

Abstract
Phylogenetic analyses were performed using multiple nuclear (ITS and ETS) and chloroplast regions (ndhF-trnL, psbA-trnH, psbD-trnT, and trnT-trnL) to test the monophyly of Aconitum subgen. Lycoctonum (Ranunculaceae) and reconstruct the phylogenetic relationships within the subgenus. The subgenus as currently circumscribed is revealed to be polyphyletic. To achieve its monophyly, sect. Galeata and sect. Fletcherum, both being unispecific and each having a unique array of characters (the latter even having the aberrant base chromosome number of x = 6), must be removed from the subgenus. The subgenus Lycoctonum should thus be redefined to include only two sections, the unispecific sect. Alatospermum and the relatively species-rich sect. Lycoctonum. The section Alatospermum, which is both morphologically and karyologically in the primitive condition, is resolved as the first diverging lineage of the subgenus Lycoctonum clade. The monophyly of sect. Lycoctonum is strongly supported, but all the ten series currently recognized within the section are revealed to be para- or poly-phyletic. Five major clades are recovered within the section. We propose to treat them as five series: ser. Crassiflora, ser. Scaposa, ser. Volubilia, ser. Longicassidata, and ser. Lycoctonia. Thus, a formal reclassification of subgen. Lycoctonum is presented, which involves segregating both sect. Galeata and sect. Fletcherum from the subgenus as two independent subgenera within the genus Aconitum, reinstating one series (ser. Crassiflora) and abolishing six series (ser. Laevia, ser. Longibracteolata, ser. Micrantha, ser. Ranunculoidea, ser. Reclinata, and ser. Umbrosa) within sect. Lycoctonum. The series affiliation of some species within the section is adjusted accordingly.
Introduction
Aconitum L. subgen. Lycoctonum (DC.) Peterm. (Ranunculaceae) consists of ca. 40 species distributed in Eurasia, northern Africa, and eastern North America [1]. It is different from its closest ally, subgen. Aconitum, by having perennial rhizomes (vs. tuberous roots) [1–3]. Another subgenus long recognized, subgen. Gymnaconitum (Stapf) Rapaics, which has annual taproots, was recently segregated as an independent genus of its own, i.e. Gymnaconitum (Stapf) Wei Wang & Z. D. Chen [4]. Many of the species in subgen. Lycoctonum are of potential medicinal value [5].
The first comprehensive classification of Aconitum subgen. Lycoctonum was proposed by Lauener and Tamura [2] and Tamura and Lauener [6] based on morphology, in which four sections and 11 series were recognized (Table 1). Later, Tamura [1] basically adopted this classification with only some minor changes in the arrangement order of the sections and series, and in the number of series as well. As shown in Table 1, he divided the subgenus into four sections: sect. Alatospermum Tamura, sect. Galeata Rapaics, sect. Fletcherum Tamura, and sect. Lycoctonum DC. The former three are all unispecific, including each the eastern Himalayan A. novoluridum Munz, the Kashmir Himalayan A. moschatum (Brühl ex Duthie) Stapf, and the eastern Himalayan A. fletcheranum G. Taylor, respectively. The section Lycoctonum, occurring nearly in the same area as subgen. Lycoctonum, comprises the remaining species which were subdivided into nine series: ser. Micrantha Steinb. ex Tamura & Lauener, ser. Scaposa W. T. Wang, ser. Laevia Tamura & Lauener, ser. Reclinata Tamura & Lauener, ser. Volubilia (Nakai) Tamura & Lauener, ser. Longibracteolata Steinb. ex Tamura & Lauener, ser. Ranunculoidea Steinb. ex Tamura & Lauener, ser. Lycoctonia Tamura & Lauener, and ser. Longicassidata (Steinb. ex Nakai) Tamura & Lauener. It is to be noted that Kadota [7] proposed ser. Umbrosa (Steinb. ex Nakai) Kadota to accommodate A. mashikense Kadota & Umezawa, A. gigas H. Lév. & Vaniot and A. umbrosum (Korsh.) Kom, the latter two of which had been previously placed in ser. Lycoctonia by Tamura and Lauener [6] and Tamura [1]. Later, Kadota [8] ascribed another six species from northern Japan to ser. Umbrosa, five of which were described as new by him.
Table 1. Historical classifications of Aconitum subgen. Lycoctonum.
| Tamura and Lauener (1979) | Tamura (1995) |
|---|---|
| subgen. Lycoctonum | subgen. Lycoctonum |
| sect. Galeata (A. moschatum) | sect. Alatospermum (A. novoluridum) |
| sect. Fletcherum (A. fletcheranum) | sect. Galeata (A. moschatum) |
| sect. Alatospermum (A. novoluridum) | sect. Fletcherum (A. fletcheranum) |
| sect. Lycoctonum | sect. Lycoctonum |
| ser. Scaposa (A. scaposum, var. chloranthum, var. patentipilum, A. cavaleriei, A. aggregatifolium) | ser. Micrantha (A. apetalum, A. sajanense, A. brevicalcaratum, var. lauenerianum, A. chrysotrichum) |
| ser. Crassiflora (A. crassiflorum) | ser. Scaposa (A. scaposum, var. chloranthum, var. patentipilum, A. cavaleriei, A. aggregatifolium, A. crassiflorum, A. rilongense*) |
| ser. Laevia (A. laeve, var. curvipilosum) | ser. Laevia (A. laeve, var. curvipilosum) |
| ser. Reclinata (A. reclinatum) | ser. Reclinata (A. reclinatum) |
| ser. Volubilia (A. alboviolaceum, A. loczyanum, A. pseudolaeve, A. quelpaertense, A. chrysopilum, A. pterocaule, var. albidum, var. glabrescens, A. pteropus, A. desoulavyi) | ser. Volubilia (A. alboviolaceum, A. loczyanum, A. pseudolaeve, A. quelpaertense, A. chrysopilum, A. pterocaule, var. albidum, var. glabrescens, A. pteropus, A. desoulavyi, A. finetianum#, A. longecassidatum#) |
| ser. Longibracteolata (A. sukaczevii) | ser. Longibracteolata (A. sukaczevii) |
| ser. Micrantha (A. apetalum, A. sajanense) | ser. Ranunculoidea (A. ranunculoides, A. ajanense, A. crassifolium) |
| ser. Brevicalcarata (A. brevicalcaratum, var. lauenerianum) | ser. Lycoctonia (A. septentrionale, A. moldavicum, A. wardii, var. hopeiense. A. angustius, A. orientale, A. iranshahrii, A. ranunculifolium, A. monticola, A. krylovii, A. puchonroenicum, A. umbrosum, A. gigas, A. vulparia, A. pauciflorum, A. leucostomum#, A. wangyedianense*, A. sinomontanum#, A. shennongjiaense*) |
| ser. Longicassidata (A. barbatum, var. puberulum, A. kirinense, var. australe, A. lasiostomum) | ser. Longicassidata (A. barbatum, var. puberulum, A. kirinense, var. australe, A. lasiostomum) |
| ser. Ranunculoidea (A. ranunculoides, A. ajanense, A. crassifolium) | |
| ser. Lycoctonia (A. septentrionale, A. moldavicum, var. sinomontanum, A. wardii, var. hopeiense, A. angustius, A. orientale, A. iranshahrii, A. ranunculifolium, A. monticola, A. krylovii, A. puchonroenicum, A. umbrosum, A. gigas, var. hondoense, A. vulparia, A. pauciflorum) |
* Species referred to the series by their author(s), not by Tamura and Lauener (1979) and Tamura (1995).
# Species reduced to synonyms by Tamura and Lauener (1979) but recognized as independent species by other authors: both Aconitum finetianum and A. longecassidatum as synonyms of A. pterocaule, A. leucostomum as a synonym of A. wardii, and A. sinomontanum as a synonym of A. moldavicum.
The circumscription of Aconitum subgen. Lycoctonum by Tamura [1] has been questioned by some authors. Kadota [9] transferred A. fletcheranum from this subgenus to his new subgenus, subgen. Tangutica (W. T. Wang) Kadota, and still maintained its independent sectional status, stating that the nectary blade of the petal in this species was not provided with a tubular portion. For the same reason he also considered that the inclusion of A. moschatum in subgen. Lycoctonum was highly doubtful, although he did not pinpoint further its systematic position [9]. Indeed, except for A. fletcheranum and A. moschatum, all the other species within subgen. Lycoctonum have a nectary blade provided with a tubular portion [1, 2, 6, 9]. The inclusion of A. fletcheranum within subgen. Lycoctonum seems also quite abnormal in terms of karyological characters. Most recently Hong et al. [10] reported the chromosome number of this species as 2n = 12. This count represents a new base chromosome number of x = 6 for the genus Aconitum which otherwise has a uniform base chromosome number of x = 8. The karyotype of A. fletcheranum is unique in Aconitum. There are two largest metacentric chromosome pairs in this species, whereas in all the other diploid taxa of the genus with available chromosomal data there is only one such chromosome pair, with the second largest pair being submetacentric [10].
Tamura’s [1] classification of the largest section (including ca. 40 species) within Aconitum subgen. Lycoctonum, i.e., sect. Lycoctonum, may also be problematic. For example, he placed A. brevicalcaratum (Finet & Gagnep.) Diels and A. chrysotrichum W. T. Wang in ser. Micrantha while A. crassiflorum Hand.-Mazz. in ser. Scaposa, but these three species, all occurring in the southern part (western Sichuan, northwestern Yunnan) of the Hengduan Mountains region in southwestern China, are morphologically most closely similar to each other [11, 12]. More importantly, they are all tetraploid (2n = 32) with almost identical karyotypes [10, 13]. Yuan and Yang [13] proposed that the three species, together with A. rilongense Kadota, also a tetraploid species from western Sichuan, should be placed in one and the same series.
Utelli et al. [14] used the chloroplast intergenic spacer psbA-trnH and nuclear ITS region to study the relationships of the Aconitum lycoctonum L. species complex from Europe and the Caucasus Mountains. No other molecular phylogenetic study focused on subgen. Lycoctonum has been made, although more or less species of the subgenus (all belonging to sect. Lycoctonum) were chosen as outgroups or placeholders in phylogenetic analyses of subgen. Aconitum [15, 16] or of the tribe Delphinieae [4, 17]. Significantly, the nine species sampled by Jabbour and Renner [17] and the 18 species sampled by Wang et al. [4] form a well-supported monophyletic clade in the phylograms obtained from combined trnL-F and ITS dataset by using maximum likelihood method. It is also noteworthy that the genus Aconitum, after the segregation of subgen. Gymnaconitum from it, is a monophyletic group, with the genera Delphinium L. and Gymnaconitum as its sister groups [4, 17]. This provides us a framework to probe further into the phylogeny of subgen. Lycoctonum.
In the present study, we sampled the majority of the known species within Aconitum subgen. Lycoctonum and used multiple nuclear (ITS and ETS) and chloroplast regions (ndhF-trnL, psbA-trnH, psbD-trnT, and trnT-trnL) to perform phylogenetic analyses on this subgenus. Our aims were to (1) test the monophyly of subgen. Lycoctonum, (2) reconstruct the phylogenetic relationships within the subgenus, and (3) provide a reclassification of the subgenus that is phylogeny driven.
Materials and methods
Taxon sampling
We sampled 61 accessions representing 41 species (ca. 87% of the recognized taxa, Table 1) which covered all the sections and series in Aconitum subgen. Lycoctonum (S1 Table). In order to settle the dispute over the phylogenetic relationships of A. fletcheranum and A. moschatum, we further sampled 34 species from subgen. Aconitum, which represent four of its five sections according to Tamura [1] (sect. Austrokoreensia Nakai, not included herein, is unispecific, including only A. austrokoreense Koidz. from southern Korea). Based on the results of Jabbour and Renner [17] and Wang et al. [4], Gymnaconitum gymnandrum (Maxim.) Wei Wang & Z.D. Chen, the single species in Gymnaconitum, and three species of Delphinium were selected as outgroups.
DNA extraction, PCR-amplification, and sequencing
Total DNA was extracted from silica gel-dried leaf tissue or herbarium specimens, using the modified CTAB method [18] or using a DNeasy Plant Mini Kit (Qiagen). Six markers were employed in this study, including ITS, ETS, psbA-trnH, psbD-trnT, trnT-trnL and ndhF-trnL. All primers used for amplification and sequencing are given in Table 2. The whole internal transcribed spacer (ITS, including ITS1, 5.8S rDNA, ITS2) was amplified with universal primers ITS5 and ITS4 [19]. For some degraded DNA from poor quality herbarium tissue, ITS1 and ITS2 regions were amplified separately with additional primers (P2 and P4) as internal primers (Table 2). For chloroplast DNA regions, primers were designed using Primer Premier 5.0 based on the complete chloroplast genome of Megaleranthis saniculifolia Ohwi [20] (Table 2). Polymerase chain reaction (PCR) was performed in a total reaction (25 μL) containing 14.75 μL ddH2O, 5 μL 5 × PrimeSTAR® Buffer (Mg2+ plus), 2 μL dNTP Mixture (2.5 mM), 0.5 μL of each primer (10 μM), 0.25 μL of PrimerSTAR® HS DNA Polymerase (2.5 U/μL), and 2 μL of template DNA (10–40 ng). All reactions were carried out as follows: predenaturation at 95°C for 3 min followed by 35 cycles of denaturation at 94°C for 30 s, annealing at 53°C for 40 s, extension at 72°C for 50 s, and a final extension at 72°C for 8 min. PCR products were checked for length and concentration on 1.5% agarose gels and sent to Shanghai Invitrogen Biotechnology for commercial sequencing. Sequences were aligned using CLUSTALX v.2.1 [21, 22] with default settings and adjusted manually using Bioedit v.7.0.5 [23]. The complete concatenated alignment of these six regions has been deposited in TreeBASE (http://www.treebase.org; accession number: S20400), but regions of poly-A/T (≥10 replicates) and ambiguous alignment (nucleotide positions 87–104, 339–370, 428–443, 659–680, 685–694, 1331–1352, 1391–1400, 1404–1414, 1891–1902, 1956–1968) were excluded from the following analyses.
Table 2. A list of the primers used in this study.
| Primer | Fragment | Sequence (5’-3’) | Reference |
|---|---|---|---|
| ITS4 | ITS2 | TCCTCCGCTTATTGATATGC | White et al. (1990) |
| P4 | ITS2 | ATTGCAGAATCCCGTGAACC | This study |
| ITS5 | ITS1 | GGAAGTAAAAGTCGTAACAAGG | White et al. (1990) |
| P2 | ITS1 | GCTACGTTCTTCATCGATGC | This study |
| 18S-ETS | ETS | ACTTACACATGCATGGCTTAATCT | Baldwin et al. (1998) |
| ETS-R | ETS | TGATTTTGGGTTTTCGATCCACTAC | This study |
| Ndh1F | Partial ndhF-trnL | ATTGTTTCCGATTCACCAGCTCTTA | This study |
| Ndh1R | Partial ndhF-trnL | GCAACTCACTCAGTTTCACAACGAA | This study |
| Ndh2F | Partial ndhF-trnL | TCGTTGTGAAACTGAGTGAGTTGCT | This study |
| RTLjR | Partial ndhF-trnL | GAGCAGCGTGTCTACCGATT | This study |
| psbA | psbA-trnH | GTTATGCATGAACGTAATGCTC | Sang et al. (1997) |
| trnH2 | psbA-trnH | CGCGCATGGTGGATTCACAATCC | Tate (2002) |
| DT-f1 | Partial psbD-trnT | ACCTCATAGCATTTTCGGGAC | This study |
| DT-r2 | Partial psbD-trnT | GATTTATCTGAAGGAAAAGGGGGAA | This study |
| TTL1F | Partial trnT-trnL | CTCTGAGCTAAGCGGGCTCGCATAA | This study |
| TTL1R | Partial trnT-trnL | CCCCCACCCTTTTGAATGAACACAG | This study |
Phylogenetic analyses
Phylogenetic analyses of the nrDNA datasets, the chloroplast datasets, and the combined datasets, were conducted using PAUP v.4.0b10 [24], GARLI (genetic algorithm for rapid likelihood inference) v.2.0 [25] and MrBayes v.3.2.1 [26]. Maximum parsimony (MP) searches were performed using heuristic search methods with tree bisection reconnection (TBR) branch swapping, and equal weighting of all characters. The analyses were repeated 1,000 times with a random order of sequence addition in an attempt to sample multiple islands of most parsimonious trees. Bootstrap tests were carried out to evaluate node support using 1,000 replicates with heuristic search settings identical to those for the original search. We determined the best-fit model of sequence evolution using the program Modeltest v.3.7 [27]. Maximum likelihood (ML) searches were carried out in GARLI v.2.0 using models selected by the Akaike information criterion (AIC) for each dataset. GARLI was run with eight replicates, using the default settings. The topology with the highest likelihood score was chosen as the best tree. For statistical support of branches, non-parametric bootstrap values were computed with 100 replicates, and support values were calculated using PAUP v.4.0b10. Bayesian inferences (BI) were conducted using the different models selected from Modeltest for each partition. Ten million generations were run to estimate parameters relating to sequence evolution and likelihood probabilities using a Markov chain Monte Carlo (MCMC) method. Trees were collected every 1000th generation. Convergence of runs was tested by inspecting whether the standard deviation of split frequencies of the runs was < 0.01 and by using the effective sample sizes (ESS) as calculated with Tracer v.1.4 [28], considering ESS values > 200 as good evidence. After removing 25% of the generations as burn in, a 50% majority rule consensus tree was calculated to generate a posterior probability for each node.
Incongruence tests
To evaluate the congruence of datasets from different gene markers we employed the incongruence length difference (ILD) test [29] implemented in PAUP v.4.0b10 [24]. We used simple taxon addition, TBR branch swapping, and heuristic searches with 999 repartitions of the data. The ILD test was carried out with pairwise partition for each gene dataset as well as with the combined dataset. P-values below 0.05 were considered as evidence of significant incongruence [29].
As the ILD test suggested a significant difference between the cpDNA and nrDNA data, we visually compared the cpDNA and nrDNA trees and located five samples that were incongruently placed with strong support. These include Aconitum apetalum (Huth) B. Fedtsch. ex Steinb., two accessions of A. barbatum Pers. var. barbatum (ZY69 and GQ150), A. fletcheranum, and A. gigas var. hondoense Nakai ex Tamura & Lauener. Both Wilcoxon signed-ranks (WSR) test [30, 31] and approximately unbiased (AU) test [32] were further employed to assess the level of contribution of these samples to the conflict between the cpDNA and nrDNA data. Aconitum moschatum was of special interest in the phylogenetic position and thus also subjected to the WSR and AU tests, although its sister relationship to subgen. Aconitum in the nrDNA tree did not receive strong support. These six samples were first pruned from the original nrDNA and cpDNA datasets and then re-added individually. For each sample, the relationship inferred from one dataset was used as a constraint topology to test against the alternative one inferred from the other data-set. For WSR tests, PAUP v.4.0b10 [24] was employed to optimize the constraint topologies using MP approach. Before AU tests, GARLI v.2.0 [25] was used to optimize the constraint topologies, and then to calculate the site-log-likelihood values for both the best and the optimized constraint trees. The AU tests were implemented in CONSEL v.0.2 [33] using the default settings. P-values below 0.05 were considered to indicate significant differences.
Results
Phylogenetic analyses of chloroplast sequence data
There was no significant incongruence among the four chloroplast regions and the p-values resulting from the ILD test of pairwise sequences are shown in Table 3. Therefore, we combined all markers into a single dataset. For the combined cpDNA dataset, information of each aligned DNA data, tree statistics for the MP analysis and the best-fit model of each region are given in Table 4.
Table 3. P-values of the partition-homogeneity tests.
The pruned datasets are reconstructed from the original datasets by excluding Acontium apetalum, two accessions of A. barbatum var. barbatum (ZY69 and GQ150), A. fletcheranum, A. gigas var. hondoense, and A. moschatum. Bold-faced values indicate rejection of the null hypothesis with 95% confidence.
| Dataset of | P-value |
|---|---|
| ETS vs. ITS | 0.088 |
| ndhF-trnL vs. psbA-trnH | 0.050 |
| ndhF-trnL vs. psbD-trnT | 0.174 |
| ndhF-trnL vs. trnT-trnL | 0.097 |
| psbA-trnH vs. psbD-trnT | 0.309 |
| psbA-trnH vs. trnT-trnL | 1.000 |
| psbD-trnT vs. trnT-trnL | 0.576 |
| cpDNA vs. nrDNA (original datasets) | 0.001 |
| cpDNA vs. nrDNA (pruned datasets) | 0.001 |
Table 4. Statistics of the nuclear and chloroplast sequence datasets.
| ITS | ETS | ndhF-trnL | psbA-trnH | psbD-trnT | trnT-trnL | nrDNA | cpDNA | Combined cpDNA and nrDNA | |
|---|---|---|---|---|---|---|---|---|---|
| Number of taxa | 97 | 83 | 99 | 98 | 81 | 68 | 98 | 99 | 99 |
| Aligned length | 670 | 318 | 944 | 301 | 638 | 417 | 988 | 2300 | 3288 |
| No. variable characters | 267 | 185 | 180 | 65 | 92 | 81 | 452 | 402 | 854 |
| No. parsimony-informative characters | 173 | 133 | 94 | 41 | 41 | 37 | 306 | 211 | 517 |
| Tree length (steps) | 523 | 394 | 265 | 93 | 110 | 91 | 929 | 555 | 1532 |
| Consistency index (CI) | 0.671 | 0.624 | 0.815 | 0.774 | 0.864 | 0.923 | 0.643 | 0.809 | 0.683 |
| Retention index (RI) | 0.921 | 0.871 | 0.922 | 0.909 | 0.943 | 0.965 | 0.900 | 0.919 | 0.895 |
| Rescaled consistency index (RC) | 0.618 | 0.544 | 0.751 | 0.704 | 0.814 | 0.891 | 0.578 | 0.744 | 0.611 |
| Model | GTR+I+G | GTR+G | TIM+I+G | HKY+G | K81uf+G | K81uf |
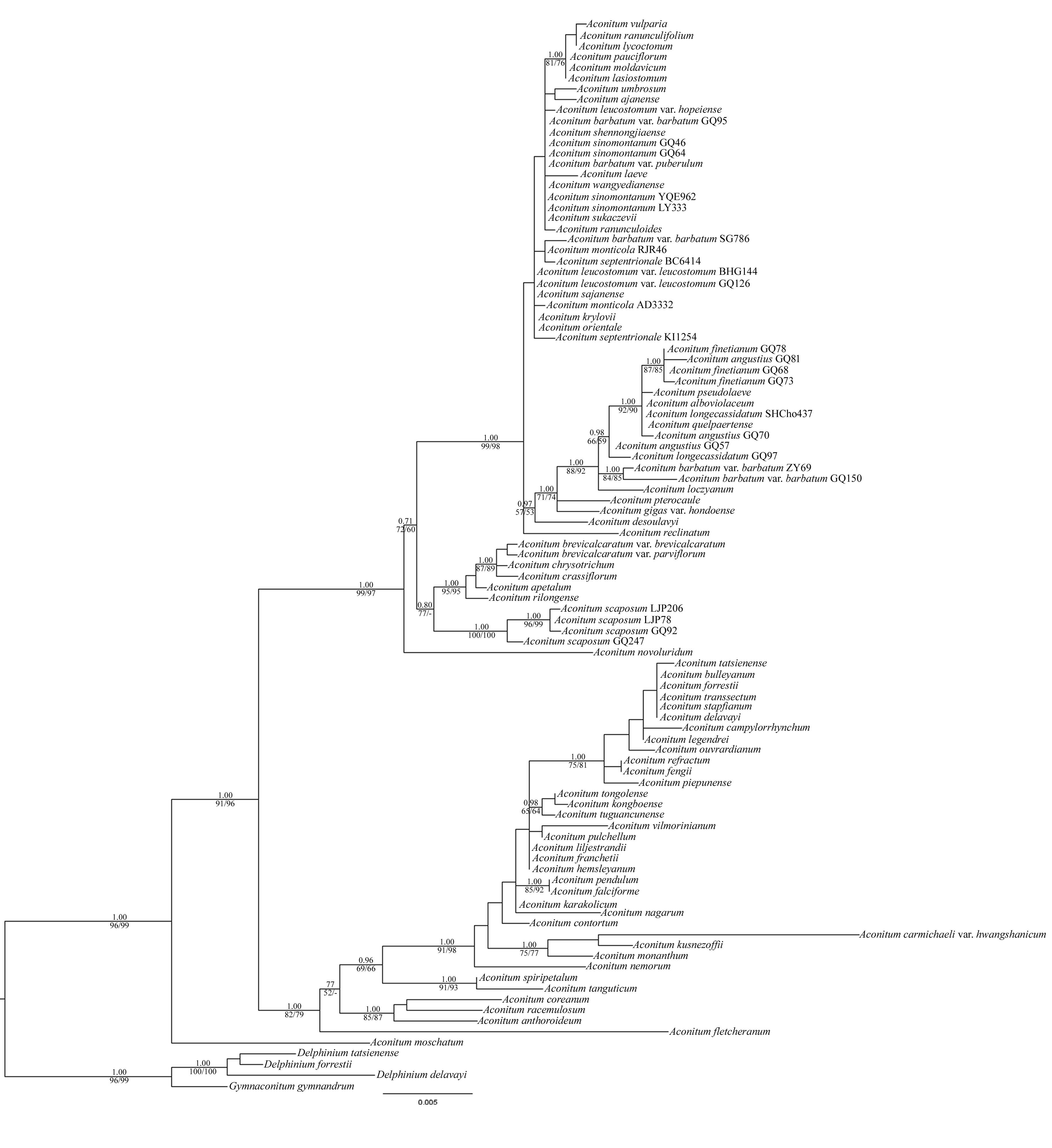
The analyses of three approaches (MP, ML, and BI) revealed largely congruent tree topologies and the ML tree is shown in Fig 1 with support values. The monophyly of the genus Aconitum was strongly supported (PP/MP/ML = 1.00/96%/99%). However, the monophyly of subgen. Lycoctonum as currently circumscribed was not supported since its two members, A. moschatum (the single species of sect. Galeata) and A. fletcheranum (the single species of sect. Fletcherum), did not cluster together with other members in the same clade. Aconitum moschatum appeared to be sister to all the other species of Aconitum (PP/MP/ML = 1.00/91%/96%), whereas A. fletcheranum formed a clade together with species of subgen. Aconitum (PP/MP/ML = 1.00/82%/79%) and held a sister position to this subgenus, although with weak support (PP/MP = 0.77/52%). Except for A. moschatum and A. fletcheranum, all the remaining taxa of subgen. Lycoctonum formed a well-supported clade (PP/MP/ML = 1.00/99%/97%). Aconitum novoluridum (the single species of sect. Altospermum) was the first diverging lineage of the clade. The section Lycoctonum was resolved as a monophyletic group, although with somewhat weak support (PP/MP/ML = 0.71/72%/60%). It was further resolved as two clades. In one of them (PP/MP = 0.80/77%), two subclades with strong support were resolved: (1) four accessions of A. scaposum Franch. (PP/MP/ML = 1.00/100%/100%); (2) A. brevicalcaratum var. brevicalcaratum, A. brevicalcaratum var. parviflorum Chen & Liu, A. chrysotrichum, A. crassiflorum, A. apetalum, and A. rilongense (PP/MP/ML = 1.00/95%/95%). The phylogenetic relationships within the other clade (PP/MP/ML = 1.00/99%/98%) were largely unresolved, although A. gigas var. hondoense, A. angustius (W. T. Wang) W. T. Wang, and two accessions of A. barbatum var. barbatum formed a subclade together with species of ser. Volubilia with weak support (PP/MP/ML = 0.97/57%/53%), and six European species formed a subclade with strong support (PP/MP/ML = 1.00/81%/76%).
Fig 1. Phylogenetic relationships in Aconitum obtained from an ML analysis of the combined cpDNA dataset.

Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses. “-” indicates that support is less than 50% bootstrap value. Tamura’s (1995) classification of subgen. Lycoctonum are shown on the right. Accessions with a different placement in the nrDNA tree are indicated in bold. The clade of subgen. Aconitum has been collapsed for saving space (see S1 Fig for the complete topology).
Phylogenetic analyses of nuclear ribosomal DNA sequence data
The p-value resulting from the ILD test between nrDNA ETS dataset and nrDNA ITS dataset showed no significant incongruence (Table 3). We thus combined them in the phylogenetic analyses. For the combined nrDNA dataset, information of each aligned DNA data, tree statistics for the MP analysis and the best-fit model of each region are given in Table 4.
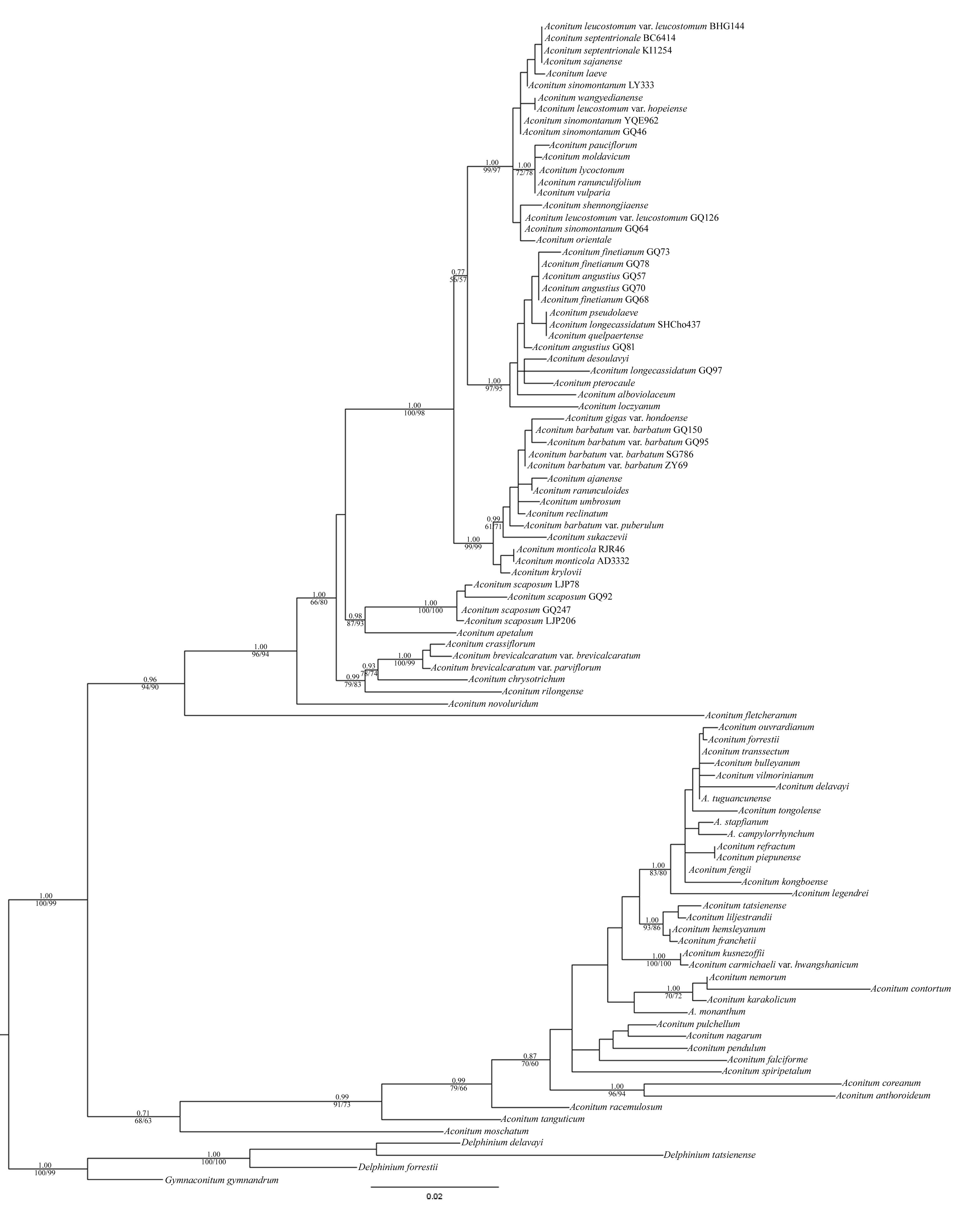
All the three analyses revealed approximately congruent tree topologies and the ML tree is shown in Fig 2 with support values. The monophyletic status of the genus Aconitum was confirmed with strong support values (PP/MP/ML = 1.00/100%/99%). However, as the case with cpDNA dataset, the monophyly of subgen. Lycoctonum as currently circumscribed was not supported. Its member A. moschatum was resolved as the sister to the subgenus Aconitum clade (PP/MP/ML = 0.99/91%/73%), although with relatively weak support (PP/MP/ML = 0.71/68%/63%). The remaining species of subgen. Lycoctonum formed a highly supported clade (PP/MP/ML = 0.96/94%/90%). Within this clade, A. fletcheranum and A. novoluridum were the first two species successively diverging. The section Lycoctonum was again resolved as a monophyletic group (PP/MP/ML = 1.00/66%/80%). Within this section, five clades with moderate to strong support were resolved. Comparison of the series classification by Tamura [1] on this section (Table 1) with our ML tree indicates that all the series recognized by him were para- or polyphyletic, although species of ser. Volubilia and of ser. Lycoctonia were almost nested together in a well-supported clade of their own respectively. The series Umbrosa was also not retrieved. It is noteworthy that the placements of A. apetalum, two accessions of A. barbatum var. barbatum, and A. gigas var. hondoense in the ML tree resulting from nrDNA dataset were different from their placements in the ML tree resulting from cpDNA dataset.
Fig 2. Phylogenetic relationships in Aconitum obtained from an ML analysis of the combined nrDNA dataset.

Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses. Tamura’s (1995) classification of subgen. Lycoctonum are shown on the right. Accessions with a different placement in the cpDNA tree are indicated in bold. The clade of subgen. Aconitum has been collapsed for saving space (see S2 Fig for the complete topology).
Phylogenetic analyses of combined cpDNA and nrDNA data
The ILD test indicated strong incongruence between nuclear markers and chloroplast markers with a p-value of 0.001 (Table 3). The results of the WSR and AU tests are provided in Table 5. For Aconitum gigas var. hondoense and two accessions of A. barbatum var. barbatum (ZY69 and GQ150), all the tests suggested significant differences between the relationships inferred respectively from cpDNA and nrDNA datasets, indicating that they contributed greatly to the conflict. These samples were thus included in phylogenetic analyses as two entries, once as a cpDNA-only entry and once as an nrDNA-only entry. For A. apetalum, A. fletcheranum and A. moschatum, at least the tests of the nrDNA dataset showed no significant difference between the relationships inferred from the cpDNA and nrDNA datasets, suggesting that they should not be the main causes of the conflict between the two datasets. We thus combined their nrDNA and cpDNA sequences for phylogenetic analyses. However, it is to be noted that, even after excluding the six samples mentioned above, the p-value of the ILD test between the cpDNA and nrDNA datasets was still less than 0.001 (Table 4). A visual comparison showed that both the original (containing all the six samples) and pruned (containing only each of the six samples) datasets suggested the same positions for the six samples (results not shown here), indicating that their placements were not influenced by the other five samples.
Table 5. P-values of the WSR and AU tests.
The pruned datasets are reconstructed from the original datasets by excluding Aconitum apetalum, two accessions of A. barbatum var. barbatum (ZY69 and GQ150), A. fletcheranum, A. gigas var. hondoense, and A. moschatum. Bold-faced values indicate rejection of the null hypothesis with 95% confidence.
| Dataset | Constraint topology (inferred from the other dataset) | WSR | AU |
|---|---|---|---|
| Pruned nrDNA dataset | |||
| + Aconitum moschatum | A. moschatum is sister to all the remaining species of Aconitum | 0.0956 | 0.311 |
| + A. fletcheranum | A. fletcheranum is sister to subgen. Aconitum | 0.0578 | 0.559 |
| + A. apetalum | A. apetalum forms a clade with the tetraploid species from the Hengduan Mountains region | 0.5637 | 0.307 |
| + A. gigas var. hondoense | A. gigas var. hondoense is a member of Clade C | 0.0010 | 0.011 |
| + A. barbatum var. barbatum ZY69 | A. barbatum var. barbatum ZY69 is a member of Clade C | 0.0033 | <0.001 |
| + A. barbatum var. barbatum GQ150 | A. barbatum var. barbatum GQ150 is a member of Clade C | 0.0021 | <0.001 |
| Pruned cpDNA dataset | |||
| + A. moschatum | A. moschatum is sister to subgen. Aconitum | 0.0253 | 0.056 |
| + A. fletcheranum | A. fletcheranum is sister to subgen. Lycoctonum | 0.1025 | 0.242 |
| + A. apetalum | A. apetalum is sister to A. scaposum | 0.0455 | 0.006 |
| + A. gigas var. hondoense | A. gigas var. hondoense is a member of Clade D | 0.0384 | <0.001 |
| + A. barbatum var. barbatum ZY69 | A. barbatum var. barbatum ZY69 is a member of Clade D | 0.0108 | <0.001 |
| + A. barbatum var. barbatum GQ150 | A. barbatum var. barbatum GQ150 is a member of Clade D | 0.0108 | <0.001 |
All the three analyses revealed approximately congruent tree topologies and the ML tree is shown in Fig 3 with support values. The genus Aconitum was confirmed again to be monophyletic with strong support values (PP/MP/ML = 1.00/100%/100%). The sister relationship of A. moschatum with the remaining species of Aconitum was supported by both BI and ML analyses (PP/ML = 1.00/91%). Aconitum fletcheranum was sister to subgen. Lycoctonum (PP/MP = 0.78/82%). The rest of species in subgen. Lycoctonum formed a well-supported clade (PP/MP/ML = 1.00/100%/100%). Aconitum novoluridum was the first split lineage. The monophyly of sect. Lycoctonum was well supported (PP/MP/ML = 1.00/93%/94%). Within this section, five major highly supported clades (Clades A–E) were resolved. These clades conformed largely to those revealed by the nrDNA dataset in taxon composition if regardless of those accessions with incongruent placements as revealed by nrDNA and cpDNA markers. All the series recognized by Tamura [1] were para- or poly-phyletic when superimposed on the ML tree (Fig 3), although Clade C consisted of taxa mostly from ser. Volubilia and Clade E was comprised of those mostly from ser. Lycoctonia. The series Umbrosa was also not recovered here, with its type species, A. umbrosum, deeply embedded in Clade D.
Fig 3. Phylogenetic relationships in Aconitum obtained from an ML analysis of the combined cpDNA and nrDNA dataset.

Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses. “-” indicates that support is less than 50% bootstrap value. Tamura’s (1995) classification and our new classification of subgen. Lycoctonum are shown on the right. Accessions with different placements between the cpDNA tree and the nrDNA tree are indicated in bold. The clade of subgen. Aconitum has been collapsed for saving space (see S3 Fig for the complete topology).
Discussion
Phylogenetic position and taxonomic status of Aconitum sect. Fletcherum
Aconitum sect. Fletcherum is unispecific, including only A. fletcheranum distributed in the eastern Himalayan region (Bhutan, southeastern Xizang in China, and Assam in northern India) [1–3, 9, 34–36]. When Tamura [34] established the section, he stated that the species was very peculiar in the short and scapose stem, the single flower terminal to the stem which was contrary to the indeterminate raceme in all other species of subgen. Lycoctonum, and in the navicular upper sepal and the 6–8 carpels as well. Lauener and Tamura [2], Wang [3], and Tamura [1] all accepted this section and its affiliation with subgen. Lycoctonum, but Kadota [9] transferred A. fletcheranum from subgen. Lycoctonum to his subgen. Tangutica, stressing that the nectary blade of the petal in this species, contrary to the other species (except for A. moschatum) within subgen. Lycoctonum but the same as members within subgen. Tangutica, was not provided with a tubular portion.
Most notably, the chromosome number of 2n = 12 in Aconitum fletcheranum, which represents a new base chromosome number of x = 6 for the genus Aconitum [10], is remarkably aberrant in the genus which otherwise has uniformly a base chromosome number of x = 8 [1, 13, 37–45]. Moreover, the karyotypic constitution of this species is also unique in the genus Aconitum. There are two largest metacentric chromosome pairs in this species, whereas in all the other diploid taxa of Aconitum there is only one such chromosome pair, with the second largest pair being submetacentric [10]. Hong et al. [10] considered that the base number of x = 6 in Aconitum may have originated from x = 8 (descending dysploidy) through asymmetric reciprocal translocations of some telocentric chromosomes. All these further suggest the abnormality of A. fletcheranum within subgen. Lycoctonum.
In our analyses, Aconitum fletcheranum is revealed to form a sister relationship to subgen. Aconitum in the cpDNA tree (PP/MP/ML = 1.00/82%/79% in Fig 1) and to all the remaining species of subgen. Lycoctonum in the nrDNA tree (PP/MP/ML = 0.96/94%/90% in Fig 2), respectively. There are several causes which may be invoked to account for this incongruence. All the topology tests show no significant difference between these two phylogenetic hypotheses, suggesting that stochastic errors, which are caused mainly by finite sequences used in studies, could not be ruled out confidently [46, 47]. Hybridization, probably combined with some other biological factors (reviewed in [48–52]), seems to be a more reasonable explanation for the incongruence of A. fletcheranum in view of its conflicting placements between the cpDNA and nrDNA trees, its aberrant chromosome number of 2n = 12 and unique karyotypic constitution. The signs of hybridization are nevertheless not evident in the nrDNA sequences of A. fletcheranum. We have failed to amplify the ETS sequence of A. fletcheranum, but we have successfully sequenced the ITS sequences of five individuals of this species. The chromatograms all contain well-formed, distinctive single peaks (no overlapping peaks) with very little background “noise”, and the sequences are different by only two nucleotides. It is regrettable that only one population of this species has been available for the present study. Sampling of more populations and further analyses using single- or low-copy nuclear genes are expected to reveal a more comprehensive and convincing pattern for this incongruence. Systematic errors, such as long-branch attraction, which result primarily from model misspecification [46], seem to be less likely a cause for the incongruence of A. fletcheranum. This is indicated by the results that, for each dataset, all the analyses (MP, ML, and BI) suggest the same placement for A. fletcheranum and that the branch length is generally even. Moreover, the positions of A. fletcheranum are stable in the cpDNA and nrDNA trees, at least not influenced by A. apetalum, the two accessions of A. barbatum var. barbatum (ZY69 and GQ150), A. gigas var. hondoense, and A. moschatum.
Our analyses of the combined data place Aconitum fletcheranum as a sister to all the remaining species of subgen. Lycoctonum (PP/MP = 0.78/82% in Fig 3), the same position as suggested by the nrDNA data. Although further studies are still needed to disentangle its evolutionary history, A. fletcheranum should deserve a subgeneric status in the genus Aconitum given that it always occupies a sister position to subgen. Aconitum (Fig 1) or to all the remaining taxa in subgen. Lycoctonum (Figs 2 and 3) and that it has a unique array of morphological and cytological characters.
Our molecular work lends no support to the establishment of Aconitum subgen. Tangutica. In all our analyses, A. tanguticum (Maxim.) Stapf, the type species of this subgenus, is always embedded in subgen. Aconitum (S1–S3 Figs), conforming to the results of previous molecular works on the genus Aconitum [15] or on the tribe Delphinieae [17]. Karyological data also indicate the membership of A. tanguticum within subgen. Aconitum. This species has the same chromosome number (2n = 16) and chromosome morphology as other taxa within subgen. Aconitum [44].
Phylogenetic position and taxonomic status of Aconitum sect. Galeata
Aconitum sect. Galeata is also unispecific, including only A. moschatum endemic in the alpine zone of Kashmir [1, 2, 9, 34, 53]. Tamura [1, 34] and Lauener and Tamura [2] accepted this section and its affiliation with subgen. Lycoctonum, although they stated that A. moschatum was very particular within subgen. Lycoctonum by having the navicular or depressed galeate helmet broader than long, lurid purple flowers, and the very obtuse spur of the petal, not opposite to the labium but a continuation of the stalk. Kadota [9], however, pointed out that A. moschatum should not be placed in subgen. Lycoctonum, stressing that its nectary blade of the petal, contrary to the other species (except for A. fletcheranum) within the subgenus, was not equipped with a tubular portion. He left the problem aside for a further study. The chromosome number of A. moschatum was reported to be 2n = 16 [54] but regrettably the chromosome morphology has yet been unknown.
In our ML tree resulting from cpDNA dataset, Aconitum moschatum is revealed as the sister to all other members of the genus Aconitum with strong support (PP/MP/ML = 1.00/91%/96% in Fig 1). In the ML tree resulting from nrDNA dataset, however, it is resolved as the sister to subgen. Aconitum, but with weak support (PP/MP/ML = 0.71/68%/63% in Fig 2). All the topology tests, except for the WSR test of the cpDNA data, show no significant difference between the two phylogenetic hypotheses (Table 5). This may suggest that the incongruence of A. moschatum between cpDNA and nrDNA is not a “hard” one [55]. Similar to the case with A. fletcheranum, systematic errors cannot be invoked to account for this incongruence due to the stable positions suggested by all the analyses (the MP, ML, and BI analyses, and the analyses based on complete or pruned datasets) and the relatively short branch length as well. We thus combine the nrDNA and cpDNA data of A. moschatum for phylogenetic analyses. The analyses of combined data show that this species is the sister to all other species of Aconitum (PP/ML = 1.00/91% in Fig 3), the same relationship as suggested by cpDNA dataset.
In summary, our phylogenetic analyses indicate that Aconitum moschatum is not closely related to any of the major groups of Aconitum revealed here. Taking into account its unique array of morphological characters, we consider it justifiable to remove sect. Galeata from subgen. Lycoctonum and treat it as an independent subgenus of its own in the genus Aconitum.
Phylogenetic position and taxonomic status of Aconitum sect. Alatospermum
Aconitum sect. Alatospermum is also unispecific, including only A. novoluridum distributed in the eastern Himalayan region (Bhutan, southeastern Xizang in China, Sikkim in northern India, and Nepal) [1–3, 34, 35]. When establishing the section under subgen. Lycoctonum based on this species, Tamura [34] pointed out that the species was clearly distinguishable from other species of the subgenus by the depressed hemi-elliptic helmet gradually descending into the long beak and much broader than long, the hammer-shaped petals with the nectary blade at a right angle to the stalk and longer than the stalk, the longitudinally winged seeds along the three ridges, and the lurid, reddish, brownish red or purple flowers. This section has since been recognized by Lauener and Tamura [2], Tamura [1], and Kadota [35] but rejected by Wang [3], who placed A. novoluridum in sect. Lycoctonum (“Paraconitum”).
Both the sectional status and subgeneric affiliation of Aconitum novoluridum are strongly supported by our molecular analyses. In our ML trees resulting from cpDNA (Fig 1), nrDNA (Fig 2) or the combined cpDNA and nrDNA dataset (Fig 3), this species is always resolved as the first split lineage in subgen. Lycoctonum and forms a sister relationship to sect. Lycoctonum.
Tamura [1] regarded Aconitum sect. Alatospermum, which has the seeds without transverse squamae and the hemi-elliptic upper sepal, as a primitive group in subgen. Lycoctonum, listing it as the first section in this subgenus (Tamura and Lauener [6] had previously listed sect. Galeata (“Galeatum”) as the first section). The primitive condition of A. novoluridum in seed morphology is confirmed by Kong et al. [56]. According to Hong et al. [10], the karyotype of A. novoluridum is the most symmetric in the subgenus and thus very probably represents a primitive condition. The primitive condition of A. novoluridum in subgen. Lycoctonum is also supported by our molecular work. This species is always the first split lineage in the subgenus in our ML trees resulting from different datasets.
Our molecular work, therefore, strongly favors the treatment of Aconitum novoluridum as an independent section within subgen. Lycoctonum, i.e. sect. Alatospermum, lending no support to its placement in sect. Lycoctonum. Evidence from morphology, karyology, and molecular phylogeny all suggests the primitive condition of sect. Alatospermum within subgen. Lycoctonum.
Phylogenetic relationships within Aconitum sect. Lycoctonum and its reclassification
Our molecular work indicates that Aconitum sect. Lycoctonum is a monophyletic group within subgen. Lycoctonum (PP/MP/ML = 0.71/72%/60% in Fig 1; PP/MP/ML = 1.00/66%/80% in Fig 2; PP/MP/ML = 1.00/93%/94% in Fig 3), agreeing with the results of past molecular analyses involving this section [4, 17]. All the nine series recognized by Tamura [1] and ser. Umbrosa proposed additionally by Kadota [7] in this section, however, are revealed to be poly- or para-phyletic in our analyses of the combined cpDNA and nrDNA dataset (Fig 3). Five major clades are nevertheless recovered within the section, and they can be conveniently treated at series rank: ser. Crassiflora, ser. Scaposa, ser. Volubilia, ser. Longicassidata, and ser. Lycoctonia.
Aconitum ser. Scaposa includes only A. scaposum (Clade A: PP/MP/ML = 1.00/100%/100% in Fig 3). This species, fairly widespread in central and southwestern China [3] and recently reported, under the name A. chloranthum Hand.-Mazz., to occur also in Bhutan [35], shows great variation in the relative development of basal and cauline leaves, the leaf shape, pedicel pubescence, and flower color. Several varieties were once described in this species, and some of them were even recognized as independent species [2, 3, 57]. Yang [11] and Luo and Yang [12] treated A. scaposum as a polymorphic species including all the varieties and species. The four accessions that we have chosen in our molecular analyses represent two types of plants in respect of the relative development of basal and cauline leaves: two accessions (LJP206 from Sichuan, GQ92 from Gansu) have cauline leaves nearly aggregated in the middle part of the stem, while the other two accessions (GQ247 from Hubei, LJP78 from Chongqing) have cauline leaves nearly equally distantly arranged along the stem. All these accessions are nested together with each other in the same clade.
Aconitum ser. Crassiflora includes six taxa: A. apetalum, A. brevicalcaratum var. brevicalcaratum, A. brevicalcaratum var. parviflorum, A. crassiflorum, A. chrysotrichum, and A. rilongense (Clade B: PP/MP/ML = 1.00/52%/84% in Fig 3). The close affinity of the latter five taxa is also indicated by gross morphology, geographical distribution, and in particular, karyology. They are very similar to each other in general appearance [11, 12, 58] and all concentrated in the southern part of the Hengduan Mountains region in southwestern China [10]. They are all tetraploid (2n = 32) and share similar chromosome size and karyotype constitution [10]. The close affinity between A. scaposum (a diploid species with 2n = 16) and A. crassiflorum as previously regarded by Tamura [1] is not supported by our molecular analyses. In fact, no diploid species are clustered with these five tetraploid taxa. All the five tetraploid taxa in this series might have originated through only one polyploidization event, but we have been unable to ascertain their type of polyploidy (autopolyploidy vs. allopolyploidy) and parental origin from morphological, karyological and molecular data currently available.
Aconitum apetalum is distributed in central Asia (Xinjiang in China, Kazakhstan, and Tajikistan). It is readily distinguishable by its long, many-flowered raceme, very small flowers, and petals with a short, capitate spur, and the upper sepal narrowly cylindrical with a prominent peak [1, 6]. Tamura [1] placed it in ser. Micrantha together with A. brevicalcaratum, A. chrysotrichum, and A. sajanense Kumin mainly due to their short spur of petals, but in our molecular analyses A. brevicalcaratum and A. chrysotrichum are shown to be members in ser. Crassiflora, while A. sajanense, a diploid species with 2n = 16 [59], is shown to be a member in ser. Lycoctonia (Fig 3). The results of our molecular analyses of A. apetalum are somewhat similar to those of A. fletcheranum. As a hexaploid species (2n = 48), A. apetalum is nested with those tetraploid taxa (2n = 32) within ser. Crassiflora in the cpDNA tree (PP/MP/ML = 1.00/95%/95% in Fig 1) and sister to A. scaposum (2n = 16) in the nrDNA tree (PP/MP/ML = 0.98/87%/93% in Fig 2). This seems to strongly support a hybrid origin followed by subsequent polyploidization for A. apetalum. However, the chromatograms of ITS and ETS of A. apetalum, with only distinctive single peaks, show no obvious evidence of hybridization. The WSR and AU tests of the nrDNA data do not significantly reject the hypothesis inferred from the cpDNA data, further indicating the probable existence of a stochastic error. We thus also combine the cpDNA and nrDNA sequences of A. apetalum for phylogenetic analyses. The analyses place A. apetalum sister to all the tetraploid taxa from the Hengduan Mountains region, a relationship similar to that suggested by the cpDNA data. We therefore tentatively refer A. apetalum to ser. Crassiflora. It is worth mentioning that our molecular results do not support the reduction of A. monticola Steinb., also a central Asian species, to the synonymy of A. apetalum [60]. Both of them are nested in different clades (Fig 3).
Aconitum ser. Volubilia defined by Tamura and Lauener [6] and Tamura [1] is largely supported by our molecular work (Figs 1–3). Nine species traditionally classified in this series are clustered in this clade together with A. angustius (Clade C: PP/MP/ML = 1.00/92%/94% in Fig 3), a species mainly distributed in southeastern and central China and previously placed, under the name A. sinomontanum Nakai var. angustius W. T. Wang, in ser. Lycoctonia [6]. Aconitum angustius has long been considered to be most closely related to A. sinomontanum [57] or even treated as a variety of it [3, 61, 62]. This is not supported by our molecular results. The two taxa are nested in different clades (Fig 3). In fact, Gao et al. [63] and Hong et al. [10] has previously revealed that A. angustius is distinct from A. sinomontanum in ploidy level (tetraploid with 2n = 32 vs. diploid with 2n = 16) and that the former is morphologically more closely similar to A. finetianum Hand.-Mazz., a diploid species mainly distributed in southeastern China and placed (as a synonym of the Japanese species A. pterocaule Koidz.) in ser. Volubilia by Tamura and Lauener [6]. Moreover, as pointed out by Gao et al. [63], some specimens of A. angustius from Guizhou and Hubei, China, had been previously misidentified as A. loczyanum Rapaics or A. pterocaule (both belonging to ser. Volubilia) by Handel-Mazzetti [64] and Tamura and Lauener [6]. From its karyotypic constitution A. angustius may be of an allopolyploid origin [63]. Considering their close morphological similarity and somewhat overlapping geographical distribution we infer that one of the parents of A. angustius is very likely A. finetianum. The origin of A. angustius is an interesting problem worthy of further investigations.
Aconitum ser. Longicassidata (Clade D: PP/MP/ML = 1.00/90%/88% in Fig 3) is the most complex group which comprises species from five different series classified by Tamura [1]: ser. Longibracteolata, ser. Longicassidata, ser. Lycoctonia, ser. Ranunculoidea, and ser. Reclinata. The series Umbrosa proposed by Kadota [7] should also be transferred to here. Among them, ser. Longibracteolata and ser. Reclinata are both unispecific. The close similarity between A. monticola and A. krylovii Steinb. (ser. Lycoctonia) in morphology has been noted by Steinberg [65]. Aconitum barbatum and A. kirinense Nakai (ser. Longicassidata) have also been considered to be closely related to each other [6]; Handel-Mazzetti [64] even placed the latter in synonymy with the former. Most notably, while Tamura and Lauener [6] established ser. Ranunculoidea to accommodate A. ranunculoides Turcz. ex Ledeb. and A. ajanense Steinb., they pointed out that the latter species was also near to A. umbrosum (ser. Lycoctonia), a species which they considered to be closely related to A. gigas var. hondoense (= A. iinumae Kadota). These opinions are largely supported by our molecular results (Fig 3). Aconitum reclinatum A. Gray is the only representative of subgen. Lycoctonum in the New World [1, 6]. Our molecular phylogeny reveals it to be closely related to several Asian species, suggesting that it may have migrated from Asia to eastern North America.
It is noteworthy that two accessions of Aconitum barbatum var. barbatum (GQ150 and ZY69) and A. gigas var. hondoense show significant incongruences between the nrDNA and cpDNA datasets. The former taxon is widespread in northeastern and central China and the Far East of Russia (Primorye). Four accessions of it are included in our analyses, including one (GQ95) from Shaanxi in central China, two (GQ150 and ZY69) from Jilin in northeastern China, and one (SG786) from Primorye. On the nrDNA tree (Fig 2), the four accessions cluster together with A. gigas var. hondoense and then are nested within ser. Longicassidata (Clade D in Fig 3). On the cpDNA tree (Fig 1), GQ95 and SG786 are still grouped with species of ser. Longicassidata (although this series do not form a clade in the cpDNA tree), whereas GQ150 and ZY69 cluster with species of ser. Volubilia (Clade C in Fig 3). All the topology tests indicate significant incongruence between the two phylogenetic hypotheses of GQ150 and ZY69 (Table 5). However, both GQ150 and ZY69 are typical A. barbatum var. barbatum in morphology, showing no obvious difference from GQ95 and SG786, and no intermediates between A. barbatum var. barbatum and any species of ser. Volubilia have thus far been found. A visual examination of the nrDNA sequences of A. barbatum var. barbatum shows that there are no more than three nucleotide differences among its four accessions, and those of ZY69 (with incongruent placements) and SG786 (with congruent placement) are even totally identical. This seems to be a typical pattern of gene tree incongruence caused by chloroplast capture: the cytoplasm of GQ150 and ZY69 has been replaced by that of a certain member of ser. Volubilia probably via introgression [66–68]. This hypothesis is further indicated by the geographical distribution of A. barbatum var. barbatum and species of ser. Volubilia. Members in ser. Volubilia occur mainly in southeastern and northeastern China, the Korean Peninsula, Japan, and the Far East of Russia, with their distribution largely overlapping with that of A. barbatum var. barbatum. In the field A. barbatum var. barbatum is often found to grow in the neighborhood of some species of ser. Volubilia, e.g., A. alboviolaceum Kom. (pers. observ.).
Similar to the case with the two accessions of Aconitum barbatum var. barbatum, GQ150 and ZY69, A. gigas var. hondoense is nested in ser. Longicassidata on the nrDNA tree (Fig 2) but in ser. Volubilia on the cpDNA tree (Fig 1). The topology tests also indicate significant incongruence between the two phylogenetic hypotheses (Table 5). Regrettably only one accession of this variety is included in this study. A more extensive sampling is needed to explore the overall phylogenetic pattern of this taxon and the exact cause(s) of the incongruence of the accession sampled here. From a morphological perspective, A. gigas var. hondoense is closely related to A. umbrosum, and thus should be placed in ser. Longicassidata construed here.
Aconitum ser. Lycoctonia (Clade E: PP/MP/ML = 1.00/88%/91% in Fig 3) includes 14 taxa sampled here. Among them, 11 have been previously placed within this series by Tamura and Lauener [6] and Tamura [1], but the remaining three placed within other series by them, one each respectively in ser. Micrantha (A. sajanense), ser. Laevia (A. laeve Royle), and ser. Longicassidata (A. lasiostomum Reichb. ex Besser). Significantly, the eight European species sampled are all nested in this clade. Six of them (A. lasiostomum, A. lycoctonum, A. moldavicum Hacquet, A. pauciflorum Host, A. ranunculifolium Reichb., and A. vulparia Reichb. ex Spreng.) cluster together in a strongly supported subclade (PP/MP/ML = 1.00/83%/88% in Fig 3). Although Tamura and Lauener [6] placed A. lasiostomum in ser. Longicassidata, they noted that this species was not typical of the series and might approach ser. Lycoctonia. Utelli et al. [14], based on their analyses on the A. lycoctonum species complex from Europe and the Caucasus Mountains using the chloroplast intergenic spacer psbA-trnH and nuclear ITS region, also considered that A. lasiostomum should belong to ser. Lycoctonia.
Taxonomic treatment
The first formal phylogeny-based classification of Aconitum subgen. Lycoctonum is presented below (also see Fig 3), which involves segregating both sect. Galeata and sect. Fletcherum from this subgenus as two independent subgenera of their own within the genus Aconitum, reinstating one series (ser. Crassiflora) and abolishing six (ser. Laevia, ser. Longibracteolata, ser. Micrantha, ser. Ranunculoidea, ser. Reclinata, and ser. Umbrosa) within sect. Lycoctonum. The series affiliation of some species within the section is adjusted accordingly. We include only the more significant synonyms, for a more complete synonymy see Tamura [1].
Aconitum subgen. Galeata (Rapaics) Y. Hong & Q. E. Yang, comb. & stat. nov.
Type. A. moschatum (Brühl ex Duthie) Stapf
Basionym. A. sect. Galeata Rapaics in Növényt. Közlem. 6: 140. 1907.
Description. Stem usually unbranched with a few cauline leaves. Basal leaves 5–7-fid, segments obovate-cuneate, incised-dentate. Inflorescence racemiform, many- flowered. Flowers lurid-purple; upper sepal navicular or depressed galeate; spur of the petal very obtuse. Carpels 3. Seeds broadly squamate. chromosome number: x = 8.
Species and Distribution. Unispecific, endemic in the alpine zone of Kashmir.
Note. Our molecular work indicates that this subgenus is the earliest diverging lineage of Aconitum and sister to all the remaining species of the genus (Fig 3). Morphologically it is readily distinguishable from subgen. Aconitum by having rhizomes. From subgen. Lycoctonum it differs in the upper sepal navicular or depressed galeate, broader than long, and in the nectary blade of the petal not provided with a tubular portion. From subgen. Fletcherum it differs in the inflorescence racemiform, many-flowered.
Aconitum subgen. Fletcherum (Tamura) Y. Hong & Q. E. Yang, comb. & stat. nov.
Type. A. fletcheranum G. Taylor
Basionym. A. sect. Fletcherum Tamura in Sci. Rep. Osaka Univ. 15: 30. 1966.
Description. Stem short, subscapose. Basal leaves rosulate, 5-partite, segments incised-lobate; cauline leaves usually 2, bract-like, 3-partite into entire lobes, remarkably sheathy at the base. Flower single, terminal to the stem; blue-violet; upper sepal navicular; spur of petal obtuse, labium inconspicuous. Carpels 6–8. Seeds unknown. Chromosome number: x = 6.
Species and Distribution. Unispecific, in the alpine zone of Bhutan, southeastern Xizang in China, and Assam in India.
Aconitum subgen. Lycoctonum (DC.) Peterm. in Deutschl. Fl. 16. 1846.
Type. A. lycoctonum L.
Basionym. A. sect. Lycoctonum DC. in Syst. Nat. 1: 367. 1817.
Description. Stem branched or unbranched, leafy. Leaves palmatifid, palmatipartite, or palmatisect. Inflorescence racemose or paniculate. Flowers blue, purple, yellow or white; upper sepal hemielliptic, cylindrical, conical to tubular, usually longer than broad; spur of petal obtuse, capitate or elongate, long or short, or absent. Carpels 3. Seeds longitudinally alate along 3 ridges, hardly squamate or transversely squamate. Chromosome number: x = 8.
Species and Distribution. Approximately 44 species in two sections distributed in Eurasia, northern Africa, and eastern North America.
Note. The subgenus defined here includes only two sections, sect. Alatospermum and sect. Lycoctonum, not identical to the concept of previous authors. Tamura and Lauener [6] and Tamura [1], placed another two sections, sect. Fletcherum and sect. Galeata, in this subgenus.
Section 1. Aconitum sect. Alatospermum Tamura in Sci. Rep. Osaka Univ. 15: 30. 1966.
Type. A. novoluridum Munz
Description. Stem usually unbranched, leafy. Leaves 5-fid to -partite, segments incised-dentate. Inflorescence racemiform, many-flowered. Flowers lurid reddish or purple; upper sepal hemielliptic with a broad beak; petal hammer-shaped, spur opposite to the lamina, produced at right angles to the short, erect, thick stalk. Carpels 3. Seeds longitudinally alate along 3 ridges, hardly squamate.
Species and Distribution. Unispecific, in Bhutan, southeastern Xizang in China, Sikkim in India, and Nepal.
Section 2. Aconitum sect. Lycoctonum DC. in Syst. Nat. 1: 367.1817.
Type. A. lycoctonum L.
Description. Stem branched or unbranched, leafy. Leaves palmatifid, palmatipartite, or palmatisect. Inflorescence racemose or paniculate. Flowers blue, purple, yellow or white; upper sepal cylindrical, conical or high-galeate, usually longer than broad, often recurved at the apex; spur of petal obtuse, capitate or elongate, long or short, or absent. Carpels 3. Seeds transversely squamate.
Species and Distribution. Approximately 43 species in six series distributed in Eurasia, northern Africa, and eastern North America.
Note. At this stage it is very difficult to give an exact estimation of the number of species in this section because most of the species are most highly variable morphologically and different authors often have quite different concepts of species. Taxonomic revision at species level is still badly needed for some series of this section, e.g., ser. Scaposa, ser. Volubilia, ser. Longicassidata, and ser. Lycoctonia.
Series 1. Aconitum ser. Scaposa W. T. Wang in Acta Phytotax. Sin. Addit. 1: 60. 1965. Type. A. scaposum Franch.
Description. Stem scapose. Leaves reniform-pentagonal, 3-parted nearly to midvein, central lobe rhombic or cuneate-rhombic, lateral lobes obliquely flabellate, unequally 2-lobed. Inflorescence racemose, many-flowered, lax; pedicels long, usually spreading hairy; bracteoles 2, ovate or oblong, near the base of pedicels. Flowers purplish, greenish or pale-yellow; upper sepal cylindric; petaline lip linear, spur coiled, longer than lip.
Species and Distribution. Unispecific and yet highly polymorphic, in Bhutan and central and southwestern China.
Series 2. Aconitum ser. Crassiflora Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 123. 1979.
Type. A. crassiflorum Hand.-Mazz.
Synonym. A. ser. Brevicalcarata Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 443. 1979.
Type. A. brevicalcaratum (Finet & Gagnep.) Diels
Synonym. A. ser. Micrantha Steinb. ex Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 442. 1979.
Type. A. apetalum (Huth) B. Fedtsch. ex Steinb.
Description. Stem erect, scapose or subscapose, leafy, branched to inflorescences. Leaves obicular-reniform or reniform, 3-parted slightly beyond middle, central lobe cuneate-obtrapezoid, obovate-rhombic or rhombic, 3-fid, lateral lobes obliquely flabellate, unequally 2- or 3-fid. Inflorescence racemose, many-flowered; pedicels spreading or appressed pubescent; bracteoles 2, linear, usually near the base of pedicels. Flowers blue, blue-purple, dull yellow or yellowish; upper sepal high-galeate, cylindrical-galeate or cylindric; petal lip linear, conspicuous, spur slightly incurved, circinate or ecalcarate.
Species and Distribution. Five species, A. apetalum, A. brevicalcaratum, A. chrysotrichum, A. crassiflorum, and A. rilongense, in the southern part (western Sichuan and northeastern Yunnan) of the Hengduan Mountains region in southwestern China, Xinjiang, China and Kazakhstan.
Series 3. Aconitum ser. Volubilia (Steinb. ex Nakai) Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 434. 1979.
Type. A. alboviolaceum Kom.
Description. Stem erect, decumbent or twining. Leaves pentagonal-reniform, 3-parted slightly beyond middle, central lobe rhombic-obtrapezoid or broadly rhombic, lateral lobes obliquely flabellate, unequally 2- or 3-fid. Inflorescence racemose, simple or branched, elongate or more or less aggregate; pedicels short, spreading or appressed pubescent; bracteoles 2, linear, below the middle or near the base of pedicels. Flowers dilute purple or white; upper sepal cylindrical, more or less recurved at the tip; petal lip linear, conspicuous, spur curved or coiled, longer than the lip.
Species and Distribution. Approximately 11 species, mainly in eastern Asia (China, Korea, and Japan), e.g., Aconitum alboviolaceum, A. angustius, A. desoulavyi Kom., A. finetianum, A. locyzanum Rapcs., A. longecassidatum Nakai, A. pseudolaeve Nakai, A. pterocaule, and A. quelpaertense Nakai.
Note. This series has been fairly well defined previously by Tamura and Lauener [6] and Tamura [1]. Although A. pteropus Nakai, a species from Korea, is not included in our molecular analyses because of unavailability of DNA material, we agree with Tamura and Lauener [6] that it should belong to this series from a morphological perspective. Morphologically this species is closely similar to A. pterocaule.
Series 4. Aconitum ser. Longicassidata (Steinb. ex Nakai) Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 444. 1979.
Type. A. barbatum Pers.
Synonym. A. ser. Reclinata Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh. 37: 434. 1979. syn. nov.
Type. A. reclinatum A. Gray
Synonym. A. ser. Logibracteolata (Steinb. ex) Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 442. 1979. syn. nov.
Type. A. sukaczevii Steinb.
Synonym. A. ser. Ranunculoidea (Steinb. ex) Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 449. 1979. syn. nov.
Type. A. ranunculoides Turcz. ex Ledeb.
Synonym. A. ser. Umbrosa (Steinb. ex Nakai) Kadota in Fl. Jpn. IIa: 268. 2006. syn. nov.
Type. A. umbrosum (Korsh.) Kom.
Description. Stem erect, subscapose or leaning, sometimes trailing. Leaves obicular-reniform or reniform-pentagonal, 3-parted or subpedatifid to partite, central leaf segment broadly rhombic or cuneate-rhombic, sometimes 3-parted nearly to midvein, lateral segments obliquely flabellate. Inflorescence lax or densely racemiform; pedicels spreading or appressed pubescent; bracteoles below the middle or near the base of pedicels. Flowers yellow, white or pale blue-violet; upper sepal elongate conical, cylindrical or tubulose, often with a beak; petal lip linear, conspicuous, spur short or elongate, circinate or semi-coiled, shorter than, nearly as long as or longer than the lip.
Species and Distribution. Approximately 11 species, in eastern and central Asia and eastern North America, e.g., Aconitum ajanense, A. barbatum, A. gigas, A. krylovii, A. monticola, A. ranunculoides, A. reclinatum, A. sukaczevii Steinb., and A. umbrosum.
Note. This series is greatly expanded to include several species which have been previously placed in four other series by Tamura and Lauener [6] and Tamura [1]: Aconitum reclinatum (the single species in ser. Reclinata), A. sukaczevii (the single species in ser. Longibracteolata), A. ajanense, A. ranunculoides (both in ser. Ranunculoidea), A. umbrosum, A. monticola and A. krylovii (all in ser. Lycoctonia). Another two species, including A. crassifolium Steinb. (ser. Ranunculoidea) from the Far East of Russia and A. puchonroenicum Uyeki & Satake (ser. Lycoctonia) from Korea, should also belong to this series, but this needs to be verified by using molecular data.
Among the nine species (Aconitum asahikawaense Kadota, A. gigas, A. hiroshi-igarashii Kadota, A. ikedae Kadota, A. mashikense, A. soyaense Kadota, A. tatewaki Miyabe, A. umezawae Kadota, A. umbrosum) placed by Kadota [7, 8] in his ser. Umbrosa, only A. gigas var. hondoense and A. umbrosum are included in our molecular analyses. It is somewhat strange to us that when Kadota [8] described A. hiroshi-igarashii as new, he compared it with both A. pterocaule and A. gigas, the former of which is a member of ser. Volubilia. Judging from their morphological characters, A. tatewaki and all the above-mentioned species described as new by Kadota are very closely related to A. gigas, and their identities need further determination.
Series 5. Aconitum ser. Lycoctonia Tamura & Lauener in Not. Bot. Gard. Edinb. 37: 451. 1979.
Type. A. lycoctonum L.
Synonym. A. ser. Laevia Tamura & Lauener in Notes Roy. Bot. Gard. Edinburgh 37: 433. 1979. syn. nov.
Type. A. laeve Royle
Description. Stem erect, sometimes decumbent. Leaves obicular-reniform, 3-parted or subpedatifid to partite, central lobe broadly rhombic or narrowly cuneate-rhombic, 3-fid, lateral lobes obliquely flabellate, unequally 3-fid. Inflorescence racemose, many flowered; pedicels spreading or appressed pubescent; bracteoles 2, below the middle or near the base of pedicels. Flowers yellow, purple or blue; upper sepal elongate conical or cylindrical; petal lip linear, conspicuous, spur elongate, curved or circinate, nearly as long as or longer than the lip.
Species and Distribution. Approximately 15 species, in Europe, Asia and northern Africa, e.g., Aconitum laeve, A. lasiostomum, A. leucostomum Vorosh., A. lycoctonum, A. moldavicum, A. orientale Mill., A. pauciflorum, A. ranunculifolium, A. sajanense, A. septentrionale, A. shennongjiaense Q. Gao & Q.E. Yang, A. sinomontanum, A. vulparia, and A. wangyedianense Y.Z. Zhao.
Note. Although the Iranian species Aconitum iranshahri H. Riedl is not included in our molecular analyses, it should belong to this series from its close morphological similarity with A. orientale.
Supporting information
Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses. “-” indicates that support is less than 50% bootstrap value.
(JPG)
{kind=link}
Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses.
(JPG)
{kind=link}
Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses. “-” indicates that support is less than 50% bootstrap value.
(JPG)
{kind=link}
(XLSX)
Acknowledgments
We are grateful to two anonymous reviewers and Prof. Liu, academic editor of PLOS ONE, for their valuable comments on the manuscript. We thank Dr. T. Ohi-Toma (University of Tokyo, Japan), Dr. Y. D. Kim (Hallym University, South Korea) and Dr. I. Illarionova (Komarov Botanical Institute, Russia) for contributing plant material.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This study was supported by the National Natural Science Foundation of China [www.nsfc.gov.cn/] grant no 31470303.
References
- 1.Tamura M. Ranunculaceae In: Hiepko P, editor. Die Natürlichen Pflanzenfamilien, Aufl. II. 17a (4). Berlin: Duncker & Humblot; 1995. pp. 272–312. [Google Scholar]
- 2.Lauener LA, Tamura M. A synopsis of Aconitum subgenus Paraconitum: I. Notes from the Royal Botanic Garden Edinburgh. 1978; 37: 113–124. [Google Scholar]
- 3.Wang WT. Aconitum L. In: Anonymous editor. Flora Reipublicae Popularis Sinicae, Vol. 27 Beijing: Science Press; 1979. pp. 113–326. [Google Scholar]
- 4.Wang W, Liu Y, Yu SX, Gao TG, Chen ZD. Gymnaconitum, a new genus of Ranunculaceae endemic to the Qinghai-Tibetan Plateau. Taxon. 2013; 62: 713–722. [Google Scholar]
- 5.Xiao PG, Wang FP, Gao F, Yan LP, Chen DL, Liu Y. A pharmacophylogenetic study of Aconitum L. (Ranunculaceae) from China. Acta Phytotaxonomica Sinica. 2006; 44: 1–46. [Google Scholar]
- 6.Tamura M, Lauener LA. A synopsis of Aconitum subgenus Lycoctonum: II. Notes from the Royal Botanic Garden Edinburgh. 1979; 37: 431–466. [Google Scholar]
- 7.Kadota Y. Aconitum L. In: Iwatsuki K, Boufford DE, Ohba H, editors. Flora of Japan, Vol. IIa Tokyo: Kodansha Ltd; 2006. pp. 267–285. [Google Scholar]
- 8.Kadota Y. Systematic studies of Asian Aconitum (Ranunculaceae) XIV. Four new species of subgenus Lycoctonum from Hokkaido, northern Japan. The Journal of Japanese Botany. 2012; 87: 1–21. [Google Scholar]
- 9.Kadota Y. Systematic studies of Asian Aconitum (Ranunculaceae) VIII. Subgenus Tangutica, the fourth new subgenus of the genus Aconitum and a new species belonging to the subgenus Tangutica. The Journal of Japanese Botany. 2001; 76: 183–192. [Google Scholar]
- 10.Hong Y, Gao Q, Luo Y, Luo JP, Zhang Y, Yuan Q, et al. Karyology of Aconitum subgenus Lycoctonum (Ranunculaceae) from China, with a report of the new base chromosome number x = 6 in the genus Aconitum. Nordic Journal of Botany. 2016; 34: 441–454. [Google Scholar]
- 11.Yang QE. Taxonomic notes on some species of Aconitum L. (Ranunculaceae) from Yunnan, China. Acta Phytotaxonomica Sinica. 1999; 37: 545–590. [Google Scholar]
- 12.Luo Y, Yang QE. Taxonomic revision of Aconitum (Ranunculaceae) from Sichuan, China. Acta Phytotaxonomica Sinica. 2005; 43: 289–386. [Google Scholar]
- 13.Yuan Q, Yang QE. Polyploidy in Aconitum subgenus Lycoctonum (Ranunculaceae). Botanical Journal of the Linnean Society. 2006; 150: 343–353. [Google Scholar]
- 14.Utelli AB, Roy BA, Baltisberger M. Molecular and morphological analyses of European Aconitum species (Ranunculaceae). Plant Systematics and Evolution. 2000; 224: 195–212. [Google Scholar]
- 15.Luo Y, Zhang FM, Yang QE. Phylogeny of Aconitum subgenus Aconitum (Ranunculaceae) inferred from ITS sequences. Plant Systematic and Evolution. 2005; 252: 11–25. [Google Scholar]
- 16.Kita Y, Ueda K, Kadota Y. Molecular phylogeny and evolution of the Asian Aconitum subgenus Aconitum (Ranunculaceae). Journal of Plant Research. 1995; 108: 429–442. [Google Scholar]
- 17.Jabbour F, Renner SS. A phylogeny of Delphinieae (Ranunculaceae) shows that Aconitum is nested within Delphinium and that Late Miocene transitions to long life cycles in the Himalayas and Southwest China coincide with bursts in diversification. Molecular Phylogenetics and Evolution. 2012; 62: 928–942. 10.1016/j.ympev.2011.12.005 [DOI] [PubMed] [Google Scholar]
- 18.Doyle JJ, Doyle JL. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin. 1987; 19: 11–15. [Google Scholar]
- 19.White TJ, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR protocols: a guide to methods and applications. San Diego: Academic Press; 1990. pp. 315–322. [Google Scholar]
- 20.Kim Y, Park C, Kim K. Complete chloroplast DNA sequence from a Korean endemic genus, Megaleranthis saniculifolia, and its evolutionary implications. Molecules and Cells. 2009; 27: 365–381. 10.1007/s10059-009-0047-6 [DOI] [PubMed] [Google Scholar]
- 21.Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. The Clustal_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research. 1997; 25: 4876–4882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007; 23: 2947–2948. 10.1093/bioinformatics/btm404 [DOI] [PubMed] [Google Scholar]
- 23.Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series. 1999; 41: 95–98. [Google Scholar]
- 24.Swofford DL. PAUP*. Phylogenetic analysis using parsimony (*and other methods), version 4.0b10. Sunderland, Massachusetts: Sinauer; 2003. [Google Scholar]
- 25.Zwickl DJ. Genetic algorithm approaches for the phylogenetic analysis of large biological sequence datasets under the maximum likelihood criterion. Ph.D. dissertation, The University of Texas, Austin, USA; 2006.
- 26.Ronquist F, Teslenko M, van der Mark P, Ayres DL, Darling A, Höhna S, et al. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology. 2012; 61: 539–542. 10.1093/sysbio/sys029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Posada D, Crandall KA. Modeltest: testing the model of DNA substitution. Bioinformatics. 1998; 14: 817–818. [DOI] [PubMed] [Google Scholar]
- 28.Drummond AJ, Rambaut A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evolutionary Biology. 2007; 7: 214 10.1186/1471-2148-7-214 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Farris JS, Källersjö M, Kluge AG, Bult C. Testing significance of incongruence. Cladistics. 1995; 10: 315–319. [Google Scholar]
- 30.Templeton AR. Phylogenetic inference from restriction endonuclease cleavage site maps with particular reference to the evolution of humans and the apes. Evolution. 1983; 37: 221–244. [DOI] [PubMed] [Google Scholar]
- 31.Mason-Gamer RJ, Kellogg EA. Testing for phylogenetic conflict among molecular data sets in the tribe Triticeae (Gramineae). Systematic Biology. 1996; 45: 524–545. [Google Scholar]
- 32.Shimodaira H. An approximately unbiased test of phylogenetic tree selection. Systematic Biology. 2002; 51: 492–508. 10.1080/10635150290069913 [DOI] [PubMed] [Google Scholar]
- 33.Shimodaira H, Hasegawa M. CONSEL: for assessing the confidence of phylogenetic tree selection. Bioinformatics. 2001; 17: 1246–1247. [DOI] [PubMed] [Google Scholar]
- 34.Tamura M. Morphology, ecology and phylogeny of the Ranunculaceae VI Science Reports, College of General Education, Osaka University; 1966; 15: 13–35. [Google Scholar]
- 35.Kadota Y. Systematic studies of Asian Aconitum (Ranunculaceae) XIII. Species of Bhutan including a new scandent species. The Journal of Japanese Botany. 2010; 85: 8–24. [Google Scholar]
- 36.Shankar U. Aconitum fletcheranum G. Taylor (Ranunculaceae) in eastern Himalayas: Occurrence and conservation. Current Science. 2003; 84: 148–151. [Google Scholar]
- 37.Langlet O. Beiträge zur Zytologie der Ranunculazeen. Svensk Botanisk Tidskrift. 1927; 21: 1–17. [Google Scholar]
- 38.Langlet O. Über Chromosomenverhältnisse und Systematik der Ranunculaceae. Svensk Botanisk Tidskrift. 1932; 26: 381–400. [Google Scholar]
- 39.Lewitsky GA. The ‘karyotype’ in systematics, on the base of karyology of the subfamily Helleboreae. Bulletin of Applied Botany, of Genetics and Plant Breeding.1931; 27: 187–240. [Google Scholar]
- 40.Schafer B, La Cour L. A chromosome survey of Aconitum L. Annals of Botany. 1934; 48: 693–713. [Google Scholar]
- 41.Gregory WC. Phylogenetic and cytological studies in the Ranunculaceae Juss. Transactions of the American Philosophical Society. 1941; 31: 443–521. [Google Scholar]
- 42.Kurita M. Chromosome studies in Ranunculaceae V. Karyotypes of the subtribe Delphininae. Reports of the Biological Institute, Ehime University. 1957; 3: 1–8.
- 43.Shang XM, Lee CL. Chromosome studies of ten species of Aconitum in China. Acta Phytotaxonomica Sinica. 1984; 22: 378–385. [Google Scholar]
- 44.Yang QE. Cytology of 12 species in Aconitum L. and of 18 species in Delphinium L. of the tribe Delphineae (Ranunculaceae) from China. Acta Phytotaxonomica Sinica. 2001; 39: 502–514. [Google Scholar]
- 45.Yang QE, Gu ZJ, Wu ZY, Hong DY. A karyomorphological study in Aconitum subgen. Aconitum (Ranunculaceae) from Yunnan, China. Cathaya. 1993; 5: 89–114. [Google Scholar]
- 46.Swofford DL, Olsen GJ, Waddell PJ, Hillis DM. Phylogenetic inference In: Hillis DM, Moritz C, Mable BK, editors. Molecular Systematics, 2nd edition Sinauer Associates: Sunderland, Massachusetts; 1996. pp. 407–514. [Google Scholar]
- 47.Philippe H, Telford MJ. Large-scale sequencing and the new animal phylogeny. Trends in Ecology and Evolution. 2006; 21: 614–620. 10.1016/j.tree.2006.08.004 [DOI] [PubMed] [Google Scholar]
- 48.Wendel JF, Doyle JJ. Phylogenetic incongruence: window into genome history and molecular evolution In: Soltis DE, Soltis PS, Doyle JJ, editors. Molecular Systematics of Plants II. Boston: Kluwer Academic Publishers; and London: Dordrecht; 1998. pp. 265–296. [Google Scholar]
- 49.Than C, Ruths D, Innan H, Nakhleh L. Confounding factors in HGT detection: Statistical error, coalescent effects, and multiple solutions. Journal of Computational Biology. 2007; 14: 517–535. 10.1089/cmb.2007.A010 [DOI] [PubMed] [Google Scholar]
- 50.Kubatko LS. Identifying hybridization events in the presence of coalescence via model selection. Systematic Biology. 2009; 58: 478–488. 10.1093/sysbio/syp055 [DOI] [PubMed] [Google Scholar]
- 51.Joly S, McLenachan PA, Lockhart PJ. A statistical approach for distinguishing hybridization and incomplete lineage sorting. The American Naturalist. 2009; 174: E54–E70. 10.1086/600082 [DOI] [PubMed] [Google Scholar]
- 52.Mallet J, Besansky N, Hahn MW. How reticulated are species? Bioessays. 2016; 38: 140–149. 10.1002/bies.201500149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chaudhary LB, Rao RR. Notes on the genus Aconitum L. (Ranunculaceae) in North-West Himalaya (India). Feddes Repertorium. 1998; 109: 527–537. [Google Scholar]
- 54.Jee V, Dhar U, Kachroo P. Aconitum moschatum Stapf In: Löve À, editor. IOPB Chromosome Number Reports LXXIX. Taxon; 1983; 32: 320–324. [Google Scholar]
- 55.Seelanan TA, Schnabel A, Wendel JF. Congruence and consensus in the cotton tribe. Systematic Botany. 1997; 22: 259–290. [Google Scholar]
- 56.Kong HH, Gao Q, Luo Y, Yang QE. Seed morphology in some Chinese species of Aconitum (Ranunculaceae) and its systematic implications. Plant Diversity and Resources. 2013; 35: 241–252. [Google Scholar]
- 57.Wang WT, Hsiao PK. Notulae de Ranunculaceis sinensibus (II). Acta Phytotaxonomica Sinica 12, Additamentum. 1965; 1: 49–103. [Google Scholar]
- 58.Kadota Y. Systematic studies of Asian Aconitum (Ranunculaceae) IV. Description of three new species and one new variety. The Journal of Japanese Botany. 1999; 74: 282–295. [Google Scholar]
- 59.Krasnikov AA, Schaulo DN. Chromosome numbers of some representatives of the families Apiaceae and Ranunculaceae from the south of Siberia. Botanicheskii Zhurnal (Moscow & Leningrad). 1986; 71: 116–117. [Google Scholar]
- 60.Borodina-Grabovskaya AE, Grubov VI, Mikhailova MA. Aconitum L. In: Borodina-Grabovskaya AE, Grubov VI, Mikhailova MA, editors. Plants of Central Asia, Vol. 12 New Hampshire: Science Publishers; 2007. pp. 22–36. [Google Scholar]
- 61.Wang WT. Ranunculaceae In: Chen PC, Wu PC, Chiu PH, Hsu PS, Chow CL, editors. Observationes ad Florulam Hwangshanicum. Shanghai: Shanghai Science and Technology Press; 1965. pp. 113–116. [Google Scholar]
- 62.Li LQ, Kadota Y. Aconitum L. In: Wu ZY, Raven PH, editors. Flora of China, Vol. 6 Beijing: Science Press and St; Louis: Missouri Botanical Garden Press; 2001. pp. 149–222. [Google Scholar]
- 63.Gao Q, Ren C, Yang QE. Taxonomic status and distributional range of Aconitum angustius (Ranunculaceae) based on cytological evidence. Nordic Journal of Botany. 2012; 30: 1–13. [Google Scholar]
- 64.Handel-Mazzetti H. Plantae sinensis XXXIII. Ranunculaceae. Acta Horti Gotoburgensis. 1939; 13: 39–219. [Google Scholar]
- 65.Steinberg EI. Aconitum L. In: Komarov VL, Shishkin BK, editors. Flora of the URSS. Vol. VII Moscow and Leningrad; 1937. pp. 183–236. [Google Scholar]
- 66.Tsitrone A, Kirkpatrick M, Levin DA. A model for chloroplast capture. Evolution. 2003; 57: 1776–1782. [DOI] [PubMed] [Google Scholar]
- 67.Fehrer J, Gemeinholzer B, Chrtek J, Bräutigam S. Incongruent plastid and nuclear DNA phylogenies reveal ancient intergeneric hybridization in Pilosella hawkweeds (Hieracium, Cichorieae, Asteraceae). Molecular Phylogenetics and Evolution. 2007; 42: 347–361. 10.1016/j.ympev.2006.07.004 [DOI] [PubMed] [Google Scholar]
- 68.Acosta MC, Premoli AC. Evidence of chloroplast capture in South American Nothofagus (subgenus Nothofagus, Nothofagaceae). Molecular Phylogenetics and Evolution. 2010; 54: 235–242. 10.1016/j.ympev.2009.08.008 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses. “-” indicates that support is less than 50% bootstrap value.
(JPG)
Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses.
(JPG)
Numbers above branches are posterior probabilities; numbers below branches are bootstrap values for maximum parsimony/maximum likelihood analyses. “-” indicates that support is less than 50% bootstrap value.
(JPG)
(XLSX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.