

Abstract
Susceptibility to tuberculosis (TB) is partially dependent on host genetic variability. SP110 and PMP22 are candidate genes identified in this study as associated with human susceptibility to TB. Here we performed an association analysis in a case-control study of a Tibetan population (217 cases and 383 controls). Using bioinformatics methods, we identified two SNPs in SP110 that may decrease susceptibility to TB (rs4327230, p<0.001, OR: 0.37, 95%CI: 0.25-0.55; rs2114591, p<0.001, OR: 0.59, 95%CI: 0.45-0.78), whereas one SNP in PMP22 appeared to increase TB risk (rs13422, p=0.003, OR: 1.45, 95%CI: 1.14-1.84). SNPs rs4327230 and rs2114591 remained significant after Bonferroni correction (p<0.00178). We found that the “GC” haplotype in SP110 was protective against TB, with a 64% reduction in disease risk. “CA” and “CG” in PMP22 were also associated with a protective effect. Our study indicates there is an association between specific gene polymorphisms and TB risk in a Tibetan population, and may help to identify those TB-affected individuals most susceptible to disease.
Keywords: SP110, PMP22, single nucleotide polymorphism (SNP), tuberculosis (TB), Tibet
INTRODUCTION
Tuberculosis (TB) is a highly infectious respiratory disease that remains one of the world's deadliest communicable diseases. According to the World Health Organization (WHO), an estimated 9 million people developed TB and 1.5 million died of the disease in 2013. Although one third of the world's population is infected with the bacillus Mycobacterium tuberculosis, the causative agent of TB, only about 10% will eventually develop clinical disease [1]. Whether or not an individual ultimately develops active disease depends upon both the host immune response and environmental aspects [2]. Genetic factors play an important role in immune system function and are thought to be the key factor in determining an individual's susceptibility to TB [2-5].
The SP110 gene on chromosome 2 encodes a leukocyte-specific nuclear body component that can function as an activator of gene transcription and may serve as a nuclear hormone receptor coactivator. SP110 is a promising candidate gene in the regulation of individual susceptibility to M. tuberculosis. First reported by Pan H, et al. [6], SP110 is the closest human homolog of the mouse gene Ipr1 (Intracellular Pathogen Resistance 1), which protects against tuberculosis by encoding Speckled proteins (SP) that regulate cell activation, division and apoptosis [7]. Previous studies associated SP110 polymorphisms and susceptibility to TB in north Indian [8], Vietnamese [9] and Chinese Han [10] populations, although some groups reported negative results [11, 12]. In addition, the PMP22 (peripheral myelin protein 22) gene on chromosome 17, previously reported to be associated with Charcot Marie Tooth (CMT) disease or hereditary neuropathy with pressure palsies (HNPP), may also play an important role in susceptibility to TB [13-15]. However, neither SP110 nor PMP22 has yet been investigated with respect to TB in the Tibetan population.
To investigate the association between SP110 and PMP22 and TB risk in Tibetans, we performed an association analysis in a case-control study. Our findings will help to improve patient-specific TB treatment and diagnosis options in the Tibetan population.
RESULTS
We recruited 217 cases (118 males and 99 females; average age at diagnosis: 49.9) and 383 controls (153 males and 230 females; average age: 20.3) for this study (Table 1). Multivariate analyses were adjusted for age and sex.
Table 1. Patient demographics.
| Variables | Case (N=217) | Control (N=383) | Total | p-value |
|---|---|---|---|---|
| Sex, No.(%) | < 0.05a | |||
| Male | 118 (43.3) | 153 (60) | 287 (52.5) | |
| Female | 99 (56.7) | 230 (40) | 260 (47.5) | |
| Mean age ±SD | 33.2 ±2.75 | 49.9 ±5.14 | < 0.05b |
P values was calculated from Pearson's chi-square tests.
P values was calculated by Welch's t tests.
To design the multiplexed SNPs MassEXTEND assay, we used Sequenom MassARRAY Assay Design 3.0 Software (PCR primers are shown in Table 2). A total of seven SNPs (four in SP110 and three in PMP22) were identified in cases and controls. All SNP call rates exceeded 98.2%, which was considered high enough to perform association analyses. SNPs in controls were all in Hardy-Weinberg equilibrium (HWE) (Table 3). We found three SNPs in SP110 (rs6436917, p=0.022, OR: 1.32, 95%CI: 1.04–1.67; rs4327230, p<0.001, OR: 0.37, 95%CI: 0.25–0.55; rs2114591, p<0.001, OR: 0.59, 95%CI: 0.45–0.78) and one in PMP22 (rs13422, p=0.003, OR: 1.45, 95%CI: 1.14–1.84) that exhibited differential allele frequency distributions in cases vs. controls. Rs4327230 and rs2114591 in SP110 remained significant after Bonferroni correction (p<0.00178).
Table 2. PCR primers used in this study.
| SNP_ID | 1st-PCR primer | 2nd-PCR primer | UEP_SEQ |
|---|---|---|---|
| rs6436917 | ACGTTGGATGGCCAATTGTAAGTGCCAAAG | ACGTTGGATGATGGCCCTAAAATGTTCCAC | ggAATGTTCCACAGTGGGC |
| rs4327230 | ACGTTGGATGACAATAGCAAAGACATGGGC | ACGTTGGATGTATGGCTGCATAGTATTCCG | ACATATTCTTTATCCAGTCTATAATT |
| rs2114591 | ACGTTGGATGGAGACGTACAGAAAAGAGGG | ACGTTGGATGCTTCCAACAACCACTGTCAC | aacaACTGTCACATCAACAGATCC |
| rs7580912 | ACGTTGGATGACACCCTCTCTCCTAACTAC | ACGTTGGATGGTCTCAAACCACAAACCACC | aaaaaCACAAACCACCAGGGCTTCC |
| rs13422 | ACGTTGGATGTTGGGATTTTGGGCTAGCTC | ACGTTGGATGTCTGAGCGTACATAGGGAAG | ggagcGGGAAGGGAGGAAGGGAAA |
| rs230915 | ACGTTGGATGCAAGAAGCCTTTCAGTTGAG | ACGTTGGATGCCTGACTCTTAGTAAGTCAA | ttttcAGTAAGTCAATAGAGTGCTGC |
| rs231024 | ACGTTGGATGAGCTGGATAAACAGGTCTGG | ACGTTGGATGCCATGCCTGGCCATCTTATT | gGGCCATCTTATTATTATTTAATGA |
UEP: Unextended mini-sequencing primer.
Table 3. Allele frequencies in cases and controls and odds ratio estimates for TB.
| SNP ID | Gene | Position | Alleles Aa/B | p value | MAF | HWE p | ORs(95%CI) | p value | |
|---|---|---|---|---|---|---|---|---|---|
| case | control | ||||||||
| rs6436917 | SP110 | 231038160 | G/A | 0.0221* | 0.514 | 0.445 | 1.0000 | 1.32 (1.04,1.67) | 0.0221* |
| rs4327230 | SP110 | 231046783 | G/C | 0.0001* | 0.078 | 0.188 | 0.7379 | 0.37 (0.25,0.55) | 0.0001* |
| rs2114591 | SP110 | 231050569 | C/T | 0.0002 * | 0.203 | 0.302 | 0.3317 | 0.59 (0.45,0.78) | 0.0002 * |
| rs7580912 | SP110 | 231081405 | C/T | 0.7104 | 0.399 | 0.388 | 0.5182 | 1.05 (0.82,0.14) | 0.7104 |
| rs13422 | PMP22 | 15134175 | A/C | 0.0025* | 0.447 | 0.358 | 0.4358 | 1.45 (1.14,1.84) | 0.0025* |
| rs230915 | PMP22 | 15148256 | G/A | 0.2531 | 0.268 | 0.299 | 0.8060 | 0.86 (0.66,1.12) | 0.2531 |
| rs231024 | PMP22 | 15157972 | A/G | 0.0700 | 0.281 | 0.332 | 0.4193 | 0.79 (0.61,1.02) | 0.0700 |
MAF, minor allelic frequency; HWE, Hardy-Weinberg Equilibrium; ORs, odds ratios; CI: confidence interval.
Minor allele; *p≤0.05.
Bonferroni's multiple adjustment was applied, with p<0.00178 (0.05/28).
We then analyzed associations between SP110 and PMP22 genotypes and TB susceptibility (Table 4). Genotypes “GG” in rs6436917 (p=0.021, OR: 1.72, 95%CI: 1.08–2.74) and “AA” in rs13422 (p=0.002, OR: 2.20, 95%CI: 1.32–3.66) were associated with TB risk. A protective effect was also associated with genotypes “GG” in rs4327230 (p=0.035, OR: 0.23, 95%CI: 0.05–1.02) and “CC” in rs2114591 (p=0.004, OR: 0.35, 95%CI: 0.17–0.73). The “GG” genotype of rs4327230 remained significant after Bonferroni correction.
Table 4. Association between SP110 and PMP22 tSNP genotypes and TB risk.
| SNP_ID | Genotype | No. (Frequency) | OR | 95%CI | p value | |
|---|---|---|---|---|---|---|
| Case | Control | |||||
| rs6436917 | GG | 61 (28.24%) | 76 (19.84%) | 1.72 | 1.08,2.74 | 0.021 |
| GA | 100 (46.30%) | 189 (49.35%) | 1.14 | 0.76,1.70 | 0.536 | |
| AA | 55 (25.46%) | 118 (30.81%) | 1 | - | ||
| rs4327230 | GG | 2 (0.92%) | 12 (3.13%) | 0.23 | 0.05,1.02 | 0.035 |
| GC | 30 (13.82%) | 120 (31.33%) | 0.34 | 0.21,0.53 | <0.001* | |
| CC | 185 (85.25%) | 251 (65.54%) | 1 | - | ||
| rs2114591 | CC | 10 (4.61%) | 39 (10.18%) | 0.35 | 0.17,0.730 | 0.004* |
| CT | 68 (31.34%) | 153 (39.95%) | 0.61 | 0.43,0.88 | 0.007* | |
| TT | 139 (64.06%) | 191 (49.87%) | 1 | - | ||
| rs7580912 | CC | 34 (15.67%) | 54 (14.10%) | 1.13 | 0.68,1.89 | 0.639 |
| CT | 105 (48.39%) | 189 (49.35%) | 1.00 | 0.69,1.44 | 0.988 | |
| TT | 78 (35.94%) | 140 (36.55%) | 1 | - | ||
| rs13422 | AA | 42 (19.35%) | 45 (11.81%) | 2.20 | 1.32,3.66 | 0.002* |
| AC | 110 (50.69%) | 183 (48.03%) | 1.41 | 0.97,2.06 | 0.069 | |
| CC | 65 (29.95%) | 153 (40.16%) | 1 | - | ||
| rs230915 | GG | 13 (6.10%) | 35 (9.26%) | 0.62 | 0.32,1.22 | 0.165 |
| GA | 88 (41.31%) | 156 (41.27%) | 0.94 | 0.66,1.34 | 0.738 | |
| AA | 112 (52.58%) | 187 (49.47%) | 1 | - | ||
| rs231024 | AA | 16 (7.37%) | 38 (9.92%) | 0.63 | 0.34.191 | 0.154 |
| AG | 90 (41.47%) | 178 (46.48%) | 0.76 | 0.54,1.08 | 0.124 | |
| GG | 111 (51.15%) | 167 (43.60%) | 1 | - | ||
OR: odd ratio; CI: confidence interval.
*p≤0.05.
Bonferroni's multiple adjustment was applied, with p<0.00178 (0.05/28).
We assumed that the minor allele of each SNP was a TB risk factor compared to the wild type allele and analyzed associations between SNPs and TB in various inheritance models (Table 5). Rs6436917 was associated with TB risk in a recessive model (p=0.019, OR: 1.59, 95%CI: 1.06–2.39), while rs4327230 was significant in a dominant model (p<0.001, OR: 0.33, 95%CI: 0.21–0.51). Two other SNPs were associated with TB in both dominant (rs2114591, p<0.001, OR: 0.56, 95%CI: 0.39–0.80; rs13422, p=0.013, OR: 1.57, 95%CI: 1.09–2.28) and recessive models (rs2114591, p=0.017, OR: 0.43, 95%CI: 0.19–0.89; rs13422, p=0.012, OR: 1.79, 95%CI: 1.10–2.91). Rs4327230 and rs2114591 remained significant in the dominant model after Bonferroni correction (p<0.00178).
Table 5. Association between SP110 and PMP22 tSNPs and TB risk based on different inheritance models.
| SNP_ID | Model | Genotype | Case | Control | OR | 95%CI | p value |
|---|---|---|---|---|---|---|---|
| rs6436917 | Dominant | G-G/G-A | 161 | 265 | 1.30 | 0.88,1.94 | 0.1656 |
| A-A | 55 | 118 | |||||
| Recessive | G-G | 61 | 76 | 1.59 | 1.06,2.39 | 0.0188* | |
| G-A/A-A | 155 | 307 | |||||
| rs4327230 | Dominant | G-G/G-C | 32 | 132 | 0.33 | 0.27,0.51 | 0.0001* |
| C-C | 185 | 251 | |||||
| Recessive | G-G | 2 | 12 | 0.29 | 0.03,1.31 | 0.0847 | |
| G-C/C-C | 215 | 371 | |||||
| rs2114591 | Dominant | C-C/C-T | 78 | 192 | 0.56 | 0.393,0.80 | 0.0008* |
| T-T | 139 | 191 | |||||
| Recessive | C-C | 10 | 39 | 0.43 | 0.19,0.89 | 0.0166* | |
| C-T/T-T | 207 | 344 | |||||
| rs7580912 | Dominant | C-C/C-T | 139 | 243 | 1.03 | 0.72,1.48 | 0.8816 |
| T-T | 78 | 140 | |||||
| Recessive | C-C | 34 | 54 | 1.13 | 0.69,1.84 | 0.6017 | |
| C-T/T-T | 183 | 329 | |||||
| rs13422 | Dominant | A-A/A-C | 152 | 228 | 1.57 | 1.08,2.28 | 0.0127* |
| C-C | 65 | 153 | |||||
| Recessive | A-A | 42 | 45 | 1.79 | 1.10,2.91 | 0.0119* | |
| A-C/C-C | 175 | 336 | |||||
| rs230915 | Dominant | G-G/G-C | 101 | 191 | 0.88 | 0.62,1.25 | 0.4676 |
| C-C | 112 | 187 | |||||
| Recessive | G-G | 13 | 35 | 0.64 | 0.30,1.27 | 0.1775 | |
| G-C/C-C | 200 | 343 | |||||
| rs231024 | Dominant | A-A/A-G | 106 | 216 | 0.74 | 0.52,1.05 | 0.0748 |
| G-G | 111 | 167 | |||||
| Recessive | A-A | 16 | 38 | 0.73 | 0.37,1.37 | 0.2946 | |
| A-G/G-G | 201 | 345 |
OR: odd ratio; CI: confidence interval.
*p≤0.05.
Bonferroni's multiple adjustment was applied, with p 0.00178 (0.05/28).
We detected two blocks in SP110 and PMP22 via haplotype analysis (Figure 1 & 2). Block 1 in SP110 included rs4327230 and rs2114591 and Block 2 in PMP22 included rs13422 and rs230915. The global result for Block 1 (rs4327230 and rs2114591) was: total case = 434, total control = 766, global χ2 = 26.416 while df = 2, Pearson's p value<0.001. The result for Block 2 (rs13422 and rs230915) was: total case = 434, total control = 766, global χ2=10.567 while df = 3, Pearson's p value=0.014. We did not perform Fisher's exact probabilities because the total sample size exceeded 40 and all theoretical frequencies were >1.
Figure 1. Haplotype block map for SP110 tSNPs.
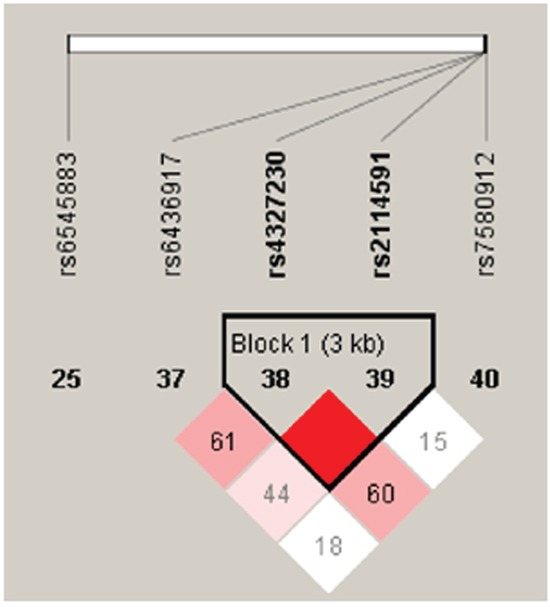
Block 1 includes rs4327230 and rs2114591. The coefficient of linkage disequilibrium (D) between two SNPs is normalized to D' (D/Dmax) (red schemes).
Figure 2. Haplotype block map for PMP22 tSNPs.
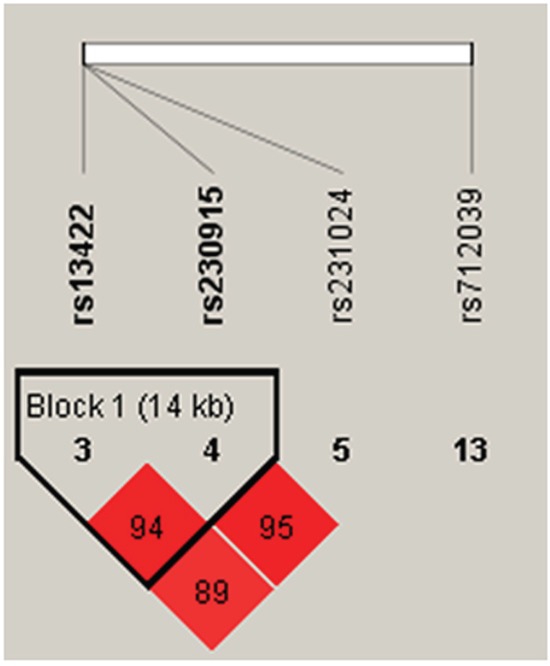
Block 2 includes rs13422 and rs230915. The coefficient of linkage disequilibrium (D) between two SNPs is normalized to D' (D/Dmax) (red schemes).
Further analyses of associations between SP110 and PMP22 haplotypes and TB risk showed that haplotype “GC” in Block 1 was protective against TB (p<0.001, OR: 0.36, 95%CI: 0.24–0.54), which remained significant after Bonferroni correction (p<0.00178) (Table 6). Both haplotypes “CA” (p=0.024, OR: 0.71, 95%CI: 0.53–0.96) and “CG” (p=0.021, OR: 0.70, 95%CI: 0.52–0.95) were also protective against TB. These results suggested that rs13422 allele A might be a risk factor for TB susceptibility.
Table 6. Association of haplotype frequencies with TB risk in cases and controls.
| Block | SNPs | Haplotypes | Frequencies | χ2 | Pearson's p | OR | 95%CI | |
|---|---|---|---|---|---|---|---|---|
| case | control | |||||||
| 1 | rs4327230 | rs2114591 | CT | 0.797 | 0.698 | - | - | 1 (reference) | - |
| GC | 0.078 | 0.188 | 26.365 | <0.001* | 0.36 | 0.24-0.54 | ||
| CC | 0.124 | 0.114 | 0.314 | 0.84 | 0.96 | 0.68-1.37 | ||
| 2 | rs13422 | rs230915 | AA | 0.429 | 0.351 | - | - | 1 (reference) | - |
| CA | 0.302 | 0.350 | 2.869 | 0.024* | 0.71 | 0.53-0.96 | ||
| CG | 0.251 | 0.291 | 2.203 | 0.021* | 0.70 | 0.52-0.95 | ||
| AG | 0.018 | 0.008 | 2.518 | 0.190 | 2.60 | 0.62-10.90 | ||
OR: odd ratio; CI: confidence interval
Bonferroni's multiple adjustment was applied, with p<0.00178 (0.05/28).
DISCUSSION
Multiple groups have investigated associations between SP110 and TB susceptibility in a variety of populations, but these study results are generally considered controversial [8-10, 12, 16-18]. In the current case-control study, we investigated seven total SNPs in a Tibetan population and found that three in SP110 (rs6436917, rs4327230, rs2114591) and one in PMP22 (rs13422) were associated with TB risk. To the best of our knowledge, this is the first study to demonstrate an association between PMP22 and TB, and the first study of SP110 in Tibetans. We found that the “GC” haplotype in SP110 was protective against TB, with a 64% reduction in disease risk. “CA” and “CG” in PMP22 were also associated with a protective effect.
Two SP110 SNPs, rs2114591 and rs6436917, were particularly notable in this study. The rs2114591 C allele in intron 11 of SP110 greatly reduced TB risk in the study population. However, a previous study by Abhimanyu, et al. [8] showed no association between rs2114591 and TB in lymph node tuberculosis in north Indians. Moreover, neither Thye, et al. [12] nor Png, et al. [17] found any association between rs2114591 and TB risk in Indonesians or the republic of Ghana, respectively. Rs6436917 was also associated with TB in our study. Abhimanyu, et al. [8] did not detect this SNP in their study, but instead associated another SNP 3636 bp upstream of rs6436917, rs6436915, with TB risk (OR=3, 95%CI, 1.3-6.3; p=0.0057). Our study and others indicate that gene expression may differ among races, and this should be taken into consideration when determining an individual patient's susceptibility to TB.
PMP22 encodes an integral membrane protein that is a major component of myelin in the peripheral nervous system. It is expressed in Schwann cells and also in non-neural cells such as the lung epithelium, and influences cell proliferation [19]. In the current study, we identified the rs13422 A allele in the PMP22 3′UTR as a TB risk factor in a Tibetan population. This is the first study to associate PMP22 and TB risk.
TB elicits a kind of type IV allergic reaction (also called delayed type hypersensitivity, DTH). The main pathological processes involved include exudation, alteration and proliferation, which depend on both the host immune system and M. tuberculosis strain virulence. Mouse models have been employed to explore the impacts of Ipr1, encoded within the sst1 locus, on TB susceptibility. Kramnik [7] showed that sst1 gene expression may be regulated by macrophages in a cell-autonomous manner. Chackerian, et al. [20] also found increased numbers of IFN-γ-producing CD4+ T cells in the lungs of sst1-resisitant C57BL/6 inbred mice, and conjectured that TB susceptibility was related to the ability to recruit mycobacteria-specific IFN-γ-producing Th1 cells to the lungs. Mechanisms of TB resistance via Ipr1 and sst1 have been extensively explored, but studies of SP110 with respect to TB in humans are rare, and no work had yet addressed PMP22, which is primarily associated with neural diseases, in TB. Though we found that PMP22 is a risk factor for TB, the mechanisms by which PMP22 affects immunity are unknown and require further study.
The Bonferroni correction is one of the most important methods used to address false discovery rates resulting from multiple testing. We found that SNPs rs4327230 and rs2114591 remained significant after Bonferroni correction, while rs6436917 and rs13422 were not associated with TB. This may due to our strict SNP filtering criteria and small sample size. Additionally, the Bonferroni correction adjusts the value of alpha based on the number of tests performed, and is thus conservative; in some cases, truly significant differences may be deemed non-significant as a result of type II errors [21].
Our study faced some intrinsic limitations. Our patient sample size (217 cases and 383 controls) was not large enough for the association analysis. Other studies included more than 1000 samples [22], and their SNP-TB association findings are therefore more reliable. Additionally, the case groups we selected included patients diagnosed with clinical tuberculosis (sputum smear-positive), regardless of their typing, which could potentially reduce the validity of our conclusions.
Our study provides new evidence supporting the importance of host genetic variability in TB susceptibility. We identified SP110 genotypes that appear protective against TB, and identified PMP22 genotypes as risk factors in TB susceptibility. Our study may help to identify those TB-affected individuals most susceptible to disease, and to improve patient-specific clinical TB diagnosis.
MATERIALS AND METHODS
Ethics statement
We strictly obeyed the World Medical Association Declaration of Helsinki when using human tissue and signing the study protocol with subjects, which was approved by the Ethical Committee of Xi'an Jiaotong University. Each participant provided written, informed consent.
Subjects
All participants in our study were Tibetan Chinese. A total of 143 TB patients were consecutively recruited between March 2013 and June 2015 in the Tangdu Hospital, affiliated with The Fourth Military Medical University in Xi'an city, China. Patients were diagnosed with TB using symptoms and signs, chest X-ray, sputum smear test for acid fast bacilli (AFB) and M. tuberculosis sputum culture. All patients were HIV negative, which was confirmed by the HIV serology test. There were no age, sex or TB classification restrictions when enrolling the case group. Controls were healthy people receiving physical examinations in other clinical departments of Tangdu Hospital. Healthy controls did not have respiratory disease, immune system disease or any other infectious disease, such as acquired immunodeficiency syndrome (AIDS), that could have affected our study results. Peripheral blood was collected from both cases and controls for DNA extraction.
Clinical data and patient demographics
A standardized epidemiological questionnaire was provided to all subjects to collect basic demographic information, including sex, age, residence, educational status, history of family cancer, history of smoking, alcohol consumption and TB contact history. Plasma carcinoembryonic antigen and alpha-fetoprotein levels were determined to ensure that no controls suffered any cancer.
SNP selection and genotyping
Candidate SNPs in the SP110 and PMP22 genes were selected from previous publications that associated polymorphisms with TB [23, 24]. SNPs with minor allele frequencies (MAF) > 5% in the HapMap CHB population were selected. We validated four SNPs in SP110 and three in PMP22. The GoldMag-Mini Purification Kit (GoldMag Co. Ltd. Xian city, China) was used to extract genomic DNA from whole blood samples. DNA concentration was measured using a DU530 UV/VIS spectrophotometer (Beckman Instruments, Fullerton, CA, USA). Using MassARRAY Assay Design 3.0 software (Sequenom, San Diego, CA, USA), we designed a multiplexed SNP MassEXTENDED assay. SNPs were genotyped using the standard protocol recommended by the MassARRAY RS1000 (Sequenom) manufacturer and data were analyzed using Typer 4.0 Software (Sequenom).
Statistical analysis
We used Microsoft Excel and SPSS 16.0 (SPSS, Chicago, IL) statistical packages to perform statistical analyses. All p-values were two-sided and p<0.05 was considered statistically significant. A t test and Chi-square test were performed to compare sex and age differences between cases and controls. Fisher's exact test was applied to each SNP in the controls to test for departure from Hardy–Weinberg Equilibrium (HWE). Odds ratios (ORs) and 95% confidence intervals (CIs) for the allele and genotype frequencies were calculated using Pearson Chi-square test adjusted by age and gender. Cochran-Armitage trend test was performed to determine associations between SNPs and TB. Sex and age subgroups were not separated due to sample size limitations.
PLINK software (http://pngu.mgh.harvard.edu/purcell/plink/) was used to assess SNP associations with TB risk in different genetic models (dominant and recessive). We used unconditional logistic regression analysis to calculate ORs and 95% CI adjusted for age and gender. Pairwise linkage disequilibrium and haplotype constructions were performed using HAPLOVIEW 4.1 (http://broad.mit.edu/mpg/haploview) [25]. All p values were Bonferroni corrected, and statistical significance was set at p<0.00178 (0.05/28).
Acknowledgments
We would like to thank all the patients and individuals in this study for their participation. We are also very grateful for the assistance of the clinicians and other hospital staff who contributed blood samples and data for this study.
Footnotes
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
GRANT SUPPORT
This work was supported by the National 863 High-Technology Research and Development Program (No. 2012AA02A519).
REFERENCES
- 1.Constantoulakis P, Filiou E, Rovina N, Chras G, Hamhougia A, Karabela S, Sotiriou A, Roussos C, Poulakis N. In vivo expression of innate immunity markers in patients with Mycobacterium tuberculosis infection. BMC infectious diseases. 2010;10:243. doi: 10.1186/1471-2334-10-243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bellamy R. Genetic susceptibility to tuberculosis. Clinics in chest medicine. 2005;26:233–246. vi. doi: 10.1016/j.ccm.2005.02.006. [DOI] [PubMed] [Google Scholar]
- 3.Casanova JL, Abel L. Genetic dissection of immunity to mycobacteria: the human model. Annual review of immunology. 2002;20:581–620. doi: 10.1146/annurev.immunol.20.081501.125851. [DOI] [PubMed] [Google Scholar]
- 4.Baghdadi JE, Orlova M, Alter A, Ranque B, Chentoufi M, Lazrak F, Archane MI, Casanova JL, Benslimane A, Schurr E, Abel L. An autosomal dominant major gene confers predisposition to pulmonary tuberculosis in adults. The Journal of experimental medicine. 2006;203:1679–1684. doi: 10.1084/jem.20060269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Casanova JL, Abel L. Inborn errors of immunity to infection: the rule rather than the exception. The Journal of experimental medicine. 2005;202:197–201. doi: 10.1084/jem.20050854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Pan H, Yan BS, Rojas M, Shebzukhov YV, Zhou H, Kobzik L, Higgins DE, Daly MJ, Bloom BR, Kramnik I. Ipr1 gene mediates innate immunity to tuberculosis. Nature. 2005;434:767–772. doi: 10.1038/nature03419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kramnik I. Genetic dissection of host resistance to Mycobacterium tuberculosis: the sst1 locus and the Ipr1 gene. Current topics in microbiology and immunology. 2008;321:123–148. doi: 10.1007/978-3-540-75203-5_6. [DOI] [PubMed] [Google Scholar]
- 8.Abhimanyu, Jha P, Jain A, Arora K, Bose M. Genetic association study suggests a role for SP110 variants in lymph node tuberculosis but not pulmonary tuberculosis in north Indians. Human immunology. 2011;72:576–580. doi: 10.1016/j.humimm.2011.03.014. [DOI] [PubMed] [Google Scholar]
- 9.Fox GJ, Sy DN, Nhung NV, Yu B, Ellis MK, Van Hung N, Cuong NK, Thi Lien L, Marks GB, Saunders BM, Britton WJ. Polymorphisms of SP110 are associated with both pulmonary and extra-pulmonary tuberculosis among the Vietnamese. PloS one. 2014;9:e99496. doi: 10.1371/journal.pone.0099496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jiang SY, Li LL, Yue J, Chen WZ, Yang C, Wan CL, He L, Cai L, Deng SL. The effects of SP110′s associated genes on fresh cavitary pulmonary tuberculosis in Han Chinese population. Clinical and experimental medicine. 2015 doi: 10.1007/s10238-015-0339-4. [DOI] [PubMed] [Google Scholar]
- 11.Babb C, Keet EH, van Helden PD, Hoal EG. SP110 polymorphisms are not associated with pulmonary tuberculosis in a South African population. Human genetics. 2007;121:521–522. doi: 10.1007/s00439-007-0335-1. [DOI] [PubMed] [Google Scholar]
- 12.Thye T, Browne EN, Chinbuah MA, Gyapong J, Osei I, Owusu-Dabo E, Niemann S, Rusch-Gerdes S, Horstmann RD, Meyer CG. No associations of human pulmonary tuberculosis with Sp110 variants. Journal of medical genetics. 2006;43:e32. doi: 10.1136/jmg.2005.037960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Bai Y, Zhang X, Katona I, Saporta MA, Shy ME, O'Malley HA, Isom LL, Suter U, Li J. Conduction block in PMP22 deficiency. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2010;30:600–608. doi: 10.1523/JNEUROSCI.4264-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Seitz HK, Stickel F. Molecular mechanisms of alcohol-mediated carcinogenesis. Nature Reviews Cancer. 2007;7:599–612. doi: 10.1038/nrc2191. [DOI] [PubMed] [Google Scholar]
- 15.Amici SA, Dunn WA, Jr, Notterpek L. Developmental abnormalities in the nerves of peripheral myelin protein 22-deficient mice. Journal of neuroscience research. 2007;85:238–249. doi: 10.1002/jnr.21118. [DOI] [PubMed] [Google Scholar]
- 16.Liang L, Zhao YL, Yue J, Liu JF, Han M, Wang H, Xiao H. Association of SP110 gene polymorphisms with susceptibility to tuberculosis in a Chinese population. Infection, genetics and evolution: journal of molecular epidemiology and evolutionary genetics in infectious diseases. 2011;11:934–939. doi: 10.1016/j.meegid.2011.02.017. [DOI] [PubMed] [Google Scholar]
- 17.Png E, Alisjahbana B, Sahiratmadja E, Marzuki S, Nelwan R, Adnan I, van de Vosse E, Hibberd M, van Crevel R, Ottenhoff TH, Seielstad M. Polymorphisms in SP110 are not associated with pulmonary tuberculosis in Indonesians. Infection, genetics and evolution: journal of molecular epidemiology and evolutionary genetics in infectious diseases. 2012;12:1319–1323. doi: 10.1016/j.meegid.2012.04.006. [DOI] [PubMed] [Google Scholar]
- 18.Tosh K, Campbell SJ, Fielding K, Sillah J, Bah B, Gustafson P, Manneh K, Lisse I, Sirugo G, Bennett S, Aaby P, McAdam KP, Bah-Sow O, Lienhardt C, Kramnik I, Hill AV. Variants in the SP110 gene are associated with genetic susceptibility to tuberculosis in West Africa. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:10364–10368. doi: 10.1073/pnas.0603340103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li J, Kleeff J, Esposito I, Kayed H, Felix K, Giese T, Buchler MW, Friess H. Expression analysis of PMP22/Gas3 in premalignant and malignant pancreatic lesions. The journal of histochemistry and cytochemistry: official journal of the Histochemistry Society. 2005;53:885–893. doi: 10.1369/jhc.4A6546.2005. [DOI] [PubMed] [Google Scholar]
- 20.Chackerian AA, Perera TV, Behar SM. Gamma interferon-producing CD4+ T lymphocytes in the lung correlate with resistance to infection with Mycobacterium tuberculosis. Infection and immunity. 2001;69:2666–2674. doi: 10.1128/IAI.69.4.2666-2674.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Perneger TV. What's wrong with Bonferroni adjustments. Bmj. 1998;316:1236–1238. doi: 10.1136/bmj.316.7139.1236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Szeszko JS, Healy B, Stevens H, Balabanova Y, Drobniewski F, Todd JA, Nejentsev S. Resequencing and association analysis of the SP110 gene in adult pulmonary tuberculosis. Human genetics. 2007;121:155–160. doi: 10.1007/s00439-006-0293-z. [DOI] [PubMed] [Google Scholar]
- 23.Jha P, Jain A, Arora K, Bose M. Genetic association study suggests a role for SP110 variants in lymph node tuberculosis but not pulmonary tuberculosis in north Indians. Human immunology. 2011;72:576–580. doi: 10.1016/j.humimm.2011.03.014. [DOI] [PubMed] [Google Scholar]
- 24.Schurz H, Daya M, Möller M, Hoal EG, Salie M. TLR1, 2, 4, 6 and 9 variants associated with tuberculosis susceptibility: a systematic review and meta-analysis. PloS one. 2015;10:e0139711. doi: 10.1371/journal.pone.0139711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yong Y, Lin H. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell research. 2005;15:97–98. doi: 10.1038/sj.cr.7290272. [DOI] [PubMed] [Google Scholar]