurring over the last few decades, compared to other food animals and crops which have been domesticated over hundreds or even thousands of years. Because of this short history of domestication, aquaculture species still segregate considerable genetic variation among strains, lines, families and individuals.
Many aquaculture phenotypes are complex and quantitative in nature. Therefore, a major goal of aquaculture genetics research is to leverage genome information to predict complex phenotypes. In aquaculture species, QTL mapping and GWAS analysis are well-established procedures for correlating genetic and phenotypic variation; however additional work is required to identify specific genetic variants responsible for phenotypic variations. The identification of the causal SNPs or the genes underlining the performance traits is not only important for aquaculture applications, but also important for understanding the molecular mechanisms of phenotypic expression.
Progress with QTL/GWAS analysis has been greatly accelerated by the application of SNP arrays. Some examples of QTL mapping and GWAS analysis in aquaculture species are listed in Table 9. Most of the work has focused on disease resistance, growth traits, tolerance to stresses, and development or sexual maturity. Some of the best examples of QTL studies are from salmon research. For instance, the resistance against infectious pancreatic necrosis (IPN) virus was mapped to a major QTL that account for vast majority of phenotypic variance [98, 99], and further analysis identified the causal gene as epithelial cadherin [100, 101]. In catfish, QTL have been identified for a number of traits including disease resistance [38, 102, 103], heat stress [104], hypoxia tolerance [105], and head size [106]. In most of these cases, QTL were mapped within a region smaller than one million base pairs, allowing speculation of candidate genes, but fine mapping will be required to identify the specific causal genes. An interesting finding of these studies is the identification of functional hubs [102, 106] linking genes with roles in the same pathway. In addition, there appears to be a high level of evolutionary conservation of genes responsible for a number of traits in various species ranging across mammals, amphibians, and fishes. For instance, genes involved in the small GTPase pathway were found to affect head size and shape in catfish, frogs, mouse, and dogs [107]. Such discoveries open the possibility of comparative quantitative genomics.
Table 9.
QTL studies in selected aquaculture species with major US aquaculture species in bold
| Species | Traits | Reference |
|---|---|---|
| Arctic charr | Body weight and sexual maturation; Salinity tolerance | Küttner et al. 2011 [185] |
| Norman et al. 2011 [186] | ||
| Asian seabass | Resistance against viral nervous necrosis disease | Liu et al. 2016 [187] |
| Growth-related traits | Wang et al. 2006 [188] | |
| Omega-3 fatty acids | Xia et al. 2014 [189] | |
| Atlantic salmon | Growth traits and flesh colour | Baranski et al. 2010 [190]; Tsai et al. 2014 [191]; 2015 [192]; Moen et al. 2009 [99]; 2015 |
| Resistance against IPN | [101]; Houston et al. 2008 [98]; 2010 [100] | |
| Late sexual maturation | Gutierrez et al. 2014 [193] | |
| Resistance to pancreas disease | Gonen et al. 2015 [194] | |
| Catfish | Columnaris disease resistance | Geng et al. 2015 [102] |
| ESC disease resistance | Wang et al. 2013 [38]; Zhou et al. 2017 [103] | |
| Hypoxia tolerance | Wang et al. 2016 [105]; | |
| Heat stress | Jin et al. 2016 [104] | |
| Head size | Geng et al. 2016 [106] | |
| Common carp | Muscle fiber traits | Zhang et al. 2011 [195] |
| Morphometric traits | Boulton et al. 2011 [196] | |
| Swimming ability | Laghari et al. 2014 [197] | |
| Eastern oyster | Disease resistance | Yu and Guo, 2006 [110] |
| European seabass | Growth, body weight | Louro et al. 2016 [198], |
| Morphometric traits and stress response | Massault et al. 2010 [199] | |
| Pacific white shrimp | Growth parameters | Andriantahina et al. 2013 [178] |
| Giant tiger prawn | Disease resistance and sex determination | Robinson et al. 2014 [200] |
| Japanese flounder | Vibrio anguillarum resistance | Wang et al. 2014 [201] |
| Pacific oyster | Growth | Guo et al. 2012 [112] |
| Resistance against summer mortality | Sauvage et al. 2010 [202] | |
| Viability | Plough & Hedgecock, 2011 [111]; Plough et al. 2016 [113] | |
| Gilthead seabream | Skeletal deformities | Negrín-Báez et al. 2015 [203] |
| Sex determination and body growth | Loukovitis et al. 2011 [204] | |
| Resistance to fish pasteurellosis | Massault et al. 2011 [205] | |
| Rainbow trout | Growth related traits | Kocmarek et al. 2015 [206]; Wringe at al., 2010 [207]; Leder at al., 2006 [208]; Easton et al. 2011 [209]; Miller et al. 2012 [210] |
| Spawning time; development rate | ||
| Upper thermal tolerance | Perry et al. 2005 [211] | |
| Whirling disease resistance | Baerwald et al. 2011 [212] | |
| Bacterial cold water disease resistance | Vallejo et al. 2014 [107]; Palti et al. 2015 [108]; Liu et al. 2015 [109]; Campbell et al. 2014 [213] | |
| IHNV disease resistance | Rodriguez et al. 2004 [214]; Campbell et al. 2014 [213] | |
| Fillet yield | Gonzalez-Pena et al. 2016 [26] | |
| Osmoregulation capacity | Le Bras et al. 2011 [215] | |
| Response to crowding stress | Rexroad et al. 2013 [216]; Liu et al. 2015 [217] | |
| Turbot | Growth traits | Sánchez-Molano et al. 2011 [218] |
| Aeromonas resistance | Rodríguez-Ramilo et al. 2011 [219] | |
| Resistance against Philasterides | Rodríguez‐Ramilo et al. 2013 [220] | |
| Resistance to viral haemorrhagic septicaemia | Rodríguez-Ramilo et al. 2014 [221] | |
| Tilapia | Growth traits | Liu et al. 2014 [222]; Wang et al. 2015 [223] |
| Sex | Palaiokostas et al. 2015 [224] |
Similarly, QTL have been identified for growth and reproductive traits, upper thermal tolerance, osmoregulation capacity, stress responses, and disease resistance in rainbow trout. Significant efforts have been devoted to the analysis of resistance to bacterial cold water disease (BCWD) [107–109]. QTL analysis and genome selection for BCWD resistance are facilitating significant genetic improvement for this trait in rainbow trout [3, 4].
Although QTL have been identified for disease resistance, viability and growth-related traits in eastern and Pacific oysters [110–113], low marker density limits QTL resolution. Candidate gene-based studies have led to the identification of variation in a serine protease inhibitor associated with Perkinsus marinus-resistance in the eastern oyster [114]. QTL analysis in tilapia, striped bass, and shrimp are at the early stages, but with the efficient genotyping systems, rapid progress is expected.
Aside from lack of genetic and genomic resources (e.g., inbred lines/families, sequenced genomes, efficient genotyping platforms) in some aquaculture species, several additional challenges face aquaculture researchers. First, unlike many livestock species where phenotypic and genotypic data can be collected on a large proportion of the cultured animals, phenotypic and genotypic data collection on the entire population of an aquaculture species is impossible. It is therefore essential that aquaculture geneticists understand QTL in all strains used in the industry, because a QTL present in one population may not be present in another. Second, fish and shellfish are outbred species with extremely large numbers of founders. Their high fecundities make QTL analysis within families extremely efficient, but whether the identified QTL are conserved across families, strains, and populations are unknown.
Genome-based technologies and regulatory framework
A number of technologies, including polyploidization, gynogenesis, androgenesis, sex reversal, gamete cryopreservation, and gene transfer, are still very useful for aquaculture breeding programs. There are opportunities for enhancing these technologies by using genomic information. At the same time, genomic research has generated new technologies that can be used for genetic enhancement of aquaculture species, including marker-assisted selection (MAS), genome selection (GS), and genome editing.
Marker-assisted selection has been successfully used in aquaculture. The best example of MAS in an aquaculture species is selection for disease resistance in Japanese flounder. A microsatellite locus, Poli9-8TUF, was mapped near the major QTL for resistance to lymphocystis disease. Additional analysis indicated that the disease resistance was controlled by a single gene, and that the resistance allele was dominant. Based on the marker linkage information, Fuji et al. [115] developed a new population of Japanese flounder using MAS with the marker Poli9-8TUF. They selected a female homozygous for the favourable allele (B-favourable) and a male with a higher growth rate and good body shape, but without the resistant allele as parents. All the progeny are heterozygotes with the resistance allele and entirely resistant to lymphocystis disease, while the control group without B-favourable alleles showed incidences of 4.5 and 6.3% of mortality due to lymphocystis disease. These results clearly demonstrate that MAS is an efficient strategy for breeding [116].
Another good example of MAS is the selection of IPN resistance in Atlantic salmon. One major QTL was mapped to linkage group 21, which accounts for 29% and 83% of the phenotypic and genetic variances, respectively. Three microsatellite markers were tightly linked to the QTL, and these markers have been used for the selection of IPN resistance [99]. Recently, the gene responsible for IPN resistance was identified as a cadherin expressed in the epithelium where the protein binds to IPNV virions [101]. Marker-assisted selection allowed production of IPN-resistant salmon, leading to a 75% reduction in the number of IPN outbreaks in the salmon farming industry [101].
Sex identification using sex markers is a special case of MAS. Sex markers have been developed and used in quite a few aquaculture species, including common carp [117], tilapia [118], catfish [119], zhikong scallop [120], half-smooth tongue sole [121], white shrimp [122], kuruma prawn [123], yellowtail [124] and rainbow trout [125]. These sex-linked markers have been useful for the identification of sex without phenotypic data.
Recent advances in genome analysis including the availability of a large number of polymorphic markers, highly efficient genotyping platforms such as SNP arrays, and the application of next generation sequencing technologies, allowed mapping of dense markers across the entire genome, which in turn enables an estimation of the genetic merit of every chromosome fragment contributing variation in a population with phenotypic observations. Not only can the merit of every chromosomal segment be estimated, but also all the traits of interest can be estimated simultaneously. Whole genome selection is based on estimating the value of every chromosomal fragment contributing variation in a population with phenotypic observations (Training), and then the results of training are used to predict the merit of new animals (Testing) that are not included in the training dataset.
Genome selection was first proposed by Meuwissen et al. [126]. Since then it has gained tremendous attention in the animal genetics community. Compared with MAS, genomic selection uses the estimated effect of many loci across the entire genome at once, not just the small number of linked loci as done with MAS. Although genome selection has been successfully used in dairy cow and beef cattle and other livestock species [127], its use in aquaculture species has been limited to just a few species [128, 129]. In rainbow trout, genome selection was carried out for the selection of bacterial cold water disease [3]. In Atlantic salmon, genome selection was used to predict breeding values for resistance to sea lice [130]. Although demonstrated to be effective, genome selection has not been commercially applied in aquaculture species primarily due to financial limitations.
Supervised machine learning is similar in concept to whole genome selection using Training and Testing datasets, and includes Support Vector Machines (SVMs) and Artificial Neural Networks (ANNs). These are systems that can be trained to recognize certain data input patterns and then can be used to predict outcomes or classify data. Machine learning has been used to classify transcriptome and proteome data by pattern recognition (expression “fingerprinting”) in an analytical bioinformatics approach [49]. Expression patterns of genes and proteins can be modelled to identify the most important ones contributing to a trait or response. Machine learning ANNs have been used to analyze tens of thousands of expressed genes in microarray and RNA-Seq studies to show that the collective changes in the expression of 233 ovary genes (less than 2% of the genes measured) explained over 90% of the variation in striped bass embryo survival [47, 49]. These trained ANNs also predict, with a correct classification rate over 80%, which female striped bass will produce fertile or infertile eggs based on gene expression profiles of ovary tissues sampled prior to ovulation. Additionally, SVMs have been used to model the striped bass ovary proteome (355 proteins) and this system can predict the specific ovary growth stage with 83% accuracy based on quantitative tandem mass spectrometry data [61]. A portion of the plasma proteome (94 proteins) also has been similarly modelled to accurately predict gender of white perch [131]. Therefore, machine learning additionally poses a potential use as a diagnostic tool, for example in identifying those females that will produce poor quality eggs, or determining reproductive state or gender. Future applications of machine learning could include modelling genomic markers, such as SNPs, to identify those most important to a particular trait and then to predict the future performance of an individual based on the presence or absence of those SNP markers.
Genome editing refers to the ability to make specific changes at targeted genomic sites [132]. With the initial zinc finger nuclease (ZFN) technology developed in 1996, genome editing technologies have evolved and become more and more efficient, with the development of TALEN (transcription activator-like effector nucleases) and CRISPR/Cas9 (clustered regulatory interspaced short palindromic repeats). These new genome editing technologies overcome the disadvantages of ZFN technology and they have become very efficient for the modification of genomes. CRISPR/Cas9 has been demonstrated to be very efficient in zebrafish [133, 134], tilapia [135] and catfish (Liu, unpublished data). Mutation rates of 70–100% can be achieved with very low levels of mosacism in channel catfish [136].
Genome editing technologies can be used to introduce an immediate improvement in a phenotype in a single generation; hence, these technologies hold great promise for improving aquaculture. However, genetically modified organisms (GMO) have encountered low public acceptance, especially with aquaculture species. As demonstrated with the lengthy approval process of AquAdvantage transgenic Atlantic salmon, decades of time and millions of dollars were spent in coping with the regulatory issues ([137]; Hackett, 2016, personal communications). It could be argued that genome editing technologies differ from traditional gene transfer technologies because no foreign DNA is introduced. The scientific community must be proactive of research in the area of regulatory issues and public perception. Escaping the GMO label is possible with genome editing. For example, recently, the USDA decided that a CRISPR-modified mushroom can be cultured and sold without passing through the agency’s regulatory process [138].
Leveraging Investments in Genomics through Integration with Germplasm Repositories
Rapid development and adaptation of genomic tools among various aquatic species is a double-edged sword in terms of how such tools may create genetic diversity and thereby limit industry options in the future. Evidence of such contractions have been demonstrated with livestock and in particular the Holstein cow and how gene banks can facilitate the alleviation of genetic bottlenecks [139, 140]. While genomic research continues to rapidly proceed among various aquaculture species, there are some major technological gaps preventing the aquaculture sector from securing and utilizing improved genetic resources. As shown in other life forms such as livestock species, there is a critical need to understand and acquire genetically diverse samples from all major aquatic species, cryopreserve those samples, and to present them in publically available databases for viewing of information about the sampled populations. Such information would include phenotypes, management system descriptors, environmental conditions, locality data, and comprehensive genomic information. The Animal-GRIN information system operated by USDA/ARS is designed in this manner and is publically accessible via the internet. Acquiring and integrating this wide range of data not only serves to make germplasm and tissue samples more useful in the present, it will also allow researchers to perform studies not foreseen today and to respond to future challenges such as disease outbreaks or losses of critical genetic diversity in cultured lines. Viewing of genetic resources (via germplasm) and its associated detailed information as a public resource serves to speed innovation, as well as to leverage the considerable investments being made in genomic research. In essence this affords us new and more cost-effective approaches to produce, maintain, and distribute genetic improvement across the breadth of cultured aquatic species [141].
To respond to these needs, there is a requirement to collect and cryogenically store gametes and tissues from a wide range of species that can be used by industry members and public researchers alike. Coupled with these samples should be the ability to store genomic information from publicly funded research, as well as from industry. Such an information system would link samples with genomic, phenotypic, locality (GIS-based), and environmental descriptors and make this information publically available through a user interface via the internet. Aquatic species researchers could use this resource for varied experimental purposes (e.g., of crossing spring and fall spawning populations) and for corrective mating. As such, the collection and curation of germplasm or tissue samples has value, just as does the determination of genomic information. It is the purposeful integration of these genetic and informational resources that provides a synergistic leveraging or expansion of value and potential utility. Indeed, the value of information or germplasm samples is directly magnified by their coupling or association in a comprehensive repository system.
Future research priorities
Economically important aquaculture species are a diverse group of organisms and research priorities vary depending on the unique biology of each species. Although fishes are the most diverse vertebrate group, aquacultured teleosts are similar enough phylogenetically and biologically that they can follow a similar research program. Invertebrates are not as uniform, and may each have special properties that require different approaches.
The research tasks needed to develop a program of genetic enhancement in aquaculture species can be divided into two phases (Fig. 1). The first phase, development of species-specific genomic resources, is the one that has been pursued for the major aquaculture species over the past 20 years. It includes the development of genetic and physical maps, annotated genome sequences, and platforms for high-throughput genotyping. Some species (e.g., catfish, tilapia, rainbow trout, and salmon) may be nearing the completion of Phase I. Other species are just beginning this phase, hopefully benefitting from the experience of other species, and taking shortcuts available with new technologies. From the perspective of genetics and breeding, generation of complete sets of heritabilities and genetic correlations is also needed.
Fig. 1.
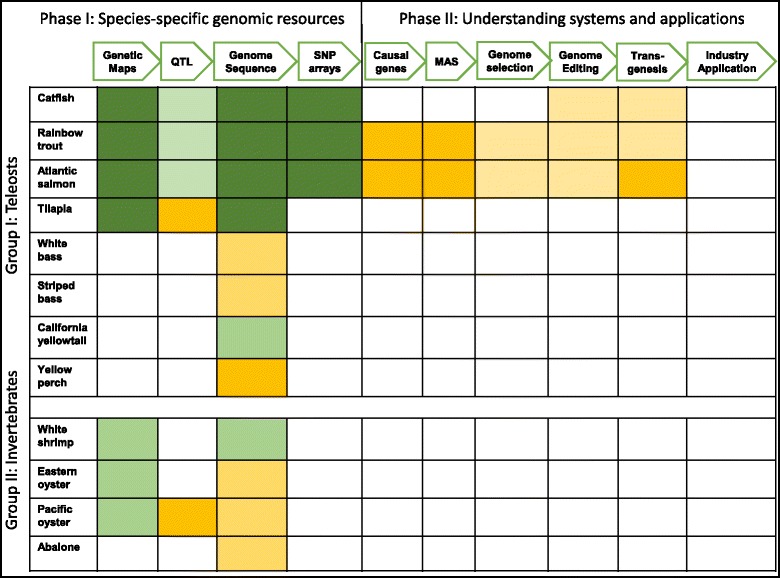
Schematic presentation of the goals and current status of aquaculture genomics and genetics research. The major aquaculture species in the United States are grouped into teleost fish and invertebrate species, with the species names listed in the first column. Major milestones of research goals are listed in the first row, while current status for each species is indicated in the appropriate cells with various colors: Dark green: good status; light green, outstanding progress has been made, but additional work still needed; dark yellow: significant progress has been made, but significant amount of additional work still needed; light yellow, some progress has been made
Within Phase I, we can make a distinction between development of resources and application of resources to commercial aquaculture. For instance, genetic maps enable QTL/MAS, and genome sequences enable genomic selection. Application of MAS/genomic selection is generally beyond the scope and funding of the academic laboratories that have participated in the development of the genomic tools that enable it, although not beyond the scope and mission of government laboratories and academic laboratories that take on genetic stock enhancement for smaller or regional aquaculture species.
In Phase II, the genomic resources developed in Phase I can be used to develop a functional understanding of animal systems. As an example, a number of laboratories are working to develop an understanding of the gene regulatory network underlying sex determination in fishes. The research program involves not only RNA-Seq to characterize patterns of gene expression in the developing gonad, but also CRISPR modifications to test linkages in the gene regulatory network model. It should be possible to develop an understanding of the gene network underlying sex determination that will be broadly applicable among aquaculture species. Similar research programs are underway to understand the genetic basis of other important traits including growth, disease resistance, etc. In each case, the goal is to develop an understanding of animal systems that can be easily transferred to related species.
When this more detailed understanding of animal systems is complete, it will become possible to make specific genetic modifications (e.g., using CRISPR) to improve animals for commercial production. The safety and effectiveness of such modifications are important topics for research, but commercial application of these technologies will require stable business models and well established regulatory frameworks that ensure the safe application of these technologies to the species being targeted, the public, and the environment.
The current status of breeding technologies in US commercial aquaculture is summarized in Table 10. With significant differences in the structures of the aquaculture industries among species, practical strategies suitable to specific situations must be developed. In addition, development of comprehensive germplasm repositories will ensure protection of valuable genetic resources of aquaculture species and the investments made in developing them.
Table 10.
Current status of breeding technologies in U.S. commercial aquaculture
| Species | Status |
|---|---|
| Catfish | Private sector efforts to conduct genetic enhancement programs appear to have been successful, but the private sector has not made a great effort in genetics and breeding. Currently, some on-farm selection is practiced, but not in a very controlled manner. Genetic improvement is primarily conducted by public sector research programs, which has resulted in 7 releases to the industry of varying impacts. Most of these fish populations were developed by mass selection and in some cases family selection with the most emphasis on growth rate. Advanced genomic tools and technologies are available but have yet to be implemented by industry. |
| The industry has widely adopted the channel female x blue male interspecific catfish hybrid which demonstrates significantly greater performance for numerous traits in comparison to the traditionally grown channel catfish with hybrids now comprising 60–70% of the industry. The vast majority of hybrids are produced with a single line of blue catfish. | |
| Atlantic salmon | Private sector breeding is integrated with a publicly funded research program. Genetic improvement is based on quantitative genetics to improve growth, fillet quality and disease traits. Due to international interest in this species advanced genome tools and technologies are widely available, their implementation in the U.S. was recently initiated in a public/private partnership with efforts to incorporate MAS for sea lice resistance. |
| In 2015 the AquAdvantage Salmon was approved for sale in the U.S. by FDA, however it is expected to reach the marketplace in 2017. | |
| Rainbow trout | Public sector breeding programs utilize quantitative genetics to select for growth performance and disease resistance in all-female populations. Chromosome set manipulation is used to provide all-female triploids for net pen operations that require sterile fish; they are also valued for their superior growth characteristics at larger sizes. |
| Publically funded research programs have released germplasm improved for growth and disease resistance characteristics. Advanced genome tools and technologies are widely available and have been implemented into the private sector. Proof of concept studies for genomic selection for disease resistance in a research population have motivated initial implementation in a commercial breeding population. | |
| Tilapia | Private sector family based breeding for Nile tilapia for improved growth, yield and disease resistance is enhanced through publicly funded research programs. Although genome tools and technologies are available, they have not yet been implemented by the private sector. |
| Striped bass | Private sector fingerling producers incorporate germplasm from wild caught and captive (domestic) populations. Significant genetic improvement has been achieved through the production of hybrids created primarily by crossing domestic striped bass males x domestic or wild caught white bass females, with parental species improvement achieved primarily via mass selection techniques. Genomic technologies are under development and have not yet incorporated into commercial breeding, although domestic striped bass and white bass are available through a publically funded research program. |
| Oysters | The Pacific oyster industry is supported through public and private programs for ploidy manipulation, family-based selection and crossbreeding. Polyploid and improved broodstocks are widely used by the U.S. West Coast industry. Genetic improvement of the eastern oyster is publically funded. For much of the past 40 years, improvements in eastern oyster growth and survival have been realized using mass-selection techniques; however, there has been a recent shift toward applying quantitative genetics and ploidy manipulation to enhance production traits. Broodstock from these breeding programs are widely used by the private sector in the Northeast and Mid-Atlantic. Genome tools for both oyster species are coming online, but have not yet been implemented. |
| Shrimp | Shrimp breeders in the public and private sector selectively breed to produce specific pathogen resistant shrimp. |
Conclusions
Based on the current status, trends, and industry needs of aquaculture genomics, genetics and breeding research, the following areas of research need to be priorities:
Phase I goals for each species
Highly contiguous and complete genome sequence
Full annotation of the genome sequence, including functional (genome to phenome) studies
Identification of genetic variants in different broodstocks, and their relationship to performance traits
Development of systems for high-throughput genotyping
Anchoring of the genome sequence to genetic maps
Identification of QTL for performance and production traits
Bioinformatic capabilities to manage these data
Training the next generation of aquaculture breeders
Establishment of high-throughput cryopreservation protocols and pathways for aquaculture species
Phase II goals for each group of species
Proof of concept demonstrations which apply genome technologies to improve production efficiency, production sustainability, animal welfare and/or product quality in the commercial sector
Development of standardized measures of organismal phenotypes
Understanding epigenetic effects that contribute to variation in gene expression
Validation of QTL, identification of the causative genetic variants underlying variations in performance, and determine the mechanisms of actions
Determine general and specific combining abilities in both intraspecific and interspecific systems
Marker-assisted selection and genome selection for production traits
Characterization of the gene regulatory networks underlying phenotypic traits important to commercial aquaculture production
Determine the genomic basis of heterosis and genomic predictors of heterosis
Identification of conserved regulatory mechanisms and pathways for growth, feed conversion efficiency, disease resistance, stress tolerance, sex and other traits among aquaculture species
Development and application of gene editing technologies and the associated regulatory frameworks, first for basic research, and eventually for commercial production
Development of tools that can be easily used by the industry
Industry applications of genome technologies
Establishment of a comprehensive germplasm repository system to protect, maintain and distribute genetic resources developed through genomic technologies
Acknowledgements
We thank Dr. Lakshmi Matukumalli and Dr. Gene Kim, national program leaders for Animal Genomics, Genetics and Breeding program, and Aquaculture Program, respectively, for their participation in the workshop, and their encouragement for writing this paper. We are grateful to the staff assistance provided by the College of Agriculture and the Office of the Vice President for Research and Economic Development at Auburn University during the workshop.
Availability of data and materials
Not applicable.
Funding
The Aquaculture Genomics, Genetics and Breeding Workshop held in Auburn, Alabama, USA was supported partially by a part of a competitive grant from the Animal Genomics, Genetics and Breeding Program of the USDA National Institute of Food and Agriculture (#2015-67015-22907), and partially by USDA NRSP-8 Aquaculture Coordinator’s funds.
Authors’ contributions
All authors are signatories of the Aquaculture Genomics, Genetics and Breeding Workshop; they have contributed to its conception and in drafting of the manuscript. ZL is the organizer of the workshop and provided the very initial draft. MG-C, XG, TK, YP, DP, BR, CR, SR, and MS wrote the first draft and contributed to initial revisions. All authors were involved in subsequent rounds of revisions, and read and approved the final manuscript. Authors are listed alphabetically based on last names.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Abbreviations
- ANN
Artificial neural networks
- CRISPR
Clustered regulatory interspaced short palindromic repeats
- ESD
Environmental sex determination
- EST
Expressed sequence tag
- FAANG
Functional Annotation of Animal Genomes
- GMO
Genetically modified organisms
- GS
Genome selection
- GSD
Genetic sex determination
- GWAS
Genome-wide association studies
- IPN
Infectious pancreatic necrosis
- IPNV
Infectious pancreatic necrosis virus
- MAS
Marker-assisted selection
- NRSP-8
National Research Support Project 8
- QTL
Quantitative trait loci
- SNP
Single nucleotide polymorphism
- SPF
Specific pathogen-free
- SSD
Sexual size dimorphism
- SVM
Support vector machines
- TALEN
Transcription activator-like effector nucleases
- ZFN
Zinc finger nuclease
References
- 1.Alcivar‐Warren A, Dunham R, Gaffney P. First aquaculture species genome mapping workshop. Anim Genet. 1997;28(6):451–452. doi: 10.1111/j.1365-2052.1997.00202.x. [DOI] [Google Scholar]
- 2.Tsai H-Y, Hamilton A, Tinch AE, Guy DR, Bron JE, Taggart JB, Gharbi K, Stear M, Matika O, Pong-Wong R. Genomic prediction of host resistance to sea lice in farmed Atlantic salmon populations. Genet Sel Evol. 2016;48:47. doi: 10.1186/s12711-016-0226-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Vallejo RL, Leeds TD, Fragomeni BO, Gao G, Hernandez AG, Misztal I, Welch TJ, Wiens GD, Palti Y. Evaluation of genome-enabled selection for bacterial cold water disease resistance using progeny performance data in rainbow trout: Insights on genotyping methods and genomic prediction models. Front Genet. 2016;7:96. doi: 10.3389/fgene.2016.00096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Vallejo RL, Leeds TD, Gao G, Parsons JE, Martin KE, Evenhuis JP, Fragomeni BO, Wiens GD, Palti Y. Genomic selection models double the accuracy of predicted breeding values for bacterial cold water disease resistance compared to a traditional pedigree-based model in rainbow trout aquaculture. Genet Select Evol. 2017;49(1):17. [DOI] [PMC free article] [PubMed]
- 5.Liu Z, Liu S, Yao J, Bao L, Zhang J, Li Y, Jiang C, Sun L, Zhang Y, Zhou T et al. The channel catfish genome sequence provides insights into the evolution of scale formation in teleosts. Nat Commun. 2016;7:11757. [DOI] [PMC free article] [PubMed]
- 6.Lien S, Koop BF, Sandve SR, Miller JR, Kent MP, Nome T, Hvidsten TR, Leong JS, Minkley DR, Zimin A, et al. The Atlantic salmon genome provides insights into rediploidization. Nature. 2016;533(7602):200–205. doi: 10.1038/nature17164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Berthelot C, Brunet F, Chalopin D, Juanchich A, Bernard M, Noël B, Bento P, Da Silva C, Labadie K, Alberti A. The rainbow trout genome provides novel insights into evolution after whole-genome duplication in vertebrates. Nat Commun. 2014;5:3657. doi: 10.1038/ncomms4657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brawand D, Wagner CE, Li YI, Malinsky M, Keller I, Fan S, Simakov O, Ng AY, Lim ZW, Bezault E, et al. The genomic substrate for adaptive radiation in African cichlid fish. Nature. 2014;513(7518):375–381. doi: 10.1038/nature13726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhang G, Fang X, Guo X, Li L, Luo R, Xu F, Yang P, Zhang L, Wang X, Qi H, et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature. 2012;490(7418):49–54. doi: 10.1038/nature11413. [DOI] [PubMed] [Google Scholar]
- 10.Zhang L, Guo X. Development and validation of single nucleotide polymorphism markers in the eastern oyster Crassostrea virginica Gmelin by mining ESTs and resequencing. Aquaculture. 2010;302(1):124–129. doi: 10.1016/j.aquaculture.2010.02.012. [DOI] [Google Scholar]
- 11.Guo X, Wang Y, Xu Z, Yang H. Chromosome set manipulation in shellfish. 2009. pp. 165–194. [Google Scholar]
- 12.Eierman LE, Hare MP. Transcriptomic analysis of candidate osmoregulatory genes in the eastern oyster Crassostrea virginica. BMC Genomics. 2014;15(1):1. doi: 10.1186/1471-2164-15-503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zeng Q, Fu Q, Li Y, Liu S, Waldbieser G, Yang Y, Bao L, Yuan Z, Li N, Liu Z. Development of 690 K SNP arrays for whole genome mapping and genetic studies in catfish. Sci Rep. 2017;7:40347. [DOI] [PMC free article] [PubMed]
- 14.Hedgecock D, Shin G, Gracey AY, Den Berg DV, Samanta MP. Second-Generation Linkage Maps for the Pacific Oyster Crassostrea gigas Reveal Errors in Assembly of Genome Scaffolds. G3: Genes/Genomes/Genetics. 2015;5(10):2007–2019. doi: 10.1534/g3.115.019570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gomez-Chiarri M, Warren WC, Guo X, Proestou D. Developing tools for the study of molluscan immunity: the sequencing of the genome of the eastern oyster, Crassostrea virginica. Fish Shellfish Immunol. 2015;46(1):2–4. doi: 10.1016/j.fsi.2015.05.004. [DOI] [PubMed] [Google Scholar]
- 16.Zhang X, Zhang Y, Scheuring C, Zhang H-B, Huan P, Wang B, Liu C, Li F, Liu B, Xiang J. Construction and characterization of a bacterial artificial chromosome (BAC) library of Pacific white shrimp, Litopenaeus vannamei. Mar Biotechnol. 2010;12(2):141–149. doi: 10.1007/s10126-009-9209-y. [DOI] [PubMed] [Google Scholar]
- 17.Liu Z, Cordes J. DNA marker technologies and their applications in aquaculture genetics. Aquaculture. 2004;238(1):1–37. doi: 10.1016/j.aquaculture.2004.05.027. [DOI] [Google Scholar]
- 18.Sun L, Liu S, Wang R, Jiang Y, Zhang Y, Zhang J, Bao L, Kaltenboeck L, Dunham R, Waldbieser G. Identification and analysis of genome-wide SNPs provide insight into signatures of selection and domestication in channel catfish (Ictalurus punctatus) PLoS One. 2014;9(10):e109666. doi: 10.1371/journal.pone.0109666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yanez JM, Naswa S, Lopez ME, Bassini L, Correa K, Gilbey J, Bernatchez L, Norris A, Neira R, Lhorente JP, et al. Genomewide single nucleotide polymorphism discovery in Atlantic salmon (Salmo salar): validation in wild and farmed American and European populations. Mol Ecol Resour. 2016;16(4):1002–1011. doi: 10.1111/1755-0998.12503. [DOI] [PubMed] [Google Scholar]
- 20.Gidskehaug L, Kent M, Hayes BJ, Lien S. Genotype calling and mapping of multisite variants using an Atlantic salmon iSelect SNP array. Bioinformatics (Oxford, England) 2011;27(3):303–310. doi: 10.1093/bioinformatics/btq673. [DOI] [PubMed] [Google Scholar]
- 21.Houston RD, Taggart JB, Cézard T, Bekaert M, Lowe NR, Downing A, Talbot R, Bishop SC, Archibald AL, Bron JE. Development and validation of a high density SNP genotyping array for Atlantic salmon (Salmo salar) BMC Genomics. 2014;15:90. doi: 10.1186/1471-2164-15-90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu S, Sun L, Li Y, Sun F, Jiang Y, Zhang Y, Zhang J, Feng J, Kaltenboeck L, Kucuktas H. Development of the catfish 250 K SNP array for genome-wide association studies. BMC Res Notes. 2014;7:135. doi: 10.1186/1756-0500-7-135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Palti Y, Gao G, Liu S, Kent M, Lien S, Miller M, Rexroad C, Moen T. The development and characterization of a 57 K single nucleotide polymorphism array for rainbow trout. Mol Ecol Resour. 2015;15(3):662–672. doi: 10.1111/1755-0998.12337. [DOI] [PubMed] [Google Scholar]
- 24.Xu J, Zhao Z, Zhang X, Zheng X, Li J, Jiang Y, Kuang Y, Zhang Y, Feng J, Li C, et al. Development and evaluation of the first high-throughput SNP array for common carp (Cyprinus carpio) BMC Genomics. 2014;15:307. doi: 10.1186/1471-2164-15-307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang J, Li L, Zhang G. A High-Density SNP Genetic Linkage Map and QTL Analysis of Growth-Related Traits in a Hybrid Family of Oysters (Crassostrea gigas × Crassostrea angulata) Using Genotyping-by-Sequencing. G3: Genes/Genomes/Genetics. 2016;6(5):1417–1426. doi: 10.1534/g3.116.026971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gonzalez-Pena D, Gao G, Baranski M, Moen T, Cleveland BM, Kenney PB, Vallejo RL, Palti Y, Leeds TD. Genome-Wide Association Study for Identifying Loci that Affect Fillet Yield, Carcass, and Body Weight Traits in Rainbow Trout (Oncorhynchus mykiss) Front Genet. 2016;7:203. doi: 10.3389/fgene.2016.00203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Ng SHS, Artieri CG, Bosdet IE, Chiu R, Danzmann RG, Davidson WS, Ferguson MM, Fjell CD, Hoyheim B, Jones SJM, et al. A physical map of the genome of Atlantic salmon. Salmo Salar Genomics. 2005;86(4):396–404. doi: 10.1016/j.ygeno.2005.06.001. [DOI] [PubMed] [Google Scholar]
- 28.Katagiri T, Kidd C, Tomasino E, Davis JT, Wishon C, Stern JE, Carleton KL, Howe AE, Kocher TD. A BAC-based physical map of the Nile tilapia genome. BMC Genomics. 2005;6:89. doi: 10.1186/1471-2164-6-89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Quiniou SM, Waldbieser GC, Duke MV. A first generation BAC-based physical map of the channel catfish genome. BMC Genomics. 2007;8:40. doi: 10.1186/1471-2164-8-40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Xu P, Wang S, Liu L, Thorsen J, Kucuktas H, Liu Z. A BAC-based physical map of the channel catfish genome. Genomics. 2007;90(3):380–388. doi: 10.1016/j.ygeno.2007.05.008. [DOI] [PubMed] [Google Scholar]
- 31.Palti Y, Luo M-C, Hu Y, Genet C, You FM, Vallejo RL, Thorgaard GH, Wheeler PA, Rexroad CE. A first generation BAC-based physical map of the rainbow trout genome. BMC Genomics. 2009;10:462. doi: 10.1186/1471-2164-10-462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Xu P, Wang J, Wang J, Cui R, Li Y, Zhao Z, Ji P, Zhang Y, Li J, Sun X. Generation of the first BAC-based physical map of the common carp genome. BMC Genomics. 2011;12:537. doi: 10.1186/1471-2164-12-537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Xia JH, Feng F, Lin G, Wang CM, Yue GH. A First Generation BAC-Based Physical Map of the Asian Seabass (Lates calcarifer) PLoS One. 2010;5(8):e11974. doi: 10.1371/journal.pone.0011974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Gaffney PM. A BAC-based physical map for the Pacific oyster genome. J Shellfish Res. 2008;27(4):1009. [Google Scholar]
- 35.Zhang X, Zhao C, Huang C, Duan H, Huan P, Liu C, Zhang X, Zhang Y, Li F, Zhang H-B, et al. A BAC-Based Physical Map of Zhikong Scallop (Chlamys farreri Jones et Preston) PLoS One. 2011;6(11):e27612. doi: 10.1371/journal.pone.0027612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Schwartz D, Li X, Hernandez L, Ramnarain S, Huff E, Wang Y. Ordered restriction maps of Saccharomyces cerevisiae chromosomes constructed by optical mapping. Science. 1993;262(5130):110–114. doi: 10.1126/science.8211116. [DOI] [PubMed] [Google Scholar]
- 37.Rajendran K, Zhang J, Liu S, Peatman E, Kucuktas H, Wang X, Liu H, Wood T, Terhune J, Liu Z. Pathogen recognition receptors in channel catfish: II. Identification, phylogeny and expression of retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) Dev Comp Immunol. 2012;37(3):381–389. doi: 10.1016/j.dci.2012.02.004. [DOI] [PubMed] [Google Scholar]
- 38.Wang R, Sun L, Bao L, Zhang J, Jiang Y, Yao J, Song L, Feng J, Liu S, Liu Z. Bulk segregant RNA-seq reveals expression and positional candidate genes and allele-specific expression for disease resistance against enteric septicemia of catfish. BMC Genomics. 2013;14:929. doi: 10.1186/1471-2164-14-929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Sun F, Peatman E, Li C, Liu S, Jiang Y, Zhou Z, Liu Z. Transcriptomic signatures of attachment, NF-κB suppression and IFN stimulation in the catfish gill following columnaris bacterial infection. Dev Comp Immunol. 2012;38(1):169–180. doi: 10.1016/j.dci.2012.05.006. [DOI] [PubMed] [Google Scholar]
- 40.Liu S, Wang X, Sun F, Zhang J, Feng J, Liu H, Rajendran K, Sun L, Zhang Y, Jiang Y. RNA-Seq reveals expression signatures of genes involved in oxygen transport, protein synthesis, folding, and degradation in response to heat stress in catfish. Physiol Genomics. 2013;45(12):462–476. doi: 10.1152/physiolgenomics.00026.2013. [DOI] [PubMed] [Google Scholar]
- 41.Valenzuela-Miranda D, Boltana S, Cabrejos ME, Yanez JM, Gallardo-Escárate C. High-throughput transcriptome analysis of ISAV-infected Atlantic salmon Salmo salar unravels divergent immune responses associated to head-kidney, liver and gills tissues. Fish Shellfish Immunol. 2015;45(2):367–377. doi: 10.1016/j.fsi.2015.04.003. [DOI] [PubMed] [Google Scholar]
- 42.Xu C, Evensen Ø, Munang’andu HM. De novo assembly and transcriptome analysis of Atlantic salmon macrophage/dendritic-like TO cells following type I IFN treatment and Salmonid alphavirus subtype-3 infection. BMC Genomics. 2015;16:96. doi: 10.1186/s12864-015-1302-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Salem M, Vallejo RL, Leeds TD, Palti Y, Liu S, Sabbagh A, Rexroad CE, III, Yao J. RNA-Seq identifies SNP markers for growth traits in rainbow trout. PLoS One. 2012;7(5):e36264. doi: 10.1371/journal.pone.0036264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Narum SR, Campbell NR. Transcriptomic response to heat stress among ecologically divergent populations of redband trout. BMC Genomics. 2015;16:103. doi: 10.1186/s12864-015-1246-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cano I, Verner-Jeffreys DW, van Aerle R, Paley RK, Peeler EJ, Green M, Rimmer GS, Savage J, Joiner CL, Bayley AE. Puffy Skin Disease Is an Emerging Transmissible Condition in Rainbow Trout Oncorhynchus mykiss Walbaum. PLoS One. 2016;11(7):e0158151. doi: 10.1371/journal.pone.0158151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Marancik D, Gao G, Paneru B, Ma H, Hernandez AG, Salem M, Yao J, Palti Y, Wiens GD. Whole-body transcriptome of selectively bred, resistant-, control-, and susceptible-line rainbow trout following experimental challenge with Flavobacterium psychrophilum. Front Genet. 2015;5:453. doi: 10.3389/fgene.2014.00453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Chapman RW, Reading BJ, Sullivan CV. Ovary transcriptome profiling via artificial intelligence reveals a transcriptomic fingerprint predicting egg quality in striped bass, Morone saxatilis. PloS One. 2014;9(5):e96818. doi: 10.1371/journal.pone.0096818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Reading BJ, Chapman RW, Schaff JE, Scholl EH, Opperman CH, Sullivan CV. An ovary transcriptome for all maturational stages of the striped bass (Morone saxatilis), a highly advanced perciform fish. BMC Res Notes. 2012;5:111. doi: 10.1186/1756-0500-5-111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sullivan CV, Chapman RW, Reading BJ, Anderson PE. Transcriptomics of mRNA and egg quality in farmed fish: Some recent developments and future directions. Gen Comp Endocrinol. 2015;221:23–30. doi: 10.1016/j.ygcen.2015.02.012. [DOI] [PubMed] [Google Scholar]
- 50.Zhao Y, Wang J, Thammaratsuntorn J, Wu J, Wei J, Wang Y, Xu J, Zhao J. Comparative transcriptome analysis of Nile tilapia (Oreochromis niloticus) in response to alkalinity stress. Genet Mol Res. 2015;14(4):17916–17926. doi: 10.4238/2015.December.22.16. [DOI] [PubMed] [Google Scholar]
- 51.Xu Z, Gan L, Li T, Xu C, Chen K, Wang X, Qin JG, Chen L, Li E. Transcriptome profiling and molecular pathway analysis of genes in association with salinity adaptation in nile tilapia Oreochromis niloticus. PLoS One. 2015;10(8):e0136506. doi: 10.1371/journal.pone.0136506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.He AY, Ning LJ, Chen LQ, Chen YL, Xing Q, Li JM, Qiao F, Li DL, Zhang ML, Du ZY. Systemic adaptation of lipid metabolism in response to low‐and high‐fat diet in Nile tilapia (Oreochromis niloticus) Physiol Rep. 2015;3(8):e12485. doi: 10.14814/phy2.12485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Clark MS, Thorne MA, Amaral A, Vieira F, Batista FM, Reis J, Power DM. Identification of molecular and physiological responses to chronic environmental challenge in an invasive species: the Pacific oyster, Crassostrea gigas. Ecol Evol. 2013;3(10):3283–3297. doi: 10.1002/ece3.719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhao X, Yu H, Kong L, Liu S, Li Q. Comparative transcriptome analysis of two oysters, Crassostrea gigas and Crassostrea hongkongensis provides insights into adaptation to hypo-osmotic conditions. PLoS one. 2014;9(11):e111915. [DOI] [PMC free article] [PubMed]
- 55.Zhao X, Yu H, Kong L, Li Q. Transcriptomic responses to salinity stress in the Pacific oyster Crassostrea gigas. PLoS One. 2012;7(9):e46244. doi: 10.1371/journal.pone.0046244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.McDowell IC, Modak TH, Lane CE, Gomez-Chiarri M. Multi-species protein similarity clustering reveals novel expanded immune gene families in the eastern oyster Crassostrea virginica. Fish Shellfish Immunol. 2016;53:13–23. doi: 10.1016/j.fsi.2016.03.157. [DOI] [PubMed] [Google Scholar]
- 57.Zhang L, Li L, Zhu Y, Zhang G, Guo X. Transcriptome analysis reveals a rich gene set related to innate immunity in the Eastern oyster (Crassostrea virginica) Mar Biotechnol. 2014;16(1):17–33. doi: 10.1007/s10126-013-9526-z. [DOI] [PubMed] [Google Scholar]
- 58.Wei J, Zhang X, Yu Y, Huang H, Li F, Xiang J. Comparative transcriptomic characterization of the early development in Pacific white shrimp Litopenaeus vannamei. PLoS One. 2014;9(9):e106201. doi: 10.1371/journal.pone.0106201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Sookruksawong S, Sun F, Liu Z, Tassanakajon A. RNA-Seq analysis reveals genes associated with resistance to Taura syndrome virus (TSV) in the Pacific white shrimp Litopenaeus vannamei. Dev Comp Immunol. 2013;41(4):523–533. doi: 10.1016/j.dci.2013.07.020. [DOI] [PubMed] [Google Scholar]
- 60.Ghaffari N, Sanchez-Flores A, Doan R, Garcia-Orozco KD, Chen PL, Ochoa-Leyva A, Lopez-Zavala AA, Carrasco JS, Hong C, Brieba LG. Novel transcriptome assembly and improved annotation of the whiteleg shrimp (Litopenaeus vannamei), a dominant crustacean in global seafood mariculture. Sci Rep. 2014;4:7081. doi: 10.1038/srep07081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Reading BJ, Williams VN, Chapman RW, Williams TI, Sullivan CV. Dynamics of the striped bass (Morone saxatilis) ovary proteome reveal a complex network of the translasome. J Proteome Res. 2013;12(4):1691–1699. doi: 10.1021/pr3010293. [DOI] [PubMed] [Google Scholar]
- 62.Schilling J, Nepomuceno A, Schaff JE, Muddiman DC, Daniels HV, Reading BJ. Compartment proteomics analysis of white perch (Morone americana) ovary using support vector machines. J Proteome Res. 2014;13(3):1515–1526. doi: 10.1021/pr401067g. [DOI] [PubMed] [Google Scholar]
- 63.Schilling J, Loziuk PL, Muddiman DC, Daniels HV, Reading BJ. Mechanisms of egg yolk formation and implications on early life history of white perch (Morone americana) PLoS One. 2015;10(11):e0143225. doi: 10.1371/journal.pone.0143225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Williams VN, Reading BJ, Amano H, Hiramatsu N, Schilling J, Salger SA, Islam Williams T, Gross K, Sullivan CV. Proportional accumulation of yolk proteins derived from multiple vitellogenins is precisely regulated during vitellogenesis in striped bass (Morone saxatilis) J Exp Zool A Ecol Genet Physiol. 2014;321(6):301–315. doi: 10.1002/jez.1859. [DOI] [PubMed] [Google Scholar]
- 65.Salem M, Kenney PB, Rexroad CE, Yao J. Proteomic signature of muscle atrophy in rainbow trout. J Proteome. 2010;73(4):778–789. doi: 10.1016/j.jprot.2009.10.014. [DOI] [PubMed] [Google Scholar]
- 66.Mennigen JA, Panserat S, Larquier M, Plagnes-Juan E, Medale F, Seiliez I, Skiba-Cassy S. Postprandial regulation of hepatic microRNAs predicted to target the insulin pathway in rainbow trout. PLoS One. 2012;7(6):e38604. doi: 10.1371/journal.pone.0038604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mennigen JA, Skiba-Cassy S, Panserat S. Ontogenetic expression of metabolic genes and microRNAs in rainbow trout alevins during the transition from the endogenous to the exogenous feeding period. J Exp Biol. 2013;216(9):1597–1608. doi: 10.1242/jeb.082248. [DOI] [PubMed] [Google Scholar]
- 68.Juanchich A, Bardou P, Rue O, Gabillard JC, Gaspin C, Bobe J, Guiguen Y. Characterization of an extensive rainbow trout miRNA transcriptome by next generation sequencing. BMC Genomics. 2016;17:164. doi: 10.1186/s12864-016-2505-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mennigen JA, Zhang D. MicroTrout: A comprehensive, genome-wide miRNA target prediction framework for rainbow trout, Oncorhynchus mykiss. Comp Biochem Physiol Part D Genomics Proteomics. 2016;20:19–26. doi: 10.1016/j.cbd.2016.07.002. [DOI] [PubMed] [Google Scholar]
- 70.Al-Tobasei R, Paneru B, Salem M. Genome-wide discovery of long non-coding RNAs in rainbow trout. PLoS One. 2016;11(2):e0148940. doi: 10.1371/journal.pone.0148940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Wang Z, Schwacke R, Kunze R. DNA damage-induced transcription of transposable elements and long non-coding RNAs in Arabidopsis is rare and ATM-dependent. Mol Plant. 2016;9(8):1142–1155. doi: 10.1016/j.molp.2016.04.015. [DOI] [PubMed] [Google Scholar]
- 72.Farlora R, Valenzuela-Miranda D, Alarcón-Matus P, Gallardo-Escárate C. Identification of microRNAs associated with sexual maturity in rainbow trout brain and testis through small RNA deep sequencing. Mol Reprod Dev. 2015;82(9):651–662. doi: 10.1002/mrd.22499. [DOI] [PubMed] [Google Scholar]
- 73.Ma H, Weber GM, Hostuttler MA, Wei H, Wang L, Yao J. MicroRNA expression profiles from eggs of different qualities associated with post-ovulatory ageing in rainbow trout (Oncorhynchus mykiss) BMC Genomics. 2015;16:201. doi: 10.1186/s12864-015-1400-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Paneru B, Al-Tobasei R, Leeds T, Kenney B, Salem M: MicroRNAs associated with muscle growth and fillet quality in rainbow trout. In: PAG-XXV Plant & Animal Genomes Conference: Plant and Animal Genome. San Diego; 2017.
- 75.Paneru B, Al-Tobasei R, Palti Y, Wiens GD, Salem M. Differential expression of long non-coding RNAs in three genetic lines of rainbow trout in response to infection with Flavobacterium psychrophilum. Sci Rep. 2016;6(36032):1–14. doi: 10.1038/srep36032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Bekaert M, Lowe NR, Bishop SC, Bron JE, Taggart JB, Houston RD. Sequencing and Characterisation of an Extensive Atlantic Salmon (Salmo salar L.) MicroRNA Repertoire. PLoS One. 2013;8(7):e70136. doi: 10.1371/journal.pone.0070136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Andreassen R, Worren MM, Høyheim B. Discovery and characterization of miRNA genes in atlantic salmon (Salmo salar) by use of a deep sequencing approach. BMC Genomics. 2013;14:482. doi: 10.1186/1471-2164-14-482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Kure EH, Saebo M, Stangeland AM, Hamfjord J, Hytterod S, Heggenes J, Lydersen E. Molecular responses to toxicological stressors: profiling microRNAs in wild Atlantic salmon (Salmo salar) exposed to acidic aluminum-rich water. Aquat Toxicol. 2013;138–139:98–104. doi: 10.1016/j.aquatox.2013.04.004. [DOI] [PubMed] [Google Scholar]
- 79.Barozai MY, Baloch IA, Din M. Identification of MicroRNAs and their targets in Helianthus. Mol Biol Rep. 2012;39(3):2523–2532. doi: 10.1007/s11033-011-1004-y. [DOI] [PubMed] [Google Scholar]
- 80.Xu Z, Qin Q, Ge J, Pan J, Xu X. Bioinformatic identification and validation of conservative microRNAs in Ictalurus punctatus. Mol Biol Rep. 2012;39(12):10395–10405. doi: 10.1007/s11033-012-1918-z. [DOI] [PubMed] [Google Scholar]
- 81.Xu Z, Chen J, Li X, Ge J, Pan J, Xu X. Identification and characterization of microRNAs in channel catfish (Ictalurus punctatus) by using Solexa sequencing technology. PLoS One. 2013;8(1):e54174. doi: 10.1371/journal.pone.0054174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Tao W, Sun L, Shi H, Cheng Y, Jiang D, Fu B, Conte MA, Gammerdinger WJ, Kocher TD, Wang D. Integrated analysis of miRNA and mRNA expression profiles in tilapia gonads at an early stage of sex differentiation. BMC Genomics. 2016;17:328. doi: 10.1186/s12864-016-2636-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Xu F, Wang X, Feng Y, Huang W, Wang W, Li L, Fang X, Que H, Zhang G. Identification of conserved and novel microRNAs in the pacific oyster Crassostrea gigas by deep sequencing. PLoS One. 2014;9(8):e104371. doi: 10.1371/journal.pone.0104371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhou Z, Wang L, Song L, Liu R, Zhang H, Huang M, Chen H. The identification and Characteristics of immune-related microRNAs in hemocytes of oyster Crassostrea gigas. PLoS One. 2014;9(2):e88397. doi: 10.1371/journal.pone.0088397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zeng D, Chen X, Xie D, Zhao Y, Yang Q, Wang H, Li Y, Chen X. Identification of highly expressed host microRNAs that respond to white spot syndrome virus infection in the Pacific white shrimp Litopenaeus vannamei (Penaeidae) Genet Mol Res. 2015;14(2):4818–4828. doi: 10.4238/2015.May.11.14. [DOI] [PubMed] [Google Scholar]
- 86.Xi Q, Xiong Y, Wang Y, Cheng X, Qi Q, Shu G, Wang S, Wang L, Gao P, Zhu X, et al. Genome-wide discovery of novel and conserved microRNAs in white shrimp (Litopenaeus vannamei) Mol Biol Rep. 2015;42(1):61–69. doi: 10.1007/s11033-014-3740-2. [DOI] [PubMed] [Google Scholar]
- 87.Andersson L, Archibald AL, Bottema CD, Brauning R, Burgess SC, Burt DW, Casas E, Cheng HH, Clarke L, Couldrey C, et al. Coordinated international action to accelerate genome-to-phenome with FAANG, the Functional Annotation of Animal Genomes project. Genome Biol. 2015;16:57. doi: 10.1186/s13059-015-0622-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Fellous A, Favrel P, Guo X, Riviere G. The Jumonji gene family in Crassostrea gigas suggests evolutionary conservation of Jmj-C histone demethylases orthologues in the oyster gametogenesis and development. Gene. 2014;538(1):164–175. doi: 10.1016/j.gene.2013.12.016. [DOI] [PubMed] [Google Scholar]
- 89.Fellous A, Favrel P, Riviere G. Temperature influences histone methylation and mRNA expression of the Jmj-C histone-demethylase orthologues during the early development of the oyster Crassostrea gigas. Mar Genomics. 2015;19:23–30. doi: 10.1016/j.margen.2014.09.002. [DOI] [PubMed] [Google Scholar]
- 90.Riviere G. Epigenetic features in the oyster Crassostrea gigas suggestive of functionally relevant promoter DNA methylation in invertebrates. Front Physiol. 2014;5(129):1–7. doi: 10.3389/fphys.2014.00129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Olson CE, Roberts SB. Genome-wide profiling of DNA methylation and gene expression in Crassostrea gigas male gametes. Front Physiol. 2014;5:224. doi: 10.3389/fphys.2014.00224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Wang X, Li Q, Lian J, Li L, Jin L, Cai H, Xu F, Qi H, Zhang L, Wu F. Genome-wide and single-base resolution DNA methylomes of the Pacific oyster Crassostrea gigas provide insight into the evolution of invertebrate CpG methylation. BMC Genomics. 2014;15:1119. doi: 10.1186/1471-2164-15-1119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Saint-Carlier E, Riviere G. Regulation of Hox orthologues in the oyster Crassostrea gigas evidences a functional role for promoter DNA methylation in an invertebrate. FEBS Lett. 2015;589(13):1459–1466. doi: 10.1016/j.febslet.2015.04.043. [DOI] [PubMed] [Google Scholar]
- 94.Morán P, Pérez-Figueroa A. Methylation changes associated with early maturation stages in the Atlantic salmon. BMC Genet. 2011;12:86. doi: 10.1186/1471-2156-12-86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Baerwald MR, Meek MH, Stephens MR, Nagarajan RP, Goodbla AM, Tomalty KMH, Thorgaard GH, May B, Nichols KM. Migration-related phenotypic divergence is associated with epigenetic modifications in rainbow trout. Mol Ecol. 2016;25(8):1785–1800. doi: 10.1111/mec.13231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Marandel L, Lepais O, Arbenoits E, Véron V, Dias K, Zion M, Panserat S. Remodelling of the hepatic epigenetic landscape of glucose-intolerant rainbow trout (Oncorhynchus mykiss) by nutritional status and dietary carbohydrates. Sci Rep. 2016;6:32187. doi: 10.1038/srep32187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kratochwil CF, Meyer A. Mapping active promoters by ChIP-seq profiling of H3K4me3 in cichlid fish- a first step to uncover cis-regulatory elements in ecological model teleosts. Mol Ecol Resour. 2015;15(4):761–771. doi: 10.1111/1755-0998.12350. [DOI] [PubMed] [Google Scholar]
- 98.Houston RD, Haley CS, Hamilton A, Guy DR, Tinch AE, Taggart JB, McAndrew BJ, Bishop SC. Major quantitative trait loci affect resistance to infectious pancreatic necrosis in Atlantic salmon (Salmo salar) Genetics. 2008;178(2):1109–1115. doi: 10.1534/genetics.107.082974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Moen T, Baranski M, Sonesson AK, Kjøglum S. Confirmation and fine-mapping of a major QTL for resistance to infectious pancreatic necrosis in Atlantic salmon (Salmo salar): population-level associations between markers and trait. BMC Genomics. 2009;10:368. doi: 10.1186/1471-2164-10-368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Houston RD, Haley CS, Hamilton A, Guy DR, Mota-Velasco JC, Gheyas AA, Tinch AE, Taggart J, Bron J, Starkey W. The susceptibility of Atlantic salmon fry to freshwater infectious pancreatic necrosis is largely explained by a major QTL. Heredity. 2010;105(3):318–327. doi: 10.1038/hdy.2009.171. [DOI] [PubMed] [Google Scholar]
- 101.Moen T, Torgersen J, Santi N, Davidson WS, Baranski M, Ødegård J, Kjøglum S, Velle B, Kent M, Lubieniecki KP. Epithelial cadherin determines resistance to infectious pancreatic necrosis virus in Atlantic salmon. Genetics. 2015;200(4):1313–1326. doi: 10.1534/genetics.115.175406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Geng X, Sha J, Liu S, Bao L, Zhang J, Wang R, Yao J, Li C, Feng J, Sun F. A genome-wide association study in catfish reveals the presence of functional hubs of related genes within QTLs for columnaris disease resistance. BMC Genomics. 2015;16:196. doi: 10.1186/s12864-015-1409-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Zhou T, Liu S, Geng X, Jin Y, Jiang C, Bao L, Yao J, Zhang Y, Zhang J, Sun L et al. QTLs for ESC disease resistance of catfish as revealed by a genome wide association study. Mol Genet Genomics. 2017;292:231–42. [DOI] [PubMed]
- 104.Jin Y, Zhou T, Geng X, Liu S, Chen A, Yao J, Jiang C, Tan S, Su B, Liu Z. A genome-wide association study of heat stress-associated SNPs in catfish. Anim Genet. 2016. doi:10.1111/age.12482. [DOI] [PubMed]
- 105.Wang X, Liu S, Jiang C, Geng X, Zhou T, Li N, Bao L, Li Y, Yao J, Yang Y et al. Multiple across-strain and within-strain QTLs suggest highly complex genetic architecture for hypoxia tolerance in channel catfish. Mol Gen Genomics. 2016. doi:10.1007/s00438-016-1256-2. [DOI] [PubMed]
- 106.Geng X, Liu S, Yao J, Bao L, Zhang J, Li C, Wang R, Sha J, Zeng P, Zhi D, et al. A Genome Wide Association Study Identifies Multiple Regions Associated with Head Size in Catfish. G3: Genes/Genomes/Genetics. 2016;6(10):3389–3398. doi: 10.1534/g3.116.032201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Vallejo RL, Palti Y, Liu S, Marancik DP, Wiens GD. Validation of linked QTL for bacterial cold water disease resistance and spleen size on rainbow trout chromosome Omy19. Aquaculture. 2014;432:139–143. doi: 10.1016/j.aquaculture.2014.05.003. [DOI] [Google Scholar]
- 108.Palti Y, Vallejo RL, Gao G, Liu S, Hernandez AG, Rexroad CE, III, Wiens GD. Detection and validation of QTL affecting bacterial cold water disease resistance in rainbow trout using restriction-site associated DNA sequencing. PLoS One. 2015;10(9):e0138435. doi: 10.1371/journal.pone.0138435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Liu S, Vallejo RL, Palti Y, Gao G, Marancik DP, Hernandez AG, Wiens GD. Identification of single nucleotide polymorphism markers associated with bacterial cold water disease resistance and spleen size in rainbow trout. Front Genet. 2015;6:298. doi: 10.3389/fgene.2015.00298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Yu Z, Guo X. Identification and mapping of disease-resistance QTLs in the eastern oyster, Crassostrea virginica Gmelin. Aquaculture. 2006;254(1):160–170. doi: 10.1016/j.aquaculture.2005.10.016. [DOI] [Google Scholar]
- 111.Plough LV, Hedgecock D. Quantitative trait locus analysis of stage-specific inbreeding depression in the Pacific oyster Crassostrea gigas. Genetics. 2011;189(4):1473–1486. doi: 10.1534/genetics.111.131854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Guo X, Li Q, Wang QZ, Kong LF. Genetic mapping and QTL analysis of growth-related traits in the Pacific oyster. Mar Biotechnol. 2012;14(2):218–226. doi: 10.1007/s10126-011-9405-4. [DOI] [PubMed] [Google Scholar]
- 113.Plough L, Shin G, Hedgecock D. Genetic inviability is a major driver of type‐III survivorship in experimental families of a highly fecund marine bivalve. Mol Ecol. 2016;25(4):895–910. doi: 10.1111/mec.13524. [DOI] [PubMed] [Google Scholar]
- 114.He Y, Yu H, Bao Z, Zhang Q, Guo X. Mutation in promoter region of a serine protease inhibitor confers Perkinsus marinus resistance in the eastern oyster (Crassostrea virginica) Fish Shellfish Immunol. 2012;33(2):411–417. doi: 10.1016/j.fsi.2012.05.028. [DOI] [PubMed] [Google Scholar]
- 115.Fuji K, Hasegawa O, Honda K, Kumasaka K, Sakamoto T, Okamoto N. Marker-assisted breeding of a lymphocystis disease-resistant Japanese flounder (Paralichthys olivaceus) Aquaculture. 2007;272(1–4):291–295. doi: 10.1016/j.aquaculture.2007.07.210. [DOI] [Google Scholar]
- 116.Ozaki A, Araki K, Okamoto H, Okauchi M, Mushiake K, Yoshida K, Tsuzaki T, Fuji K, Sakamoto T, Okamoto N. Progress of DNA marker-assisted breeding in maricultured finfish. Bull Fisheries Res Agency. 2012;35:31–37. [Google Scholar]
- 117.Chen J, Wang Y, Yue Y, Xia X, Du Q, Chang Z. A novel male-specific DNA sequence in the common carp, Cyprinus carpio. Mol Cell Probes. 2009;23(5):235–239. doi: 10.1016/j.mcp.2009.04.004. [DOI] [PubMed] [Google Scholar]
- 118.Lee BY, Penman DJ, Kocher TD. Identification of a sex-determining region in Nile tilapia (Oreochromis niloticus) using bulked segregant analysis. Anim Genet. 2003;34(5):379–383. doi: 10.1046/j.1365-2052.2003.01035.x. [DOI] [PubMed] [Google Scholar]
- 119.Ninwichian P, Peatman E, Liu H, Kucuktas H, Somridhivej B, Liu S, Li P, Jiang Y, Sha Z, Kaltenboeck L. Second-generation genetic linkage map of catfish and its integration with the BAC-based physical map. G3: Genes| Genomes| Genetics. 2012;2(10):1233–1241. doi: 10.1534/g3.112.003962. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Li L, Xiang J, Liu X, Zhang Y, Dong B, Zhang X. Construction of AFLP-based genetic linkage map for Zhikong scallop, Chlamys farreri Jones et Preston and mapping of sex-linked markers. Aquaculture. 2005;245(1–4):63–73. doi: 10.1016/j.aquaculture.2004.12.015. [DOI] [Google Scholar]
- 121.Chen S, Li J, Deng S, Tian Y, Wang Q, Zhuang Z, Sha Z, Xu J. Isolation of Female-Specific AFLP Markers and Molecular Identification of Genetic Sex in Half-Smooth Tongue Sole (Cynoglossus semilaevis) Mar Biotechnol. 2007;9(2):273–280. doi: 10.1007/s10126-006-6081-x. [DOI] [PubMed] [Google Scholar]
- 122.Pérez F, Erazo C, Zhinaula M, Volckaert F, Calderón J. A sex-specific linkage map of the white shrimp Penaeus (Litopenaeus) vannamei based on AFLP markers. Aquaculture. 2004;242(1–4):105–118. doi: 10.1016/j.aquaculture.2004.09.002. [DOI] [Google Scholar]
- 123.Li Y, Byrne K, Miggiano E, Whan V, Moore S, Keys S, Crocos P, Preston N, Lehnert S. Genetic mapping of the kuruma prawn Penaeus japonicus using AFLP markers. Aquaculture. 2003;219(1–4):143–156. doi: 10.1016/S0044-8486(02)00355-1. [DOI] [Google Scholar]
- 124.Fuji K, Yoshida K, Hattori K, Ozaki A, Araki K, Okauchi M, Kubota S, Okamoto N, Sakamoto T. Identification of the sex-linked locus in yellowtail, Seriola quinqueradiata. Aquaculture. 2010;308:S51–S55. doi: 10.1016/j.aquaculture.2010.06.035. [DOI] [Google Scholar]
- 125.Felip A, Young WP, Wheeler PA, Thorgaard GH. An AFLP-based approach for the identification of sex-linked markers in rainbow trout (Oncorhynchus mykiss) Aquaculture. 2005;247(1–4):35–43. doi: 10.1016/j.aquaculture.2005.02.014. [DOI] [Google Scholar]
- 126.Meuwissen THE, Hayes BJ, Goddard ME. Prediction of Total Genetic Value Using Genome-Wide Dense Marker Maps. Genetics. 2001;157(4):1819–1829. doi: 10.1093/genetics/157.4.1819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Hayes BJ, Bowman PJ, Chamberlain AJ, Goddard ME. Invited review: Genomic selection in dairy cattle: progress and challenges. J Dairy Sci. 2009;92(2):433–443. doi: 10.3168/jds.2008-1646. [DOI] [PubMed] [Google Scholar]
- 128.Ragavendran A, Muir WM. Genomic Selection in Aquaculture: Methods and Practical Considerations. In: Next Generation Sequencing and Whole Genome Selection in Aquaculture. Ames: Wiley-Blackwell; 2011. p. 165–183.
- 129.Sonesson AK. Genomic Selection for Aquaculture: Principles and Procedures. In: Next Generation Sequencing and Whole Genome Selection in Aquaculture. Ames: Wiley-Blackwell; 2011. p. 151–163.
- 130.Ødegård J, Moen T, Santi N, Korsvoll SA, Kjøglum S, Meuwissen THE. Genomic prediction in an admixed population of Atlantic salmon (Salmo salar) Front Genet. 2014;5:402. doi: 10.3389/fgene.2014.00402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Schilling J, Nepomuceno AI, Planchart A, Yoder JA, Kelly RM, Muddiman DC, Daniels HV, Hiramatsu N, Reading BJ. Machine learning reveals sex‐specific 17β‐estradiol‐responsive expression patterns in white perch (Morone americana) plasma proteins. Proteomics. 2015;15(15):2678–2690. doi: 10.1002/pmic.201400606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Nemudryi AA, Valetdinova KR, Medvedev SP, Zakian SM. TALEN and CRISPR/Cas Genome Editing Systems: Tools of Discovery. Acta Nat. 2014;6(3):19–40. [PMC free article] [PubMed] [Google Scholar]
- 133.Hwang WY, Fu Y, Reyon D, Maeder ML, Tsai SQ, Sander JD, Peterson RT, Yeh JRJ, Joung JK. Efficient genome editing in zebrafish using a CRISPR-Cas system. Nat Biotech. 2013;31(3):227–229. doi: 10.1038/nbt.2501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Jao LE, Wente SR, Chen W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc Natl Acad Sci. 2013;110(34):13904–13909. doi: 10.1073/pnas.1308335110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Li M, Yang H, Zhao J, Fang L, Shi H, Li M, Sun Y, Zhang X, Jiang D, Zhou L. Efficient and heritable gene targeting in tilapia by CRISPR/Cas9. Genetics. 2014;197(2):591–599. doi: 10.1534/genetics.114.163667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Elaswad A. Genetic Technologies for Disease Resistance Research and Enhancement in Catfish. PhD Dissertation. Auburn: Auburn University; 2016.
- 137.Buchanan J. Regulatory Approval of Genetically Engineered AquAdvantage Salmon. In: Plant and Animal Genome XXIV Conference: Plant and Animal Genome. San Diego; 2016.
- 138.Waltz E. Gene-edited CRISPR mushroom escapes US regulation. Nat News. 2016;532:293. doi: 10.1038/nature.2016.19754. [DOI] [PubMed] [Google Scholar]
- 139.Danchin-Burge C, Hiemstra S, Blackburn H. Ex situ conservation of holstein-friesian cattle: comparing the Dutch, French, and US germplasm collections. J Dairy Sci. 2011;94(8):4100–4108. doi: 10.3168/jds.2010-3957. [DOI] [PubMed] [Google Scholar]
- 140.Yue XP, Dechow C, Liu WS. A limited number of Y chromosome lineages is present in North American Holsteins. J Dairy Sci. 2015;98(4):2738–2745. doi: 10.3168/jds.2014-8601. [DOI] [PubMed] [Google Scholar]
- 141.Tiersch TR, Green CC. Cryopreservation in aquatic species. 2. Baton Rouge: World Aquaculture Society; 2011. [Google Scholar]
- 142.Star B, Nederbragt AJ, Jentoft S, Grimholt U, Malmstrøm M, Gregers TF, Rounge TB, Paulsen J, Solbakken MH, Sharma A. The genome sequence of Atlantic cod reveals a unique immune system. Nature. 2011;477(7363):207–210. doi: 10.1038/nature10342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.McGaugh SE, Gross JB, Aken B, Blin M, Borowsky R, Chalopin D, Hinaux H, Jeffery WR, Keene A, Ma L, et al. The cavefish genome reveals candidate genes for eye loss. Nat Commun. 2014;5:5307. doi: 10.1038/ncomms6307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Amemiya CT, Alfoldi J, Lee AP, Fan S, Philippe H, MacCallum I, Braasch I, Manousaki T, Schneider I, Rohner N, et al. The African coelacanth genome provides insights into tetrapod evolution. Nature. 2013;496(7445):311–316. doi: 10.1038/nature12027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Xu P, Zhang X, Wang X, Li J, Liu G, Kuang Y, Xu J, Zheng X, Ren L, Wang G. Genome sequence and genetic diversity of the common carp, Cyprinus carpio. Nat Genet. 2014;46(11):1212–1219. doi: 10.1038/ng.3098. [DOI] [PubMed] [Google Scholar]
- 146.Wang Y, Lu Y, Zhang Y, Ning Z, Li Y, Zhao Q, Lu H, Huang R, Xia X, Feng Q. The draft genome of the grass carp (Ctenopharyngodon idellus) provides insights into its evolution and vegetarian adaptation. Nat Genet. 2015;47(6):625–631. doi: 10.1038/ng.3280. [DOI] [PubMed] [Google Scholar]
- 147.Smith JJ, Kuraku S, Holt C, Sauka-Spengler T, Jiang N, Campbell MS, Yandell MD, Manousaki T, Meyer A, Bloom OE, et al. Sequencing of the sea lamprey (Petromyzon marinus) genome provides insights into vertebrate evolution. Nat Genet. 2013;45(4):415–421. doi: 10.1038/ng.2568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Kasahara M, Naruse K, Sasaki S, Nakatani Y, Qu W, Ahsan B, Yamada T, Nagayasu Y, Doi K, Kasai Y, et al. The medaka draft genome and insights into vertebrate genome evolution. Nature. 2007;447(7145):714–719. doi: 10.1038/nature05846. [DOI] [PubMed] [Google Scholar]
- 149.Schartl M, Walter RB, Shen Y, Garcia T, Catchen J, Amores A, Braasch I, Chalopin D, Volff J-N, Lesch K-P, et al. The genome of the platyfish, Xiphophorus maculatus, provides insights into evolutionary adaptation and several complex traits. Nat Genet. 2013;45(5):567–572. doi: 10.1038/ng.2604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Tine M, Kuhl H, Gagnaire P-A, Louro B, Desmarais E, Martins RS, Hecht J, Knaust F, Belkhir K, Klages S. European sea bass genome and its variation provide insights into adaptation to euryhalinity and speciation. Nat Commun. 2014;5:5770. doi: 10.1038/ncomms6770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Venkatesh B, Lee AP, Ravi V, Maurya AK, Lian MM, Swann JB, Ohta Y, Flajnik MF, Sutoh Y, Kasahara M, et al. Elephant shark genome provides unique insights into gnathostome evolution. Nature. 2014;505(7482):174–179. doi: 10.1038/nature12826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Chen S, Zhang G, Shao C, Huang Q, Liu G, Zhang P, Song W, An N, Chalopin D, Volff J-N. Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle. Nat Genet. 2014;46(3):253–260. doi: 10.1038/ng.2890. [DOI] [PubMed] [Google Scholar]
- 153.Jones FC, Grabherr MG, Chan YF, Russell P, Mauceli E, Johnson J, Swofford R, Pirun M, Zody MC, White S, et al. The genomic basis of adaptive evolution in threespine sticklebacks. Nature. 2012;484(7392):55–61. doi: 10.1038/nature10944. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Jaillon O, Aury J-M, Brunet F, Petit J-L, Stange-Thomann N, Mauceli E, Bouneau L, Fischer C, Ozouf-Costaz C, Bernot A, et al. Genome duplication in the teleost fish Tetraodon nigroviridis reveals the early vertebrate proto-karyotype. Nature. 2004;431(7011):946–957. doi: 10.1038/nature03025. [DOI] [PubMed] [Google Scholar]
- 155.Figueras A, Robledo D, Corvelo A, Hermida M, Pereiro P, Rubiolo JA, Gómez-Garrido J, Carreté L, Bello X, Gut M et al. Whole Genome Sequencing of Turbot (Scophthalmus maximus; Pleuronectiformes): A Fish Adapted to Demersal Life. DNA Res. 2016;23(3):181–192. [DOI] [PMC free article] [PubMed]
- 156.Wu C, Zhang D, Kan M, Lv Z, Zhu A, Su Y, Zhou D, Zhang J, Zhang Z, Xu M. The draft genome of the large yellow croaker reveals well-developed innate immunity. Nat Commun. 2014;5:5227. doi: 10.1038/ncomms6227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Howe K, Clark MD, Torroja CF, Torrance J, Berthelot C, Muffato M, Collins JE, Humphray S, McLaren K, Matthews L, et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature. 2013;496(7446):498–503. doi: 10.1038/nature12111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Liu S, Zhang Y, Zhou Z, Waldbieser G, Sun F, Lu J, Zhang J, Jiang Y, Zhang H, Wang X. Efficient assembly and annotation of the transcriptome of catfish by RNA-Seq analysis of a doubled haploid homozygote. BMC Genomics. 2012;13:595. doi: 10.1186/1471-2164-13-595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Palti Y, Gao G, Miller MR, Vallejo RL, Wheeler PA, Quillet E, Yao J, Thorgaard GH, Salem M, Rexroad CE. A resource of single‐nucleotide polymorphisms for rainbow trout generated by restriction‐site associated DNA sequencing of doubled haploids. Mol Ecol Resour. 2014;14(3):588–596. doi: 10.1111/1755-0998.12204. [DOI] [PubMed] [Google Scholar]
- 160.Christensen KA, Brunelli JP, Lambert MJ, DeKoning J, Phillips RB, Thorgaard GH. Identification of single nucleotide polymorphisms from the transcriptome of an organism with a whole genome duplication. BMC Bioinformatics. 2013;14:325. doi: 10.1186/1471-2105-14-325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Al-Tobasei R: Allelic-imbalance analysis in pooled RNA-Seq samples identifies muscle-associated genetic markers in rainbow trout: improved bioinformatics practices. In: Plant and Animal Genome XXIV Conference: Plant and Animal Genome. San Diego; 2016.
- 162.Van Bers N, Crooijmans R, Groenen M, Dibbits B, Komen J. SNP marker detection and genotyping in tilapia. Mol Ecol Resour. 2012;12(5):932–941. doi: 10.1111/j.1755-0998.2012.03144.x. [DOI] [PubMed] [Google Scholar]
- 163.Li C, Beck BH, Fuller SA, Peatman E. Transcriptome annotation and marker discovery in white bass (Morone chrysops) and striped bass (Morone saxatilis) Anim Genet. 2014;45(6):885–887. doi: 10.1111/age.12211. [DOI] [PubMed] [Google Scholar]
- 164.Yu Y, Wei J, Zhang X, Liu J, Liu C, Li F, Xiang J. SNP discovery in the transcriptome of white Pacific shrimp Litopenaeus vannamei by next generation sequencing. PLoS One. 2014;9(1):e87218. doi: 10.1371/journal.pone.0087218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Wang CM, Bai ZY, He XP, Lin G, Xia JH, Sun F, Lo LC, Feng F, Zhu ZY, Yue GH. A high-resolution linkage map for comparative genome analysis and QTL fine mapping in Asian seabass, Lates calcarifer. BMC Genomics. 2011;12:174. doi: 10.1186/1471-2164-12-174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Lien S, Gidskehaug L, Moen T, Hayes BJ, Berg PR, Davidson WS, Omholt SW, Kent MP. A dense SNP-based linkage map for Atlantic salmon (Salmo salar) reveals extended chromosome homeologies and striking differences in sex-specific recombination patterns. BMC Genomics. 2011;12:615. doi: 10.1186/1471-2164-12-615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Gharbi K, Gautier A, Danzmann RG, Gharbi S, Sakamoto T, Høyheim B, Taggart JB, Cairney M, Powell R, Krieg F, et al. A linkage map for brown trout (Salmo trutta): chromosome homeologies and comparative genome organization with other salmonid fish. Genetics. 2006;172(4):2405–2419. doi: 10.1534/genetics.105.048330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Li Y, Liu S, Qin Z, Waldbieser G, Wang R, Sun L, Bao L, Danzmann RG, Dunham R, Liu Z. Construction of a high-density, high-resolution genetic map and its integration with BAC-based physical map in channel catfish. DNA Res. 2015;22(1):39–52. doi: 10.1093/dnares/dsu038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Liu S, Li Y, Qin Z, Geng X, Bao L, Kaltenboeck L, Kucuktas H, Dunham R, Liu Z. High‐density interspecific genetic linkage mapping provides insights into genomic incompatibility between channel catfish and blue catfish. Anim Genet. 2016;47(1):81–90. doi: 10.1111/age.12372. [DOI] [PubMed] [Google Scholar]
- 170.Peng W, Xu J, Zhang Y, Feng J, Dong C, Jiang L, Feng J, Chen B, Gong Y, Chen L, et al. An ultra-high density linkage map and QTL mapping for sex and growth-related traits of common carp (Cyprinus carpio) Sci Rep. 2016;6:26693. doi: 10.1038/srep26693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Chistiakov DA, Tsigenopoulos CS, Lagnel J, Guo YM, Hellemans B, Haley CS, Volckaert FAM, Kotoulas G. A combined AFLP and microsatellite linkage map and pilot comparative genomic analysis of European sea bass Dicentrarchus labrax L. Anim Genet. 2008;39(6):623–634. doi: 10.1111/j.1365-2052.2008.01786.x. [DOI] [PubMed] [Google Scholar]
- 172.Xia JH, Liu F, Zhu ZY, Fu J, Feng J, Li J, Yue GH. A consensus linkage map of the grass carp (Ctenopharyngodon idella) based on microsatellites and SNPs. BMC Genomics. 2010;11(1):554. doi: 10.1186/1471-2164-11-554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Castaño-Sánchez C, Fuji K, Ozaki A, Hasegawa O, Sakamoto T, Morishima K, Nakayama I, Fujiwara A, Masaoka T, Okamoto H, et al. A second generation genetic linkage map of Japanese flounder (Paralichthys olivaceus) BMC Genomics. 2010;11(1):554. doi: 10.1186/1471-2164-11-554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Guyomard R, Boussaha M, Krieg F, Hervet C, Quillet E. A synthetic rainbow trout linkage map provides new insights into the salmonid whole genome duplication and the conservation of synteny among teleosts. BMC Genet. 2012;13:15. doi: 10.1186/1471-2156-13-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Jiao W, Fu X, Dou J, Li H, Su H, Mao J, Yu Q, Zhang L, Hu X, Huang X et al. High-resolution linkage and quantitative trait locus mapping aided by genome survey sequencing: building up an integrative genomic framework for a bivalve mollusc. DNA Res. 2013. doi:10.1093/dnares/dst043. [DOI] [PMC free article] [PubMed]
- 176.Tsigenopoulos CS, Louro B, Chatziplis D, Lagnel J, Vogiatzi E, Loukovitis D, Franch R, Sarropoulou E, Power DM, Patarnello T, et al. Second generation genetic linkage map for the gilthead sea bream Sparus aurata L. Mar Genomics. 2014;18 Pt A:77–82. doi: 10.1016/j.margen.2014.09.008. [DOI] [PubMed] [Google Scholar]
- 177.Baranski M, Gopikrishna G, Robinson NA, Katneni VK, Shekhar MS, Shanmugakarthik J, Jothivel S, Gopal C, Ravichandran P, Kent M, et al. The Development of a High Density Linkage Map for Black Tiger Shrimp (Penaeus monodon) Based on cSNPs. PLoS One. 2014;9(1):e85413. doi: 10.1371/journal.pone.0085413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Andriantahina F, Liu X, Huang H. Genetic map construction and quantitative trait locus (QTL) detection of growth-related traits in Litopenaeus vannamei for selective breeding applications. PLoS One. 2013;8(9):e75206. doi: 10.1371/journal.pone.0075206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Yu Y, Zhang X, Yuan J, Li F, Chen X, Zhao Y, Huang L, Zheng H, Xiang J. Genome survey and high-density genetic map construction provide genomic and genetic resources for the Pacific White Shrimp Litopenaeus vannamei. Sci Rep. 2015;5:15612. doi: 10.1038/srep15612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Lee BY, Lee WJ, Streelman JT, Carleton KL, Howe AE, Hulata G, Slettan A, Stern JE, Terai Y, Kocher TD. A second-generation genetic linkage map of tilapia (Oreochromis spp.) Genetics. 2005;170(1):237–244. doi: 10.1534/genetics.104.035022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Liu F, Sun F, Li J, Xia JH, Lin G, Tu RJ, Yue GH. A microsatellite-based linkage map of salt tolerant tilapia (Oreochromis mossambicus x Oreochromis spp.) and mapping of sex-determining loci. BMC Genomics. 2013;14:58. doi: 10.1186/1471-2164-14-58. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Ohara E, Nishimura T, Nagakura Y, Sakamoto T, Mushiake K, Okamoto N. Genetic linkage maps of two yellowtails (Seriola quinqueradiata and Seriola lalandi) Aquaculture. 2005;244(1–4):41–48. doi: 10.1016/j.aquaculture.2004.10.022. [DOI] [Google Scholar]
- 183.Aoki J-y, Kai W, Kawabata Y, Ozaki A, Yoshida K, Koyama T, Sakamoto T, Araki K. Second generation physical and linkage maps of yellowtail (Seriola quinqueradiata) and comparison of synteny with four model fish. BMC Genomics. 2015;16:406. doi: 10.1186/s12864-015-1600-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Ozaki A, Aoki J-y, Usuki H, Yoshida K, Noda T, Hotta T, Mizuochi H, Chujo T, Shima Y, Araki K: A high density genetic linkage map for yellowtail (Seriola quinqueradiata) containing EST-based SNPs. In: Plant and Animal Genome XXIV Conference: Plant and Animal Genome. San Diego; 2016
- 185.Küttner E, Moghadam HK, Skúlason S, Danzmann RG, Ferguson MM. Genetic architecture of body weight, condition factor and age of sexual maturation in Icelandic Arctic charr (Salvelinus alpinus) Mol Gen Genomics. 2011;286(1):67–79. doi: 10.1007/s00438-011-0628-x. [DOI] [PubMed] [Google Scholar]
- 186.Norman JD, Danzmann RG, Glebe B, Ferguson MM. The genetic basis of salinity tolerance traits in Arctic charr (Salvelinus alpinus) BMC Genet. 2011;12:81. doi: 10.1186/1471-2156-12-81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Liu P, Wang L, Wan ZY, Ye BQ, Huang S, Wong S-M, Yue GH. Mapping QTL for resistance against viral nervous necrosis disease in Asian seabass. Mar Biotechnol. 2016;18(1):107–116. doi: 10.1007/s10126-015-9672-6. [DOI] [PubMed] [Google Scholar]
- 188.Wang CM, Lo LC, Zhu ZY, Yue GH. A genome scan for quantitative trait loci affecting growth-related traits in an F1 family of Asian seabass (Lates calcarifer) BMC Genomics. 2006;7:274. doi: 10.1186/1471-2164-7-274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Xia JH, Lin G, He X, Yunping B, Liu P, Liu F, Sun F, Tu R, Yue GH. Mapping quantitative trait loci for omega-3 fatty acids in Asian seabass. Mar Biotechnol. 2014;16(1):1–9. doi: 10.1007/s10126-013-9524-1. [DOI] [PubMed] [Google Scholar]
- 190.Baranski M, Moen T, Våge DI. Mapping of quantitative trait loci for flesh colour and growth traits in Atlantic salmon (Salmo salar) Genet Sel Evol. 2010;42:17. doi: 10.1186/1297-9686-42-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Tsai H, Hamilton A, Guy D, Houston R. Single nucleotide polymorphisms in the insulin‐like growth factor 1 (IGF1) gene are associated with growth‐related traits in farmed Atlantic salmon. Anim Genet. 2014;45(5):709–715. doi: 10.1111/age.12202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Tsai HY, Hamilton A, Guy DR, Tinch AE, Bishop SC, Houston RD. The genetic architecture of growth and fillet traits in farmed Atlantic salmon (Salmo salar) BMC Genet. 2015;16:51. doi: 10.1186/s12863-015-0215-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Gutierrez AP, Lubieniecki KP, Fukui S, Withler RE, Swift B, Davidson WS. Detection of quantitative trait loci (QTL) related to grilsing and late sexual maturation in Atlantic salmon (Salmo salar) Mar Biotechnol. 2014;16(1):103–110. doi: 10.1007/s10126-013-9530-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Gonen S, Baranski M, Thorland I, Norris A, Grove H, Arnesen P, Bakke H, Lien S, Bishop SC, Houston R. Mapping and validation of a major QTL affecting resistance to pancreas disease (salmonid alphavirus) in Atlantic salmon (Salmo salar) Heredity. 2015;115(5):405–414. doi: 10.1038/hdy.2015.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Zhang Y, Xu P, Lu C, Kuang Y, Zhang X, Cao D, Li C, Chang Y, Hou N, Li H. Genetic linkage mapping and analysis of muscle fiber-related QTLs in common carp (Cyprinus carpio L.) Mar Biotechnol. 2011;13(3):376–392. doi: 10.1007/s10126-010-9307-x. [DOI] [PubMed] [Google Scholar]
- 196.Boulton K, Massault C, Houston RD, de Koning DJ, Haley CS, Bovenhuis H, Batargias C, Canario AV, Kotoulas G, Tsigenopoulos CS. QTL affecting morphometric traits and stress response in the gilthead seabream (Sparus aurata) Aquaculture. 2011;319(1):58–66. doi: 10.1016/j.aquaculture.2011.06.044. [DOI] [Google Scholar]
- 197.Laghari MY, Lashari P, Zhang X, Xu P, Narejo NT, Liu Y, Mehboob S, Al-Ghanim K, Zhang Y, Sun X. Mapping QTLs for swimming ability related traits in Cyprinus carpio L. Mar Biotechnol. 2014;16(6):629–637. doi: 10.1007/s10126-014-9578-8. [DOI] [PubMed] [Google Scholar]
- 198.Louro B, Kuhl H, Tine M, de Koning D-J, Batargias C, Volckaert FA, Reinhardt R, Canario AV, Power DM. Characterization and refinement of growth related quantitative trait loci in European sea bass (Dicentrarchus labrax) using a comparative approach. Aquaculture. 2016;455:8–21. doi: 10.1016/j.aquaculture.2016.01.004. [DOI] [Google Scholar]
- 199.Massault C, Hellemans B, Louro B, Batargias C, Van Houdt J, Canario A, Volckaert F, Bovenhuis H, Haley C, De Koning D. QTL for body weight, morphometric traits and stress response in European sea bass Dicentrarchus labrax. Anim Genet. 2010;41(4):337–345. doi: 10.1111/j.1365-2052.2009.02010.x. [DOI] [PubMed] [Google Scholar]
- 200.Robinson NA, Gopikrishna G, Baranski M, Katneni VK, Shekhar MS, Shanmugakarthik J, Jothivel S, Gopal C, Ravichandran P, Gitterle T. QTL for white spot syndrome virus resistance and the sex-determining locus in the Indian black tiger shrimp (Penaeus monodon) BMC Genomics. 2014;15:731. doi: 10.1186/1471-2164-15-731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Wang L, Fan C, Liu Y, Zhang Y, Liu S, Sun D, Deng H, Xu Y, Tian Y, Liao X. A genome scan for quantitative trait loci associated with Vibrio anguillarum infection resistance in Japanese flounder (Paralichthys olivaceus) by bulked segregant analysis. Mar Biotechnol. 2014;16(5):513–521. doi: 10.1007/s10126-014-9569-9. [DOI] [PubMed] [Google Scholar]
- 202.Sauvage C, Boudry P, De Koning DJ, Haley CS, Heurtebise S, Lapègue S. QTL for resistance to summer mortality and OsHV‐1 load in the Pacific oyster (Crassostrea gigas) Anim Genet. 2010;41(4):390–399. doi: 10.1111/j.1365-2052.2009.02018.x. [DOI] [PubMed] [Google Scholar]
- 203.Negrín-Báez D, Navarro A, Afonso JM, Ginés R, Zamorano MJ. Detection of QTL associated with three skeletal deformities in gilthead seabream (Sparus aurata L.): Lordosis, vertebral fusion and jaw abnormality. Aquaculture. 2015;448:123–127. doi: 10.1016/j.aquaculture.2015.05.025. [DOI] [Google Scholar]
- 204.Loukovitis D, Sarropoulou E, Tsigenopoulos CS, Batargias C, Magoulas A, Apostolidis AP, Chatziplis D, Kotoulas G. Quantitative trait loci involved in sex determination and body growth in the gilthead sea bream (Sparus aurata L.) through targeted genome scan. PLoS One. 2011;6(1):e16599. doi: 10.1371/journal.pone.0016599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Massault C, Franch R, Haley C, De Koning D, Bovenhuis H, Pellizzari C, Patarnello T, Bargelloni L. Quantitative trait loci for resistance to fish pasteurellosis in gilthead sea bream (Sparus aurata) Anim Genet. 2011;42(2):191–203. doi: 10.1111/j.1365-2052.2010.02110.x. [DOI] [PubMed] [Google Scholar]
- 206.Kocmarek AL, Ferguson MM, Danzmann RG. Co-localization of growth QTL with differentially expressed candidate genes in rainbow trout. Genome. 2015;58(9):393–403. doi: 10.1139/gen-2015-0047. [DOI] [PubMed] [Google Scholar]
- 207.Wringe BF, Devlin RH, Ferguson MM, Moghadam HK, Sakhrani D, Danzmann RG. Growth-related quantitative trait loci in domestic and wild rainbow trout (Oncorhynchus mykiss) BMC Genet. 2010;11:63. doi: 10.1186/1471-2156-11-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Leder E, Danzmann R, Ferguson M. The candidate gene, clock, localizes to a strong spawning time quantitative trait locus region in rainbow trout. J Hered. 2006;97(1):74–80. doi: 10.1093/jhered/esj004. [DOI] [PubMed] [Google Scholar]
- 209.Easton A, Moghadam H, Danzmann R, Ferguson M. The genetic architecture of embryonic developmental rate and genetic covariation with age at maturation in rainbow trout Oncorhynchus mykiss. J Fish Biol. 2011;78(2):602–623. doi: 10.1111/j.1095-8649.2010.02881.x. [DOI] [PubMed] [Google Scholar]
- 210.Miller MR, Brunelli JP, Wheeler PA, Liu S, REXROAD CE, Palti Y, Doe CQ, Thorgaard GH. A conserved haplotype controls parallel adaptation in geographically distant salmonid populations. Mol Ecol. 2012;21(2):237–249. doi: 10.1111/j.1365-294X.2011.05305.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Perry GM, Martyniuk CM, Ferguson MM, Danzmann RG. Genetic parameters for upper thermal tolerance and growth-related traits in rainbow trout (Oncorhynchus mykiss) Aquaculture. 2005;250(1):120–128. doi: 10.1016/j.aquaculture.2005.04.042. [DOI] [Google Scholar]
- 212.Baerwald M, Petersen J, Hedrick R, Schisler G, May B. A major effect quantitative trait locus for whirling disease resistance identified in rainbow trout (Oncorhynchus mykiss) Heredity. 2011;106(6):920–926. doi: 10.1038/hdy.2010.137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Campbell NR, LaPatra SE, Overturf K, Towner R, Narum SR. Association mapping of disease resistance traits in rainbow trout using restriction site associated DNA sequencing. G3: Genes/Genomes/Genetics. 2014;4(12):2473–2481. doi: 10.1534/g3.114.014621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Rodriguez MF, LaPatra S, Williams S, Famula T, May B. Genetic markers associated with resistance to infectious hematopoietic necrosis in rainbow and steelhead trout (Oncorhynchus mykiss) backcrosses. Aquaculture. 2004;241(1):93–115. doi: 10.1016/j.aquaculture.2004.08.003. [DOI] [Google Scholar]
- 215.Le Bras Y, Dechamp N, Krieg F, Filangi O, Guyomard R, Boussaha M, Bovenhuis H, Pottinger TG, Prunet P, Le Roy P. Detection of QTL with effects on osmoregulation capacities in the rainbow trout (Oncorhynchus mykiss) BMC Genet. 2011;12(1):46. doi: 10.1186/1471-2156-12-46. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Rexroad CE, Vallejo RL, Liu S, Palti Y, Weber GM. Quantitative trait loci affecting response to crowding stress in an F2 generation of rainbow trout produced through phenotypic selection. Mar Biotechnol. 2013;15(5):613–627. doi: 10.1007/s10126-013-9512-5. [DOI] [PubMed] [Google Scholar]
- 217.Liu S, Vallejo RL, Gao G, Palti Y, Weber GM, Hernandez A, Rexroad CE., III Identification of single-nucleotide polymorphism markers associated with cortisol response to crowding in rainbow trout. Mar Biotechnol. 2015;17(3):328–337. doi: 10.1007/s10126-015-9621-4. [DOI] [PubMed] [Google Scholar]
- 218.Sánchez-Molano E, Cerna A, Toro MA, Bouza C, Hermida M, Pardo BG, Cabaleiro S, Fernández J, Martínez P. Detection of growth-related QTL in turbot (Scophthalmus maximus) BMC Genomics. 2011;12:473. doi: 10.1186/1471-2164-12-473. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Rodríguez-Ramilo ST, Toro MA, Bouza C, Hermida M, Pardo BG, Cabaleiro S, Martínez P, Fernández J. QTL detection for Aeromonas salmonicida resistance related traits in turbot (Scophthalmus maximus) BMC Genomics. 2011;12:541. doi: 10.1186/1471-2164-12-541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Rodríguez‐Ramilo S, Fernández J, Toro M, Bouza C, Hermida M, Fernández C, Pardo B, Cabaleiro S, Martínez P. Uncovering QTL for resistance and survival time to Philasterides dicentrarchi in turbot (Scophthalmus maximus) Anim Genet. 2013;44(2):149–157. doi: 10.1111/j.1365-2052.2012.02385.x. [DOI] [PubMed] [Google Scholar]
- 221.Rodríguez-Ramilo ST, De La Herrán R, Ruiz-Rejón C, Hermida M, Fernández C, Pereiro P, Figueras A, Bouza C, Toro MA, Martínez P. Identification of quantitative trait loci associated with resistance to viral haemorrhagic septicaemia (VHS) in turbot (Scophthalmus maximus): a comparison between bacterium, parasite and virus diseases. Mar Biotechnol. 2014;16(3):265–276. doi: 10.1007/s10126-013-9544-x. [DOI] [PubMed] [Google Scholar]
- 222.Liu F, Sun F, Xia JH, Li J, Fu GH, Lin G, Tu RJ, Wan ZY, Quek D, Yue GH. A genome scan revealed significant associations of growth traits with a major QTL and GHR2 in tilapia. Sci Rep. 2014;4:7256. doi: 10.1038/srep07256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Wang R, Li L, Huang Y, Luo F, Liang W, Gan X, Huang T, Lei A, Chen M, Chen L. Comparative genome analysis identifies two large deletions in the genome of highly-passaged attenuated Streptococcus agalactiae strain YM001 compared to the parental pathogenic strain HN016. BMC Genomics. 2015;16:897. doi: 10.1186/s12864-015-2026-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Palaiokostas C, Bekaert M, Khan MG, Taggart JB, Gharbi K, McAndrew BJ, Penman DJ. A novel sex-determining QTL in Nile tilapia (Oreochromis niloticus) BMC Genomics. 2015;16:171. doi: 10.1186/s12864-015-1383-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.