

Abstract
Due to the rise in multi-drug resistant pathogens and other diseases, there is renewed interest in marine sponge endosymbionts as a rich source of natural products (NPs). The South African marine environment is rich in marine biota that remains largely unexplored and may represent an important source for the discovery of novel NPs. We first investigated the bacterial diversity associated with five South African marine sponges, whose microbial populations had not previously been investigated, and select the two sponges (Isodictya compressa and Higginsia bidentifera) with highest species richness to culture bacteria. By employing 33 different growth conditions 415 sponge-associated bacterial isolates were cultured and screened for antibacterial activity. Thirty-five isolates showed antibacterial activity, twelve of which exhibited activity against the multi-drug resistant Escherichia coli 1699, implying that some of the bioactive compounds could be novel. Genome sequencing of two of these isolates confirmed that they harbour uncharacterized biosynthetic pathways that may encode novel chemical structures.
Keywords: Isodictya compressa, Higginsia bidentifera, natural products, marine sponge, antimicrobial
1. Introduction
The marine environment has been identified as a rich source of bioactive compounds with interesting chemical diversity, and in particular those produced by marine sponges [1]. Sponges belong to the phylum Porifera, and are some of the oldest metazoans on Earth [2,3]. There are four classes of marine sponges, namely Calcarea, Hexactinellida, Homoscleromorpha and Demospongiae with the Demospongiae being most abundant, representing 83% of described species [4]. These filter feeding sessile organisms occur primarily in marine environments where they survive under immense competition and predation [5,6,7].
There is evidence that many of the previously identified marine sponge secondary metabolites are of microbial origin [1,8,9,10]. Their ability to survive in this competitive environment could be due to their own adaptation, in addition to or perhaps because of the microbial communities they harbor, which can account for up to 40% of the sponge’s wet weight [5,7,11]. Furthermore, these microbes are proven to produce medically relevant compounds including ecteinascidin-743 (Yondelis®), dolastatin-10 (Adcetris®) and halichondrin B (Halaven®) [12]. As a result, marine sponge-associated microbes are sought after for natural product discovery [13,14,15,16] due to their potential for delivering novel bioactive compounds [17,18]. In general, the large scale production of microbial-derived secondary metabolites for study and use is hampered by the lack of these bacteria in culture collections, either through an inability to culture them or identify the microbes responsible for producing an observed bioactivity [19]. It is therefore important to develop techniques to cultivate these marine sponge symbionts in order to access and exploit their secondary metabolite capabilities [20]. The use of novel culture media, such as very minimal nutrient content [21], application of diffusion chambers [20,22] or microencapsulation [23], have successfully resulted in the isolation of previously uncultured marine bacteria.
South Africa has a unique coastline that is dominated by two major current systems: the cold Benguela current and the warm Agulhas current [24,25]. As a result of the physico-chemical differences introduced by the two currents, the coastline has an exceptionally rich and varied marine life which remains largely unexplored. The few reported studies performed on South African marine sponges thus far do not take their associated microorganisms into account [26]. The latrunculid sponge, Tsitsikamma favus, endemic to the coast of South Africa, represents one of the few specimens whose microbial diversity has been assessed and serves as an example of what South African sponges may have to offer [8]. Unique to this sponge is the production of pyrroloiminoquinones, known as tsitsikammamines, that mediate cytotoxicity through the cleavage of DNA due to inhibition of topoisomerase I [27] and it is thought that these secondary metabolites are of microbial origin [9]. The South African coast therefore represents a potentially resourceful environment for the exploration of novel bioactive compounds.
This study assesses the bacterial diversity associated with five South African marine sponges, namely Waltherarndtia caliculatum, Higginsia bidentifera, Isodictya compressa, Spongia and Axinella species. All five sponges belong to the class Demospongiae. The Higginsia genus was first described by Higgin in 1877 during a scientific expedition aboard the ship Argo, with the species Higginsia bidentifera described in 1886 and according to the World Porifera Database, appears to be endemic to South African coastal waters [28,29]. Isodictya compressa was first described by Esper in 1794, while Waltherarndtia caliculatum was described by Kirkpatrick in 1903, both of which are endemic to South Africa [30,31,32,33] and the bacteria associated with these three marine sponges have not been investigated. Axinella and Spongia species have a global distribution and the microbial populations of some species of these sponge genera have been studied [34,35,36,37]. We first determined the bacterial diversity associated with these five sponges, then isolated bacteria from two of these and demonstrated their ability to produce antibacterial compounds. Here we report a new collection of South African marine sponge-associated bacteria that show antibacterial activity when subjected to a variety of culture conditions.
2. Materials and Methods
Unless otherwise specified, chemicals and reagents used in this study were supplied by Sigma Aldrich Chemical Company (Darmstadt, Germany), Merck Chemical and Laboratory Supplies (Darmstadt, Germany) and Kimix Chemical and Laboratory Supplies (Cape Town, South Africa). The enzymes, DNA size markers and polymerases were purchased from Fermentas Life Sciences Ltd. (Vilnius, Lithuania).
2.1. Sample Collection
The five marine sponges (Waltherarndtia caliculatum, PE05; Higginsia bidentifera, PE07; Isodictya compressa, PE09; an Axinella species, PE11; and a Spongia species PE08) were collected in January and April 2013 in Algoa Bay, Port Elizabeth, South Africa (longitude 34°00.366 S and latitude 25°43.209 E; 15 °C), and kept on ice in sterile sea water. The sponge samples (Figure S1) were shipped to the Institute for Microbial Biotechnology and Metagenomics, Cape Town, where the sponge material was stored at –20 °C and –80 °C in 20% glycerol until further processing. Voucher specimens of all sponge samples are housed in the South African Institute for Biodiversity (SAIAB) in Grahamstown.
2.2. Terminal Restriction Fragment Length Polymorphism (T-RFLP) Analysis
The five sponge samples were thawed on an ice/water slurry and cut with a sterile blade into 1 g aliquots for further processing. The UltraClean Soil DNA Isolation kit (MOBIO Laboratories, Carlsbad, CA, USA) was used to isolate DNA according to the manufactures protocol. Metagenomic DNA (mDNA) was further purified as described by Liles et al. [38]. The NucleoSpin® Gel and PCR Clean-up kit (Macherey-Nagel, Düren, Germany) was used to extract the mDNA from the agarose plug.
The PCR reaction mixture to generate amplicons for T-RFLP was as follows: 1 × Phusion DNA polymerase buffer, 0.2 mM dNTP mix, 0.5 µM of each forward primer E9F (5′-GAGTTTGATCCTGGCTCAG-3′) labelled at the 5′ end with fluorescein amidite (FAM) and reverse primer U1510R (5′-GGTTACCTTGTTGTTACACTT-3′), 0.02 U/µL of Phusion DNA polymerase (Fermentas, Lithuania), 100 ng of template mDNA and ultraviolet (UV) treated water [39,40]. The following cycling conditions were used: initial denaturation of 98 °C for 2 min followed by 35 cycles of 98 °C for 20 s, 54 °C for 30 s, 72 °C for 1 min and final elongation at 72 °C for 5 min. Duplicate amplifications were conducted for each sample, pooled and purified using the NucleoSpin® Gel and PCR Clean-up kit (Macherey-Nagel). Amplicons (1 µg) were digested in 30 µL volumes containing 3 µL of 10 × Tango buffer, 2 µL of HhaI (10 U/µL), 4 µL HaeIII (10 U/µL), and UV treated water. Digestions were performed in a Bio-Rad T100 Thermal Cycler (Bio-Rad, Hercules, CA, USA) overnight at 37 °C, followed by inactivation at 80 °C for 20 min. Capillary electrophoresis and analysis of the fluorescently labelled terminal restriction fragments (T-RFs) was carried out on an automated ABI3130XL genetic analyzer (Applied Biosystems, Foster City, CA, USA) by the Central Analytical Facility at the University of Stellenbosch, South Africa. The marker used for the analysis was GS500LIZ. T-RFLP profiles were analysed using freeware Peak ScannerTM Software Version 1.0 (Applied Biosystems). Peak height was used to characterize unique T-RFs. Peaks shorter than 25 bp and larger than 1200 bp were excluded from further analysis. Valid peaks were identified and aligned using the online T-REX software [41] to create an operational taxonomic unit (OTU) data matrix. In theory, each OTU represents an individual T-RF where one T-RF represents one distinct ribotype [42]. It should be noted however that, one peak may represent two different bacterial species or two peaks may be one bacterial species depending on the recognition site of restriction enzymes on the 16S ribosomal RNA (rRNA) gene sequence [43,44,45]. The OTU matrix was analysed using Primer 6, Version 6.1.11 (Primer E, Plymouth, UK) software and diversity indices for all the samples were determined.
2.3. Bacterial Culturing
Thawed sponge material (1 g) was homogenised by grinding with a sterile pestle and mortar in 9 mL of sterile sea water. The sponge homogenates were serially diluted in sterile sea water to 10−6, and from each dilution, 100 μL aliquots were plated on 23 general media (Table S1). The full culturing scheme is depicted in Figure S2. In addition, aliquots of the sponge homogenates underwent heat treatment to select for endospore forming bacteria [15] and to inhibit the growth of Gram negative fast growing bacteria [46]. The heat treatment process was adapted from Phelan et al. (2012) [15] with minor modifications. An aliquot of 100 μL of the 10−1 dilution of the sponge homogenate was incubated at 80 °C for 10 min and serially diluted to 10−6. Aliquots of 100 μL were aseptically plated on three media Shivji Nutrient Agar (SNA), Tryptic Soy Agar (TSA) and Zobell Agar (ZBA). Liquid enrichment was used to select for cyanobacteria [47] and nitrogen fixing bacteria [48]. For liquid enrichment cultures, 100 μL of the 10−1 dilutions of the sponge homogenate were added to 9 mL of Ashby’s Nitrogen Free Media (ANFA), MagMin Semi-Solid Medium (MMM) and Blue Green Algae (BG11) liquid media. The liquid enrichment cultures were incubated at 15 °C for up to six weeks on a shaker at 150 rpm. Subsequently, 100 μL of the liquid enriched cultures were plated on ZBA plates, as ZBA is one of the rich media which can generally support proliferation of a range of different bacteria. After the initial isolations, three media Glucose Yeast Media (GYM), TSA and ZBA were selected as the “standard/rich media” for routine culturing and characterization of the bacterial isolates. Isolates from Trypticase Soya Agar pH 4.5 (TS4), TSA and SNA, inclusive of those which underwent different treatments, were cultured on TSA. Isolates from Medium A Agar (MAA), GYM, Modified 172F medium (172), Oatmeal Agar (OMA), Actinomyces Isolation Medium (AIM), and Planctomycetes Medium (PMM) along with their different treatments were cultured on GYM. Isolates from the thirteen other media with their respective treatments were also cultured on ZBA. Antibiotic treated sponge homogenates were prepared by adding 20 μg/mL of streptomycin [49] to the homogenate. The homogenates were agitated on a shaker (150 rpm) for 1 h at room temperature. The sponge homogenates were centrifuged at 11,000× g for 3 min and the supernatant was discarded. Sterile sea water (1 mL) was used to re-suspend the pellet and the suspension was serially diluted to 10−6. Aliquots of 100 μL of the 10−1 to 10−6 dilutions were plated on eight media (OMA, OMA + NaCl (1.8% wt/vol), AIM, GYM, 172, MAA, Yeast Extract-Malt Extract (YEME) and Starch Casein Nitrate Agar (SCN)) containing cyclohexamide (100 μg/mL) and nalidixic acid (50 μg/mL) [50]. All plates were incubated at 15 °C for up to four weeks. Colonies were examined and picked on a daily basis selecting for colonies with different morphologies (shape, colour, texture, and size).
2.4. Antimicrobial Screening
The marine sponge-associated isolates were screened, using agar overlay assays, for antibacterial activity against the Gram-positive strains Mycobacterium smegmatis LR222, Bacillus cereus ATCC10702, Staphylococcus epidermidis ATCC14990 and the Gram negative strains Pseudomonas putida ATCC27853 and the multidrug resistant Escherichia coli 1699 (Cubist, Lexington, MA, USA). Isolates were cultivated on four media (ZBA, GYM, TSA and Activated Charcoal Medium (ACM)) for two weeks at room temperature. On the day prior to carrying out the overlay assays, the test strains were cultured overnight in 10 mL Luria Broth (LB) at 37 °C with aeration at 250 rpm. To ensure that approximately the same concentration of cells (using optical density measurements, OD600) were present in the sloppy agar, the following formulae were used to calculate the volume of culture to add to 6 mL of sloppy agar (LB): OD600 × X μL = 4 (for E. coli). OD600 × X μL =160 (for other test strains). The sloppy agar containing the test strains was poured over the marine isolate growth, incubated at 30 °C and examined for zones of inhibition after 16 h and again after 48 h.
For induction of antimicrobial activities, a one-strain-many-active-compounds (OSMAC) approach was followed. A total of 29 isolates were selected; 12 that gave hits against the panel of test strains in the overlay assays and an additional 17 isolates that did not show antibacterial activity in the first round of screening (Table S2). A matrix incorporating the following growth modifications or treatments was developed: heat or cold shock treatment, co-culturing with sponge-associated bacteria, and varying carbon (mannitol, succinic acid and starch at 5 mM) and nitrogen (NH4Cl or NaNO3 at 0.02% w/v) sources or phosphate concentrations (KH2PO4 at 0.1 µM or 0.5 mM). The base medium employed was a minimal media containing per liter: 18 g NaCl, 2 g MgCl2, 0.525 g KCl, 0.075 g CaCl and 2.38 g HEPES buffer. The experiment was set up in a 96-well format on large petri dish plates (20 mm × 150 mm). The method of plating in 96-well format was similar to that described in [51] with some modification. Single colonies from the selected isolates were inoculated into 96-well plates containing 70 µL sterile water in each well, and shaken at 100 rpm on a shaker for 10 min at room temperature (RT). Cultures were then plated aseptically on the different matrix media using a 96-pin hedgehog and allowed to grow for 2 weeks at RT. The order of inoculation of isolates was interchanged between isolates that gave positive hits on standard medium in the first round of screening and those which did not. All isolates were inoculated in triplicate on each 96-well plate and therefore located at different positions resulting in different neighboring isolates which resulted in a co-culture setup (Figure S3). On the day prior to performing overlays, cold and heat shock treatments were carried out. The heat shock method used was adapted from [52] with minor changes. Heat shock plates were incubated for 1 h at 42 °C, whereas cold shock treatment was carried out at 4 °C for 1 h. The heat and cold shock treatments were repeated on the day of the antibiotic overlay assay. The day prior to carrying out the overlay assay, single colonies of each of the test strains was inoculated into Luria Broth (10 mL) and the cultures were incubated at 37 °C overnight while shaking at 250 rpm. The optical density (OD600) of the cultures were determined after which they were diluted to achieve the same overall concentration of cells. An overlay assay similar to that described above was carried out using 24 mL of agar for the large format plates (20 mm × 150 mm).
2.5. DNA Sequencing and Phylogenetic Analysis
For 16S rRNA identification of the bioactive isolates, genomic DNA extraction was performed using the GeneJET Genomic DNA Purification Kit (Thermo Scientific, Waltham, MA, USA). A single colony was inoculated in 10 mL TSA broth or ¼ strength ZBA broth and incubated on a shaker (150 rpm) at room temperature for 2 to 5 days depending on growth rate. DNA extraction was carried out according to the manufacturer’s instructions for Gram-negative bacteria. Minor modifications were made for Gram-positive bacteria where 200 µL of lysis buffer (25 mM Tris-HCl pH8, 50 mM glucose, 10 mM EDTA and 25 mg/mL lysozyme) was added to the pelleted bacterial cells and the solutions were incubated at 37 °C for 30 min. Thereafter the manufacturer’s instructions were followed. PCR amplification was performed in 50 µL volumes which consisted of 1 × Dream Taq buffer, 0.2 mM of dNTP mix, 1 µM of each of the reverse and forward primers, 100 ng template DNA, 1.25 U DreamTaq DNA polymerase (Fermentas), and sterile distilled water. The universal primers E9F and U1510R were used to amplify the 16S rRNA gene sequences of the marine sponge-associated bacteria. The following cycling conditions were used: initial denaturation 95 °C for 5 min, 35 cycles of 95 °C for 45 s, 55 °C for 45 s, 72 °C for one and a half min and final elongation of 72 °C for 10 min.
The DNA sequence determination of purified 16S rRNA gene products were carried out by the Central Analytical Facility at Stellenbosch University using an ABI PRISM 377 automated sequencer (Applied Biosystems). The data from the sequencing unit was processed using Chromas Pro 1.5a software (Technelysium, South Brisbane, QLD, Australia) for alignment and manual editing of sequences. The neighbor-joining phylogenetic tree was constructed using MEGA 7 software [53]. Bootstrap tests were performed 1000 times.
2.6. Genome Sequencing
Whole genome sequencing of PE09-72 and PE09-221 was performed at the sequencing facility within the Institute for Microbial Biotechnology and Metagenomics, University of the Western Cape, South Africa. Genomic DNA was prepared using standard phenol:chloroform extraction. Sequencing libraries were prepared using the Illumina Nextera XT library prep kit according to the manufacturer’s instructions and sequencing was performed using the Illumina MiSeq Reagent kit V3 (Illumina, San Diego, CA, Country) which included a 10% phiX V3 spike [54]. This resulted in 2 × 300 base pair (bp) paired end reads (forward and reverse). Analysis of the sequence data was carried out using CLC Genomics version 7.5.1 (CLC, Aarhus, Denmark). Raw reads were reference assembled to phiX V3 genome used in the sequencing reaction for quality control. Unmapped reads were mapped to the human genome to remove any contaminating sequences. A de novo assembly was carried out on the remaining unmapped reads using default parameters. Basic analysis of the assembled contigs were carried out using the RAST server [55] and BLAST analysis against the NCBI database [56]. The genomes have been deposited in the Genbank database under accession numbers MIJA00000000 and MIJB00000000 for PE09-72 and PE09-221 respectively.
3. Results and Discussion
3.1. Bacterial Diversity Analysis and Culturing of Marine Sponge-Associated Bacteria
Five sponges (PE05, PE07, PE08, PE09, PE11) were investigated for their bacterial diversity, using 16S rRNA based T-RFLP analysis to identify those with highest diversity and unique operational taxonomic units (OTUs) to improve the chance of isolating novel bacteria. T-RFLP profiles revealed that there was a total of 179 OTUs from the five marine sponges, 90 of which were unique (Figure 1). Studies have classified microbial OTUs as either species specific OTUs (defined as OTUs unique to a certain sponge species) or sponge specific OTUs (defined as OTUs found in sponges but not found in other environments) [57,58]. Notably, five of the OTUs were shared among all five sponges indicating that the representative OTUs could be sponge specific [59,60].
Figure 1.

Venn diagram of species specific and shared operational taxonomic units (OTUs) (at genus level) in the selected marine sponges.
The Simpson index of the bacterial species from PE05 and the PE11 was the least even, indicating the presence of more dominating OTUs in these samples, while the other sponges had a more even distribution of species abundance (Table 1). PE07 and PE09 yielded the most OTUs with 43 and 44 respectively and had the highest evenness. Although seemingly low, the OTUs from the present study were similar to the numbers of T-RFLP-based OTUs reported from other studies where 45 and 32 OTUs were identified in Hymeniacidon heliophila and 36 and 22 OTUs were identified in Haliclona tubifera depending on the restriction enzymes used [61]. PE09 had the highest number of species specific OTUs (14) with the second highest being PE05 with 11 OTUs. Although PE07 had the second highest number of OTUs only three were specific to this sample which indicated that fewer H. bidentifera-specific OTUs were isolated from the sample. Given the higher diversity and even representation of species present in both PE07 and PE09, as well as the uniqueness of PE09 profiles and the representative nature of PE07, these were selected for bacterial isolation studies.
Table 1.
Diversity indices of the sponge-associated microbial populations.
| Sample ID | Marine Sponge ID | Species Specific OTUs | S | 1 − λ |
|---|---|---|---|---|
| PE05 | W. caliculatum | 11 | 36 | 0.6923 |
| PE07 | H. bidentifera | 3 | 43 | 0.9388 |
| PE08 | Spongia (spongia) sp. 001RSASPN | 7 | 24 | 0.8514 |
| PE09 | I. compressa | 14 | 44 | 0.9306 |
| PE11 | Axiinella sp. 007RSASPN | 7 | 32 | 0.7706 |
Index: species richness (S) = OTUs; Simpson index (1 − λ) = evenness.
A total of 415 unique bacterial colonies, as judged by their morphology (size, shape, colour and texture), were isolated from the two sponge samples with PE09 yielding 273 isolates and 142 bacterial isolates from PE07, respectively. The fewest number of isolates were obtained from samples growing in liquid enrichment medium and from the heat treated medium, which yielded 5 and 12 isolates respectively. This could be due to the selectivity in the liquid enrichment medium which was meant to select for nitrogen fixing bacteria. In the case of heat treatment, the low viable microbial numbers post treatment indicate that the number of spore-forming bacteria are low in these marine sponges or that sporulation had not been induced in spore formers at the time of harvest [15]. The antibiotic treatment yielded 37 isolates while the non-treated medium resulted in 361 bacterial isolates. GYM was the medium which resulted in the highest number of bacterial isolates (40) which accounted for 10% of the total isolates. AIM and ZBA resulted in 39 (10%) and 37 (9%) isolates respectively (Figure 2). The medium which resulted in the fewest bacterial isolates were MMM and Seawater Agar (SWA) since each contributed only one bacterial isolate. Both media discourage growth of fast growing bacteria thus resulting in fewer numbers of microbial isolates.
Figure 2.

Total number (and percentage) of bacterial isolates cultured from the marine sponge samples PE07 and PE09 per medium type. The number shown inside each pie section indicates the raw number of isolates from that medium type. Only 1 isolate was recorded from MMM and SWA respectively.
3.2. Antimicrobial Screening and Induction
The antibacterial properties of microbial isolates from PE07 and PE09 were investigated in an agar overlay assay against five test strains (Table 2). Most isolates only showed a narrow spectrum of activity, being active against only one test strain when cultured on standard media (ZBA, GYM, TSA and ACM). However, isolates PE07-7, PE07-144, PE07-207, PE09-87, PE09-105 and PE09-119 displayed activity against two or more test strains. Notably, PE07-7 and PE09-119 only displayed activity against Gram-positive test strains, while PE07-144, PE07-207 and PE09-105 had activity against both Gram-positive and Gram-negative test strains. Most activity was recorded against B. cereus with 17 out of 32 strains showing activity against this test strain, while seven isolates gave activity against the highly antibiotic resistant E. coli 1699 (resistant to 52 known antibiotics). It was also noted that none of the isolates displayed antibacterial activity on more than two growth media, suggesting inducible genes or pathways. It is also important to note that all isolates grew to some degree on the media used and lack of activity is not due to the inability of the isolate(s) to grow.
Table 2.
Antibacterial activities against test strains in standard medium.
| Isolate | Test Strains | ||||
|---|---|---|---|---|---|
| Gram Negative | Gram Positive | ||||
| E. coli 1699 | P. putida | M. smegmatis | B. cereus | S. epidermidis | |
| PE07-5 | GYM | GYM | ZBA | ||
| PE07-7 | TSA, ZBA | TSA, ZBA | |||
| PE07-86 | TSA | ||||
| PE07-105 | GYM | ||||
| PE07-133 | GYM | ||||
| PE07-143 | TSA, ZBA | ||||
| PE07-144 | GYM | GYM | |||
| PE07-172 | TSA | ||||
| PE07-200 | GYM | ||||
| PE07-201 | GYM | ||||
| PE07-204 | GYM | ||||
| PE07-207 | GYM | GYM | |||
| PE09-7 | TSA | ||||
| PE09-73 | ACM | ||||
| PE09-87 | GYM | GYM | GYM | ||
| PE09-100 | ZBA | ||||
| PE09-105 | GYM, TSA | TSA, GYM | TSA | ||
| PE09-108 | GYM | ||||
| PE09-110 | ZBA | ||||
| PE09-119 | ZBA, GYM | TSA, ZBA | ACM, TSA | ||
| PE09-124 | ACM | ||||
| PE09-140 | ACM | ||||
| PE09-142 | ACM | ||||
| PE09-168 | ACM | ||||
| PE09-197 | ACM | ||||
| PE09-210 | ACM | ||||
| PE09-213 | ACM | ||||
| PE09-221 | ZBA | ||||
| PE09-228 | ACM | ||||
| PE09-229 | ACM | ||||
| PE09-235 | ACM | ||||
| PE09-266 | ZBA | ||||
| Total hits | 7 | 5 | 8 | 17 | 6 |
To further induce antimicrobial activities, an OSMAC approach was taken for a subset of the isolates. From screening 29 microbial isolates in the matrix of 36 culture/treatment conditions and co-culture, six isolates showed induced antibacterial activities (Table 3). Most notably, isolate PE09-72 produced activity against all test strains in mannitol containing medium, especially with the addition of heat shock, whereas only one activity was produced with succinic acid as the carbon source. This indicates that the pathway(s) responsible for the activities are likely subject to carbon catabolite repression (CCR) [62]. Although no medium composition induced activities against all test strains, some medium compositions (NH4Cl-LP, NH4Cl-HP and NaNO3-HP) induced activity in PE09-72 against four of the test strains when mannitol was used as carbon source and the culture heat shocked.
Table 3.
Antibacterial activities of six selected sponge-associated isolates under matrix screening conditions. Three of the isolates did not display activity in the initial screening, whereas the other three isolates did. RT: Room Temperature (27 °C); HS: Heat Shock treatment (42 °C); CS: Cold shock treatment (4 °C).
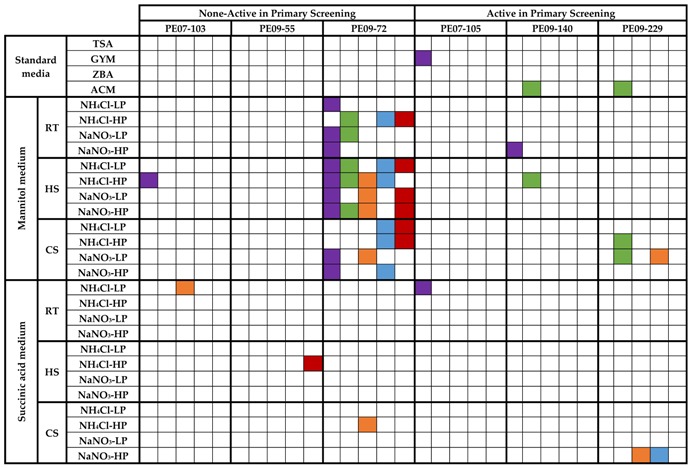
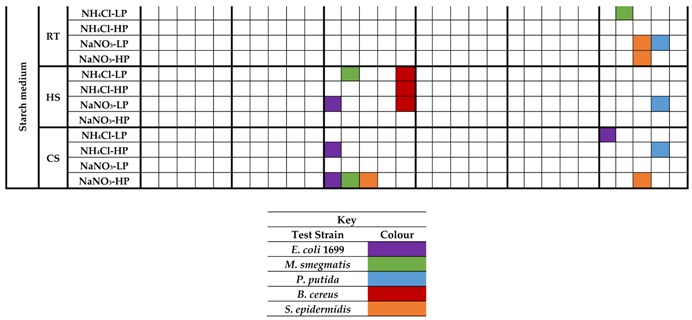
Differential antimicrobial activities were observed in relation to the media the isolates were cultured on. For instance, the isolate PE07-5 showed antibacterial activities against E. coli 1699 and M. smegmatis in GYM medium and only against B. cereus on ZBA medium. This is a well-known phenomenon regarding microbial secondary metabolite production [8,63].
3.3. 16S rRNA Gene Sequence Analysis of Bacteria Producing Antimicrobial Activity
Twenty-six of the bacterial isolates which showed antibacterial activities against the test strains were identified by determination of the sequence of the 16S rRNA gene sequence (Table 4). The identities ranged from 96.5% to 100% identity over a region of 1340 bp and the bacteria belonged to the following phyla: Actinobacteria, Firmicutes and Proteobacteria (Alphaproteobacteria and Gammaproteobacteria). Unexpectedly the Firmicutes and Gammaproteobacteria were the phyla which showed the most antibacterial activities as opposed to the Actinobacteria, well-known for their antibiotic activities [9,64,65].
Table 4.
16S rRNA gene taxonomic affiliation of 26 of the sponge-associated bacterial isolates showing antibacterial activities against one or more indicator strains.
| Phylum/Class | Closest Relative (as Present in EzTaxon) | Identity (%) | Isolate | Isolation Medium |
|---|---|---|---|---|
| Actinobacteria | Aeromicrobium erythreum NRRL B-3381 | 96.5 | PE09-221 | GYM |
| Arthrobacter citreus DSM 20133 | 99.9 | PE09-119 | ACM | |
| Brevibacterium luteolum CF87 | 100 | PE09-168 | GYM | |
| Citricoccus nitrophenolicus PNP1 | 99.7 | PE09-100 | SCN | |
| Citricoccus nitrophenolicus PNP1 | 99.8 | PE09-110 | 172 | |
| Curtobacterium oceanosedimentum ATCC 31317 | 99.3 | PE09-7 | GYM | |
| Micrococcus yunnanensis YIM 65004 | 99.7 | PE07-133 | R2A | |
| Firmicutes | Bacillus kochii WCC 4582 | 99.5 | PE07-103 | MAA |
| Bacillus marisflavi TF-11 | 100 | PE07-7 | ZBA | |
| Bacillus vietnamesis B-23890 | 99.7 | PE07-5 | ZBA | |
| Bacillus zhangzhouensis DW5-4 | 99.9 | PE09-72 | ZBA | |
| Bacillus vietnamesis B-23890 | 99.7 | PE07-144 | ZBA | |
| Oceanobacillus picturae LMG 19492 | 99.5 | PE07-105 | MAA | |
| Oceanobacillus picturae LMG 19492 | 100 | PE09-108 | ZBA | |
| Sporosarcina aquimarina SW28 | 98.8 | PE07-172 | 172 | |
| Staphylococcus cohnii ATCC 29974 | 100 | PE07-204 | ZBA | |
| Staphylococcus epidermidis ATCA 1490 | 99.9 | PE07-201 | TSA | |
| Staphylococcus warneri ATCC 27836 | 99.9 | PE07-200 | TSA | |
| Gammaproteobacteria | Halomonas titanicae | 99.9 | PE09-210 | CCA |
| Halomonas titanicae | 99.9 | PE09-228 | SEM | |
| Halomonas titanicae | 99.9 | PE09-229 | SEM | |
| Kushneria pakistanensis NCCP-934 | 98.9 | PE09-73 | PMM | |
| Kushneria pakistanensis NCCP-934 | 98.9 | PE09-124 | ACM | |
| Kushneria pakistanensis NCCP-934 | 98.9 | PE09-266 | ANFA | |
| Pseudomonas fulva NBRC 16637 | 100 | PE09-140 | SNA | |
| Pseudomonas fulva NBRC 16637 | 100 | PE09-197 | ANFA | |
| Psychrobacter alimentarius JG-100 | 99.9 | PE09-87 | ACM | |
| Alphaproteobacteria | Pseudovibrio ascidiaceicola DSM 16392 | 100 | PE09-55 | TSA |
In some instances, isolates that shared 99% 16S rRNA gene sequence identity differed in their bioactivity profiles and were thus considered to be different OTUs. An example of this was observed with isolates PE09-100 and PE09-110 which were both identified as Citricoccus nitrophenolicus whereas their bioactivities differed (Table 2). Similar analyses of five strains of Pseudoalteromonas luteoviolacea revealed that the strains were closely related based on 16S rRNA gene sequences however they produced different bioactivity profiles [66,67].
Isolate PE09-72, which showed most activity as described above, was identified as a Bacillus zhangzhouensis strain, a newly described bacterial species [68] that belongs to the Bacillus pumilus clade, a bacterium known for its antibiotic producing ability [69,70,71,72]. Bacillus marisflavi, Psychrobacter alimentarius and Arthrobacter citreus which PE07-7, PE09-87 and PE09-119 were identified as respectively, are here, for the first time, shown to display antimicrobial activity. Arthrobacter citreus is often used as test organism against which to test for antimicrobial activity, yet here it displays antibacterial properties against both Gram-positive and Gram-negative bacteria [73,74], while B. marisflavi and P. alimentarius are relatively newly described species [75,76].
The most novel isolate at the time of isolation was PE09-221, which has 96.5% identity to Aeromicrobium erythreum. It has subsequently been shown to be a strain of the recently described Aeromicrobium camelliae species which is also closely related to another newly described species Aeromicrobium halotolerans (Figure 3) [77,78]. Thus, our study represents only the second isolation of this species. Of the thirteen described Aeromicrobium species, four have been isolated from marine environments with one, Aeromicrobium halocynthiae, isolated from a marine ascidian. A. camelliae, to which PE09-221 is most closely related, was isolated from Pu’er tea demonstrating that these organisms are not sponge specific and can occupy a wide range of habitats.
Figure 3.

Neighbor-joining tree generated by analysing partial (approximately 1340 bp) 16S rRNA gene sequences of Aeromicrobium sp. compared with isolate PE09-221.
3.4. PE09-72 and PE09-221 Genome Sequence and Secondary Metabolism
Two isolates were selected for whole genome sequencing: PE09-72, which was the most bioactive isolate under the widest range of test conditions; and PE09-221, due to its novelty at the 16S rRNA sequence level and no genome sequence being available for any isolate of this species.
The draft genome sequence of PE09-72 is 3.69 Mbp consisting of 109 contigs with an N50 value of 68,385 bp and the largest contig was 316,332 bp. BAGEL3 and antiSMASH analysis of the contigs revealed the presence of eight putative secondary metabolite pathways which could be responsible for the range of activities observed. These include three nonribosomal peptides (NRP), one of which is a hybrid nonribosomal peptide/polyketide operon; a terpene; two bacteriocins; a Type III polyketide and a predicted siderophore-terpene hybrid. Derivatives of all these pathways can be found in Bacillus genome sequences currently on the Genbank database with similarity between 40% and 85%. The hybrid NRP/PKS pathway shows weak similarity with the Zwittermycin-A pathway and likely produces a novel compound.
Two of the pathways are related to known antimicrobial compound producing pathways, namely lichenysin and bacilysin, showing 85% similarity in each case. Interestingly the lichenysin-like pathway contains two biosynthetic genes in addition to those found in the characterized lichenysin and related surfactin pathways (Figure 4). Each of these genes harbours a ketoreductase (KR) domain, which is an unusual part of the domain structure of NRP biosynthetic genes. However, when present, they are thought to perform chiral reduction of α-ketoacyl-S carrier proteins generating an ester bond [79,80]. Taken together, the additional biosynthetic genes and unusual domain structure indicate that a novel lichenysin-like compound could be produced from this pathway. These same two open reading frames can be identified in similar pathways on several Bacillus species genomes currently on the Genbank database, indicating that this modified, uncharacterized pathway is likely common in a range of Bacilli. Even so, differences can still be observed between the PE09-72 pathway and its closest relative (Figure 4).
Figure 4.

tBLASTx comparison of lichenysin-like pathway identified in PE09-72 with the lichenysin and surfactin pathways as well as the most similar pathway identified on a genome currently on Genbank. Gene annotations are as follow: Pink—condensation domain containing protein; Green—condensation domain containing protein; Dark blue—condensation domain containing protein which includes a thioesterase domain; Light blue—two “extra” genes present in the pathway from PE09-72 which contain ketoreductase domains, the downstream gene also contains a thioesterase domain; Orange—other ORF’s (biosynthetic and transport related) identified by antiSMASH which delineates the extent of the pathways. Similarity at the amino acid level in the repeated domain structures results in the criss-cross pattern observed when comparing the biosynthetic genes.
Of the seven genes necessary (bacA-E, ywfA and ywfG) for production of bacilysin, only six appear to be present. A bacE homologue (bacilysin export protein) could not be detected on the PE09-72 genome. Considering the genes downstream (short chain dehydrogenase, NAD-dependent epimerase, glucose-1-phosphate adenylyl/thymidylyl transferase, glycosyltransferase, aldehyde dehydrogenase and polyprenol-monophosphomannose synthase) of the bacilysin-like genes, it is possible that they could form part of a larger pathway than that delineated by antiSMASH to produce a bacilysin-like compound.
As mentioned previously, the pathway(s) responsible for the observed antibacterial activity in PE09-72 may be subject to CCR. A search for the B. pumilus cre (WTGNAARCGNWWWCA) consensus [81] indicated a potential site (TTGAAAACGAATTCA) located between two ORFs (14,389–14,403 bp) on contig 33 on which the bacilysin-like pathway was identified with a beta-glucoside transporter located 7 kb downstream on the same contig. Bacilysin production in Bacillus subtilis is however not subject to glucose repression [82]. Another putative cre site (ATGTAAGCGGTAACA) was identified directly upstream (5936–5950 bp) of a four ORF operon, with the translational start at 5968 bp, predicted to be part of the bacteriocin cluster on contig 38. The ORFs encode an acetyl-CoA acetyltransferase, a 3-hydroxybutyryl-CoA dehydrogenase and two acyl-CoA dehydrogenases. No Bacillus-like CodY binding sites (AATTTTCWGAAAATT) could be identified in any of the secondary metabolite pathways identified, nor could the MtlR (Transcriptional activator of mannitol metabolism) recognition site (TTGTCACANNNNNTGTGCCAA) be identified anywhere except upstream of the mtlR gene. The observation that activity was turned on against all test strains under mannitol, as opposed to little or no activity on succinic acid, either suggests that one pathway, the product of which shows broad-spectrum activity, was activated, or demonstrates that several of the pathways are subject to CCR.
BAGEL3 identified the presence of a plantazolicin-like peptide and the pathway responsible for its maturation (contig 17 from 98,257 bp to 108,183 bp). These small ribosomally encoded peptides belong to the thiazole/oxazole-modified microcins (TOMM) class of natural products and show a very narrow-spectrum of activity against related Bacillus species. Based on the similarity in the core region of the precursor peptide as well as high amino acid identity with the enzymes involved in maturation of the peptide from other B. pumilus strains, PE09-72 likely produces the exact same molecule described by Molohon and coworkers [83]. It is therefore probable that the activity against B. cereus observed here could be as a consequence of the production of this molecule.
The draft genome of PE09-221 (A. camelliae) is 4.01 Mbp and consists of 128 contigs greater than 450 bp with an N50 value of 147,047 bp, with the largest contig being 284,975 bp. The genome sequence of PE09-221 represents the first genome for an A. camelliae isolate. The G+C content is 67.5% in line with that found for A. camelliae (66%), but lower than for most other Aeromicrobium species 71%–74% [84,85], with the exception of A. halotolerans (44.7%). As intra genus G+C content does not vary greatly (±10%), the low G+C content observed in A. halotolerans, suggests that the genus may yet be split into more genera [80,86]. As no genome sequence for A. camelliae or A. halotolerans are available for comparison, comparison of the PE09-221 genome could only be made with the next closest relatives as identified through comparison of the 16S rRNA sequence. Average amino acid identity is 64.8% when compared with A. erythreum AR18 and 64.4% when compared with Aeromicrobium massiliense JC14. Similarly, the average nucleotide identities were well below the 94-96% cut-off used to delineate new species with values of 79.1% and 78.7% when compared with A. erythreum AR18 and A. massiliense JC14, respectively [87]. This shows the utility of culturing, despite its limitations, in still isolating novel species. Unlike many other actinobacteria, Aeromicrobium species are not well known for their antibiotic production, with the exception of the broad-spectrum macrolide, erythromycin (A. erythreum). PE09-221 did show activity against B. cereus and it is possible that one of the two NRP pathways present, one of which is a NRP/PKS hybrid, are responsible for this observed activity. Neither pathway shows similarity to any currently characterized pathway.
4. Conclusions
Here we report the first study of bacteria associated with two endemic South African marine sponges and provide the first draft genome for the recently described species A. camelliae. Although no entirely novel bacteria were isolated, many of those that were are related to recently described and therefore mostly uncharacterized species. This suggests that these sponges harbour species not routinely isolated. As evidenced by the antimicrobial activities observed for the bacterial isolates and the genome sequences of two of these isolates, endemic South African marine sponges could offer a wealth of new, microbial derived chemistry. In particular, the range and number of the pathways identified for PE09-72 indicates that it is an organism with an impressive arsenal, subject to stringent control. The study serves to further demonstrate the utility of culture-dependent approaches as a valuable tool to tap into marine-derived natural products.
Acknowledgments
This work was funded by the Medical Research Council (MRC) of South Africa and by the European Union PharmaSea consortium as part of the European Commission seventh framework programme (312184).
Supplementary Materials
The following are available online at www.mdpi.com/1660-3397/15/2/47/s1, Figure S1: Sponges used in this study: PE05—Waltherarndtia caliculatum, PE07—Higginsia bidentifera, PE08—Spongia sp., PE09—Isodictya compressa, PE11—Axinella sp., Figure S2: Flow diagram showing the culturing process used in this study, Figure S3: 96-well plate setup for matrix screening. Bioactivities of the isolate PE09-72 when cultured on Na5HM under co-culture conditions with 3 and 4 “immediate” neighbouring isolates in 96 well-plate set up, Table S1: Media used to isolate sponge symbionts, Table S2: Panel of 29 isolates selected for the matrix study; 12 isolates which showed antibacterial activities after primary screening and 17 isolates which did not show any bioactivities.
Author Contributions
M.T. and M.T.D.-C. conceived the study, S.P.-N. collected sponge specimens and performed identification. R.M.M. performed all experiments except genome sequencing and helped draft the manuscript. L.J.v.Z. performed genome sequencing and analysis as well as wrote the bulk of the manuscript. All authors contributed to editing of, and approved of, the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- 1.Piel J. Bacterial symbionts: Prospects for the sustainable production of invertebrate-derived pharmaceuticals. Curr. Med. Chem. 2006;13:39–50. doi: 10.2174/092986706775197944. [DOI] [PubMed] [Google Scholar]
- 2.Hentschel U., Usher K.M., Taylor M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006;55:167–177. doi: 10.1111/j.1574-6941.2005.00046.x. [DOI] [PubMed] [Google Scholar]
- 3.Hardoim C.C.P., Costa R., Araújo F.V., Hajdu E., Peixoto R., Lins U., Rosado A.S., van Elsas J.D. Diversity of bacteria in the marine sponge Aplysina fulva in Brazilian coastal waters. Appl. Environ. Microbiol. 2009;75:3331–3343. doi: 10.1128/AEM.02101-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Van Soest R.W.M., Boury-Esnault N., Vacelet J., Dohrmann M., Erpenbeck D., De Voogd N.J., Santodomingo N., Vanhoorne B., Kelly M., Hooper J.N.A. Global diversity of sponges (Porifera) PLoS ONE. 2012;7:e35105. doi: 10.1371/journal.pone.0035105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Schmitt S., Weisz J.B., Lindquist N., Hentschel U. Vertical transmission of a phylogenetically complex microbial consortium in the viviparous sponge Ircinia felix. Appl. Environ. Microbiol. 2007;73:2067–2078. doi: 10.1128/AEM.01944-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bayer K., Scheuermayer M., Fieseler L., Hentschel U. Genomic mining for novel FADH2-dependent halogenases in marine sponge-associated microbial consortia. Mar. Biotechnol. 2013;15:63–72. doi: 10.1007/s10126-012-9455-2. [DOI] [PubMed] [Google Scholar]
- 7.Zeng Z., Zhao J., Ke C., Wang D. Antimicrobial activities of novel cultivable bacteria isolated from marine sponge Tedania anhelans. Chin. J. Oceanol. Limnol. 2013;31:581–590. doi: 10.1007/s00343-013-2223-y. [DOI] [Google Scholar]
- 8.Muscholl-Silberhorn A., Thiel V., Imhoff J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microb. Ecol. 2008;55:94–106. doi: 10.1007/s00248-007-9255-9. [DOI] [PubMed] [Google Scholar]
- 9.Walmsley T.A., Matcher G.F., Zhang F., Hill R.T., Davies-Coleman M.T., Dorrington R.A. Diversity of bacterial communities associated with the Indian Ocean sponge Tsitsikamma favus that contains the bioactive pyrroloiminoquinones, tsitsikammamine A and B. Mar. Biotechnol. 2012;14:681–691. doi: 10.1007/s10126-012-9430-y. [DOI] [PubMed] [Google Scholar]
- 10.Xi L., Ruan J., Huang Y. Diversity and biosynthetic potential of culturable actinomycetes associated with marine sponges in the China seas. Int. J. Mol. Sci. 2012;13:5917–5932. doi: 10.3390/ijms13055917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fieseler L., Horn M., Wagner M., Hentschel U. Discovery of the novel candidate phylum Poribacteria in marine sponges. Appl. Environ. Microbiol. 2004;70:3724–3732. doi: 10.1128/AEM.70.6.3724-3732.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Webster N.S., Taylor M.W. Marine sponges and their microbial symbionts: Love and other relationships. Environ. Microbiol. 2012;14:335–346. doi: 10.1111/j.1462-2920.2011.02460.x. [DOI] [PubMed] [Google Scholar]
- 13.Gerwick W.H., Fenner A.M. Drug Discovery from Marine Microbes. Microb. Ecol. 2013;65:800–806. doi: 10.1007/s00248-012-0169-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kennedy J., Codling C.E., Jones B.V., Dobson A.D., Marchesi J.R. Diversity of microbes associated with the marine sponge, Haliclona simulans, isolated from Irish waters and identification of polyketide synthase genes from the sponge metagenome. Environ. Microbiol. 2008;10:1888–1902. doi: 10.1111/j.1462-2920.2008.01614.x. [DOI] [PubMed] [Google Scholar]
- 15.Fusetani N. Antifungal peptides in marine invertebrates. Invert. Surv. J. 2010;7:53–66. [Google Scholar]
- 16.Phelan R.W., O’Halloran J.A., Kennedy J., Morrissey J.P., Dobson A.D., O’Gara F., Barbosa T.M. Diversity and bioactive potential of endospore-forming bacteria cultured from the marine sponge Haliclona simulans. J. Appl. Microbiol. 2012;112:65–78. doi: 10.1111/j.1365-2672.2011.05173.x. [DOI] [PubMed] [Google Scholar]
- 17.Skariyachan S., Rao A.G., Patil M.R., Saikia B., Bharadwaj V.K.N., Rao J.G.S. Antimicrobial potential of metabolites extracted from bacterial symbionts associated with marine sponges in coastal area of Gulf of Mannar Biosphere, India. Lett. Appl. Microbiol. 2014;58:231–241. doi: 10.1111/lam.12178. [DOI] [PubMed] [Google Scholar]
- 18.Bhatnagar I., Kim S.K. Immense essence of excellence: Marine microbial bioactive compounds. Mar. Drugs. 2010;8:2673–2701. doi: 10.3390/md8102673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Woodhouse J.N., Fan L., Brown M.V., Thomas T., Neilan B.A. Deep sequencing of non-ribosomal peptide synthetases and polyketide synthases from the microbiomes of Australian marine sponges. ISME J. 2013;7:1842–1851. doi: 10.1038/ismej.2013.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Bergman O., Haber M., Mayzel B., Anderson M.A., Shpigel M., Hill R.T., Ilan M. Marine-based cultivation of diacarnus sponges and the bacterial community composition of wild and maricultured sponges and their larvae. Mar. Biotechnol. 2011;13:1169–1182. doi: 10.1007/s10126-011-9391-6. [DOI] [PubMed] [Google Scholar]
- 21.Steinert G., Whitfield S., Taylor M.W., Thoms C., Schupp P.J. Application of diffusion growth chambers for the cultivation of marine sponge-associated bacteria. Mar. Biotechnol. 2014;16:594–603. doi: 10.1007/s10126-014-9575-y. [DOI] [PubMed] [Google Scholar]
- 22.Song J., Oh H.M., Cho J.C. Improved culturability of SAR11 strains in dilution-to-extinction culturing from the East Sea, West Pacific Ocean. FEMS Microbiol. Lett. 2009;295:141–147. doi: 10.1111/j.1574-6968.2009.01623.x. [DOI] [PubMed] [Google Scholar]
- 23.Bollmann A., Lewis K., Epstein S.S. Incubation of environmental samples in a diffusion chamber increases the diversity of recovered isolates. Appl. Environ. Microbiol. 2007;73:6386–6390. doi: 10.1128/AEM.01309-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Joint I., Mühling M., Querellou J. Culturing marine bacteria—An essential prerequisite for biodiscovery. Microb. Biotechnol. 2010;3:564–575. doi: 10.1111/j.1751-7915.2010.00188.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Branch G.M., Griffiths C.L., Branch M.L., Beckley L.E. Two Oceans. A Guide to the Marine Life of Southern Africa. Struik Nature; Cape Town, South Africa: 2010. [Google Scholar]
- 26.Griffiths C.L., Robinson T.B., Lange L., Mead A. Marine biodiversity in South Africa: An evaluation of current states of knowledge. PLoS ONE. 2010;5:e12008. doi: 10.1371/journal.pone.0012008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Antunes E.M., Beukes D.R., Kelly M., Samaai T., Barrows L.R., Marshall K.M., Sincich C., Davies-Coleman M.T. Cytotoxic pyrroloiminoquinones from four new species of South African Latrunculid sponges. J. Nat. Prod. 2004;67:1268–1276. doi: 10.1021/np034084b. [DOI] [PubMed] [Google Scholar]
- 28.Samaai T., Gibbons M.J., Kerwath S., Yemane D., Sink K. Sponge richness along a bathymetric gradient within the Isimangaliso Wetland Park, South Africa. Mar. Biodivers. 2010;40:205–217. doi: 10.1007/s12526-010-0046-z. [DOI] [Google Scholar]
- 29.Higgin T. Description of some Sponges obtained during a Cruise of the Steam-Yacht ‘Argo’ in the Caribbean and neighbouring Seas. J. Nat. Hist. 1877;19:291–299. doi: 10.1080/00222937708682143. [DOI] [Google Scholar]
- 30.Ridley S.O., Dendy A. Preliminary Report on the Monaxonida collected by H.M.S. ‘Challenger’. Ann. Mag. Nat. Hist. 1886;18:325–351, 470–493. doi: 10.1080/00222938609459982. [DOI] [Google Scholar]
- 31.Esper E.C.J. Die Pflanzenthiere in Abbildungen Nach der Natur Mit Farben Erleuchtet, Nebst Beschreibungen, Zweyter Theil. Raspe; Nürnberg, Germany: 1794. pp. 1–303. [Google Scholar]
- 32.Topsent E. Éponges de Lamarck conservées au Muséum de Paris. Archives du Muséum National D’histoire Naturelle. 1931;5:1–56. [Google Scholar]
- 33.Kirkpatrick R. Descriptions of South African Sponges. Part III. Mar. Investig. S. Afr. 1903;2:233–264. [Google Scholar]
- 34.Vimala A., Xavier Innocent B., Huxley V.A.J. Phylogenetic investigations on the endosymbiotic bacteria of Axinella donnani. J. Microb. Biochem. Technol. 2015;7:318–322. [Google Scholar]
- 35.Haber M., Ilan M. Diversity and antibacterial activity of bacteria cultured from Mediterranean Axinella spp. sponges. J. Appl. Microbiol. 2014;116:519–532. doi: 10.1111/jam.12401. [DOI] [PubMed] [Google Scholar]
- 36.Bauvais C., Zirah S., Piette L., Chaspoul F., Domart-Coulon I., Chapon V., Gallice P., Rebuffat S., Perez T., Bourguet-Kondracki M.-L. Sponging up metals: Bacteria associated with the marine sponge Spongia officinalis. Mar. Environ. Res. 2015;104:20–30. doi: 10.1016/j.marenvres.2014.12.005. [DOI] [PubMed] [Google Scholar]
- 37.Noyer C., Casamayor E.O., Becerro M.A. Environmental heterogeneity and microbial inheritance influence sponge-associated bacterial composition of Spongia lamella. Microb. Ecol. 2014;68:611–620. doi: 10.1007/s00248-014-0428-z. [DOI] [PubMed] [Google Scholar]
- 38.Liles M.R., Williamson L.L., Rodbumrer J., Torsvik V., Goodman R.M., Handelsman J. Recovery, purification, and cloning of high-molecular-weight DNA from soil microorganisms. Appl. Environ. Microbiol. 2008;74:3302–3305. doi: 10.1128/AEM.02630-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Hansen M.C., Tolker-Nelson T., Givskov M., Molin S. Biased 16S rDNA PCR amplification caused by interference from DNA flanking template region. FEMS Microbiol. Ecol. 1998;15:25–36. doi: 10.1111/j.1574-6941.1998.tb00500.x. [DOI] [Google Scholar]
- 40.Baker G.C., Smith J.J., Cowan D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Meth. 2003;55:541–555. doi: 10.1016/j.mimet.2003.08.009. [DOI] [PubMed] [Google Scholar]
- 41.Culman S.W., Bukowski R., Gauch H.G., Cadillo-Quiroz H., Buckley D.H. T-REX: Software for the processing and analysis of T-RFLP data. [(accessed on 3 February 2017)];BMC Bioinform. 2009 10:171. doi: 10.1186/1471-2105-10-171. Available online: http://trex.biohpc.org/ [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Blackwood C.B., Hudleston D., Zak D.R., Buyer J.S. Interpreting ecological diversity indices applied to terminal restriction fragment length polymorphism data: Insights from simulated microbial communities. Appl. Environ. Microbiol. 2007;73:5276–5283. doi: 10.1128/AEM.00514-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nocker A., Burr M., Camper A.K. Genotypic microbial community profiling: A critical technical review. Microb. Ecol. 2007;54:276–289. doi: 10.1007/s00248-006-9199-5. [DOI] [PubMed] [Google Scholar]
- 44.Schütte U.M.E., Abdo Z., Bent S.J., Shyu C., Williams C.J., Pierson J.D., Forney L.J. Advances in the use of terminal restriction fragment length polymorphism (T-RFLP) analysis of 16S rRNA genes to characterize microbial communities. Appl. Microbiol. Biotechnol. 2008;80:365–380. doi: 10.1007/s00253-008-1565-4. [DOI] [PubMed] [Google Scholar]
- 45.Ramond J.-B., Tshabuse F., Bopda C.W., Cowan D.A., Tuffin M.I. Evidence of variability in the structure and recruitment of rhizospheric and endophytic bacterial communities associated with arable sweet sorghum (Sorghum bicolor (L) Moench) Plant Soil. 2013;372:265–278. doi: 10.1007/s11104-013-1737-6. [DOI] [Google Scholar]
- 46.Lavy A., Keren R., Haber M., Schwartz I., Ilan M. Implementing sponge physiological and genomic information to enhance the diversity of its culturable associated bacteria. FEMS Microbiol. Ecol. 2014;87:486–502. doi: 10.1111/1574-6941.12240. [DOI] [PubMed] [Google Scholar]
- 47.Atlas R.M. Handbook of Microbiological Media. 4th ed. CRC Press; Florence, KY, USA: 2010. pp. 1–2040. [Google Scholar]
- 48.Yang Y., Zhang J., Wu H., Gong M. Genetic diversity analysis of Azotobacter in cotton soil of Yangtze River Basin. J. Chem. Pharm. Res. 2013;5:54–58. [Google Scholar]
- 49.Sipkema D., Schippers K., Maalcke W.J., Yang Y., Salim S., Blanch H.W. Multiple approaches to enhance the cultivability of bacteria associated with the marine sponge Haliclona (Gellius) sp. Appl. Environ. Microbiol. 2011;77:2130–2140. doi: 10.1128/AEM.01203-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Intra B., Mungsuntisuk I., Nihira T., Igarashi Y., Panbangred W. Identification of actinomycetes from plant rhizospheric soils with inhibitory activity against Colletotrichum spp., the causative agent of anthracnose disease. BMC Res. Notes. 2011;4:98. doi: 10.1186/1756-0500-4-98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bills G.F., Platas G., Fillola A., Jiménez M.R., Collado J., Vicente F., Martín J., González A., Bur-Zimmermann J., Tormo J.R., et al. Enhancement of antibiotic and secondary metabolite detection from filamentous fungi by growth on nutritional arrays. J. Appl. Microbiol. 2008;104:1644–1658. doi: 10.1111/j.1365-2672.2008.03735.x. [DOI] [PubMed] [Google Scholar]
- 52.Nakata K., Yoshimoto A., Yamada Y. Promotion of antibiotic production by high ethanol, high NaCl concentration, or heat shock in Pseudomonas fluorescens S272. Biosci. Biotechnol. Biochem. 1999;63:293–297. doi: 10.1271/bbb.63.293. [DOI] [PubMed] [Google Scholar]
- 53.Kumar S., Stecher G., Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. [(accessed on 8 February 2017)];Mol. Biol. Evol. 2016 33:1870–1874. doi: 10.1093/molbev/msw054. Available online: http://www.megasoftware.net/ [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Illumina Nextera® XT DNA Sample Preparation Guide, Part # 15031942 Rev. C, 2012. [(accessed on 8 February 2017)]. Available online: https://tinyurl.com/jfd867j.
- 55.Aziz R.K., Bartels D., Best A.A., DeJongh M., Disz T., Edwards R.A., Formsma K., Gerdes S., Glass E.M., Kubal M., et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008;9:75. doi: 10.1186/1471-2164-9-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.NCBI Resource Coordinators Database resources of the National Center for Biotechnology Information. [(accessed on 8 February 2017)];Nucleic Acids Res. 2016 44:7–19. doi: 10.1093/nar/gkv1290. Available online: https://www.ncbi.nlm.nih.gov/ [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Schmitt S., Tsai P., Bell J., Fromont J., Ilan M., Lindquist N., Perez T., Rodrigo A., Schupp P.J., Vacelet J., et al. Assessing the complex sponge microbiota: Core, variable and species-specific bacterial communities in marine sponges. ISME J. 2012;6:564–576. doi: 10.1038/ismej.2011.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Taylor M.W., Tsai P., Simister R.L., Deines P., Botte E., Ericson G., Schmitt S., Webster N.S. “Sponge-specific” bacteria are widespread (but rare) in diverse marine environments. ISME J. 2013;7:438–443. doi: 10.1038/ismej.2012.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Simister R.L., Taylor M.W., Tsai P., Fan L., Bruxner T.J., Crowe M.L., Webster N. Thermal stress responses in the bacterial biosphere of the great barrier reef sponge, Rhopaloeides odorabile. Environ. Microbiol. 2012;14:3232–3246. doi: 10.1111/1462-2920.12010. [DOI] [PubMed] [Google Scholar]
- 60.Simister R.L., Deines P., Botté E.S., Webster N.S., Taylor M.W. Sponge-specific clusters revisited: A comprehensive phylogeny of sponge-associated microorganisms. Environ. Microbiol. 2012;14:517–524. doi: 10.1111/j.1462-2920.2011.02664.x. [DOI] [PubMed] [Google Scholar]
- 61.Erwin P.M., Olson J.B., Thacker R.W. Phylogenetic diversity, host-specificity and community profiling of sponge-associated bacteria in the northern Gulf of Mexico. PLoS ONE. 2011;6:e26806. doi: 10.1371/journal.pone.0026806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sanchez S., Demain A.L. Metabolic regulation of fermentation processes. Enzyme Microb. Technol. 2002;31:895–906. doi: 10.1016/S0141-0229(02)00172-2. [DOI] [Google Scholar]
- 63.Rateb M.E., Houssen W.E., Harrison W.T., Deng H., Okoro C.K., Asenjo J.A., Andrews B.A., Bull A.T., Goodfellow M., Ebel R., et al. Diverse metabolic profiles of a Streptomyces strain isolated from a hyper-arid environment. J. Nat. Prod. 2011;74:1965–1971. doi: 10.1021/np200470u. [DOI] [PubMed] [Google Scholar]
- 64.Sun W., Dai S., Jiang S., Wang G., Liu G., Wu H., Li X. Culture-dependent and culture-independent diversity of Actinobacteria associated with the marine sponge Hymeniacidon perleve from the South China Sea. Antonie van Leeuwenhoek. 2010;98:65–75. doi: 10.1007/s10482-010-9430-8. [DOI] [PubMed] [Google Scholar]
- 65.Ouyang Y., Wu H., Xie L., Wang G., Dai S., Chen M., Yang K., Li X. A method to type the potential angucycline producers in actinomycetes isolated from marine sponges. Antonie van Leeuwenhoek. 2011;99:807–815. doi: 10.1007/s10482-011-9554-5. [DOI] [PubMed] [Google Scholar]
- 66.Vynne N.G., Mansson M., Gram L. Gene sequence based clustering assists in dereplication of Pseudoalteromonas luteoviolacea strains with identical inhibitory activity and antibiotic production. Mar. Drugs. 2012;10:1729–1740. doi: 10.3390/md10081729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Duncan K.R., Haltli B., Gill K.A., Correa H., Berrué F., Kerr R.G. Exploring the diversity and metabolic potential of actinomycetes from temperate marine sediments from Newfoundland, Canada. J. Ind. Microbiol. Biotechnol. 2015;42:57–72. doi: 10.1007/s10295-014-1529-x. [DOI] [PubMed] [Google Scholar]
- 68.Liu Y., Lai Q., Du J., Shao Z. Bacillus zhangzhouensis sp. nov. and Bacillus australimaris sp. nov. Int. J. Syst. Evol. Microbiol. 2016;66:1193–1199. doi: 10.1099/ijsem.0.000856. [DOI] [PubMed] [Google Scholar]
- 69.Prieto M.L., O’Sullivan L., Tan S.P., McLoughlin P., Hughes H., O’Connor P.M., Cotter P.D., Lawlor P.G., Gardiner G.E. Assessment of the bacteriocinogenic potential of marine bacteria reveals lichenicidin production by seaweed-derived Bacillus spp. Mar. Drugs. 2012;10:2280–2299. doi: 10.3390/md10102280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sumi C.D., Yang B.W., Yeo I.-C., Hahm Y.T. Antimicrobial peptides of the genus Bacillus: A new era for antibiotics. Can. J. Microbiol. 2014;61:93–103. doi: 10.1139/cjm-2014-0613. [DOI] [PubMed] [Google Scholar]
- 71.Berrue F., Ibrahim A., Boland P., Kerr R.G. Newly isolated marine Bacillus pumilus (SP21): A source of novel lipoamides and other antimicrobial agents. Pure Appl. Chem. 2009;81:1027–1031. doi: 10.1351/PAC-CON-08-09-25. [DOI] [Google Scholar]
- 72.Ouoba L.I.I., Diawara B., Jespersen L., Jakobsen M. Antimicrobial activity of Bacillus subtilis and Bacillus pumilus during the fermentation of African locust bean (Parkia biglobosa) for Soumbala production. J. Appl. Microbiol. 2007;102:963–970. doi: 10.1111/j.1365-2672.2006.03156.x. [DOI] [PubMed] [Google Scholar]
- 73.Clark G.J., Langley D., Bushell M.E. Oxygen limitation can induce microbial secondary metabolite formation: Investigations with miniature electrodes in shaker and bioreactor culture. Microbiology. 1995;141:663–669. doi: 10.1099/13500872-141-3-663. [DOI] [Google Scholar]
- 74.Jami M., Ghanbari M., Kneifel W., Domig K.J. Phylogenetic diversity and biological activity of culturable Actinobacteria isolated from freshwater fish gut microbiota. Microbiol. Res. 2015;175:6–15. doi: 10.1016/j.micres.2015.01.009. [DOI] [PubMed] [Google Scholar]
- 75.Yoon J.H., Kim I.G., Kang K.H., Oh T.K., Park Y.H. Bacillus marisflavi sp. nov. and Bacillus aquimaris sp. nov., isolated from sea water of a tidal flat of the Yellow Sea in Korea. Int. J. Syst. Evol. Microbiol. 2003;53:1297–1303. doi: 10.1099/ijs.0.02365-0. [DOI] [PubMed] [Google Scholar]
- 76.Yoon J.H., Yeo S.H., Oh T.K., Park Y.H. Psychrobacter alimentarius sp. nov., isolated from squid jeotgal, a traditional Korean fermented seafood. Int. J. Syst. Evol. Microbiol. 2005;55:171. doi: 10.1099/ijs.0.63140-0. [DOI] [PubMed] [Google Scholar]
- 77.Niu L., Xiong M., Tang T., Song L., Hu X., Zhao M., Zhang K. Aeromicrobium camelliae sp. nov., isolated from Pu’er tea. Int. J. Syst. Evol. Microbiol. 2015;65:4369–4373. doi: 10.1099/ijsem.0.000583. [DOI] [PubMed] [Google Scholar]
- 78.Yan Z.-F., Lin P., Chu X., Kook M.C., Li C.-T., Yi T.-H. Aeromicrobium halotolerans sp. nov., isolated from desert soil sample. Arch. Microbiol. 2016;198:423–427. doi: 10.1007/s00203-016-1196-6. [DOI] [PubMed] [Google Scholar]
- 79.Magarvey N.A., Ehling-Schulz M., Walsh C.T. Characterization of the cereulide NRPS α-hydroxy acid specifying modules: Activation of α-keto Acids and chiral reduction on the assembly line. J. Am. Chem. Soc. 2006;128:10698–10699. doi: 10.1021/ja0640187. [DOI] [PubMed] [Google Scholar]
- 80.Huang T., Li L., Brock N.L., Deng Z., Lin S. Functional characterization of PyrG, an unusual nonribosomal peptide synthetase module from the pyridomycin biosynthetic pathway. Chem. Biol. Chem. 2016 doi: 10.1002/cbic.201600156. [DOI] [PubMed] [Google Scholar]
- 81.Heravi K.M., Shali A., Naghibzadeh N., Ahmadian G. Characterization of cis-acting elements residing in the chitinase promoter of Bacillus pumilus SG2. World J. Microbiol. Biotechnol. 2014;30:1491–1499. doi: 10.1007/s11274-013-1569-9. [DOI] [PubMed] [Google Scholar]
- 82.Ozcengiz G., Alaeddinoglu N.G., Demain A.L. Regulation of biosynthesis of bacilysin by Bacillus subtilis. J. Ind. Microbiol. 1990;6:91–100. doi: 10.1007/BF01576428. [DOI] [PubMed] [Google Scholar]
- 83.Molohon K.J., Melby J.O., Lee J., Evans B.S., Dunbar K.L., Bumpus S.B., Kelleher N.L., Mitchell D.A. Structure determination and interception of biosynthetic intermediates for the plantazolicin class of highly discriminating antibiotics. ACS Chem. Biol. 2011;6:1307–1313. doi: 10.1021/cb200339d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Lee D.W., Lee S.D. Aeromicrobium ponti sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2008 doi: 10.1099/ijs.0.65575-0. [DOI] [PubMed] [Google Scholar]
- 85.Ramasamy D., Kokcha S., Lagier J.-C., Nguyen T.-T., Raoult D., Fournier P.-E. Genome sequence and description of Aeromicrobium massiliense sp. nov. Stand. Genom. Sci. 2012;7:246–257. doi: 10.4056/sigs.3306717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Lightfield J., Fram N.R., Ely B. Across bacterial phyla, distantly-related genomes with similar genomic GC content have similar patterns of amino acid usage. PLoS ONE. 2011;6:e17677. doi: 10.1371/journal.pone.0017677. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Tu Q., Lin L. Gene content dissimilarity for subclassification of highly similar microbial strains. BMC Genom. 2016;17:647. doi: 10.1186/s12864-016-2991-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.