

Abstract
Background
In Europe, bank voles (Myodes glareolus) are widely distributed and can transmit Puumala virus (PUUV) to humans, which causes a mild to moderate form of haemorrhagic fever with renal syndrome, called nephropathia epidemica. Uncovering the link between host and virus dynamics can help to prevent human PUUV infections in the future. Bank voles were live trapped three times a year in 2010–2013 in three woodland plots in each of four regions in Germany. Bank vole population density was estimated and blood samples collected to detect PUUV specific antibodies.
Results
We demonstrated that fluctuation of PUUV seroprevalence is dependent not only on multi-annual but also on seasonal dynamics of rodent host abundance. Moreover, PUUV infection might affect host fitness, because seropositive individuals survived better from spring to summer than uninfected bank voles. Individual space use was independent of PUUV infections.
Conclusions
Our study provides robust estimations of relevant patterns and processes of the dynamics of PUUV and its rodent host in Central Europe, which are highly important for the future development of predictive models for human hantavirus infection risk.
Keywords: Myodes glareolus, Population dynamics, Puumala virus seroprevalence, Space use, Survival
Background
Hantaviruses are zoonotic and emerging pathogens for humans with rodent and other small mammal reservoirs [1]. Currently, hantaviruses are known to occur in two rodent families (Cricetidae and Muridae), in two families of insectivores (Soricidae and Talpidae) and in three families of bats (Rhinolophidae, Nycteridae and Vespertilionidae) [2, 3]. All hantaviruses seem to be associated exclusively with one or a few closely related mammal reservoir species and mostly follow the geographical distribution of the reservoir [2, 4, 5].
In Europe, five rodent-borne hantaviruses (Puumala, Tula, Tatenale, Dobrava-Belgrade, and Seoul virus) and four insectivore-borne hantaviruses (Seewis, Asikkala, Boginia and Nova virus) have been identified [6, 7]. According to current knowledge, only some rodent-borne hantaviruses cause significant disease in humans. Infections occur via inhalation of aerosolised virus particles, which are shed through urine, faeces or saliva. Hantavirus disease, namely haemorrhagic fever with renal syndrome (HFRS), occurs in Eurasia and has been known since the 1930s [8]. Puumala virus (PUUV) is the most important hantavirus in Northern, Central and Western Europe [9, 10]. PUUV-caused disease in humans is termed nephropatica epidemica (NE), which is a mild to moderate form of HFRS. The disease is mainly characterized by renal dysfunction or renal failure. Main symptoms are fever, headache, backpain and gastrointestinal symptoms [11]. The much more severe hantavirus cardiopulmonary syndrome is restricted to the Americas [12].
The first detection of PUUV in Germany was in the 1980s during a Belgian military exercise [13]. Since 2001, human hantavirus infections have been notifiable in Germany and a total of 10,403 cases were recorded until December 2016 (Robert Koch Institute: SurvStat@RKI 2.0, https://survstat.rki.de, data status: 31.12.2016). The geographical distribution of human cases in Germany is heterogeneous, with the majority reported from highly endemic regions in southern, western and north-western Germany [5, 14–16]. In outbreak years, the number of human NE cases rises to about 2000 in Germany (SurvStat@RKI 2.0, https://survstat.rki.de, data status: 17.05.2016).
The bank vole (Myodes glareolus) is the main, and in Central Europe exclusive, reservoir of the PUUV. The species is distributed all over Germany and in many other European and Asian countries. In Central Europe it inhabits mainly deciduous broad-leaved forests of oak (Quercus robur) and beech (Fagus sylvatica) [17], but can also occur in hedges, parks, and urban gardens. In temperate forests, food availability for bank voles fluctuates greatly because of seed masting of beech and other tree species, which is triggered by climatic conditions [18–21]. During beech mast years, bank vole populations grow rapidly reaching peak population densities the following year [19, 22–24]. Earlier studies show that in European broad-leaved forests, bank vole populations fluctuate seasonally and multi-annually with population peaks in summer and population outbreaks every 4–7 years [19]. More recent studies demonstrate bank vole outbreaks every 2–3 years [25] due to shortening beech mast intervals [22]. The increase in the number of human PUUV infections is associated with bank vole population peaks [14, 22, 26, 27]. However, the interplay of vole and hantavirus dynamics is much less understood and this may be crucial for hantavirus transmission within host populations ultimately affecting human infection risk. Due to the potentially severe course of the disease and the large number of people affected, understanding the occurrence of human PUUV infections is highly relevant for public health management.
We present the results of a temporally and spatially replicated live trapping study of bank vole populations that link the presence of PUUV specific antibodies in bank voles to their population dynamics, space use and survival. Patterns of bank vole population dynamics are presented for four, and PUUV dynamics for two, geographical regions (PUUV virtually absent from 2 regions) in Germany from 2010 to 2013 covering two vole outbreaks. Results may be relevant for a future development of an early warning system to minimize the risk of hantavirus infections and related adverse effects on public health in Central Europe.
Methods
Rodent trapping
Rodent monitoring was conducted in spring, summer, and autumn of 2010–2013 in four study areas in Germany; West (North Rhine-Westphalia, Billerbeck, 51°59.63′N, 7°18.99′E), South (Baden-Wuerttemberg, Weissach, 48°49.88′N, 8°57.71′E), North (Mecklenburg-Western Pomerania, Jeeser, 54°9.75′N, 13°15.55′E), and East (Thuringia, Gotha, 50°57.38′N, 10°39.13′E). Habitats surveyed were mainly beech forests or mixed deciduous forests.
Ugglan multiple live traps (Grahnab®, Gnosjö, Sweden) were baited with apple, rodent pellets, rolled oats and peanut curls and wood shavings were provided for insulation. On each plot, a trap grid of 49 (7 × 7) traps with 10 m spacing was set. Traps were pre-baited for 3 days and checked twice a day for 2–3 consecutive days, in the early morning and late afternoon. Thus, each trapping session consisted of multiple (3–7) trapping occasions. In total, three replicate woodland plots per study area (region) (0.2–2.2 km apart) and trapping session were sampled. Every captured vole was marked with a passive integrated transponder (PIT) tag (LUX-IDent s.r.o.®, Lanškroun, Czech Republic) for individual identification in the scruff. Species and sex were morphologically determined and animals were weighed to the nearest gram with a 50 g-spring scale (PESOLA AG®, Schindellegi, Switzerland). Blood samples (20–40 µl) were taken from the facial vein or the retro-orbital sinus and stored at −20 °C until analysis for PUUV-specific antibodies. After processing, animals were released at the point of capture.
Density estimation
Bank vole populations were assumed to be closed because immigration, emigration, births and mortality were thought to be minimal during a 3 day trapping period. Closed population densities were estimated using program DENSITY 5.0 (http://www.otago.ac.nz/density/) with the spatial detection model SECR (spatially explicit capture-recapture; [28]) using maximum likelihood (ML). If recapture rates or movement were too limited to calculate density estimates, the minimum number alive multiplied by the trapping area (0.36 ha) was used. Density estimates are stated as individuals per hectare throughout.
Serological PUUV analyses
Blood was analysed in an immunoglobulin G (IgG) enzyme linked immunosorbent assay (ELISA), which uses a yeast-expressed PUUV nucleocapsid protein to test for PUUV antibody presence in the sample. The investigations followed a previously established protocol [29, 30]. Scoring of reactivities (positive, equivocal, negative) followed a previously described decision tree [31]. In further analyses explicit positive or negative results were required, hence equivocal test results were classified as negative. For each state and season mean seroprevalences ± standard deviation were calculated from plots where >4 samples were obtained.
Space use
The minimum distance moved among traps during a trapping session divided by the number of recaptures was used as a measure of space use to investigate potential cause and consequence of PUUV infection in bank voles [32]. Animals with first captures in the traps at the margin of the trapping grid were excluded to avoid edge effects [33] and to increase the probability of including grid residents. Individuals captured only once were excluded from the analysis because no information about movement was available.
Seroconversion and survival
Recaptured bank voles, which were tested for PUUV-specific antibodies in a preceding session, could be identified by their individual marking. This allowed studying seroconversion rates and survival in relation to PUUV infection status. Seroconversion is defined as the occurrence of PUUV-specific antibodies in animals found to be seronegative during previous trapping.
Statistical analyses
Bank vole population density and PUUV seroprevalence
Annual and seasonal fluctuations in population density as well as PUUV seroprevalence were analysed by univariate analyses of variance (ANOVA) with subsequent post hoc tests (Tukey’s HSD). Population density or PUUV seroprevalence were dependent variables and year, season as well as study area fixed factors. Analyses were performed using R software (Version 3.2.5., 2016, R Core Team, Vienna, Austria). Level of significance was α < 5%.
Correlation of PUUV seroprevalence and bank vole population density
The influence of bank vole population density on PUUV-seroprevalence was statistically analysed using a generalized linear mixed model (GLMM) with binomial distribution and a logit link function (level of significance α < 5%) using R software. PUUV seroprevalence as a proportional response variable (2-vector variable) was generated from the number of PUUV seropositive bank voles and the difference between the number of tested bank voles and the number of PUUV seropositive bank voles (=number of PUUV seronegative bank voles). Population density and the density of the previous session (standardised by z-transformation (z = (x − mean)/sd)), both in interaction with season (factorial variable), were included as covariates. Plot nested in study area and year were included as random factors. Further, an observation-level random effect was added to account for overdispersion [34], which was tested a priori using package ‘blmeco’ and function ‘dispersion_glmer’. The number of observations was N = 58 and the best model was chosen according to Akaike information criterion (AIC). We used function ‘r.squaredGLMM’ from the ‘MuMIn’-package to estimate a pseudo-R2 for GLMMs (R2 conditional = variance explained by fixed and random factors) [35, 36].
Space use
Individual differences in space use were investigated under the premise to be either the cause or consequence of PUUV infection. For this the effect of the mean minimum distance moved between recaptures on PUUV infection status (space use = cause) was analysed with a GLMM with binomial distribution and a logit link function (level of significance α < 5%) using R software. PUUV infection status was the dependent variable and ‘mean minimum distance moved’ the fixed factor. Plot nested in study area was included as a random factor. The number of observations was N = 405.
Further, the effect of PUUV infection status on the mean minimum distance moved between recaptures (space use = consequence) was analysed with a GLMM with gamma distribution and a log link function (level of significance α < 5%) using R software. The dependent variable ‘mean minimum distance moved’ was transformed by adding 0.1 m to each value to eliminate zero values. In the initial model, PUUV infection status, sex, weight, interaction of sex as well as weight with PUUV infection status, and season were included as covariates. Plot nested in study area was included as a random factor. The number of observations was N = 405 and the best model was chosen according to AIC.
Seroconversion and survival
Changes in PUUV infection status of recaptures (seroconversion) were analysed by Chi square tests (χ2). Analyses were performed using SPSS (IBM SPSS Statistics for Windows, Version 22.0, 2013, IBM Corp., Armonk, New York). Level of significance was α < 5%.
Survival of bank voles was analysed by a GLMM with binomial distribution and a logit link function (level of significance α < 5%) using R software (if necessary, post hoc tests were performed using ‘multcomp’ (Tukey contrasts) and ‘lsmeans’ (pairwise comparisons) packages). Survival (factorial variable; yes/no) was the dependent variable and PUUV infection status, sex, weight, interaction of sex as well as weight with PUUV infection status, and season were the initial fixed factors. Plot nested in study area was included as a random factor. The number of observations was N = 1263 and the best model was chosen according to AIC.
Results
Population density
In total 3301 bank voles were trapped during 2010–2013 which were roughly equally distributed among study areas. Population density tended to increase from spring to summer or autumn (Fig. 1). There were statistically significant differences in population densities between years (ANOVA: F = 34.54, p < 0.001) and seasons (F = 5.83, p = 0.004) but not between study areas (F = 0.50, p = 0.681). In 2010 and 2012, population densities were significantly higher than in 2011 and 2013 (Tukey’s HSD: 2010/2011 p < 0.001; 2010/2012 p = 0.933; 2010/2013 p < 0.001; 2011/2012 p < 0.001; 2011/2013 p = 0.043; 2012/2013 p < 0.001). Population density in summer was significantly higher compared to spring (p = 0.003) but not to autumn (p = 0.439) or between spring and autumn (p = 0.095). However, maximum densities were observed in summer of 2010 (61–121 ind/ha) and 2012 (72–82 ind/ha), respectively, indicating bank vole population outbreaks. The highest bank vole population density was estimated for summer 2010 in the South with on average 121 ind/ha (Fig. 1). In spring 2011 and 2013 population densities were lowest with 1–19 ind/ha (2011) and 0–5 ind/ha (2013).
Fig. 1.
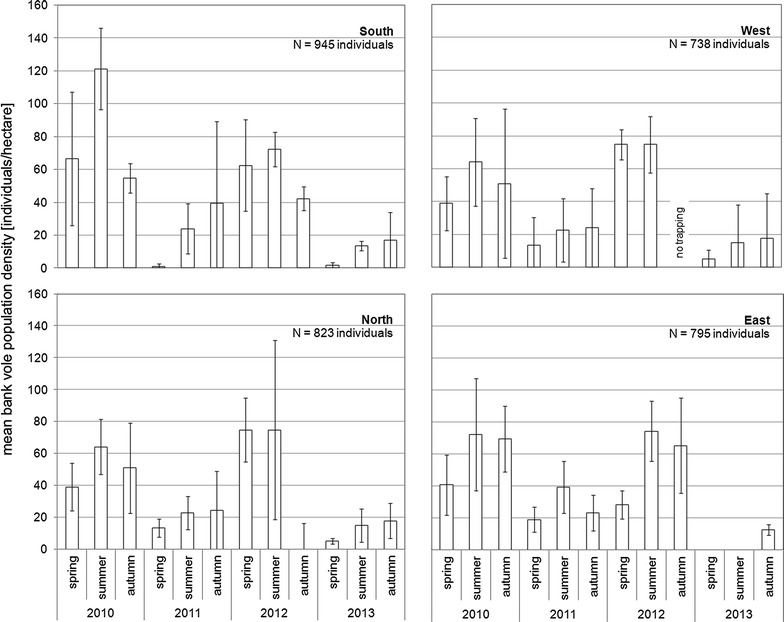
Population dynamics of bank voles in Germany from 2010 to 2013. Estimated mean population densities ± standard deviation as individuals per hectare from three replicate woodland plots per study area (N = total number of trapped bank voles)
PUUV seroprevalence
2800 bank voles were tested for PUUV-reactive antibodies of which 566 (20%) were PUUV seropositive. Almost 99% (561 individuals) of those PUUV-seropositive bank voles were trapped in the South or West. In the North and East only 5 bank voles were tested positive in the PUUV IgG ELISA. Hence, PUUV is most likely not present at the trapping sites in North and East, which is in accordance with a large-scale study on bank voles from eastern Germany [15]. Drewes et al. [15] sampled about 1200 snap-trapped voles from multiple sites in Germany partly overlapping with the sites North and East from this study. All voles were PUUV-negative in IgG ELISA, conventional and real-time RT-PCR. Therefore, the positive test results from the two study areas reported here were most probably due to spillover infections of PUUV-related Tula virus (TULV) or false positive reactions in the ELISA. Because of the general absence of PUUV, North and East were not further included in data analyses regarding PUUV seroprevalence in bank voles.
Serological investigations were performed for 1460 bank voles from West and South. Seroprevalence did not vary between study areas (ANOVA: F = 2.17, p = 0.145) but significantly differed between seasons (F = 4.35, p = 0.017) and years (F = 19.11, p < 0.001) (Fig. 2). Seroprevalences in 2010 and 2012 were significantly higher compared to 2011 and 2013 (Tukey’s HSD: 2010/2011 p < 0.001; 2010/2012 p = 0.998; 2010/2013 p < 0.001; 2011/2012 p < 0.001; 2011/2013 p = 0.367; 2012/2013 p < 0.001). Seroprevalence in spring was significantly higher compared to autumn (p = 0.018) but not to summer (p = 0.097) or between summer and autumn (p = 0.721). However, highest seroprevalence was found in spring 2010 (West: 64 ± 29%, South: 49 ± 32% per plot and trapping session) and 2012 (West: 59 ± 2%, South: 64 ± 40%). In these years, seroprevalence further decreased from spring to summer and from summer to autumn. In 2011 and 2013, the rate of PUUV-seropositive individuals in spring was much lower (<29%), and even zero, compared to the previous year. During those years, PUUV seroprevalence remained at a low or zero level or slightly increased towards autumn.
Fig. 2.
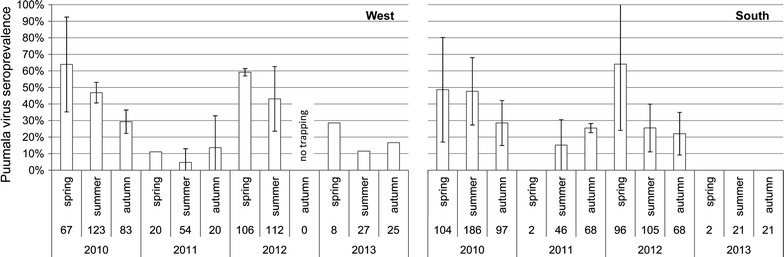
PUUV seroprevalence in bank vole populations in two regions in Germany from 2010 to 2013. Mean seroprevalence ± standard deviation (%) in spring, summer and autumn each year estimated from three replicate woodland plots per study area. Numerical values per season are total numbers of tested individuals of all plots in each study area
Correlation of PUUV seroprevalence and bank vole population density
We performed a GLMM to test for the effect of bank vole population density on the variance in PUUV seroprevalence within populations. The higher the bank vole population density in a trapping session in spring the higher the PUUV seroprevalence (p < 0.001) (Fig. 3) (Table 1). A similar but weaker relation was found in summer (p = 0.027), but not in autumn (p = 0.570) (Table 1) (Fig. 3). There was no effect of the interaction of population density of the previous trapping session and season on PUUV seroprevalence (p > 0.05; removed from model). The random factor plot nested in study area (SA: plot) did not explain the variance in PUUV seroprevalence, but year did (0.31 ± 0.56; variance ± standard deviation). The best model explained almost one-third of the variance in PUUV seroprevalence ( = 0.28).
Fig. 3.
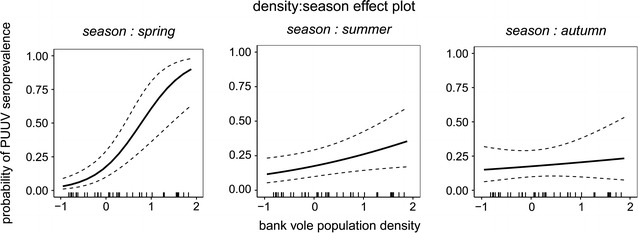
Seasonal effects of bank vole population density (z-transformed) on PUUV seroprevalence in the host population. Black bars on x-axis represent the distribution of population density values per trapping session
Table 1.
Effects of bank vole population density in interaction with season on PUUV seroprevalence in the host population
| Parameter | Estimate | (SE) | z | p |
|---|---|---|---|---|
| Intercept | −1.55 | (0.34) | −4.61 | <0.001 |
| Bank vole population density:season ‘spring’ | 1.99 | (0.44) | 4.56 | <0.001 |
| Bank vole population density:season ‘summer’ | 0.51 | (0.23) | 2.21 | 0.027 |
| Bank vole population density:season ‘autumn’ | 0.19 | (0.34) | 0.57 | 0.570 |
| Random factor | Variance | (SD) | ||
|---|---|---|---|---|
| SA: plot | 0.00 | (0.00) | ||
| Year | 0.31 | (0.56) | ||
Parameter coefficients of generalized linear mixed model (GLMM) with binomial distribution
Number of observations = 58, degrees of freedom = 7
Italic values indicate significance of p value (p < 0.05)
Space use
Analysing the effect of space use as a cause for PUUV infections in bank voles using a GLMM showed no significant effect of the mean minimum distance moved on PUUV infection status (p = 0.205). The random factor plot nested in study area (SA: plot) could explain only a small portion of the variance in the probability of PUUV infection (0.01 ± 0.10; variance ± standard deviation) (Table 2; Fig. 4a).
Table 2.
Model results of GLMMs investigating space use as cause (a) or consequence (b) of PUUV infections in bank voles
| (a) Space use = cause | ||||
|---|---|---|---|---|
| Parameter | Estimate | (SE) | z | p |
| Intercept | −0.38 | (0.17) | −2.20 | 0.028 |
| Mean minimum distance moved | −0.02 | (0.01) | −1.27 | 0.205 |
| Random factor | Variance | (SD) | ||
|---|---|---|---|---|
| SA: plot | 0.01 | (0.10) | ||
| (b) Space use = consequence | ||||
|---|---|---|---|---|
| Parameter | Estimate | (SE) | t | p |
| Intercept | 2.23 | (0.38) | 5.82 | <0.001 |
| PUUV infections status ‘positive’ | −0.21 | (0.13) | −1.55 | 0.120 |
| Sex ‘male’ | −0.90 | (0.59) | −1.52 | 0.129 |
| Random factor | Variance | (SD) | ||
|---|---|---|---|---|
| SA: plot | 0.00 | (0.00) | ||
Parameter coefficients of generalized linear mixed models (GLMM) with binomial distribution (a) and gamma distribution (b)
Number of observations = 405, degrees of freedom = 3 (a) and 4 (b)
Space use = mean minimum distance moved between recaptures
Italic values indicate significance of p value (p < 0.05)
Fig. 4.
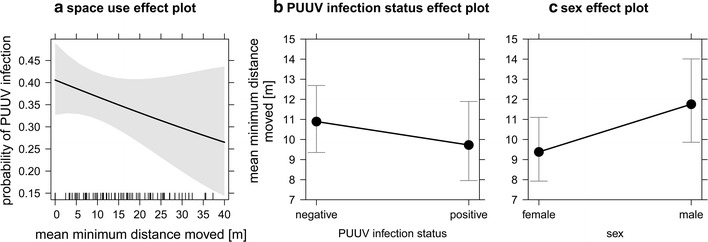
Effect plots of space use as potential cause (a) and consequence (b, c) of PUUV infections (and sex-c) in bank voles. Space use of bank voles = mean minimum distance moved. Black bars on x-axis (a) represent the distribution of individual ‘mean minimum distance moved’ values
We also performed a GLMM to test for the effect of PUUV infection status on the mean minimum distance moved (space use = consequence). First of all, model selection excluded season, weight as well as all interactions from further consideration. No significant effect of PUUV infection status on space use was found (p = 0.378) (Table 2), although the mean minimum distance moved was 11.9% longer for individuals without PUUV-reactive antibodies (Fig. 4b). Males tended to move farther distances than females (p = 0.068) (Fig. 4c). The random factor plot nested in study area (SA: plot) could not explain the variance in the mean minimum distance moved.
Seroconversion and survival
Most individuals trapped more than once were captured in two (122) and some in three (14) or four (2) trapping sessions. We recorded a total of 156 recaptures of individuals in multiple seasons. Usually, recaptures occurred in the following trapping session, but in two cases (one in each study area), voles first captured in summer were not recorded in autumn, but recaptured in the next spring. In both study areas, females were more frequently recaptured than males (West: 33/61 = 54%; South: 56/95 = 59%).
Most recaptures remained seronegative (West 44%; South 46%) followed by bank voles that stayed seropositive (West 34%; South 34%) (Fig. 5). In the West 11 (18%) of recaptures indicated a PUUV seroconversion. In the South 16 voles (17%) seroconverted (Fig. 5). Two voles in the West (1%) and 3 voles in the South (2%) converted from seropositive to seronegative. More females than males remained seronegative (West: 14/27 = 52%; South: 26/44 = 59%) or seropositive (West: 15/21 = 71%; South: 21/32 = 66%). There was no sex difference in seroconversion (p > 0.05).
Fig. 5.
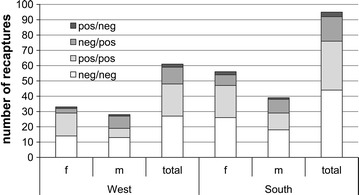
Number of recaptured bank voles. Recaptures per study area (West, South) in total and per sex (f female, m male) subdivided in recaptures that remained seronegative (neg/neg) or seropositive (pos/pos), seroconverted (neg/pos) or seemed to have lost antibodies (pos/neg)
Most recaptures occurred in 2010 and 2012 (78%). Seasonal variation in survival rates (Fig. 6) mirrored fluctuations in bank vole population density in outbreak years with peaks in summer (Fig. 1). A GLMM was performed to investigate the effect of PUUV infection status on survival of bank voles. First of all, the model showed significantly lower survival occurring from autumn to spring (over winter) in comparison to within year survival from spring to summer and summer to autumn (spring/summer z = −1.08, p = 0.520; spring/autumn z = −3.43, p = 0.002; summer/autumn z = −2.86, p = 0.011). This was most probably due to trapping intervals between seasons (twice as long over winter). Hence, separate GLMMs were performed for each season.
Fig. 6.
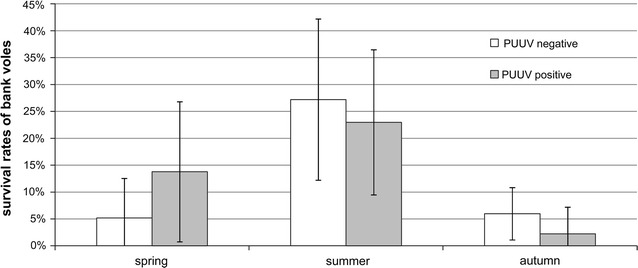
Survival rates of bank voles according to PUUV seroprevalence. Mean values ± standard deviation per season of first capture
Model selection excluded weight from further consideration in each season and all interactions in autumn (Table 3). From spring to summer, survival was significantly higher for PUUV-seropositive voles (p = 0.044) (Table 3; Fig. 7a) but did not vary between sexes. From summer to autumn, survival was significantly lower for seropositive (pos) males (M) in comparison to seropositive or seronegative (neg) females (F) (negF/posF z = −0.09, p = 1.000; negF/negM z = 0.67, p = 0.903; negF/posM z = 2.77, p = 0.027; posF/negM z = 0.63, p = 0.919; posF/posM z = 2.64, p = 0.038; negM/posM z = 2.38, p = 0.076) (Table 3; Fig. 7b). Over winter (autumn to spring), no effect of PUUV infection status or sex on survival was found (Table 3).
Table 3.
Seasonal effects of PUUV infection status, sex and their interaction (not in c) on the survival of bank voles
| (a) Spring | ||||
|---|---|---|---|---|
| Parameter | Estimate | (SE) | z | p |
| Intercept | −2.35 | (0.45) | −5.21 | <0.001 |
| PUUV infections status ‘positive’ | 0.91 | (0.45) | 2.01 | 0.044 |
| Sex ‘male’ | 0.42 | (0.51) | 0.82 | 0.414 |
| PUUV infections status ‘positive’:sex ‘male’ | −1.01 | (0.63) | −1.59 | 0.112 |
| Random factor | Variance | (SD) | ||
|---|---|---|---|---|
| SA: plot | 0.32 | (0.56) | ||
| Number of observations = 367, degrees of freedom = 5 | ||||
| (b) Summer | ||||
|---|---|---|---|---|
| Parameter | Estimate | (SE) | z | p |
| Intercept | −1.82 | (0.30) | −6.04 | <0.001 |
| PUUV infections status ‘positive’ | 0.03 | (0.34) | 0.09 | 0.930 |
| Sex ‘male’ | −0.19 | (0.29) | −0.67 | 0.502 |
| PUUV infections status ‘positive’:sex ‘male’ | −1.35 | (0.65) | −2.07 | 0.038 |
| Random factor | Variance | (SD) | ||
|---|---|---|---|---|
| SA: plot | 0.25 | (0.50) | ||
| Number of observations = 599, degrees of freedom = 5 | ||||
| (c) Autumn | ||||
|---|---|---|---|---|
| Parameter | Estimate | (SE) | z | p |
| Intercept | −2.86 | (0.37) | −7.75 | <0.001 |
| PUUV infections status ‘positive’ | −1.07 | (0.76) | −1.41 | 0.160 |
| Sex ‘male’ | 0.87 | (0.47) | 1.87 | 0.062 |
| Random factor | Variance | (SD) | z | p |
|---|---|---|---|---|
| SA: plot | 0.00 | (0.00) | ||
| Number of observations = 297, degrees of freedom = 4 | ||||
Parameter coefficients of generalized linear mixed models (GLMM) with binomial distribution
Italic values indicate significance of p value (p < 0.05)
Fig. 7.
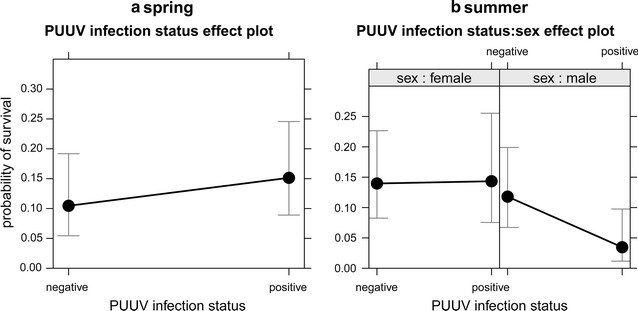
Effect of individual PUUV infection status on survival of bank voles from spring (a) to summer and from summer (b) to autumn. Significant results according to Table 3 are shown
Discussion
The processes involved in hantavirus transmission have been mostly studied in Northern Europe [26, 27, 37, 38]. Knowledge about mechanisms in Central Europe is sparse [17, 21, 39–43]. With our recent study, we covered two spatially replicated bank vole outbreaks (2010 and 2012) and two low phases (2011 and 2013), which is likely to provide robust estimation of relevant patterns and processes.
Population dynamics of bank voles in Central Europe are driven by seed masting of beech trees [18–20, 22, 23]. As beech mast events have recently occurred every 2–3 years [17, 20, 22], bank vole outbreaks have also occured every 2–3 years. This 2–3 year cycle has a major effect on PUUV dynamics within the rodent host population and hence on the number of human PUUV infections.
In our study, PUUV seroprevalence (reliably detected only in two regions) temporally fluctuated depending on host population abundance. Highest seroprevalences were found in 2010 and 2012, when rodent host abundance peaked (Fig. 1, upper graphs) triggered by beech mast events in 2009 and 2011 [22], and coincided with the highest numbers of human PUUV infections ever recorded since the disease became notifiable in 2001 (SurvStat@RKI 2.0, https://survstat.rki.de, data status: 17.05.2016). In bank vole outbreak years, PUUV seroprevalence peaked in spring, when populations consisted of old overwintered animals [37, 44]. These animals most probably die by summer, leading in part to the decrease in PUUV seroprevalence, although population density increases [37, 44]. Further, despite increasing absolute numbers of infected individuals, PUUV seroprevalence decreases (Fig. 2) indicating that population growth rate outperforms transmission rate in peak reproductive phase. Hence, uninfected young of the year lower PUUV seroprevalence during the year [45, 46] (see below). Accordingly, this decrease in PUUV seroprevalence was observed to proceed until autumn. In 2011 and 2013, when bank vole densities collapsed in the West and South, seroprevalence also drastically decreased.
Density-dependence of virus occurrence in the rodent host population was demonstrated for hantavirus [45–47]. However, we found also clear differences in PUUV seroprevalence among seasons. There was a strong direct density-dependence in spring. This might be due to the presence of many overwintered individuals that contracted PUUV in the previous year or during winter and represent the founder population for the upcoming outbreak, while in years with smaller spring populations (not leading to an outbreak), PUUV seroprevalence was much lower. In summer, when uninfected newborns without maternal antibodies (for details see Kallio et al. [48]) enter the population, PUUV seroprevalence is lowered (the juvenile dilution effect—[46]) and consequently density-dependence was decreased, but still significant. In autumn, there was no density effect and density-dependence seems to be diluted over the course of the year likely due to a more complex transmission scenario during the reproduction phase based on reproductive behaviour (e.g. aggression, territoriality; [49]). Our data underlines the dependence of PUUV seroprevalence on seasonal and multi-annual density dynamics of the rodent host in Central-Europe [37, 47].
Considerable temporal variation and the geographic differences in host and virus dynamics were detected indicating that the PUUV-bank vole-human epidemiological system is even more complex than previously assumed. Perhaps most strikingly, PUUV is virtually absent from the German east and north although bank voles are present [15] and fluctuations in abundance are also related to beech masting [22]. Bank voles in the North and East belong to the Eastern and Carpathian evolutionary lineages, whereas bank voles in the South and West belong to the Western evolutionary lineage [15]. The almost complete absence of PUUV-seroreactive bank voles in the current study from sites in North and East is in line with results of a previous study where bank voles from northern and eastern parts of Germany were found to be free of PUUV as determined by IgG-ELISA and RT-PCR analyses [15]. The PUUV absence in the eastern and northern part of Germany is caused by the association of PUUV with the Western evolutionary lineage of the bank vole and its distribution following the postglacial recolonization of Germany [15]. The few PUUV-seroreactive bank voles found in the current study are probably the result of spillover infections, most likely by TULV. TULV is the only hantavirus present in other vole species which is antigenically closely related to PUUV and PUUV- and TULV-specific antibodies cannot be differentiated by ELISA [50, 51]. However, spillover infections seem to occur only rarely (Drewes et al. unpublished data) [2, 30] and hence did not affect the present results.
Male bank voles move farther than female bank voles [39, 52], which could not be confirmed statistically in our study, although males seemed to be more mobile (25.4% farther mean minimum distance moved). More active males are more prone to infection [44, 53], which could lead to the hypothesis that PUUV seroprevalence is positively correlated with space use causing an increased infection probability. Nevertheless, no effect of space use as a cause for PUUV infections in bank voles could be found. A reverse connection of both parameters (reciprocal effect; [32]) might indicate a possible sublethal effect on rodent host fitness (here presented by space use) as a consequence of PUUV infection. However, an effect of PUUV infection status on space use could not be detected, which contrasts with earlier findings that PUUV infection affects host fitness with regards to survival and reproduction [42, 54]. In general, shorter mean minimum distances moved seem to be associated with a higher probability of PUUV infection in bank voles (Fig. 4a, b), but it remains unclear whether PUUV infections affects spatial activity or vice versa. The individual movement data generated by recapture studies are rather crude and more detailed data from telemetry work is needed to further elucidate cause as well as consequences of PUUV infection on rodent host fitness and behaviour.
The proportion of PUUV-seronegative and -seropositive voles or voles with a seroconversion corresponded to earlier findings [39, 55]. Seroconversion in our study (17–18%) was smaller compared to former results (32%; [39]). This is most probably an effect of trapping intervals, which were mostly three months in our study and 6 months in Escutenaire et al. [39], decreasing the probability of seroconversion in our study. ‘Inverted seroconversion’ from positive to negative in three juveniles (first capture ≤15 g) was most likely a consequence of loss of maternal antibodies [56]. Although the presence of PUUV-reactive antibodies is assumed to persist lifelong [57], the loss of antibodies in recaptured animals might indicate a virus clearance [58], as discussed also for another hantavirus [59], or might be caused by an oscillation of the antibody titer below the detection limit of the ELISA used.
Females were more frequently recaptured than males (male:female = 1:1.3). Total capture of females was also higher than males (male:female = 1:1.2) and can be explained by stronger territoriality of females than males [60]. Seroconversion did not differ between sexes. Hence, general assumptions that males have a higher infection risk due to their behavior [44, 47, 53] or that females probably better survive with subject to PUUV infection [54] were not supported. Thus, supposed effects of PUUV on vole fitness [54] require further investigations.
Recaptures between trapping sessions reflecting survival were more frequent in outbreak years when the initial population abundance was higher. Survival also followed seasonal patterns because there was higher survival at the end of the reproductive phase (summer to autumn) for PUUV seronegative and seropositive captures. Over-winter survival was lowest, which is most likely an effect of trapping interval (over winter twice as long as within year) and hence survival according to PUUV infection status was analysed separately for each season. PUUV-seropositive voles survived better from spring to summer, which might indicate a positive effect on host fitness. However, an alternative plausible explanation is the effect of residency. In spring, especially in outbreak years, PUUV seroprevalence is increased reflected by more seropositive than seronegative voles. PUUV-seropositive animals that survived are most probably local residents, which got infected in the previous year and overwintered. Residents are more likely to be recaptured leading to more recapture-based survival than in seronegatives. The latter may include transient or immigrating voles that are recaptured less often than residents. From summer to autumn, PUUV-seropositive males survived less compared to females of either infection status. Again, this might have been an effect of residency. In summer, PUUV seroprevalence is decreased and proportionally more individuals are seronegative most probably due to uninfected young voles. Not only residents but also females (territoriality; [60]) are more likely to be recaptured. Accordingly, males are more likely to disperse, which could have led to less recaptures (apparent reduced survival). Over winter, no effect of PUUV infection on survival was found. Seasonal survival has not been explored so far. Former studies investigating over-winter survival report no effect of PUUV infection on survival [44, 61], but see Kallio et al. [54]. Maternal antibodies did not matter in our study because the number of juvenile recaptured with potential maternal antibodies was negligible (spring to summer N = 1, summer to autumn N = 4). Little is known about sublethal effects of hantavirus infection on behaviour and fitness of rodent hosts that may have consequences for survival and population dynamics. Clarifying such impacts could shed light on our inconsistent findings about the relation of PUUV infection status on survival such as apparent increased survival of PUUV seropositives from spring to summer (maybe induced by an increased PUUV seroprevalence in spring) and apparent decreased survival of seropositive males from summer to autumn. Therefore, further research—preferably in controlled environments—is warranted.
Conclusion
Our study revealed relevant patterns and processes in the dynamics of PUUV and its rodent host in Central Europe. Seasonal and multi-annual fluctuations of PUUV seroprevalence depend on host abundance. This knowledge can facilitate the future development of early warning systems to lower the risk of human hantavirus infections.
Effects of PUUV infection on rodent host space use and survival could not be conclusively clarified. Further research is required to test for such impacts possibly by comparing behaviour and survival between endemic and non-endemic regions.
Authors’ contributions
DR, CI and JJ conceived and designed the study. DR, UMR, CI, SS and JJ collected data, DR, UMR, CI and SS analysed samples and DR and CI analysed the data. DR, CI, RGU, JAE and JJ interpreted results and wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The support of Nastasja G. Spierling in rodent trapping, of Dörte Kaufmann in PUUV ELISA analyses, of Dr. Doreen Gabriel in statistical analyses and of Alice Kenney in proofreading is kindly acknowledged.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics
All procedures involving animals were conducted according to relevant legislation and by permission of the federal authorities (permits for South: Regierungspräsidium Stuttgart 35-9185.82/0261, West: Landesamt für Natur, Umwelt und Verbraucherschutz Nordrhein-Westfalen 8.87-51.05.20.09.210, North: Landesamt für Landwirtschaft, Lebensmittel-sicherheit und Fischerei Mecklenburg-Vorpommern 7221.3-030/09, East: Thüringer Landesamt für Lebensmittelsicherheit und Verbraucherschutz 22-2684-04-15-107/09).
Funding
This study was commissioned and funded by the Federal Environment Agency (UBA) within the Environment Research Plan of the German Federal Ministry for the Environment, Nature Conservation, Building and Nuclear Safety (BMUB) (Grant Number 3709 41 401 and Grant Number 3713 48 401 to JJ). The work was supported by the Federal Ministry of Education and Research (BMBF) through the National Research Platform for Zoonoses (Network ‘Rodent-borne pathogens’; Project Numbers 01KI1018 and 01KI1303, to RGU), by the Robert Koch-Institut with funds of the German Ministry of Public Health (Grant Number 1362/1-924 and 1362/1-980 to RGU), Deutsche Forschungsgemeinschaft (SPP 1596 ‘Ecology and Species Barriers in Emerging Viral Diseases’, UL 405/1-1 to RGU) and the EU grant FP7-261504 EDENext (to RGU) and is catalogued by the EDENext Steering Committee as EDENext472 (http://www.edenext.eu). The contents of this publication are the sole responsibility of the authors and do not necessarily reflect the views of the European Commission.
Footnotes
Daniela Reil and Ulrike M. Rosenfeld contributed equally to this work
Contributor Information
Daniela Reil, Phone: 0049 251 8710642, Email: daniela.reil@julius-kuehn.de.
Ulrike M. Rosenfeld, Email: ulrike.rosenfeld@gmx.de
Christian Imholt, Email: christian.imholt@julius-kuehn.de.
Sabrina Schmidt, Email: sabrina05schmidt@gmail.com.
Rainer G. Ulrich, Email: rainer.ulrich@fli.de
Jana A. Eccard, Email: eccard@uni-potsdam.de
Jens Jacob, Email: jens.jacob@julius-kuehn.de.
References
- 1.Krüger DH, Figueiredo LTM, Song J-W, Klempa B. Hantaviruses—globally emerging pathogens. J Clin Virol. 2015;64:128–136. doi: 10.1016/j.jcv.2014.08.033. [DOI] [PubMed] [Google Scholar]
- 2.Schlegel M, Jacob J, Krüger DH, Rang A, Ulrich RG. Hantavirus emergence in rodents, insectivores and bats: what comes next? In: Johnson N, editor. The role of animals in emerging viral diseases. Boston: Academic Press; 2014. pp. 235–292. [Google Scholar]
- 3.Holmes EC, Zhang YZ. The evolution and emergence of hantaviruses. Curr Opin Virol. 2015;10:27–33. doi: 10.1016/j.coviro.2014.12.007. [DOI] [PubMed] [Google Scholar]
- 4.Plyusnin A, Morzunov SP. Virus evolution and genetic diversity of hantaviruses and their rodent hosts. In: Schmaljohn CS, Nichol ST, editors. Hantaviruses. Berlin: Springer-Verlag; 2001. pp. 47–75. [DOI] [PubMed] [Google Scholar]
- 5.Drewes S, Turni H, Rosenfeld U, Obiegala A, Straková P, Imholt C, Glatthaar E, Dressel K, Pfeffer M, Jacob J et al. Reservoir-driven heterogeneous distribution of recorded human Puumala virus cases in South–West Germany. Zoonoses Public Health. 2016. [DOI] [PubMed]
- 6.Klempa B, Radosa L, Krüger DH. The broad spectrum of hantaviruses and their hosts in Central Europe. Acta Virol. 2013;57(2):130–137. doi: 10.4149/av_2013_02_130. [DOI] [PubMed] [Google Scholar]
- 7.Pounder KC, Begon M, Sironen T, Henttonen H, Watts PC, Voutilainen L, Vapalahti O, Klempa B, Fooks AR, McElhinney LM. Novel hantavirus in field vole, United Kingdom. Emerg Infect Dis. 2013;19(4):673–675. doi: 10.3201/eid1904.121057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lee HW, Lee PW, Johnson KM. Isolation of the ethiologic agent of Korean hemorrhagic fever. J Infect Dis. 1978;137(3):298–308. doi: 10.1093/infdis/137.3.298. [DOI] [PubMed] [Google Scholar]
- 9.Heyman P, Ceianu C, Christova I, Tordo N, Beersma M, Joao Alves M, Lundkvist A, Hukic M, Papa A, Tenorio A, et al. A five-year perspective on the situation of haemorrhagic fever with renal syndrome and status of the hantavirus reservoirs in Europe, 2005–2010. Euro Surveill. 2011;16(36):15–22. doi: 10.2807/ese.16.36.19961-en. [DOI] [PubMed] [Google Scholar]
- 10.Brummer-Korvenkontio M, Henttonen H, Vaheri A. Hemorrhagic fever with renal syndrome in Finland: ecology and virology of nephropathia epidemica. Scand J Infect Dis. 1982;36:88–91. [PubMed] [Google Scholar]
- 11.Settergren B. Clinical aspects of nephropathia epidemica (Puumala virus infection) in Europe: a review. Scand J Infect Dis. 2000;32(2):125–132. doi: 10.1080/003655400750045204. [DOI] [PubMed] [Google Scholar]
- 12.Figueiredo LTM, de Souza WM, Ferres M, Enria DA. Hantaviruses and cardiopulmonary syndrome in South America. Virus Res. 2014;187:43–54. doi: 10.1016/j.virusres.2014.01.015. [DOI] [PubMed] [Google Scholar]
- 13.Clement J, Van der Groen G. Acute hantavirus nephropathy in Belgium: preliminary results of a sero-epidemiological study. Adv Exp Med Biol. 1987;212:251–263. doi: 10.1007/978-1-4684-8240-9_32. [DOI] [PubMed] [Google Scholar]
- 14.Ettinger J, Hofmann J, Enders M, Tewald F, Oehme RM, Rosenfeld UM, Sheikh Ali H, Schlegel M, Essbauer S, Osterberg A, et al. Multiple synchronous outbreaks of Puumala virus, Germany, 2010. Emerg Infect Dis. 2012;18(9):1461–1464. doi: 10.3201/eid1809.111447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Drewes S, Sheikh Ali H, Saxenhofer M, Rosenfeld UM, Binder F, Cuypers F, Schlegel M, Röhrs S, Heckel G, Ulrich RG. Host-associated absence of human Puumala virus infections in northern and eastern Germany. Emerg Infect Dis. 2017;23(1):83–86. doi: 10.3201/eid2301.160224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ulrich RG, Meisel H, Schütt M, Schmidt J, Kunz A, Klempa B, Niedrig M, Pauli G, Krüger DH, Koch J. Verbreitung von Hantavirusinfektionen in Deutschland. Bundesgesundheitsbl-Gesundheitsforsch-Gesundheitsschutz. 2004;47(7):661–670. doi: 10.1007/s00103-004-0858-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Clement J, Maes P, de Strihou CV, van der Groen G, Barrios JM, Verstraeten WW, Van Ranst M. Beechnuts and outbreaks of nephropathia epidemica (NE): of mast, mice and men. Nephrol Dial Transplant. 2010;25(6):1740–1746. doi: 10.1093/ndt/gfq122. [DOI] [PubMed] [Google Scholar]
- 18.Imholt C, Reil D, Eccard JA, Jacob D, Hempelmann N, Jacob J. Quantifying the past and future impact of climate on outbreak patterns of bank voles (Myodes glareolus) Pest Manag Sci. 2015;71:166–172. doi: 10.1002/ps.3838. [DOI] [PubMed] [Google Scholar]
- 19.Pucek Z, Jedrzejewski W, Jedrzejewska B, Pucek M. Rodent population dynamics in a primeval deciduous forest (Bialowieza National Park) in relation to weather, seed crop, and predation. Acta Theriologica. 1993;38(2):199–232. doi: 10.4098/AT.arch.93-18. [DOI] [Google Scholar]
- 20.Tersago K, Verhagen R, Servais A, Heyman P, Ducoffre G, Leirs H. Hantavirus disease (nephropathia epidemica) in Belgium: effects of tree seed production and climate. Epidemiol Infect. 2008;137(2):250–256. doi: 10.1017/S0950268808000940. [DOI] [PubMed] [Google Scholar]
- 21.Clement J, Vercauteren J, Verstraeten WW, Ducoffre G, Barrios JM, Vandamme A-M, Maes P, Van Ranst M. Relating increasing hantavirus incidences to the changing climate: the mast connection. Int J Health Geogr. 2009;8(1):1–11. doi: 10.1186/1476-072X-8-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Reil D, Imholt C, Eccard JA, Jacob J. Beech fructification and bank vole population dynamics—combined analyses of promoters of human Puumala virus infections in Germany. PLoS ONE. 2015;10(7):1–14. doi: 10.1371/journal.pone.0134124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jensen TS. Seed production and outbreaks of non-cyclic rodent populations in deciduous forests. Oecologia. 1982;54:184–192. doi: 10.1007/BF00378391. [DOI] [PubMed] [Google Scholar]
- 24.Hansson L, Jedrzejewska B, Jedrzejewski W. Regional differences in dynamics of bank vole populations in Europe. Pol J Ecol. 2000;48:163–177. [Google Scholar]
- 25.Imholt C, Reil D, Plašil P, Rödiger K, Jacob J. Long-term population patterns of forest rodents and associated damage in German forestry. Pest Manag Sci. 2016. [DOI] [PubMed]
- 26.Kallio ER, Begon M, Henttonen H, Koskela E, Mappes T, Vaheri A, Vapalahti O. Cyclic hantavirus epidemics in humans—predicted by rodent host dynamics. Epidemics. 2009;1(2):101–107. doi: 10.1016/j.epidem.2009.03.002. [DOI] [PubMed] [Google Scholar]
- 27.Olsson GE, Dalerum F, Hörnfeldt B, Elgh F, Palo TR, Juto P, Ahlm C. Human hantavirus infections, Sweden. Emerg Infect Dis. 2003;9(11):1395. doi: 10.3201/eid0911.030275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Borchers DL, Efford M. Spatially explicit maximum likelihood methods for capture–recapture studies. Biometrics. 2008;64(2):377–385. doi: 10.1111/j.1541-0420.2007.00927.x. [DOI] [PubMed] [Google Scholar]
- 29.Mertens M, Kindler E, Emmerich P, Esser J, Wagner-Wiening C, Wölfel R, Petraityte-Burneikiene R, Schmidt-Chanasit J, Zvirbliene A, Groschup MH, et al. Phylogenetic analysis of Puumala virus subtype Bavaria, characterization and diagnostic use of its recombinant nucleocapsid protein. Virus Genes. 2011;43(2):177–191. doi: 10.1007/s11262-011-0620-x. [DOI] [PubMed] [Google Scholar]
- 30.Essbauer S, Schmidt J, Conraths FJ, Friedrich R, Koch J, Hautmann W, Pfeffer M, Wolfel R, Finke J, Dobler G, et al. A new Puumala hantavirus subtype in rodents associated with an outbreak of nephropathia epidemica in south–east Germany in 2004. Epidemiol Infect. 2006;134(6):1333–1344. doi: 10.1017/S0950268806006170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mertens M, Wölfel R, Ullrich K, Yoshimatsu K, Blumhardt J, Römer I, Esser J, Schmidt-Chanasit J, Groschup MH, Dobler G, et al. Seroepidemiological study in a Puumala virus outbreak area in South-East Germany. Med Microbiol Immun. 2009;198(2):83–91. doi: 10.1007/s00430-009-0106-9. [DOI] [PubMed] [Google Scholar]
- 32.Blanchet S, Thomas F, Loot G. Reciprocal effects between host phenotype and pathogens: new insights from an old problem. Trends Parasitol. 2009;25(8):364–369. doi: 10.1016/j.pt.2009.05.005. [DOI] [PubMed] [Google Scholar]
- 33.Hansson L. Home range, population structure and density estimates at removal catches with edge effect. Acta Theriologica. 1969;14:153–160. doi: 10.4098/AT.arch.69-11. [DOI] [Google Scholar]
- 34.Harrison XA. Using observation-level random effects to model overdispersion in count data in ecology and evolution. PeerJ. 2014;2:19. doi: 10.7717/peerj.616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Nakagawa S, Schielzeth H. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol Evol. 2013;4(2):133–142. doi: 10.1111/j.2041-210x.2012.00261.x. [DOI] [Google Scholar]
- 36.Johnson PC. Extension of Nakagawa & Schielzeth’s R2GLMM to random slopes models. Methods Ecol Evol. 2014;5(9):944–946. doi: 10.1111/2041-210X.12225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Voutilainen L, Kallio ER, Niemimaa J, Vapalahti O, Henttonen H. Temporal dynamics of Puumala hantavirus infection in cyclic bank vole populations. Sci Rep-UK. 2016;6. [DOI] [PMC free article] [PubMed]
- 38.Kallio ER, Begon M, Henttonen H, Koskela E, Mappes T, Vaheri A, Vapalahti O. Hantavirus infections in fluctuating host populations: the role of maternal antibodies. Proc Biol Sci. 2010;277(1701):3783–3791. doi: 10.1098/rspb.2010.1022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Escutenaire S, Chalon P, De Jaegere F, Karelle-Bui L, Mees G, Brochier B, Rozenfeld F, Pastoret PP. Behavioral, physiologic, and habitat influences on the dynamics of Puumala virus infection in bank voles (Clethrionomys glareolus) Emerg Infect Dis. 2002;8(9):930–936. doi: 10.3201/eid0809.010537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Escutenaire S, Chalon P, Verhagen R, Heyman P, Thomas I, Karelle-Bui L, Avsic-Zupanc T, Lundkvist A, Plyusnin A, Pastoret PP. Spatial and temporal dynamics of Puumala hantavirus infection in red bank vole (Clethrionomys glareolus) populations in Belgium. Virus Res. 2000;67(1):91–107. doi: 10.1016/S0168-1702(00)00136-2. [DOI] [PubMed] [Google Scholar]
- 41.Piechotowski I, Brockmann SO, Schwarz C, Winter CH, Ranft U, Pfaff G. Emergence of hantavirus in South Germany: rodents, climate and human infections. Parasitol Res. 2008;103:131–137. doi: 10.1007/s00436-008-1055-8. [DOI] [PubMed] [Google Scholar]
- 42.Tersago K, Crespin L, Verhagen R, Leirs H. Impact of Puumala virus infection on maturation and survival in bank voles: a capture-mark-recapture analysis. J Wildl Dis. 2012;48(1):148–156. doi: 10.7589/0090-3558-48.1.148. [DOI] [PubMed] [Google Scholar]
- 43.Schwarz AC, Ranft U, Piechotowski I, Childs JE, Brockmann SO. Risk factors for human infection with Puumala virus, southwestern Germany. Emerg Infect Dis. 2009;15(7):1032–1039. doi: 10.3201/eid1507.081413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bernshtein A, Apekina N, Mikhailova T, Myasnikov YA, Khlyap L, Korotkov YS, Gavrilovskaya I. Dynamics of Puumala hantavirus infection in naturally infected bank voles (Clethrinomys glareolus) Arch Virol. 1999;144(12):2415–2428. doi: 10.1007/s007050050654. [DOI] [PubMed] [Google Scholar]
- 45.Olsson GE, Ahlm C, Elgh F, Verlemyr A-C, White N, Juto P, Palo RT. Hantavirus antibody occurrence in bank voles (Clethrionomys glareolus) during a vole population cycle. J Wildl Dis. 2003;39(2):299–305. doi: 10.7589/0090-3558-39.2.299. [DOI] [PubMed] [Google Scholar]
- 46.Mills JN, Ksiazek TG, Peters CJ, Childs JE. Long-term studies of hantavirus reservoir populations in the southwestern United States: a synthesis. Emerg Infect Dis. 1999;5(1):135–142. doi: 10.3201/eid0501.990116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Olsson GE, White N, Ahlm C, Elgh F, Verlemyr AC, Juto P, Palo RT. Demographic factors associated with hantavirus infection in bank voles (Clethrionomys glareolus) Emerg Infect Dis. 2002;8(9):924. doi: 10.3201/eid0809.020037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kallio ER, Poikonen A, Vaheri A, Vapalahti O, Henttonen H, Koskela E, Mappes T. Maternal antibodies postpone hantavirus infection and enhance individual breeding success. Proc Biol Sci. 2006;273(1602):2771–2776. doi: 10.1098/rspb.2006.3645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Tersago K, Verhagen R, Leirs H. Temporal variation in individual factors associated with hantavirus infection in bank voles during an epizootic: implications for Puumala virus transmission dynamics. Vector Born Zoonotic Dis. 2011;11(6):715–721. doi: 10.1089/vbz.2010.0007. [DOI] [PubMed] [Google Scholar]
- 50.Heider H, Ziaja B, Priemer C, Lundkvist Å, Neyst J, Krüger DH, Ulrich R. A chemiluminescence detection method of hantaviral antigens in neutralisation assays and inhibitor studies. J Virol Methods. 2001;96(1):17–23. doi: 10.1016/S0166-0934(01)00314-7. [DOI] [PubMed] [Google Scholar]
- 51.Schmidt S, Saxenhofer M, Drewes S, Schlegel M, Wanka KM, Frank R, Klimpel S, Blanckenhagen F, Maaz D, Herden C, et al. High genetic structuring of Tula hantavirus. Arch Virol. 2016;161(5):1135–1149. doi: 10.1007/s00705-016-2762-6. [DOI] [PubMed] [Google Scholar]
- 52.Kozakiewicz M, Chołuj A, Kozakiewicz A. Long-distance movements of individuals in a free-living bank vole population: an important element of male breeding strategy. Acta Theriologica. 2007;52(4):339–348. doi: 10.1007/BF03194231. [DOI] [Google Scholar]
- 53.Deter J, Chaval Y, Galan M, Gauffre B, Morand S, Henttonen H, Laakkonen L, Voutilainen N, Charbonnel N, Cosson JF. Kinship, dispersal and hantavirus transmission in bank and common voles. Arch Virol. 2008;153(3):435–444. doi: 10.1007/s00705-007-0005-6. [DOI] [PubMed] [Google Scholar]
- 54.Kallio ER, Voutilainen L, Vapalahti O, Vaheri A, Henttonen H, Koskela E, Mappes T. Endemic hantavirus infection impairs the winter survival of its rodent host. Ecology. 2007;88(8):1911–1916. doi: 10.1890/06-1620.1. [DOI] [PubMed] [Google Scholar]
- 55.WeberdeMelo V, Sheikh Ali H, Freise J, Kühnert D, Essbauer S, Mertens M, Wanka KM, Drewes S, Ulrich RG, Heckel G. Spatiotemporal dynamics of Puumala hantavirus associated with its rodent host, Myodes glareolus. Evol Appl. 2015;8(6):545–559. doi: 10.1111/eva.12263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kallio ER, Henttonen H, Koskela E, Lundkvist A, Mappes T, Vapalahti O. Maternal antibodies contribute to sex-based difference in hantavirus transmission dynamics. Biol Lett. 2013;9(6):20130887. doi: 10.1098/rsbl.2013.0887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Voutilainen L, Sironen T, Tonteri E, Tuiskunen Back A, Razzauti M, Karlsson M, Wahlstrom M, Niemimaa J, Henttonen H, Lundkvist A. Life-long shedding of Puumala hantavirus in wild bank voles (Myodes glareolus) J Gen Virol. 2015;96:1238–1247. doi: 10.1099/vir.0.000076. [DOI] [PubMed] [Google Scholar]
- 58.Alexeyev OA, Ahlm C, Elgh F, Aava B, Palo T, Settergren B, Tärnvik A, Wadell G, Juto P. A minority of seropositive wild bank voles (Clethrionomys glareolus) show evidence of current Puumala virus infection. Epidemiol Infect. 1998;121(02):419–425. doi: 10.1017/S0950268898001307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kuenzi AJ, Douglass RJ, Bond CW, Calisher CH, Mills JN. Long-term dynamics of Sin Nombre viral RNA and antibody in deer mice in Montana. J Wildl Dis. 2005;41(3):473–481. doi: 10.7589/0090-3558-41.3.473. [DOI] [PubMed] [Google Scholar]
- 60.Koskela E, Mappes T, Ylönen H. Territorial behaviour and reproductive success of bank vole Clethrionomys glareolus females. J Anim Ecol. 1997;66:341–349. doi: 10.2307/5980. [DOI] [Google Scholar]
- 61.Verhagen R, Leirs H, Tkachenko E, Vandergroen G. Ecological and epidemiologic data on Hantavirus in bank vole populations in Belgium. Arch Virol. 1986;91(3–4):193–205. doi: 10.1007/BF01314280. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.