

Abstract
The data herein is in support of our research article by McDougall et al. (2017) [1], in which we used our zebrafish model of embryonic vitamin E (VitE) deficiency to study the consequences of VitE deficiency during development. Adult 5D wild-type zebrafish (Danio rerio), fed defined diets without (E–) or with VitE (E+, 500 mg RRR-α-tocopheryl acetate/kg diet), were spawned to obtain E– and E+ embryos that we evaluated using metabolomics and specific lipid analyses (each measure at 24, 48, 72, 120 hours-post-fertilization, hpf), neurobehavioral development (locomotor responses at 96 hpf), and rescue strategies. Rescues were attempted using micro-injection into the yolksac using VitE (as a phospholipid emulsion containing d6-α-tocopherol at 0 hpf) or D-glucose (in saline at 24 hpf).
Keywords: Alpha-tocopherol, Docosahexaenoic acid, Choline, Glucose, Ferroptosis
Specifications Table
| Subject area | Biology |
| More specific subject area | Nutrition; Antioxidants |
| Type of data | Graphs; Figures |
| How data was acquired | LC-MS (liquid chromatography-mass spectrometry) using a Shimadzu Nexera system (Shimadzu; Columbia, MD, USA) coupled to a high-resolution hybrid quadrupole–time–of-flight mass spectrometer (TripleTOF® 5600; SCIEX; Framingham, MA, USA); Embryos were assessed for viability, developmental progression and spontaneous movements (earliest behavior in zebrafish), using the zebrafish acquisition and analysis program (ZAAP). Locomotor Response Assay using a Viewpoint ZebraBox (software version 3.0, Viewpoint Life Sciences, Lyon, France). |
| Data format | Analyzed |
| Experimental factors | Adult 5D zebrafish fed defined diets without (E–) or with VitE (E+, 500 mg RRR-α-tocopheryl acetate/kg diet) were spawned to obtain E– and E+ embryos that were evaluated up to 120 hours-post-fertilization (hpf) |
| Experimental features | Lipid and metabolomics analyses were performed in developing E– and E+ zebrafish embryos collected daily from 24 to 120 hpf. Mortality and neurobehavioral outcomes were assessed at 96 hpf in two separate rescue experiments using E– and E+ embryo populations micro-injected into the yolk (1) at 0 hpf with VitE or (2) at 24 hpf with D-glucose |
| Data source location | Oregon State University, Corvallis, OR 97330 |
| Data accessibility | Data is within this article |
Value of the data
-
•
Fatty acid quantification and peroxidation data during zebrafish embryonic development in E- vs. E+ zebrafish embryos may be used by other researchers to investigate antioxidant effects of VitE with respect to specific lipids.
-
•
The metabolomics dataset may be utilized by other researchers to investigate the secondary metabolic effects of VitE deficiency.
-
•
Rescue studies using microinjection into the yolksac may be compared to other methods of compound/nutrient delivery to developing zebrafish.
1. Data
Fig. 1. shows data from quantitative analyses of LA (linoleic acid, 18:2, omega-6); ARA (arachidonic acid, 20:4, omega-6); EPA (eicosapentaenoic acid, 20:5, omega-3); DHA (docosahexaenoic acid, 22:6, omega-3) in fatty acid extracts from samples with and without alcoholic saponification of E– and E+ embryos collected at 24, 48, 72, and 120 hpf. Tables 1and 2 provide detailed targeted metabolomics datasets for E– and E+ embryos collected at 24, 48, 72, and 120 hpf. Relative response intensity metabolomics data for choline and methylation pathway intermediates in E– and E+ embryos are shown in Fig. 2. Relative response intensities of antioxidant network components from metabolomic analyses, as well as quantification of α-tocopherol and ascorbic acid, in E– and E+ embryos (pmol/embryo) are shown in Fig. 3. Relative response intensities of glycolytic and tricarboxylic acid cycle intermediates in E– and E+ embryos are shown in Fig. 4. Relative response intensities of free saturated fatty acids and coenzyme A from metabolomics data in E– and E+ embryo are shown in Fig. 5. Fig. 6 shows locomotor activity data from E– and E+ embryos micro-injected into the yolksac at 0 hpf with either saline or a VitE–emulsion. Fig. 7 shows locomotor activity data from E– and E+ embryos micro-injected into the yolksac at 24 hpf with either saline or D-glucose.
Fig. 1.

Quantified levels of total and free (unesterified) fatty acids in E– vs. E+ embryos. Area counts normalized using internal standards (n=3 samples/group, with n=10–15 embryos/sample for total lipids; n=4 samples/group with n=15-30 embryo/sample for free fatty acids). Shown are saponified (upper row) or extracted only (lower row) samples, means±SEM; p-values are for VitE x Age interactions, unless main effects (VitE or Age) are indicated (Tukey’s post-test, p<0.05 for bars bearing different letters). Abbreviations: LA (linoleic acid); ARA (arachidonic acid); EPA (eicosapentaenoic acid); DHA (docosahexaenoic acid).
Fig. 2.

Relative response intensities of choline and methylation pathway intermediates. E– and E+ embryo (n=15/sample; 4 samples/group) data were normalized against QC sample intensities (n=4) for each individual metabolite. Statistical significance (p<0.05) was calculated using 2-way ANOVA with Sidak’s post-test for multiple comparisons of normalized and natural log-transformed intensity values. Shown are means ± SEM; p-Values are for VitE x Age interactions, unless indicated otherwise. Paired comparison, p-values are indicated as *<0.05, **<0.005, ***<0.001, **** <0.0001.
Fig. 3.

Relative response intensities of antioxidant network components from metabolomics and quantification of α-tocopherol and ascorbic acid. A. E– and E+ embryo (n=15/sample; 4 samples/group) relative response data was normalized against QC sample intensities (n=4) for each individual metabolite. B. Quantified levels of α-tocopherol and ascorbic acid, according to established protocols (31) and (33), respectively. Statistical significance (p<0.05) was calculated using 2-way ANOVA with Sidak’s post-test for multiple comparisons of normalized and natural log-transformed intensity values. Shown are means ± SEM; p-Values are for VitE x Age interactions, unless indicated otherwise. Paired comparisons p-values are indicated as *<0.05, **<0.005, ***<0.001, **** <0.0001. C. Antioxidant network scheme showing interaction of antioxidants with lipid radicals and consumption or NADPH.
Fig. 4.

Relative response intensities of glycolytic and tricarboxylic acid cycle intermediates. E– and E+ embryo (n=15/sample; 4 samples/group) data were normalized against QC sample intensities (n=4) for each individual metabolite. Statistical significance (p<0.05) was calculated using 2-way ANOVA with Sidak’s post-test for multiple comparisons of normalized and natural log-transformed intensity values. Shown are means ± SEM; p-values are for VitE x Age interactions. Paired comparisons p-values are indicated as *<0.05, **<0.005, ***<0.001, **** <0.0001.
Fig. 5.
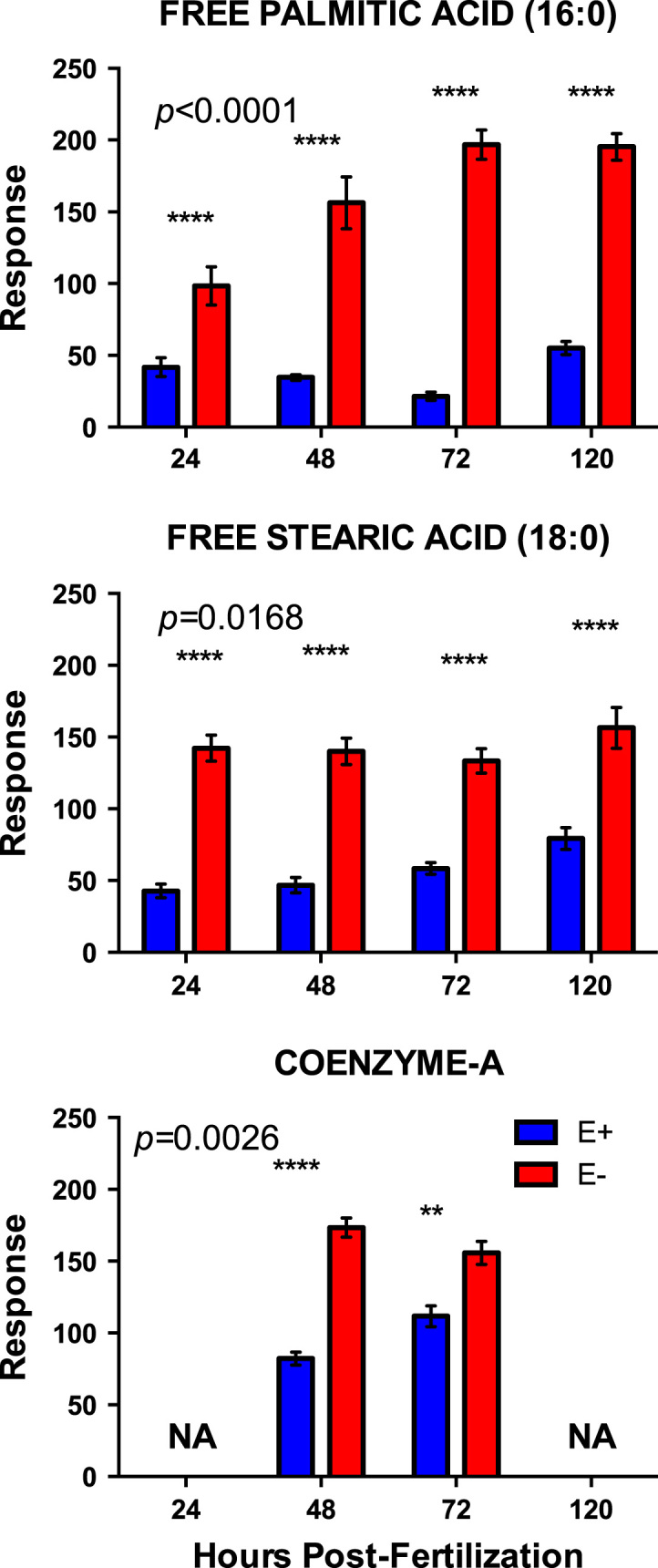
Relative response intensities of free saturated fatty acids and coenzyme A from metabolomic analyses. E– and E+ embryo (n=15/sample; 4 samples/group) data were normalized against QC sample intensities (n=4) for each individual metabolite. Statistical significance (p<0.05) was calculated using 2-way ANOVA with Sidak’s post-test for multiple comparisons of normalized and natural log-transformed intensity values. Shown are means±SEM; p-values are for VitE x Age interactions. Paired comparison p-values are indicated as *<0.05, **<0.005, ***<0.001, **** <0.0001.
Fig. 6.

E– compared with E+ embryos have impaired behavior when injected with saline (upper panel), but restored responses when injected with VitE (lower panel). A. Embryos were analyzed in 96-well plates (128 embryos per group). Locomotor activities following a series of light stimuli (a stimulus every 6 for 24 min) were measured as distance moved (mm) over time (seconds). At 96 hpf, E- (red) embryos treated with saline (upper panel) were 47% less responsive to light than were E+ embryos (E– area-under-curve, AUC: 2040±178; E+ AUC: 3877±228; p<0.0001). Embryos with morphological defects were not included in data analysis. E- behavior was restored using VitE injection into the yolk at the 1 cell stage (lower panel E– AUC: 2970±280; E+ AUC: 3340±226, not significantly different). B. Bar chart comparisons of respective time-course data. VitE (tocopherol)-injected E– and E+ embryo locomotor activities were not significantly different.
Fig. 7.

Locomotor response assay activity data showing neurobehavioral impairment. E– and E+ embryos (96 hpf) were analyzed in 96-well plates (128 embryos per group). Locomotor activities following a series of light stimuli (every 6 for 24 min) were measured as distance moved (mm) over time (seconds). E– (red) embryos treated with saline (upper panel) were 84% less responsive to light than were E+ (blue) embryos (E– area-under-curve, AUC: 572±72 E+ AUC: 3580±387; p<0.0001). Embryos with morphological defects were not included in data analysis. E– behavior was partially restored by approximately 50% following glucose injection into the yolk at 24 hpf (lower panel; E– AUC: 2502±150; E+ AUC: 3734±359; p<0.0001). Statistical significance was determined using a Kolmogorov–Smirnov test (p<0.01).
2. Experimental design, materials and methods
2.1. Study design
All experiments (i.e. lipid quantifications, targeted metabolomics analyses, and micro-injection rescue studies) were performed in duplicate and have been reported in detail [1].
2.2. Zebrafish husbandry and diets
The Institutional Animal Care and Use Committee of Oregon State University approved this protocol (ACUP Number: 4344). Tropical 5D strain zebrafish were housed in the Sinnhuber Aquatic Research Laboratory and complete details of the housing and husbandry have been reported [1].
2.3. Analyses
Diet and embryo α-tocopherol [2] and ascorbic acid [3] were determined using high-pressure liquid chromatography with electrochemical detection as reported [1].
Extraction and sample preparation for metabolomic analysis were performed following 24, 48, 72, and 120 hpf, embryos (n=15 per replicate, n=4 replicates per group), as described [1]. Chromatography was performed with a Shimadzu Nexera system (Shimadzu; Columbia, MD, USA) coupled to a high-resolution hybrid quadrupole–time–of-flight mass spectrometer (TripleTOF® 5600; SCIEX; Framingham, MA, USA). Two different LC analyses using reverse phase and HILIC columns were used, as described [1].
Analysis of total DHA, EPA, ARA, and LA were performed as described [2] with modifications, as described [1]. Chromatographic separations were carried out on 4.6×250 mm J׳sphere ODS-H80 (4 µm, YMC Co, Kyoto, Japan) for negative ion analysis. TOF-MS and TOF-MS/MS were operated with same parameters as for metabolomics, as described [1].
2.4. Microinjection rescue studies
Embryos were microinjected as described and criteria used to assess supplementation tolerance of zebrafish embryos using ZAAP at 24, 48, and 120 hpf, as described [1].
2.5. Behavioral assessments
Locomotor activity was measured in a total of n=128 embryos per group using Viewpoint Zebrabox [4], [5], as described [1].
2.6. Data processing and statistical analyses
All data processing and statistical analyses were performed as described in [4], [5], [6], with modification made as reported [1].
Acknowledgements
The authors thank Carrie Barton, Greg Gonnerman, Andrea Knecht, Jane La Du, Scott Leonard, and Lisa Truong for providing outstanding technical assistance. National Institutes of Health Grants S10RR027878 (MGT and JFS) and NIEHS P30 ES000210 (RT) supported this work. MM was supported in part by National Science Foundation Grant DGE 0965820. H-KK sabbatical support provided by The Catholic University of Korea. MGT supported in part by the Helen P Rumbel endowment to the Linus Pauling Institute.
Footnotes
Transparency data associated with this article can be found in the online version at doi:10.1016/j.dib.2017.02.046.
Supplementary data associated with this article can be found in the online version at doi:10.1016/j.dib.2017.02.046.
Transparency document. Supplementary material
Supplementary material
Appendix A. Supplementary material
Supplementary material
References
- 1.McDougall M., Choi J., Kim H.K., Bobe G., Stevens J.F., Cadenas E., Tanguay R., Traber M.G. Lethal dysregulation of energy metabolism during embryonic vitamin E deficiency. Free Radic. Biol. Med. 2017;104:324–332. doi: 10.1016/j.freeradbiomed.2017.01.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lebold K.M., Kirkwood J.S., Taylor A.W., Choi J., Barton C.L., Miller G.W., La Du J., Jump D.B., Stevens J.F., Tanguay R.L., Traber M.G. Novel liquid chromatography-mass spectrometry method shows that vitamin E deficiency depletes arachidonic and docosahexaenoic acids in zebrafish (Danio rerio) embryos. Redox Biol. 2013;2:105–113. doi: 10.1016/j.redox.2013.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Frei B., England L., Ames B.N. Ascorbate is an outstanding antioxidant in human blood plasma. Proc. Natl. Acad. Sci USA. 1989;86:6377–6381. doi: 10.1073/pnas.86.16.6377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Saili K.S., Corvi M.M., Weber D.N., Patel A.U., Das S.R., Przybyla J., Anderson K.A., Tanguay R.L. Neurodevelopmental low-dose bisphenol A exposure leads to early life-stage hyperactivity and learning deficits in adult zebrafish. Toxicology. 2012;291:83–92. doi: 10.1016/j.tox.2011.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Noyes P.D., Haggard D.E., Gonnerman G.D., Tanguay R.L. Advanced morphological – behavioral test platform reveals neurodevelopmental defects in embryonic zebrafish exposed to comprehensive suite of halogenated and organophosphate flame retardants. Toxicol. Sci. 2015;145:177–195. doi: 10.1093/toxsci/kfv044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Benjamini Y., Krieger A.M., Yekutieli D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika. 2006;93:491–507. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material
Supplementary material