

Abstract
Osteogenesis and adipogenesis of bone marrow mesenchymal stem cells (BMSCs) are regarded as being of great importance in the regulation of bone remodeling. In this study, rat BMSCs were exposed to different levels of cyclic mechanical stress generated by liquid drops and cultured in general medium or adipogenic medium. Markers of osteogenic (Runx2 and Collagen I) and adipogenic (C/EBPα, PPARγ, and lipid droplets) differentiation were detected using Western blot and histological staining. The protein levels of members of the phosphatidylinositol 3-kinase (PI3K)/Akt/glycogen synthase kinase 3β (GSK-3β)/β-catenin signaling pathway were also examined. Results showed that small-magnitude stress significantly upregulated Runx2 and Collagen I and downregulated PPARγ and C/EBPα expression in BMSCs cultured in adipogenic medium, while large-magnitude stress reversed the effect when compared with unloading groups. The PI3K/Akt signaling pathway could be strongly activated by mechanical stimulation; however, large-magnitude stress led to decreased activation of the signaling pathway when compared with small-magnitude stress. Activation of β-catenin with LiCl led to increased expression of Runx2 and Collagen I and reduction of C/EBPα and PPARγ expression in BMSCs. Inhibition of PI3K/Akt signaling partially blocked the expression of β-catenin. Taken together, our results indicate that mechanical stress-regulated osteogenesis and adipogenesis of rat BMSCs are mediated, at least in part, by the PI3K/Akt/GSK-3β/β-catenin signaling pathway.
1. Introduction
Mesenchymal stem cells (MSCs) have multipotent differentiation capabilities and have been confirmed to differentiate into various lineages, including osteoblasts, adipocytes, chondrocytes, myocytes, and fibroblasts [1, 2]. Thus they hold great potential for regenerative medicine and tissue-engineering applications. It has been demonstrated that differentiation of MSCs into these various lineages is controlled by their particular biochemical microenvironment [2]. Additionally, mechanical stress has also been demonstrated to aid MSCs differentiation into various mature cells [3, 4]. Tension [5] and fluid shear stress [6, 7] have both been reported to promote osteogenic lineage commitment, while compression [8, 9] and hydrostatic pressure [10] increase both chondrogenic and osteogenic differentiation.
In bones, osteoblasts and adipocytes are derived from a common progenitor cell, the bone marrow MSCs (BMSCs) [11]. In particular, osteogenic differentiation of BMSCs is essential for the maintenance of bone quality and quantity. However, reduced osteogenic potential and relatively increased adipogenic differentiation are associated with various pathological conditions such as osteoporosis, resulting in reduced bone mass, increased bone fragility, and increased susceptibility to fracture [12]. Therefore, the commitment of BMSCs to the adipocyte and osteoblast lineages appears to be of great importance in the regulation of bone remodeling.
It has been widely accepted that physiological loading is beneficial in maintaining skeletal integrity and increasing bone strength [13], while osteoporosis or bone resorption is often observed under unloading conditions such as in bedridden patients, in conditions of microgravity, following fracture of a limb and in other conditions of skeletal unloading [14, 15]. Concentration of excessive mechanical stress often induces fatigue fractures and periprosthetic fractures. Physiological mechanical stress enhances the proliferation, differentiation, and extracellular matrix formation of osteoblasts [16, 17], while pathological mechanical loading has been reported to exert an apoptotic effect on osteoblasts [18–20], leading to different outcomes of bone metabolism and bone strength.
The commitment of MSCs to different lineages in response to mechanical stress results from different types, magnitudes, and/or frequencies of forces [21]. Intensive studies using different types of stress such as tensile stretch, fluid flow, compression, and cyclic hydrostatic pressure have been reported to induce bone formation and limit adipogenesis of MSCs, which occur as a concerted response [22]. However, much more remains to be discovered about the effect of different levels of mechanical stimulation on the adipogenic and osteogenic differentiation of MSCs.
Over decades of study, it has become more and more clear that the adipogenic differentiation and osteogenic differentiation of MSCs are competing and reciprocal [23]. Our previous research has demonstrated that the apoptotic or antiapoptotic effect of mechanical stress on the osteoblast depends on the levels of the stress loading on it [20]. Just as different apoptotic effects on osteoblasts are observed in response to different levels of mechanical stress, we hypothesized that osteogenic and adipogenic differentiation of MSCs may reverse with changes in the levels of mechanical loading. The purpose of the present study was to investigate the effects of different magnitudes of cyclic mechanical stress on the osteogenic and adipogenic differentiation of BMSCs and to clarify the underlying mechanism. Markers of osteogenic and adipogenic differentiation were observed in MSCs stimulated by cyclic compression stress generated by liquid drops, cultured in both general and adipogenic medium in vitro. Furthermore, changes in the phosphatidylinositol 3-kinase (PI3K)/Akt/glycogen synthase kinase 3β (GSK-3β)/β-catenin signaling pathway were investigated to illustrate the mechanism. Our present study thus provides a preliminary insight into the mechanical stress-induced changes of osteogenic and adipogenic differentiation in BMSCs and illustrates the role played by the PI3K/Akt/GSK-3β/β-catenin signaling pathway in the process.
2. Materials and Methods
2.1. Antibodies and Reagents
Antibodies against Runx2, C/EBPα, phosphor-Akt (p-Akt), Akt, GSK-3β, β-actin, and the appropriate anti-rabbit horseradish peroxidase- (HRP-) conjugated secondary antibody were purchased from Cell Signaling Technology, USA. Antibodies against Collagen I, PPARγ, β-catenin, CD29, CD34, CD44, and CD45 were purchased from Abcam, UK. Reagent Sources including LY294002 (PI3K/Akt inhibitor), LiCl, dexamethasone, 3-isobutyl-1-methylxanthine (IBMX), indomethacin, and insulin were purchased from Sigma, USA. The enhanced chemiluminescence (ECL) detection substrate was purchased from Thermo Fisher Scientific, USA.
2.2. Isolation, Expansion, and Identification of BMSCs
Animal study was approved by the animal research committee of PLA Second Military Medical University. Neonatal Sprague-Dawley rats (7 days) were used for the isolation of BMSCs. Bone marrow was isolated aseptically by flushing the femurs and tibias of rats and then suspended in DMEM/F12 supplemented with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin in a humidified atmosphere incubator containing 95% air and 5% CO2 at 37°C. After 4 days, nonadherent cells were removed and medium was changed. Cells were passaged as the first passage when they were 90–95% confluent by trypsinization and seeded at a density of approximately 104 cells/cm2 on round glasses (φ = 25 mm, Thickness = 0.17 mm) covered with polylysine. BMSCs phenotypes were confirmed by flow cytometry and analysis of cell surface molecules. Briefly, cells at passage one were washed in PBS and incubated with PE-labeled anti-CD29 (1 : 5), anti-CD34 (1 : 100) anti-CD44 (1 : 50), and FITC-labeled anti-CD45 (1 : 50) for 30 minutes. Then the cells were analyzed on a Flow Cytometer (BD Biosciences). Apoptotic cells were excluded from analysis using propidium iodide (PI).
2.3. Groups and Parameter Settings
According to absence or presence of different levels of stress intervention and different medium cells cultured in, the cells were divided into experimental and control groups, respectively. Stress parameters were set as follows: 0.15 Hz × 8 cm as small-magnitude and 0.6 Hz × 8 cm as large-magnitude for 30 min. The general medium was DMEM/F12 supplemented with 10% FBS and 1% penicillin-streptomycin, while the adipogenic medium consisted of basic high glucose DMEM supplemented with 1.0 μM dexamethasone, 0.5 mM IBMX, 200 μM indomethacin, 10 μM insulin, 10% FBS, and 1% penicillin-streptomycin.
2.4. Cyclic Mechanical Stress Application
Cells on glasses of experimental groups were exposed to cyclic mechanical stress when they were 90–95% confluent. Briefly, thin cover glass plates were placed over the confluent cell layers on the glasses. The cyclic mechanical stress was adjusted by drops of phosphate buffer solution (PBS) to the glass plates. Cells were subjected to different frequency (Hz) (controlled by drops per second) and height (cm) of mechanical stress (Figure 1). Control cells were covered with a thin glass plate without any liquid dropping on it.
Figure 1.
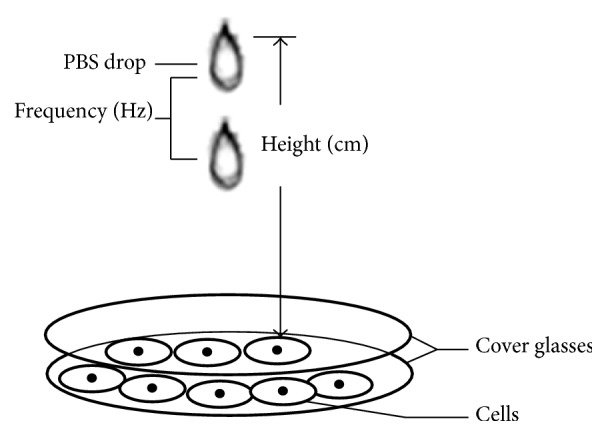
Diagrammatic representation for cyclic mechanical stress generated by PBS drops. Parameters of mechanical stress consisted of frequency (Hz) (controlled by drops per second) and height (cm) (perpendicular distance from drip nozzle of PBS to cover glass).
2.5. PI3K/Akt/GSK-3β/β-Catenin Signaling Modulation
For inhibition of PI3K/Akt signaling, cells were seeded on glasses as described and when they were 90–95% confluent, the medium was changed by DMEM/F12 with or without 10 μM LY294002 for 1 h followed by mechanical stimulation (0.15 Hz × 8 cm) for 30 min. Cells without stress intervention were used as controls. Then cells were collected and protein expression levels of signaling pathway (p-Akt, Akt, GSK-3β, and β-catenin) were measured by Western blot. For activation of β-catenin, cells were cultured with either 10 mM LiCl or 10 mM sodium chloride (NaCl) (control) in adipogenic medium for 7 days. Then expression levels of adipogenic and osteogenic marker proteins in BMSCs were examined by Western blot.
2.6. Western Blot
Total proteins were extracted from cells by using lysis buffer (Beyotime, Shanghai, China), according to the manufacturer's instructions. Proteins from each sample were separated using 10% SDS-PAGE gels and transferred to polyvinylidene fluoride (PVDF) membranes using an electroblotting apparatus (Bio-Rad, Hercules, CA, USA). After blocking in 5% nonfat milk/TBS-Tween 20 (TBST) solution at room temperature for 1 h, membranes were then incubated with primary antibodies specific for Runx2 (1 : 1000), PPARγ (1 : 1000), Collagen I (1 : 1000), C/EBPα (1 : 1000), p-Akt (1 : 1000), Akt (1 : 1000), GSK-3β (1 : 1000), β-catenin (1 : 1000), and β-actin(1 : 2000) at 4°C overnight. After washing with TBST, blots were then incubated at room temperature for 1 h with HRP-conjugated secondary antibodies diluted to 1 : 5000 in 5% nonfat milk/TBST. Protein bands were visualized using ECL reagents according to the manufacturer's instructions. The intensity values of each phosphorylated kinase were normalized to the corresponding total protein bands. Unless otherwise stated, β-actin was used as an internal control.
2.7. Histological Staining
Accumulation of lipid droplets was used to denote adipogenic differentiation of BMSCs. After fixation in 4% paraformaldehyde, cells were rinsed with 60% isopropanol, followed by freshly prepared Oil Red O solution (Sigma, 0.3% in isopropanol mixed 3 : 2 with deionized water).
2.8. Statistical Analysis
All data were presented as means ± standard deviation (SD). Statistical significance was determined using single-factor analysis of variance (ANOVA) followed by post hoc Bonferroni test to compare between groups. P < 0.05 was considered statistically significant.
3. Results
3.1. BMSCs Identification
BMSCs identification was done by the phenotypic analysis of the cells. Flow cytometric characterization analysis showed that the cells were homogenously positive for CD29 (97.3%) and CD44 (92.9%) and negative for CD34 (3.41%) and CD45 (1.11%) (Figure 2).
Figure 2.

Flow cytometric analysis of surface molecule expression in BMSCs. Cells were positive for the surface antigens CD29 (97.3%) and CD44 (92.9%) and negative for CD34 (3.41%) and CD45 (1.11%).
3.2. Effects of Mechanical Stress on BMSCs Differentiation
The adipogenesis of BMSCs was confirmed by Oil Red O staining of the droplets in cells. Significant development of lipid droplets was observed in BMSCs after exposure to adipogenic medium for 7 days compared with control groups. Application of small-magnitude mechanical stress reduced intracellular lipid droplets while large-magnitude mechanical stress led to an increasing of lipid droplets compared with unstressed groups (Figure 3(a)).
Figure 3.

Effects of different levels of mechanical stress on BMSCs differentiation. BMSCs were exposed to different levels of mechanical stress for 30 min and cultured in adipogenic medium or general medium for 7 days. (a) The adipogenesis of BMSCs was confirmed by Oil Red O staining. (A) No obvious intracellular lipid droplet formed in the control group; (B) a moderate amount of intracellular lipid droplet formed in the group of unloading + adipogenic medium; (C) a slight amount of intracellular lipid droplet formed in the small-magnitude stress + adipogenic medium group; (D) a substantial amount of intracellular lipid droplet formed in the large-magnitude stress + adipogenic medium group. Bars = 50 μm. (b) The expression levels of Runx2, Collagen I, C/EBPα, and PPARγ in BMSCs were examined by Western blot. (c) Quantitative analysis. ∗P < 0.05 versus control group; #P < 0.05 versus unloading + adipogenic medium group; &P < 0.05 versus small-magnitude stress + adipogenic medium group. Each value was presented as mean ± SD of three separate experiments and data of the treatment group was expressed as fold change versus that of control group (labeled as “1.00”). S represented small-magnitude stress; L represented large-magnitude stress. Cells cultured in general medium without mechanical loading as control.
The expressions of Runx2, Collagen I, C/EBPα, and PPARγ in BMSCs that were cultured for 7 days with or without mechanical stress were determined by Western blot. The results showed that compared with that of control, Runx2 and Collagen I were upregulated (P < 0.05) while C/EBPα and PPARγ were downregulated, respectively (P < 0.05) after BMSCs were cultured in adipogenic medium for 7 days (Figures 3(b) and 3(c)).
After culture in adipogenic medium for 7 days, small-magnitude stress stimulation led to a significant downregulation of C/EBPα and PPARγ and upregulation of Runx2 and Collagen I, respectively (P < 0.05) in BMSCs compared with unstressed groups. However, as the mechanical stress increased, the effect was reversed. Expressions of C/EBPα and PPARγ in BMSCs increased significantly (P < 0.05) which were even higher than that of unstressed groups. Additionally, large-magnitude stress resulted in decreased expressions of Runx2 and Collagen I when compared with small-magnitude stress groups and unstressed groups, respectively (P < 0.05) (Figures 3(b) and 3(c)).
3.3. Effect of Mechanical Stress on Activation of PI3K/Akt/GSK-3β/β-Catenin Signaling
The activation of PI3K/Akt/GSK-3β/β-catenin signaling was evaluated by Western blot analysis. The results showed that small-magnitude stress significantly upregulated the levels of p-Akt and β-catenin and inhibited the expression of GSK-3β compared with control cells (P < 0.05). In contrast, with the increasing of mechanical forces, levels of p-Akt (P < 0.05) and β-catenin decreased and GSK-3β (P < 0.05) increased, respectively (Figure 4). These results reflected that the manner of PI3K/Akt/GSK-3β/β-catenin signaling activation in BMSCs depended on the levels of mechanical stress loaded on them.
Figure 4.

Effects of mechanical stress on PI3K/Akt/GSK-3β/β-catenin signaling pathway. BMSCs were stimulated with different levels of mechanical stress for 30 min. (a) Expression levels of PI3K/Akt/GSK-3β/β-catenin signaling proteins were detected by Western blot. (b–d) Quantitative analysis. ∗P < 0.05 versus control group; #P < 0.05 versus small-magnitude stress group. Each value was presented as mean ± SD of three separate experiments and data of the treatment group was expressed as fold change versus that of control group (labeled as “1.00”). S represented small-magnitude stress; L represented large-magnitude stress.
3.4. β-Catenin Induces Osteogenesis and Inhibits Adipogenesis of BMSCs
Firstly, activation of β-catenin in BMSCs was investigated after treatment with LiCl. Western blot revealed that levels of β-catenin significantly increased after cells were treated with LiCl for 24 h (P < 0.05), suggesting that β-catenin could be activated by LiCl. To explore the influence of β-catenin on the differentiation of BMSCs, cells were cultured in adipogenic medium for a 7-day period in presence or absence of LiCl and adipogenic and osteogenic marker proteins in BMSCs were analyzed by Western blot. As shown in Figure 5, the results demonstrated that expressions of Runx2 and Collagen I increased with LiCl treatment (P < 0.05), accompanied by reduction of C/EBPα and PPARγ expressions in BMSCs (P < 0.05). In all experiments, NaCl acted as a negative control for LiCl, which had no effect on β-catenin levels or differentiation of BMSCs [24].
Figure 5.

Effects of activation of β-catenin on BMSCs differentiation. (a) Cells were incubated with or without 10 mM LiCl for 24 h. β-Catenin in BMSCs was detected by Western blot. Cells were cultured in adipogenic medium for 7 days with or without LiCl. Expression levels of Runx2, Collagen I, C/EBPα, and PPARγ were detected by Western blot. (b) Quantitative analysis. ∗P < 0.05 versus control group. Each value was presented as mean ± SD of three separate experiments and data of the treatment group was expressed as fold change versus that of control group (labeled as “1.00”).
3.5. β-Catenin-Regulated Different Differentiation in BMSCs Is PI3K/Akt-Dependent
Pretreatment with 10 μM LY294002 significantly blocked the mechanical stress-induced phosphorylation of Akt compared to the stress loaded cells without inhibitor (P < 0.05). Furthermore, inhibition of PI3K/Akt signaling partially blocked the expression of β-catenin by inactivation of GSK-3β (P < 0.05) under mechanical stress. However, LY294002 itself did not necessarily lead to any significant changes of p-Akt, β-catenin, and GSK-3β in the absence of mechanical stimulation (Figure 6). Taken together, the above findings indicated that mechanical stress regulated β-catenin via a PI3K/Akt-dependent signaling pathway.
Figure 6.

Activation of PI3K/Akt/GSK-3β/β-catenin signaling by the treatment with LY294002 and mechanical stress. MSCs were exposed to mechanical stress (0.15 Hz × 8 cm) for 30 min in the presence or absence of LY294002. (a) Western blot analysis of the protein levels of p-Akt, GSK-3β, and β-catenin. (b–d) Quantitative analysis. ∗P < 0.05 versus control group; #P < 0.05 versus stress without inhibitor group. Each value was presented as mean ± SD of three separate experiments and data of the treatment group was expressed as fold change versus that of control group (labeled as “1.00”).
4. Discussion
Although factors such as magnitude, frequency, and application time varied between studies, different types of mechanical stimuli, including tension [25–27], compression [8, 28], fluid shear stress [6], and hydrostatic pressure [29] have been confirmed to induce osteogenic differentiation with or without inhibition of adipogenesis of MSCs. However, MSCs experiencing tension at high levels exhibit reduced expression of osteogenic markers such as Runx2, alkaline phosphatase (ALP), and Collagen I [30]. In previous studies, mechanical loading was reported to inhibit the adipogenic commitment of MSCs. Findings of Yanagisawa et al. demonstrated that a high compressive force converts the differentiation pathway of C2C12 cells into that of the adipogenic lineage by upregulating the expression of PPARγ [28]. However, this interesting phenomenon was just reported without discussion of any mechanism and to date little information is available regarding the effect of mechanical stress on the enhanced adipogenesis of MSCs. The results of the present study confirmed the hypothesis that the osteogenic and adipogenic features of BMSCs changed as the levels of mechanical stress increased and the effect was partially mediated by the PI3K/Akt/GSK-3β/β-catenin signaling pathway.
Adipogenesis is driven by a complex signaling pathway consisting of several key transcription factors including PPARγ and C/EBPα [11]. PPARγ is regarded as the central regulator of adipogenesis because no other factors have yet been reported to induce adipogenesis without it [31]. Several critical transcription factors such as Runx2, Osterix, and DLX2 play important roles in osteogenesis [23]. Of these, Runx2 is essential for the commitment of MSCs to the osteoblast lineage as Runx2 controls other osteoblast-related genes such as Osterix and Collagen I [32] as well as autoregulating itself [33]. Therefore, we investigated the commitment of MSCs to the osteoblast and adipocyte lineages by detecting expressions of PPARγ, C/EBPα, Runx2, and Collagen I in the present study. In monolayer culture, BMSCs tend to differentiate into osteoblasts spontaneously instead of adipocytes when cultured in general medium. In order to investigate the effect of mechanical loading on the adipogenic differentiation of BMSCs, cells were cultured in adipogenic medium after mechanical application. By examining the specific markers, we demonstrated that large-magnitude mechanical loading increased adipogenesis and inhibited osteogenesis of MSCs, which was in accord with the findings of Yanagisawa et al. [28].
Cytoskeletal and focal adhesion have been reported to play an important role in mechanotransduction and to contribute to MSCs differentiation [1]. Cellular tension is generated through the interface of the external cellular environment with the internal actin cytoskeleton via integrins [34, 35]. Stimulation such as mechanical stress activates integrins, which results in phosphorylation of focal adhesion kinase and activation of a series of signaling pathways including PI3K/Akt, mitogen-activated protein kinases (MAPKs), protein kinase C, and GTPases of the Rho family [36]. Of the several signaling pathways, the PI3K/Akt pathway attracted our attention since we have previously examined the response of this pathway to different levels of cyclic mechanical stress and demonstrated that Akt is activated by small-magnitude mechanical stress, while phosphorylation of Akt is significantly reduced when large-magnitude mechanical stress is applied [20]. In the present report, we chose to subject the cells to two representative levels of mechanical stress, 0.15 Hz × 8 cm as small-magnitude and 0.6 Hz × 8 cm as large-magnitude. Our results showed the same trend of phosphorylation of Akt as our previous study.
The differentiation of MSCs is an intensely complex process, involving a large number of signaling pathways, cytokines, and transcription factors. Studies in recent years have demonstrated that a number of critical signaling pathways mediate MSCs differentiation, including transforming growth factor-beta (TGF-β)/bone morphogenic protein (BMP), MAPKs, wingless-type MMTV integration site (Wnt) signaling, Hedgehogs (Hh), Notch, and RhoA/ROCK pathways [22, 37, 38]. Wnt signaling is an important pathway that regulates osteogenesis. In the canonical Wnt pathway, β-catenin acts as a key transcriptional coactivator and transmits extracellular signals to the nucleus to activate target genes [39]. When the Wnt signaling pathway is inactivated, GSK-3β is active and β-catenin is constitutively phosphorylated by a destruction complex composed of Axin, APC, and GSK-3β, resulting in low cellular levels of β-catenin. Inactivation of GSK-3β induces the accumulation of cytosolic β-catenin and translocation to the nucleus to activate downstream target genes such as Runx2 and PPARγ [39–41]. In MSCs, increased osteogenesis and repression of adipogenesis are dependent on activation of β-catenin by inhibition of GSK-3β. Either GSK-3β inactivation or β-catenin activation block the ability of mechanical signals to induce adipogenesis [42]. It has been reported that inactivation of β-catenin in MSCs results in decreased osteogenic differentiation in vivo and in vitro [43]. Results of a study by Sen et al. showed that inhibition of GSK-3β is essential for mechanical inhibition of adipogenesis [44]. Our results revealed that upregulation of β-catenin enhanced osteogenesis, while inhibiting adipogenesis of MSCs, even under adipogenic culture conditions, which is in accordance with their studies.
The PI3K/Akt pathway is reported to be activated by multiple types of mechanical stress [16, 45], which can inhibit GSK-3β and activate β-catenin [46]. Our present study showed that the changes of GSK-3β and β-catenin under different levels of mechanical loading were in response to changes in Akt. Akt activation was responsible for the consequent GSK-3β inhibition, as the expression of GSK-3β increased significantly in the presence of PI3K/Akt inhibitor.
Taken together, these results suggest that osteogenic and adipogenic differentiation of BMSCs induced by mechanical stress is regulated by the PI3KAkt/GSK-3β/β-catenin signaling pathway. However, we hypothesized that the effect of changes from the osteogenic to the adipogenic differentiation of BMSCs when mechanical stress increased is partially mediated by the pathway. In fact, decreased activation of β-catenin by large-magnitude mechanical stress could indeed lead to increased adipogenic differentiation relative to osteogenesis, but the phenomenon that large-magnitude mechanical stress induced more adipogenesis and less osteogenesis than that in the control group indicated that there were some other signaling pathways participating in the process. Specifically, mechanical loading leads to activation of the MAPKs, resulting in increased Runx2 and subsequent expression of osteoblast-specific genes [47, 48]. Furthermore, it is reported that several pathways mediate osteogenic differentiation of MSCs, including activation of ERK, PI3K/Akt [44], and insulin-like growth factor 1 (IGF1) signaling [49]. Our future studies will focus on other signaling pathways such as MAPKs and TGF-β/BMP signaling and related downstream key factors that may mediate the regulation of stress-induced differentiation of MSCs, which will promote our understanding of the networks of signaling pathways involved in this process and thus make beneficial use of appropriate mechanical stress and avoid the deleterious effects of pathological forces.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (no. 81371974).
Competing Interests
The authors declare that they have no competing interests.
Authors' Contributions
Fanglong Song, Dawei Jiang, and Tianchen Wang contributed equally to this study.
References
- 1.Mathieu P. S., Loboa E. G. Cytoskeletal and focal adhesion influences on mesenchymal stem cell shape, mechanical properties, and differentiation down osteogenic, adipogenic, and chondrogenic pathways. Tissue Engineering—art B: Reviews. 2012;18(6):436–444. doi: 10.1089/ten.teb.2012.0014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Liu Z.-J., Zhuge Y., Velazquez O. C. Trafficking and differentiation of mesenchymal stem cells. Journal of Cellular Biochemistry. 2009;106(6):984–991. doi: 10.1002/jcb.22091. [DOI] [PubMed] [Google Scholar]
- 3.Discher D. E., Janmey P., Wang Y.-L. Tissue cells feel and respond to the stiffness of their substrate. Science. 2005;310(5751):1139–1143. doi: 10.1126/science.1116995. [DOI] [PubMed] [Google Scholar]
- 4.Vogel V., Sheetz M. Local force and geometry sensing regulate cell functions. Nature Reviews Molecular Cell Biology. 2006;7(4):265–275. doi: 10.1038/nrm1890. [DOI] [PubMed] [Google Scholar]
- 5.Haasper C., Jagodzinski M., Drescher M., et al. Cyclic strain induces FosB and initiates osteogenic differentiation of mesenchymal cells. Experimental and Toxicologic Pathology. 2008;59(6):355–363. doi: 10.1016/j.etp.2007.11.013. [DOI] [PubMed] [Google Scholar]
- 6.Arnsdorf E. J., Tummala P., Castillo A. B., Zhang F., Jacobs C. R. The epigenetic mechanism of mechanically induced osteogenic differentiation. Journal of Biomechanics. 2010;43(15):2881–2886. doi: 10.1016/j.jbiomech.2010.07.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Li Y. J., Batra N. N., You L., et al. Oscillatory fluid flow affects human marrow stromal cell proliferation and differentiation. Journal of Orthopaedic Research. 2004;22(6):1283–1289. doi: 10.1016/j.orthres.2004.04.002. [DOI] [PubMed] [Google Scholar]
- 8.Jagodzinski M., Breitbart A., Wehmeier M., et al. Influence of perfusion and cyclic compression on proliferation and differentiation of bone marrow stromal cells in 3-dimensional culture. Journal of Biomechanics. 2008;41(9):1885–1891. doi: 10.1016/j.jbiomech.2008.04.001. [DOI] [PubMed] [Google Scholar]
- 9.Pelaez D., Arita N., Cheung H. S. Extracellular signal-regulated kinase (ERK) dictates osteogenic and/or chondrogenic lineage commitment of mesenchymal stem cells under dynamic compression. Biochemical and Biophysical Research Communications. 2012;417(4):1286–1291. doi: 10.1016/j.bbrc.2011.12.131. [DOI] [PubMed] [Google Scholar]
- 10.Wagner D. R., Lindsey D. P., Li K. W., et al. Hydrostatic pressure enhances chondrogenic differentiation of human bone marrow stromal cells in osteochondrogenic medium. Annals of Biomedical Engineering. 2008;36(5):813–820. doi: 10.1007/s10439-008-9448-5. [DOI] [PubMed] [Google Scholar]
- 11.Chen Q., Shou P., Zheng C., et al. Fate decision of mesenchymal stem cells: adipocytes or osteoblasts? Cell Death and Differentiation. 2016;23(7):1128–1139. doi: 10.1038/cdd.2015.168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Harvey N., Dennison E., Cooper C. Osteoporosis: impact on health and economics. Nature Reviews Rheumatology. 2010;6(2):99–105. doi: 10.1038/nrrheum.2009.260. [DOI] [PubMed] [Google Scholar]
- 13.Frost H. M. A 2003 update of bone physiology and Wolff's law for clinicians. Angle Orthodontist. 2004;74(1):3–15. doi: 10.1043/0003-3219(2004)074<0003:AUOBPA>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- 14.Lafage-Proust M.-H., Collet P., Dubost J. M., Laroche N., Alexandre C., Vico L. Space-related bone mineral redistribution and lack of bone mass recovery after reambulation in young rats. American Journal of Physiology. 1998;274(2, part 2):R324–R334. doi: 10.1152/ajpregu.1998.274.2.R324. [DOI] [PubMed] [Google Scholar]
- 15.Dehority W., Halloran B. P., Bikle D. D., et al. Bone and hormonal changes induced by skeletal unloading in the mature male rat. American Journal of Physiology—Endocrinology and Metabolism. 1999;276(1):E62–E69. doi: 10.1152/ajpendo.1999.276.1.e62. [DOI] [PubMed] [Google Scholar]
- 16.Bin G., Bo Z., Jing W., et al. Fluid shear stress suppresses TNF-α-induced apoptosis in MC3T3-E1 cells: involvement of ERK5-AKT-FoxO3a-Bim/FasL signaling pathways. Experimental Cell Research. 2016;343(2):208–217. doi: 10.1016/j.yexcr.2016.03.014. [DOI] [PubMed] [Google Scholar]
- 17.Yan Y.-X., Gong Y.-W., Guo Y., et al. Mechanical strain regulates osteoblast proliferation through integrin-mediated ERK activation. PLoS ONE. 2012;7(4) doi: 10.1371/journal.pone.0035709.e35709 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Matsui H., Fukuno N., Kanda Y., et al. The expression of Fn14 via mechanical stress-activated JNK contributes to apoptosis induction in osteoblasts. Journal of Biological Chemistry. 2014;289(10):6438–6450. doi: 10.1074/jbc.M113.536300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang L., Pan J., Wang T., Song M., Chen W. Pathological cyclic strain-induced apoptosis in human periodontal ligament cells through the RhoGDIα/caspase-3/PARP pathway. PLoS ONE. 2013;8(10) doi: 10.1371/journal.pone.0075973.e75973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Song F., Wang Y., Jiang D., et al. Cyclic compressive stress regulates apoptosis in rat osteoblasts: involvement of PI3K/Akt and JNK MAPK signaling pathways. PLOS ONE. 2016;11(11) doi: 10.1371/journal.pone.0165845.e0165845 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Maul T. M., Chew D. W., Nieponice A., Vorp D. A. Mechanical stimuli differentially control stem cell behavior: morphology, proliferation, and differentiation. Biomechanics and Modeling in Mechanobiology. 2011;10(6):939–953. doi: 10.1007/s10237-010-0285-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Chen J. C., Jacobs C. R. Mechanically induced osteogenic lineage commitment of stem cells. Stem Cell Research and Therapy. 2013;4(5, article 107) doi: 10.1186/scrt318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kokabu S., Lowery J. W., Jimi E. Cell fate and differentiation of bone marrow mesenchymal stem cells. Stem Cells International. 2016;2016:7. doi: 10.1155/2016/3753581.3753581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang J., Liu B., Gu S., Liang J. Effects of Wnt/β-catenin signalling on proliferation and differentiation of apical papilla stem cells. Cell Proliferation. 2012;45(2):121–131. doi: 10.1111/j.1365-2184.2012.00806.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Huang S.-C., Wu T.-C., Yu H.-C., et al. Mechanical strain modulates age-related changes in the proliferation and differentiation of mouse adipose-derived stromal cells. BMC Cell Biology. 2010;11, article 18 doi: 10.1186/1471-2121-11-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Li R., Liang L., Dou Y., et al. Mechanical strain regulates osteogenic and adipogenic differentiation of bone marrow mesenchymal stem cells. BioMed Research International. 2015;2015:10. doi: 10.1155/2015/873251.873251 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Li R., Liang L., Dou Y., et al. Mechanical stretch inhibits mesenchymal stem cell adipogenic differentiation through TGFβ1/Smad2 signaling. Journal of Biomechanics. 2015;48(13):3665–3671. doi: 10.1016/j.jbiomech.2015.08.013. [DOI] [PubMed] [Google Scholar]
- 28.Yanagisawa M., Suzuki N., Mitsui N., Koyama Y., Otsuka K., Shimizu N. Effects of compressive force on the differentiation of pluripotent mesenchymal cells. Life Sciences. 2007;81(5):405–412. doi: 10.1016/j.lfs.2007.06.004. [DOI] [PubMed] [Google Scholar]
- 29.Liu J., Zhao Z., Li J., et al. Hydrostatic pressures promote initial osteodifferentiation with ERK1/2 not p38 MAPK signaling involved. Journal of Cellular Biochemistry. 2009;107(2):224–232. doi: 10.1002/jcb.22118. [DOI] [PubMed] [Google Scholar]
- 30.Jagodzinski M., Drescher M., Zeichen J., et al. Effects of cyclic longitudinal mechanical strain and dexamethasone on osteogenic differentiation of human bone marrow stromal cells. European Cells & Materials. 2004;7:35–41. doi: 10.22203/ecm.v007a04. [DOI] [PubMed] [Google Scholar]
- 31.Tontonoz P., Hu E., Spiegelman B. M. Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor. Cell. 1994;79(7):1147–1156. doi: 10.1016/0092-8674(94)90006-x. [DOI] [PubMed] [Google Scholar]
- 32.Stein G. S., Lian J. B., van Wijnen A. J., et al. Runx2 control of organization, assembly and activity of the regulatory machinery for skeletal gene expression. Oncogene. 2004;23(24):4315–4329. doi: 10.1038/sj.onc.1207676. [DOI] [PubMed] [Google Scholar]
- 33.Drissi H., Luc Q., Shakoori R., et al. Transcriptional autoregulation of the bone related CBFA1/RUNX2 gene. Journal of Cellular Physiology. 2000;184(3):341–350. doi: 10.1002/1097-4652(200009)184:3<341::aid-jcp8>3.0.co;2-z. [DOI] [PubMed] [Google Scholar]
- 34.Shemesh T., Geiger B., Bershadsky A. D., Kozlov M. M. Focal adhesions as mechanosensors: a physical mechanism. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(35):12383–12388. doi: 10.1073/pnas.0500254102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Roca-Cusachs P., del Rio A., Puklin-Faucher E., Gauthier N. C., Biais N., Sheetz M. P. Integrin-dependent force transmission to the extracellular matrix by α-actinin triggers adhesion maturation. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(15):E1361–E1370. doi: 10.1073/pnas.1220723110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Marie P. J., Haÿ E., Saidak Z. Integrin and cadherin signaling in bone: role and potential therapeutic targets. Trends in Endocrinology and Metabolism. 2014;25(11):567–575. doi: 10.1016/j.tem.2014.06.009. [DOI] [PubMed] [Google Scholar]
- 37.Kelly D. J., Jacobs C. R. The role of mechanical signals in regulating chondrogenesis and osteogenesis of mesenchymal stem cells. Birth Defects Research Part C: Embryo Today: Reviews. 2010;90(1):75–85. doi: 10.1002/bdrc.20173. [DOI] [PubMed] [Google Scholar]
- 38.Wang Y.-K., Yu X., Cohen D. M., et al. Bone morphogenetic protein-2-induced signaling and osteogenesis is regulated by cell shape, RhoA/ROCK, and cytoskeletal tension. Stem Cells and Development. 2012;21(7):1176–1186. doi: 10.1089/scd.2011.0293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kim W., Kim M., Jho E.-H. Wnt/β-catenin signalling: from plasma membrane to nucleus. Biochemical Journal. 2013;450(1):9–21. doi: 10.1042/bj20121284. [DOI] [PubMed] [Google Scholar]
- 40.MacDonald B. T., Tamai K., He X. Wnt/β-catenin signaling: components, mechanisms, and diseases. Developmental Cell. 2009;17(1):9–26. doi: 10.1016/j.devcel.2009.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Saidak Z., LeHenaff C., Azzi S., et al. Wnt/β-catenin signaling mediates osteoblast differentiation triggered by peptide-induced α5β1 integrin priming in mesenchymal skeletal cells. Journal of Biological Chemistry. 2015;290(11):6903–6912. doi: 10.1074/jbc.M114.621219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Sen B., Styner M., Xie Z., Case N., Rubin C. T., Rubin J. Mechanical loading regulates NFATc1 and β-catenin signaling through a GSK3β control node. Journal of Biological Chemistry. 2009;284(50):34607–34617. doi: 10.1074/jbc.M109.039453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Day T. F., Guo X., Garrett-Beal L., Yang Y. Wnt/β-catenin signaling in mesenchymal progenitors controls osteoblast and chondrocyte differentiation during vertebrate skeletogenesis. Developmental Cell. 2005;8(5):739–750. doi: 10.1016/j.devcel.2005.03.016. [DOI] [PubMed] [Google Scholar]
- 44.Sen B., Guilluy C., Xie Z., et al. Mechanically induced focal adhesion assembly amplifies anti-adipogenic pathways in mesenchymal stem cells. Stem Cells. 2011;29(11):1829–1836. doi: 10.1002/stem.732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pavalko F. M., Gerard R. L., Ponik S. M., Gallagher P. J., Jin Y., Norvell S. M. Fluid shear stress inhibits TNF-α-induced apoptosis in osteoblasts: a role for fluid shear stress-induced activation of PI3-kinase and inhibition of caspase-3. Journal of Cellular Physiology. 2003;194(2):194–205. doi: 10.1002/jcp.10221. [DOI] [PubMed] [Google Scholar]
- 46.Case N., Ma M., Sen B., Xie Z., Gross T. S., Rubin J. β-Catenin levels influence rapid mechanical responses in osteoblasts. The Journal of Biological Chemistry. 2008;283(43):29196–29205. doi: 10.1074/jbc.m801907200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ge C., Xiao G., Jiang D., Franceschi R. T. Critical role of the extracellular signal-regulated kinase-MAPK pathway in osteoblast differentiation and skeletal development. Journal of Cell Biology. 2007;176(5):709–718. doi: 10.1083/jcb.200610046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Xiao G., Jiang D., Thomas P., et al. MAPK pathways activate and phosphorylate the osteoblast-specific transcription factor, Cbfa1. Journal of Biological Chemistry. 2000;275(6):4453–4459. doi: 10.1074/jbc.275.6.4453. [DOI] [PubMed] [Google Scholar]
- 49.Tahimic C. G. T., Long R. K., Kubota T., et al. Regulation of ligand and shear stress-induced insulin-like growth factor 1 (IGF1) signaling by the integrin pathway. Journal of Biological Chemistry. 2016;291(15):8140–8149. doi: 10.1074/jbc.M115.693598. [DOI] [PMC free article] [PubMed] [Google Scholar]