

Abstract
Background
Ischemia and the following reperfusion damage are critical mechanisms of spinal cord injury. Statins have been reported to decrease ischemia–reperfusion injury in many organs including the spinal cord. Anti-oxidative effect is one of the main protective mechanisms of statin against neuronal death and cytotoxicity. We hypothesized that statins’ anti-oxidative property would yield neuroprotective effects on spinal cord ischemia–reperfusion injury
Methods
Primary cultured spinal cord motor neurons were isolated from Sprague–Dawley rat fetuses. Ischemia–reperfusion injury model was induced by 60 min of oxygen and glucose deprivation (OGD) and 24 h of reoxygenation. Healthy and OGD cells were treated with simvastatin at concentrations of 0.1, 1, and 10 μM for 24 h. Cell viability was assessed using water-soluble tetrazolium salt (WST)-8, cytotoxicity with LDH, and production of free radicals with DCFDA (2′,7′-dichlorofluorescein diacetate).
Results
OGD reduced neuronal viability compared to normoxic control by 35.3%; however, 0.1–10 μM of simvastatin treatment following OGD improved cell survival. OGD increased LDH release up to 214%; however, simvastatin treatment attenuated its cytotoxicity at concentrations of 0.1–10 μM (p < 0.001 and p = 0.001). Simvastatin also reduced deteriorated morphological changes of motor neurons following OGD. Oxidative stress was reduced by simvastatin (0.1–10 μM) compared to untreated cells exposed to OGD (p < 0.001).
Conclusions
Simvastatin effectively reduced spinal cord neuronal death and cytotoxicity against ischemia–reperfusion injury, probably via modification of oxidative stress.
Keywords: Ischemia–reperfusion injury, Neuroprotection, Oxidative stress, Oxygen–glucose deprivation, Simvastatin, Spinal cord injury
Background
Spinal cord injury (SCI) can lead to devastating complications, including permanent neurological damage [1–3]. In acute traumatic spinal cord injury, ischemia and the following reperfusion play a critical role in primary mechanical and secondary pathophysiological mechanisms [4–6]. After the initial rapid compression and trauma, spinal cord ischemia occurs via various mechanisms, including direct injury to the microvasculature, reduced spinal cord blood flow, and disrupted spinal cord autoregulation [5]. In the following passages, restoration of vascular perfusion and surgical intervention are an essential part of the treatment approach to avoid persistent compression; however, reperfusion per se can cause further damage [7–9].
Spinal cord ischemia–reperfusion damage is also an important cause of postoperative neurological deficits following decompression surgery, which are rare but very serious. For example, 1.5–6.3% of patients with cervical spondylotic myelopathy suffer postoperative delayed paraplegia related to ischemia–reperfusion injury of the spinal cord [7–9]. In such cases, neuroprotection against ischemia–reperfusion is crucial to prevent spinal cord injury.
Statins, 3-hydroxy-3-methyl-glutaryl-coenzyme A (HMG-CoA) reductase inhibitors, have been shown to minimize the severity of ischemia–reperfusion injury in many organs including the brain, heart, kidney, and lung [10–14]. Statins attenuate neuronal injury and promote neurologic recovery after cerebral ischemia in experimental animal models and in vitro cellular models [15–18]. Statins are frequently used as cholesterol-lowering agents, but their protective effect against ischemia depends on other actions as well [19], including modification of oxidative stress [16, 20–22], anti-inflammatory effects, and immunomodulation [15, 18].
Statins have been repeatedly reported to be neuroprotective against spinal cord injury, demonstrating neurologic and histopathologic improvements [23–27]. Especially simvastatin, since it readily crosses the blood–spinal cord barrier, could be widely used to treat spinal cord injuries in clinical practice [24]. As yet, the underlying mechanism has not been fully studied. In models of cerebral ischemia, simvastatin attenuated neuronal death by reducing the production and toxicity of oxidative stress-related markers [28, 29]. However, statins’ beneficial antioxidant properties in spinal cord neurons have not yet been investigated.
In this study, we sought the efficacy of simvastatin in attenuation of SCI-induced pathology. We first demonstrated that ischemia–reperfusion injury elicits motor neuron death and cytotoxicity in this model of SCI, and then investigated whether simvastatin treatment recovers those deteriorations of spinal cord neurons against oxidative stress as its neuroprotective mechanism of action.
Methods
Primary culture of spinal cord neuron
The animal procedures were carried out in Seoul National University Bundang Hospital according to an approved animal research protocol (IRB number 63-13-034). Timed-pregnant Sprague–Dawley rats were obtained, and primary rat spinal cord neurons were isolated from embryonic day 14–15 rat fetuses using a previously described method [30]. Briefly, embryonic vertebral canals were opened, meninges and blood vessels were cleared away using sterile fine-tipped forceps, and the embryonic spinal cords were sliced into small pieces using a scalpel. After microdissection and trituration, the isolated cells were seeded on poly-l-lysine (200 μg/mL) (PLL) (Sigma-Aldrich, St. Louis, MO, USA) coated plates at a concentration of 105 cells/well and maintained in a 5% CO2 incubator at 37 °C. Cells were cultured in neurobasal medium (Gibco, Carlsbad, CA, USA) supplemented with 2% B27 supplement (Gibco) and 2 mM glutamine (Gibco). After 3 days in vitro (DIV), 5 μM cytosine-β-d-arabinofuranoside (AraC) (Sigma-Aldrich, St. Louis, MO, USA) was added into the medium to inhibit non-neuronal cell proliferation. One half of the culture medium was replaced by a fresh medium every 3 days.
Oxygen and glucose deprivation (OGD) followed by reoxygenation
OGD and reoxygenation were carried out in cultures after 7 DIV as described previously [31]. Briefly, on the seventh day, the original media was removed and replaced with glucose-free DMEM. The cultures were then transferred to an anaerobic incubator containing a mixture of 95% N2 and 5% CO2 at 37 °C. Several pilot experiments with various durations of OGD and reoxygenation indicated that 60 min of OGD and 24-h recovery led to sufficient injury for this study.
Treatment with simvastatin
Simvastatin (Sigma-Aldrich, St Louis, MO, USA) of 4 mg was dissolved in 100 μL of ethanol, with subsequent addition of 150 μL of 0.1 N NaOH. This solution was incubated at 50 °C for 2 h and then neutralized with HCl to pH 7. The resulting solution had a final volume of 1 mL with sterile phosphate-buffered saline [32]. To examine the toxicity of simvastatin on motor neurons, various concentrations of simvastatin (0.1–50 μM) were applied to healthy motor neurons for 24 h. In the main experiment, simvastatin was applied to the ischemia–reperfusion-injured motor neurons at concentrations of 0, 0.1, 1, and 10 μM for 24 h. Injured control cultures (OGD only) were given equal volume of phosphate-buffered vehicle. Each dose of simvastatin was applied from the start of the OGD and was maintained during the following 24-h reoxygenation period.
Determination of cellular viability
Cellular viability was assessed with tetrazolium salt reduction assay. This is a colorimetric method for determining the number of living cells. The viable cells containing NADH or NADPH can convert tetrazolium compound, WST-8, 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H tetrazolium, into formazan product that is soluble in tissue culture medium [33, 34]. According to the manufacturer’s instruction (cell counting kit 8 (CCK-8), Sigma-Aldrich, St. Louis, MO, USA), 10 μL of WST-8 solution was added to each well in 96-well plates. After 2-h incubation at 37 °C, absorbance at 450 nm was measured using a microplate reader. The quantity of formazan product is directly proportional to the number of living cells in the culture, and the results are expressed as the percentage of viable cells relative to untreated controls.
Determination of cytotoxicity
Lactate dehydrogenase (LDH) is a stable cytoplasmic enzyme that is released into the cell culture supernatant when the cytoplasmic membrane is ruptured [28, 35, 36]. LDH activity in the medium was determined using a colorimetric diagnostic kit (CytoTox96® Non-Radioactive Cytotoxicity Assay, Promega, Madison, WI, USA). In brief, 50-μL aliquots of the culture medium and 50 μL of reconstituted substrate were mixed and incubated for 30 min at 37 °C in the dark, after which the reaction was terminated with a stop solution (1 M HCl, 50 μL). The color intensity is directly proportional to the number of lysed cells and can be qualified by reading the absorbance at 490 nm. The percentage of cytotoxicity is determined by calculating the optical density at 490 nm (OD490nm) and subtracting from the absorbance value obtained in the background control.
Measurement of free radical production
The production of free radicals was assessed by using DCFDA (2′,7′-dichlorofluorescein diacetate), which is one of the most widely used techniques for direct measuring of cell redox state [37]. DCFDA is a fluorogenic dye for highly selective detection of hydroxyl, peroxyl, and any other intracellular reactive oxygen species (ROS) activity. DCFDA is diffused into cells and is deacetylated by cellular esterases into a non-fluorescent compound that is subsequently oxidized by ROS into DCF (2′,7′-dichlorofluorescein). DCF is a highly fluorescent compound and hence is detectable by fluorescence spectroscopy.
The DCFDA assay was performed according to the manufacturer’s instruction (DCFDA Cellular ROS Detection Kit, Abcam, Cambridge, UK). In brief, 10 μL of DCFDA was added to cells and incubated for 30 min. After being washed out with PBS, the intensity of fluorescence was examined by flow cytometry. Accumulation of the oxidized fluorescent derivate (DCF) in the cells was measured at emission and excitation wavelengths of 530 and 485 nm, respectively.
Statistical analysis
Data are expressed as mean ± SEM. Statistical comparisons between groups were done using t tests, and comparisons between three or more groups were performed by one-way ANOVA followed by Dunnett T3 post hoc comparisons. A p value less than 0.05 was considered significant. All statistical analyses were performed using SPSS 19.0 software for Windows (SPSS, Chicago, IL, USA).
Results
Motoneuron cultures and effects of simvastatin on healthy motor neuron
After 7 to 8 days in culture, cells showed neuronal morphological characteristics and an extensive meshwork of neurite outgrowth (Fig. 1a, b).
Fig. 1.
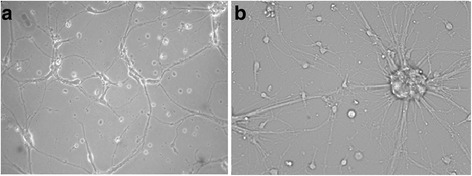
Typical morphology of healthy primary spinal cord neurons from rats. a The healthy neurons showed extensive neurite outgrowth and neurons with a dense meshwork on day 4. b Much denser meshwork can be seen on day 7 compared to (a)
Concentrations of simvastatin from 1 to 50 μM were applied to healthy control cells. Concentrations of 1, 5, and 10 μM did not affect the viability of cultured spinal neurons as indicated by WST-8. In contrast, at a concentration of 50 μM, cell viability was statistically decreased compared to the control cells (p = 0.002). Since the 50 μM concentration seemed to be toxic, we applied simvastatin at concentrations of 1 to 10 μM in further experiments (Fig. 2).
Fig. 2.
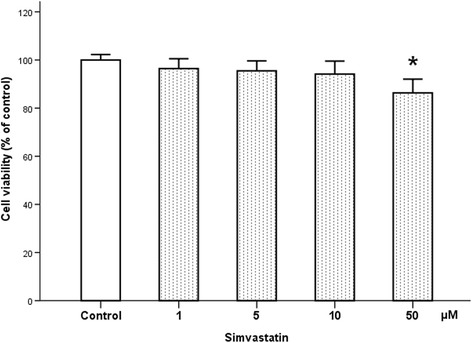
Effect of simvastatin on the viability of healthy motor neuron. Effect of simvastatin on the viability of motor neuron was assessed with WST-8 assay. There were no differences in the viability of healthy cells depending on the concentration of simvastatin. Simvastatin showed little or no effect on normal motoneuron ranges from 1 to 10 μM. However, at concentration of 50 μM, WST-8 statistically decreased compared to control cell. Data are expressed as the percentage of total WST-8 and represent the mean ± SEM. Asterisk denotes significant difference from control (n = 16; p < 0.05)
The effect of simvastatin on ischemia–reperfusion-induced cell death and cytotoxicity
Spinal cord ischemia–reperfusion injury was simulated by 60 min of OGD and 24 h of reoxygenation. This duration of injury resulted in moderate loss of the neural meshwork and morphological changes in the remaining cells (Fig. 3).
Fig. 3.
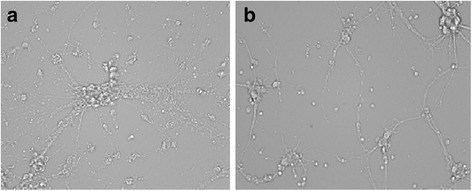
Morphological change in the motor neuron following ischemia–reperfusion injury with or without simvastatin. a Without simvastatin. Ischemia–reperfusion damage resulted in massive cell loss and destruction of neural networks. b With simvastatin. Treatment of simvastatin markedly increased neuronal survival after ischemia–reperfusion damage. Cells treated with simvastatin showed preserved morphological features of neurons with a denser meshwork of neurites when compared to (a)
The ischemia–reperfusion injury dramatically reduced cell survival (35% decrease) measured by WST-8 assay compared to the control culture. Simvastatin markedly ameliorated OGD/reoxygenation-evoked cell death at concentrations of 1 and 10 μM (p = 0.032 and p = 0.008, respectively) when compared to injured controls (Fig. 4). The protective effect was shown within a range of 0.1 to 10 μM. However, simvastatin did not fully restore viability of ischemia–reperfusion-injured motor neurons to that of healthy controls.
Fig. 4.
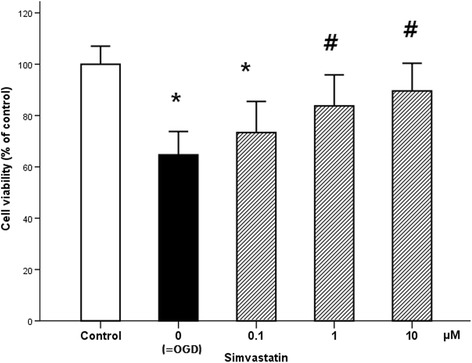
Effect of simvastatin on the viability of the motor neuron following ischemia–reperfusion injury. Effect of OGD and simvastatin on the viability of motor neuron was assessed with WST-8 assay. OGD resulted in marked reduction of cellular viability. Simvastatin, treated at the indicated concentrations, resulted in an increase in cell survival, particularly at 1 and 10 μM. Values are represented as means ± SEM; asterisk denotes significant difference from control. Number sign denotes significant difference from the OGD cells (n = 16; p < 0.05). OGD oxygen and glucose deprivation
The effects of simvastatin on the motor neuron damage are shown in Fig. 5. The ischemia–reperfusion damage resulted in cell death, as evidenced by an increase in LDH (214%). Within a range of 0.1 to 10 μM, simvastatin-treated motor neurons showed significantly lower values for LDH when compared to injured controls (p < 0.001 and p = 0.001, respectively), though cell death was not completely blocked.
Fig. 5.
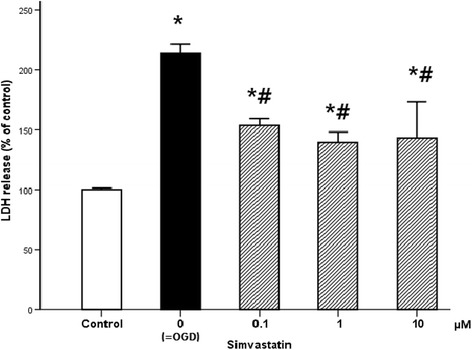
Effect of simvastatin on the cytotoxicity following ischemia–reperfusion injury. Cytotoxicity of OGD was assessed with LDH release. Cells were subjected to 60 min of OGD and 24 h of recovery time. LDH significantly increased after OGD compared to healthy cells. Simvastatin treatment (0.1, 1, and 10 μM) effectively attenuated LDH release. Values are represented as means ± SEM; asterisk denotes significant difference from control. Number sign denotes significant difference from the OGD cells (n = 16; p < 0.05). LDH lactate dehydrogenase, OGD oxygen and glucose deprivation
The effect of simvastatin on ischemia–reperfusion-induced oxidative stress
OGD and reoxygenation dramatically increased DCFDA value, implying higher levels of oxidative stress. Simvastatin effectively reduced the oxidative stress. All tested doses of simvastatin (0.1–10 μM) attenuated the rise in DCFDA level (p < 0.001 at all experimental concentrations), but not to a value as low as that of the healthy controls (Fig. 6).
Fig. 6.
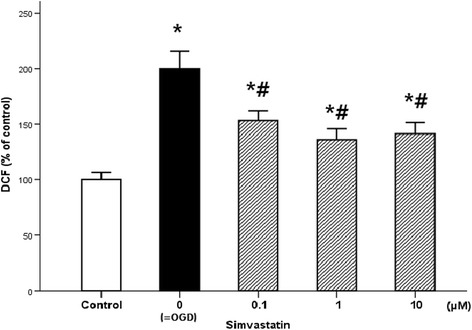
Effect of simvastatin on the motor neuron oxidative stress induced with ischemia–reperfusion. Oxidative stress of the motor neuron was assessed with DCFDA assay. OGD led to marked increase in DCF labeling of cells. Simvastatin treatment (0.1, 1, and 10 μM) effectively reduced DCF labeling following OGD. Values are represented as means ± SEM; asterisk denotes difference from control, and number sign denotes difference from OGD cells (n = 16; p < 0.05). OGD oxygen and glucose deprivation, DCFDA 2′,7′-dichlorofluorescein diacetate, DCF 2′,7′-dichlorofluorescein
Discussion
The present study demonstrated that motor neuron death, cytotoxicity, and oxidative stress induced by ischemia–reperfusion injury are significantly attenuated by simvastatin. To our knowledge, this is the first study to suggest the protective role of statins against ischemic injury of spinal cord neurons devising a primary motor neuron culture.
OGD and reoxygenation are well established and reliable neuronal cellular injury models that mimic changes that occur after ischemic insult in vivo [38–40]. Neurons are the cell type most sensitive to ischemic injury. Previous studies of cerebral ischemia–reperfusion have demonstrated that ischemia–reperfusion injury leads to obvious morphological neuronal changes, decreased cell survival, and dramatic increases in LDH release [38–40]. Our model of motor neuron ischemia–reperfusion yielded similar damages; motor neurons of primary cultured rat spinal cord resulted in a significant decrease in cell viability as evidenced by WST-8. After OGD, almost twice as much LDH had leaked out through the injured cell membrane as was leaked from healthy cells, implying increased neurotoxicity. Also OGD and reoxygenation dramatically increased DCFDA value, implying enhanced free radical generation in cells.
Statins reduce OGD and reoxygenation-induced neuronal injury [28, 40]. Statins reduce cerebral infarct volume [41, 42], improve perfusion deficits [43], and facilitate cognitive improvement [17] mostly using the experimental preclinical stroke models [39, 43]. Protective effect of statins against ischemic injury has also been reported in other organs, such as myocardial ischemia–reperfusion injury [11, 44], ischemic acute kidney injury [14], and intestinal ischemia–reperfusion injury [45]. We obtained similar results; simvastatin effectively attenuated ischemia–reperfusion-induced spinal cord motor neuron death. Quantitative analysis showed an increase in cell survival after simvastatin treatment at concentrations from 0.1 to 10 μM.
Similarly, ischemia–reperfusion-evoked LDH release was reduced by simvastatin in a dose-dependent manner, which supports the protective effect of simvastatin against ischemia–reperfusion-induced cytotoxicity. LDH leakage indicates cytotoxicity as a result of cell membrane disintegration [20]. The elevated LDH following OGD in our study significantly decreased with simvastatin treatment at concentrations of 0.1, 1, and 10 μM, indicating that it attenuates the cellular injury/death induced by ischemia. Our basic research performed at the cellular level demonstrated the safety margin of simvastatin in terms of motor neuron protection.
Ischemic injury leads to production of massive amounts of ROS that directly damage the main cellular constituents [18, 22]. In addition, reperfusion to an ischemic organ, a restoration of oxygen levels in hypoxic tissues, also stimulates ROS production [46, 47]. However, the CNS is extremely sensitive to oxidative stress due to delicate lipid layers of its cell membranes and low levels of antioxidant enzymes [48]. Oxidative stress responses in the CNS vary among different cell types. Neurons have relatively low antioxidant capacity and limited scope to upregulate it upon increased oxidative stress, so they are much more vulnerable to oxidative damage than other cells in CNS [49, 50]. Previous studies investigated the correlation between increased production of ROS and neuronal death following ischemia/hypoxia [51, 52]. Simvastatin has been reported to have pleiotropic effects, including reducing oxidative stress [18, 19, 53].
In our study, simvastatin significantly decreased free radical production induced by ischemia–reperfusion, implying that the drug modifies ischemia–reperfusion-induced oxidative stress. Numerous studies have reported that statins decrease ROS production in tissue and in vivo models and in cultured neurons derived from embryonic rat brain tissue [18, 20, 29] and following major vascular surgery [54] including thoracoabdominal aneurysm repair [55, 56] and decompression surgery of degenerative changes of the spine including cervical spondylotic myelopathy [57–59]. Our study results agree with these findings, showing that simvastatin reduces the production of DCFDA in spinal cord cells exposed to OGD and reoxygenation. DCFDA is one of the most widely used techniques for directly measuring the redox state of a cell [60].
Despite numerous reports of the neuroprotective effects of statins [19, 28, 29, 61–63], some investigators have reported that they are toxic to neurons in vitro [64, 65]. The toxicity versus protective effects of statins are dependent upon various factors, including the pharmacological characteristics of the individual statin agent, concentration of the drug, and cholesterol content of the neural cell used in the experiment [66]. Therefore, before the main experiment, we evaluated simvastatin’s toxicity to healthy motor neurons using concentrations ranging from 0.1 to 50 μM, based on the previous studies [28, 66–70]. Cellular viability, assessed by WST-8, was not affected at any experimental simvastatin dose from 0.1 to 10 μM. However, with a dose of 50 μM, the viability of the normal motor neuron was significantly reduced. This finding is consistent with that of previous reports and suggests possible cytotoxicity of high-dose statins [71]; that high dose was not applied in the main experiments.
The limitation of this study is that we did not determine the precise mechanism of action of simvastatin. Our capabilities did not allow us to examine whether the protective effects were due to direct reduction of cholesterol or whether indirect/nonspecific mechanisms of the statins modify cellular signaling. Further studies will help to elucidate the exact mechanism and the long-term effects of statins for clinical application. Second, there were no additional experiments to show antagonizing the neuroprotective effect of simvastatin, by coapplication of other drugs, such as mevalonate, one of the downstream products of HMG-CoA reductase. Third, it is significant that our spinal cord cells were cultured from embryonic 14–15 fetus because only embryonic cells can be used for extended culture. However, younger cells are more sensitive to neurotoxins, and therefore, the results should generalize to adults with caution.
Conclusions
In conclusion, simvastatin reduces ischemia and reperfusion-induced injury of spinal cord motor neurons through its antioxidant effects. Our results could lead to clinical use of simvastatin to treat this type of spinal cord injury.
Acknowledgements
The English in this document has been checked by at least two professional editors, both native speakers of English. For a certificate, please see http://www.textcheck.com/certificate/KFgcse
Funding
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (grant number: 800-2014-0159), Republic of Korea.
Availability of data and materials
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
Authors’ contributions
HMS performed all the experiments and prepared the initial draft of the manuscript. JYH Acquisition of data, Study concept and design. JHR Administrative and material support, Drafting of the manuscript. JHK Technical support, Study concept, Revision of the manuscript. SJP Analysis and interpretation of data. JWP Acquisition of data, Statistical analysis. SHH Data analysis, Supervision of all the studies. All authors reviewed, edited, and approved the final content of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable
Ethics approval
The animal procedures were carried out in Seoul National University Bundang Hospital according to an approved animal research protocol (IRB number 63-13-034).
Abbreviations
- DCFDA
2′,7′-dichlorofluorescein diacetate
- DIV
Days in vitro
- LDH
Lactate dehydrogenase
- OGD
Oxygen and glucose deprivation
- ROS
Reactive oxygen species
- SCI
Spinal cord injury
- WST
Water-soluble tetrazolium salt
Contributor Information
Hye-Min Sohn, Email: sophiesohn21@gmail.com.
Jin-Young Hwang, Email: mistyblue15@gmail.com.
Jung-Hee Ryu, Email: jinaryu74@gmail.com.
Jinhee Kim, Email: Anesing1@snu.ac.kr.
Seongjoo Park, Email: struka@snubh.org.
Jin-woo Park, Email: 72994@snubh.org.
Sung-Hee Han, Phone: 82-31-787-7499, Email: 21ambi@hanmail.net.
References
- 1.Tator CH, Edmonds VE. Acute spinal cord injury: analysis of epidemiologic factors. Can J Surg. 1979;22:575–8. [PubMed] [Google Scholar]
- 2.Celik M, Gokmen N, Erbayraktar S, Akhisaroglu M, Konakc S, Ulukus C, Genc S, Genc K, Sagiroglu E, Cerami A, Brines M. Erythropoietin prevents motor neuron apoptosis and neurologic disability in experimental spinal cord ischemic injury. Proc Natl Acad Sci U S A. 2002;99:2258–63. doi: 10.1073/pnas.042693799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sekhon LH, Fehlings MG. Epidemiology, demographics, and pathophysiology of acute spinal cord injury. Spine (Phila Pa 1976) 2001;26:S2–12. doi: 10.1097/00007632-200112151-00002. [DOI] [PubMed] [Google Scholar]
- 4.McVicar JP, Luce JM. Management of spinal cord injury in the critical care setting. Crit Care Clin. 1986;2:747–58. [PubMed] [Google Scholar]
- 5.Tator CH, Fehlings MG. Review of the secondary injury theory of acute spinal cord trauma with emphasis on vascular mechanisms. J Neurosurg. 1991;75:15–26. doi: 10.3171/jns.1991.75.1.0015. [DOI] [PubMed] [Google Scholar]
- 6.Aki T, Toya S. Experimental study on changes of the spinal-evoked potential and circulatory dynamics following spinal cord compression and decompression. Spine (Phila Pa 1976) 1984;9:800–9. doi: 10.1097/00007632-198411000-00006. [DOI] [PubMed] [Google Scholar]
- 7.Basaran R, Kaner T. C5 nerve root palsy following decompression of cervical spine with anterior versus posterior types of procedures in patients with cervical myelopathy. Eur Spine J. 2016;25:2050–9. doi: 10.1007/s00586-016-4567-4. [DOI] [PubMed] [Google Scholar]
- 8.Madhavan K, Chieng LO, Foong H, Wang MY. Surgical outcomes of elderly patients with cervical spondylotic myelopathy: a meta-analysis of studies reporting on 2868 patients. Neurosurg Focus. 2016;40:E13. doi: 10.3171/2016.3.FOCUS1657. [DOI] [PubMed] [Google Scholar]
- 9.Hasegawa K, Homma T, Chiba Y. Upper extremity palsy following cervical decompression surgery results from a transient spinal cord lesion. Spine (Phila Pa 1976) 2007;32:E197–202. doi: 10.1097/01.brs.0000257576.84646.49. [DOI] [PubMed] [Google Scholar]
- 10.Vaughan CJ, Delanty N. Neuroprotective properties of statins in cerebral ischemia and stroke. Stroke. 1999;30:1969–73. doi: 10.1161/01.STR.30.9.1969. [DOI] [PubMed] [Google Scholar]
- 11.Jones SP, Trocha SD, Lefer DJ. Pretreatment with simvastatin attenuates myocardial dysfunction after ischemia and chronic reperfusion. Arterioscler Thromb Vasc Biol. 2001;21:2059–64. doi: 10.1161/hq1201.099509. [DOI] [PubMed] [Google Scholar]
- 12.Naidu BV, Woolley SM, Farivar AS, Thomas R, Fraga C, Mulligan MS. Simvastatin ameliorates injury in an experimental model of lung ischemia-reperfusion. J Thorac Cardiovasc Surg. 2003;126:482–9. doi: 10.1016/S0022-5223(03)00699-8. [DOI] [PubMed] [Google Scholar]
- 13.Sironi L, Cimino M, Guerrini U, Calvio AM, Lodetti B, Asdente M, Balduini W, Paoletti R, Tremoli E. Treatment with statins after induction of focal ischemia in rats reduces the extent of brain damage. Arterioscler Thromb Vasc Biol. 2003;23:322–7. doi: 10.1161/01.ATV.0000044458.23905.3B. [DOI] [PubMed] [Google Scholar]
- 14.Devarajan P. Update on mechanisms of ischemic acute kidney injury. J Am Soc Nephrol. 2006;17:1503–20. doi: 10.1681/ASN.2006010017. [DOI] [PubMed] [Google Scholar]
- 15.Sutherland BA, Minnerup J, Balami JS, Arba F, Buchan AM, Kleinschnitz C. Neuroprotection for ischaemic stroke: translation from the bench to the bedside. Int J Stroke. 2012;7:407–18. doi: 10.1111/j.1747-4949.2012.00770.x. [DOI] [PubMed] [Google Scholar]
- 16.Hayashi T, Rani PJ, Fukatsu A, Matsui-Hirai H, Osawa M, Miyazaki A, Tsunekawa T, Kano-Hayashi H, Iguchi A, Sumi D, Ignarro LJ. A new HMG-CoA reductase inhibitor, pitavastatin remarkably retards the progression of high cholesterol induced atherosclerosis in rabbits. Atherosclerosis. 2004;176:255–63. doi: 10.1016/j.atherosclerosis.2003.12.034. [DOI] [PubMed] [Google Scholar]
- 17.Balduini W, Mazzoni E, Carloni S, De Simoni MG, Perego C, Sironi L, Cimino M. Prophylactic but not delayed administration of simvastatin protects against long-lasting cognitive and morphological consequences of neonatal hypoxic-ischemic brain injury, reduces interleukin-1beta and tumor necrosis factor-alpha mRNA induction, and does not affect endothelial nitric oxide synthase expression. Stroke. 2003;34:2007–12. doi: 10.1161/01.STR.0000080677.24419.88. [DOI] [PubMed] [Google Scholar]
- 18.Cimino M, Gelosa P, Gianella A, Nobili E, Tremoli E, Sironi L. Statins: multiple mechanisms of action in the ischemic brain. Neuroscientist. 2007;13:208–13. doi: 10.1177/1073858406297121. [DOI] [PubMed] [Google Scholar]
- 19.van der Most PJ, Dolga AM, Nijholt IM, Luiten PG, Eisel UL. Statins: mechanisms of neuroprotection. Prog Neurobiol. 2009;88:64–75. doi: 10.1016/j.pneurobio.2009.02.002. [DOI] [PubMed] [Google Scholar]
- 20.Hayashi T, Hamakawa K, Nagotani S, Jin G, Li F, Deguchi K, Sehara Y, Zhang H, Nagano I, Shoji M, Abe K. HMG CoA reductase inhibitors reduce ischemic brain injury of Wistar rats through decreasing oxidative stress on neurons. Brain Res. 2005;1037:52–8. doi: 10.1016/j.brainres.2004.12.051. [DOI] [PubMed] [Google Scholar]
- 21.Hong H, Zeng JS, Kreulen DL, Kaufman DI, Chen AF. Atorvastatin protects against cerebral infarction via inhibition of NADPH oxidase-derived superoxide in ischemic stroke. Am J Physiol Heart Circ Physiol. 2006;291:H2210–5. doi: 10.1152/ajpheart.01270.2005. [DOI] [PubMed] [Google Scholar]
- 22.Manzanero S, Santro T, Arumugam TV. Neuronal oxidative stress in acute ischemic stroke: sources and contribution to cell injury. Neurochem Int. 2013;62:712–8. doi: 10.1016/j.neuint.2012.11.009. [DOI] [PubMed] [Google Scholar]
- 23.Pannu R, Barbosa E, Singh AK, Singh I. Attenuation of acute inflammatory response by atorvastatin after spinal cord injury in rats. J Neurosci Res. 2005;79:340–50. doi: 10.1002/jnr.20345. [DOI] [PubMed] [Google Scholar]
- 24.Esposito E, Rinaldi B, Mazzon E, Donniacuo M, Impellizzeri D, Paterniti I, Capuano A, Bramanti P, Cuzzocrea S. Anti-inflammatory effect of simvastatin in an experimental model of spinal cord trauma: involvement of PPAR-alpha. J Neuroinflammation. 2012;9:81. doi: 10.1186/1742-2094-9-81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Anderson DK, Means ED, Waters TR, Spears CJ. Spinal cord energy metabolism following compression trauma to the feline spinal cord. J Neurosurg. 1980;53:375–80. doi: 10.3171/jns.1980.53.3.0375. [DOI] [PubMed] [Google Scholar]
- 26.Hwang J, Han JI, Han S. Effect of pretreatment with simvastatin on spinal cord ischemia-reperfusion injury in rats. J Cardiothorac Vasc Anesth. 2013;27:79–85. doi: 10.1053/j.jvca.2012.01.025. [DOI] [PubMed] [Google Scholar]
- 27.Saito T, Tsuchida M, Umehara S, Kohno T, Yamamoto H, Hayashi J. Reduction of spinal cord ischemia/reperfusion injury with simvastatin in rats. Anesth Analg. 2011;113:565–71. doi: 10.1213/ANE.0b013e318224ac35. [DOI] [PubMed] [Google Scholar]
- 28.Lim JH, Lee JC, Lee YH, Choi IY, Oh YK, Kim HS, Park JS, Kim WK. Simvastatin prevents oxygen and glucose deprivation/reoxygenation-induced death of cortical neurons by reducing the production and toxicity of 4-hydroxy-2E-nonenal. J Neurochem. 2006;97:140–50. doi: 10.1111/j.1471-4159.2006.03715.x. [DOI] [PubMed] [Google Scholar]
- 29.Domoki F, Kis B, Gaspar T, Snipes JA, Parks JS, Bari F, Busija DW. Rosuvastatin induces delayed preconditioning against oxygen-glucose deprivation in cultured cortical neurons. Am J Physiol Cell Physiol. 2009;296:C97–105. doi: 10.1152/ajpcell.00366.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Jiang XY, Fu SL, Nie BM, Li Y, Lin L, Yin L, Wang YX, Lu PH, Xu XM. Methods for isolating highly-enriched embryonic spinal cord neurons: a comparison between enzymatic and mechanical dissociations. J Neurosci Methods. 2006;158:13–8. doi: 10.1016/j.jneumeth.2006.05.014. [DOI] [PubMed] [Google Scholar]
- 31.Huang X, Sun J, Zhao T, Wu KW, Watanabe K, Xiao ZC, Zhu LL, Fan M. Loss of NB-3 aggravates cerebral ischemia by impairing neuron survival and neurite growth. Stroke. 2011;42:2910–6. doi: 10.1161/STROKEAHA.110.609560. [DOI] [PubMed] [Google Scholar]
- 32.Brown M, Hart C, Tawadros T, Ramani V, Sangar V, Lau M, Clarke N. The differential effects of statins on the metastatic behaviour of prostate cancer. Br J Cancer. 2012;106:1689–96. doi: 10.1038/bjc.2012.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Yamamoto Y, Mochida J, Sakai D, Nakai T, Nishimura K, Kawada H, Hotta T. Upregulation of the viability of nucleus pulposus cells by bone marrow-derived stromal cells: significance of direct cell-to-cell contact in coculture system. Spine (Phila Pa 1976) 2004;29:1508–14. doi: 10.1097/01.BRS.0000131416.90906.20. [DOI] [PubMed] [Google Scholar]
- 34.Deng X, Luan Q, Chen W, Wang Y, Wu M, Zhang H, Jiao Z. Nanosized zinc oxide particles induce neural stem cell apoptosis. Nanotechnology. 2009;20:115101. doi: 10.1088/0957-4484/20/11/115101. [DOI] [PubMed] [Google Scholar]
- 35.Johnson-Anuna LN, Eckert GP, Franke C, Igbavboa U, Muller WE, Wood WG. Simvastatin protects neurons from cytotoxicity by up-regulating Bcl-2 mRNA and protein. J Neurochem. 2007;101:77–86. doi: 10.1111/j.1471-4159.2006.04375.x. [DOI] [PubMed] [Google Scholar]
- 36.Burd JF, Usategui-Gomez M. A colorimetric assay for serum lactate dehydrogenase. Clin Chim Acta. 1973;46:223–7. doi: 10.1016/0009-8981(73)90174-5. [DOI] [PubMed] [Google Scholar]
- 37.Jayalakshmi K, Sairam M, Singh SB, Sharma SK, Ilavazhagan G, Banerjee PK. Neuroprotective effect of N-acetyl cysteine on hypoxia-induced oxidative stress in primary hippocampal culture. Brain Res. 2005;1046:97–104. doi: 10.1016/j.brainres.2005.03.054. [DOI] [PubMed] [Google Scholar]
- 38.Amarenco P, Moskowitz MA. The dynamics of statins: from event prevention to neuroprotection. Stroke. 2006;37:294–6. doi: 10.1161/01.STR.0000201856.90105.ab. [DOI] [PubMed] [Google Scholar]
- 39.Liu L, Liu H, Yang F, Chen G, Zhou H, Tang M, Zhang R, Dong Q. Tissue kallikrein protects cortical neurons against hypoxia/reoxygenation injury via the ERK1/2 pathway. Biochem Biophys Res Commun. 2011;407:283–7. doi: 10.1016/j.bbrc.2011.02.112. [DOI] [PubMed] [Google Scholar]
- 40.Zacco A, Togo J, Spence K, Ellis A, Lloyd D, Furlong S, Piser T. 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitors protect cortical neurons from excitotoxicity. J Neurosci. 2003;23:11104–11. doi: 10.1523/JNEUROSCI.23-35-11104.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nagotani S, Hayashi T, Sato K, Zhang W, Deguchi K, Nagano I, Shoji M, Abe K. Reduction of cerebral infarction in stroke-prone spontaneously hypertensive rats by statins associated with amelioration of oxidative stress. Stroke. 2005;36:670–2. doi: 10.1161/01.STR.0000155732.27333.3c. [DOI] [PubMed] [Google Scholar]
- 42.Lathia JD, Chigurupati S, Thundyil J, Selvaraj PK, Mughal MR, Woodruff TM, Chan SL, Karamyan VT, Mattson MP, Arumugam TV. Pivotal role for beta-1 integrin in neurovascular remodelling after ischemic stroke. Exp Neurol. 2010;221:107–14. doi: 10.1016/j.expneurol.2009.10.007. [DOI] [PubMed] [Google Scholar]
- 43.Shabanzadeh AP, Shuaib A, Wang CX. Simvastatin reduced ischemic brain injury and perfusion deficits in an embolic model of stroke. Brain Res. 2005;1042:1–5. doi: 10.1016/j.brainres.2005.01.105. [DOI] [PubMed] [Google Scholar]
- 44.Lefer AM, Scalia R, Lefer DJ. Vascular effects of HMG CoA-reductase inhibitors (statins) unrelated to cholesterol lowering: new concepts for cardiovascular disease. Cardiovasc Res. 2001;49:281–7. doi: 10.1016/S0008-6363(00)00247-9. [DOI] [PubMed] [Google Scholar]
- 45.Koh JY, Choi DW. Quantitative determination of glutamate mediated cortical neuronal injury in cell culture by lactate dehydrogenase efflux assay. J Neurosci Methods. 1987;20:83–90. doi: 10.1016/0165-0270(87)90041-0. [DOI] [PubMed] [Google Scholar]
- 46.Kalogeris T, Baines CP, Krenz M, Korthuis RJ. Cell biology of ischemia/reperfusion injury. Int Rev Cell Mol Biol. 2012;298:229–317. doi: 10.1016/B978-0-12-394309-5.00006-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Collard CD, Gelman S. Pathophysiology, clinical manifestations, and prevention of ischemia-reperfusion injury. Anesthesiology. 2001;94:1133–8. doi: 10.1097/00000542-200106000-00030. [DOI] [PubMed] [Google Scholar]
- 48.Marttila RJ, Roytta M, Lorentz H, Rinne UK. Oxygen toxicity protecting enzymes in the human brain. J Neural Transm. 1988;74:87–95. doi: 10.1007/BF01245142. [DOI] [PubMed] [Google Scholar]
- 49.Makar TK, Nedergaard M, Preuss A, Gelbard AS, Perumal AS, Cooper AJ. Vitamin E, ascorbate, glutathione, glutathione disulfide, and enzymes of glutathione metabolism in cultures of chick astrocytes and neurons: evidence that astrocytes play an important role in antioxidative processes in the brain. J Neurochem. 1994;62:45–53. doi: 10.1046/j.1471-4159.1994.62010045.x. [DOI] [PubMed] [Google Scholar]
- 50.Cafe C, Torri C, Bertorelli L, Tartara F, Tancioni F, Gaetani P, Rodriguez y Baena R, Marzatico F. Oxidative events in neuronal and glial cell-enriched fractions of rat cerebral cortex. Free Radic Biol Med. 1995;19:853–7. doi: 10.1016/0891-5849(95)00086-D. [DOI] [PubMed] [Google Scholar]
- 51.Kontos HA, George E. Brown memorial lecture. Oxygen radicals in cerebral vascular injury. Circ Res. 1985;57:508–16. doi: 10.1161/01.RES.57.4.508. [DOI] [PubMed] [Google Scholar]
- 52.Traystman RJ, Kirsch JR, Koehler RC. Oxygen radical mechanisms of brain injury following ischemia and reperfusion. J Appl Physiol (1985) 1991;71:1185–95. doi: 10.1152/jappl.1991.71.4.1185. [DOI] [PubMed] [Google Scholar]
- 53.Liao JK, Laufs U. Pleiotropic effects of statins. Annu Rev Pharmacol Toxicol. 2005;45:89–118. doi: 10.1146/annurev.pharmtox.45.120403.095748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Schouten O, Bax JJ, Dunkelgrun M, Feringa HH, van Urk H, Poldermans D. Statins for the prevention of perioperative cardiovascular complications in vascular surgery. J Vasc Surg. 2006;44:419–24. doi: 10.1016/j.jvs.2006.04.039. [DOI] [PubMed] [Google Scholar]
- 55.Moulakakis KG, Matoussevitch V, Borgonio A, Gawenda M, Brunkwall J. Evidence that statins protect renal function during endovascular repair of AAAs. Eur J Vasc Endovasc Surg. 2010;40:608–15. doi: 10.1016/j.ejvs.2010.05.006. [DOI] [PubMed] [Google Scholar]
- 56.Guzik B, Sagan A, Ludew D, Mrowiecki W, Chwala M, Bujak-Gizycka B, Filip G, Grudzien G, Kapelak B, Zmudka K, Mrowiecki T, Sadowski J, Korbut R, Guzik TJ. Mechanisms of oxidative stress in human aortic aneurysms--association with clinical risk factors for atherosclerosis and disease severity. Int J Cardiol. 2013;168:2389–96. doi: 10.1016/j.ijcard.2013.01.278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Karadimas SK, Gatzounis G, Fehlings MG. Pathobiology of cervical spondylotic myelopathy. Eur Spine J. 2015;24(Suppl 2):132–8. doi: 10.1007/s00586-014-3264-4. [DOI] [PubMed] [Google Scholar]
- 58.Hu S, Dong HL, Li YZ, Luo ZJ, Sun L, Yang QZ, Yang LF, Xiong L. Effects of remote ischemic preconditioning on biochemical markers and neurologic outcomes in patients undergoing elective cervical decompression surgery: a prospective randomized controlled trial. J Neurosurg Anesthesiol. 2010;22:46–52. doi: 10.1097/ANA.0b013e3181c572bd. [DOI] [PubMed] [Google Scholar]
- 59.Yaman O, Yaman B, Aydin F, Var A, Temiz C. Hyperbaric oxygen treatment in the experimental spinal cord injury model. Spine J. 2014;14:2184–94. doi: 10.1016/j.spinee.2014.02.013. [DOI] [PubMed] [Google Scholar]
- 60.Abramov AY, Scorziello A, Duchen MR. Three distinct mechanisms generate oxygen free radicals in neurons and contribute to cell death during anoxia and reoxygenation. J Neurosci. 2007;27:1129–38. doi: 10.1523/JNEUROSCI.4468-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Jung YS, Oh AY, Park HP, Hwang JW, Lim YJ, Jeon YT. Post-ischemic administration of pravastatin reduces neuronal injury by inhibiting Bax protein expression after transient forebrain ischemia in rats. Neurosci Lett. 2015;594:87–92. doi: 10.1016/j.neulet.2015.03.038. [DOI] [PubMed] [Google Scholar]
- 62.Bosel J, Gandor F, Harms C, Synowitz M, Harms U, Djoufack PC, Megow D, Dirnagl U, Hortnagl H, Fink KB, Endres M. Neuroprotective effects of atorvastatin against glutamate-induced excitotoxicity in primary cortical neurones. J Neurochem. 2005;92:1386–98. doi: 10.1111/j.1471-4159.2004.02980.x. [DOI] [PubMed] [Google Scholar]
- 63.Loane DJ, Faden AI. Neuroprotection for traumatic brain injury: translational challenges and emerging therapeutic strategies. Trends Pharmacol Sci. 2010;31:596–604. doi: 10.1016/j.tips.2010.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tanaka T, Tatsuno I, Uchida D, Moroo I, Morio H, Nakamura S, Noguchi Y, Yasuda T, Kitagawa M, Saito Y, Hirai A. Geranylgeranyl-pyrophosphate, an isoprenoid of mevalonate cascade, is a critical compound for rat primary cultured cortical neurons to protect the cell death induced by 3-hydroxy-3-methylglutaryl-CoA reductase inhibition. J Neurosci. 2000;20:2852–9. doi: 10.1523/JNEUROSCI.20-08-02852.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Marz P, Otten U, Miserez AR. Statins induce differentiation and cell death in neurons and astroglia. Glia. 2007;55:1–12. doi: 10.1002/glia.20422. [DOI] [PubMed] [Google Scholar]
- 66.Wood WG, Eckert GP, Igbavboa U, Muller WE. Statins and neuroprotection: a prescription to move the field forward. Ann N Y Acad Sci. 2010;1199:69–76. doi: 10.1111/j.1749-6632.2009.05359.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Bu DX, Tarrio M, Grabie N, Zhang Y, Yamazaki H, Stavrakis G, Maganto-Garcia E, Pepper-Cunningham Z, Jarolim P, Aikawa M, Garcia-Cardena G, Lichtman AH. Statin-induced Kruppel-like factor 2 expression in human and mouse T cells reduces inflammatory and pathogenic responses. J Clin Invest. 2010;120:1961–70. doi: 10.1172/JCI41384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kureishi Y, Luo Z, Shiojima I, Bialik A, Fulton D, Lefer DJ, Sessa WC, Walsh K. The HMG-CoA reductase inhibitor simvastatin activates the protein kinase Akt and promotes angiogenesis in normocholesterolemic animals. Nat Med. 2000;6:1004–10. doi: 10.1038/79510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Ponce J, de la Ossa NP, Hurtado O, Millan M, Arenillas JF, Davalos A, Gasull T. Simvastatin reduces the association of NMDA receptors to lipid rafts: a cholesterol-mediated effect in neuroprotection. Stroke. 2008;39:1269–75. doi: 10.1161/STROKEAHA.107.498923. [DOI] [PubMed] [Google Scholar]
- 70.Qi XF, Zheng L, Lee KJ, Kim DH, Kim CS, Cai DQ, Wu Z, Qin JW, Yu YH, Kim SK. HMG-CoA reductase inhibitors induce apoptosis of lymphoma cells by promoting ROS generation and regulating Akt, Erk and p38 signals via suppression of mevalonate pathway. Cell Death Dis. 2013;4:e518. doi: 10.1038/cddis.2013.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Hsieh CH, Rau CS, Hsieh MW, Chen YC, Jeng SF, Lu TH, Chen SS. Simvastatin-induced heme oxygenase-1 increases apoptosis of Neuro 2A cells in response to glucose deprivation. Toxicol Sci. 2008;101:112–21. doi: 10.1093/toxsci/kfm258. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.