

Abstract
Background
Recent researches have been focusing on mucosal immune adjuvants, which play the key roles in mucosal immunization and have become the limitation for non-injected vaccine development. Escherichia coli heat-labile enterotoxin B subunit (LTB) was regarded as a promising mucosal adjuvant for its nontoxicity and potent activity. LTB preparation issues have always been recurring, in part owing to that the recombinant LTB expressed by E. coli does not act as its native form.
Results
We constructed an engineered Lactococcus lactis strain using a food-grade expression system. The LTB secreted by the engineered strain was detected in the culture supernatant, constituting 10.3% of the supernatant proteins, and recognized by mouse anti-LTB antibodies. The engineered strain, co-administered orally to SPF BALB/c mice with a H. pylori vaccine candidate expressing Lpp20 antigen, could significantly enhance the Lpp20-induced mucosal SIgA antibody responses against H. pylori.
Conclusions
This is the first report that LTB was efficiently produced and delivered via using a food-grade lactococcal expression system, which offers a novel production and utilization mode of this crucial mucosal adjuvant. The engineered L. lactis strain secreting LTB has considerable potential for oral vaccine formulation owing to its outstanding safety, adjuvant activity and high-level production.
Electronic supplementary material
The online version of this article (doi:10.1186/s12896-017-0345-6) contains supplementary material, which is available to authorized users.
Keywords: Helicobactor pylori, Lactococcus lactis, Lpp20, Food-grade, Oral vaccine, LTB
Highlights
This is the first report that LTB was efficiently produced and delivered via using a food-grade lactococcal expression system.
This study offers a novel production and utilization mode of this crucial mucosal adjuvant.
The engineered L. lactis strain secreting LTB has considerable potential for oral vaccine formulation owing to its outstanding safety, adjuvant activity and high-level production.
Background
Gastroenteric infections cause an estimated two million deaths worldwide per year, and remain severe public health issues [1, 2]. As antibiotic resistance has been continually increasing, researches currently focus on developing vaccines against the causative agents, such as Helicobacter pylori, Shigella and enterotoxigenic E. coli (ETEC), for which no commercial vaccines are available [1].
Presently, most commercial vaccines are administered by parenteral routes [3]. However, recent studies demonstrated mucosal vaccination as the most effective strategy against the pathogens that colonize or invade mucosa to initiate lesions [3–5]. Although parenteral immunizations can protect against causative agents parasitizing host tissues via stimulation of serum antibody and cellular immune responses, they can hardly elicit mucosal immunity against noninvasive pathogens [3, 6]. Mucosal vaccination can stimulate secretory antibody responses preventing infection by the pathogens from the mucosal surface [5]. Additionally, mucosal immunizations have the advantages of simple manipulation, less invasion, lowered risks of disease transmissions and ease of manufacture over parenteral inoculations.
However, mucosal vaccinations with antigens alone are commonly insufficient to induce marked immune responses, unless the antigens can reach the mucosal inductive sites as cholera toxins [3, 7]. As proved, mucosal adjuvants or microbial delivery vectors are required for effective mucosal immune responses [8]. Therefore, recent researches have emphasized screening and preparation of adjuvants and the biotic delivery vehicles which possess adjuvant activity [3].
E. coli heat-labile enterotoxin B subunit (LTB) is a promising mucosal adjuvant, owing to its nontoxicity and potent mucosal adjuvant activity [9]. Nevertheless, LTB preparation issues have always been existing because it is impractical to purify LTB from ETEC for production of vaccines, and the activity of recombinant LTB (rLTB) was greatly affected by the expression hosts employed. Previous studies have indicated that preparation of LTB by using a E. coli expression system are not only inefficient but also costly [6]. The reasons involve the recurring formation of insoluble inclusion bodies, lower yields of bioactive rLTB, the cost of protein purification and the risk of pollution with unbeneficial bacterial components like lipopolysaccharide.
To address these issues, such bacteria as attenuated pathogens and probiotics have been exploited as expression hosts and live vectors for LTB production and delivery. A study compared expression efficacy of rLTB in Pichia pastoris with that in E. coli, demonstrating that a higher expression level and adjuvant activity of rLTB could be obtained by using P. pastoris as the expression host [6]. Another study proved that LTB expressed in fusion with antigens in engineered Lactobacillus can significantly enhance the local and systemic immune responses to the antigens [10, 11]. Recently, increasing evidences supported that food-grade L. lactis expression systems, through expression and delivery of antigens/adjuvants, are promising oral vaccine vectors, particularly owing to their outstanding safety, avoidance of protein purification, reduced antigen degradation and efficient delivery of immunogens to the mucosal inductive sites [12, 13]. However, successful expression of LTB in food-grade L. lactis has not been reported to date. Therefore, construction of a food-grade L. lactis strain producing LTB should be a considerable step toward the goal of effective and safe mucosal vaccines.
In this work, a food-grade L. lactis strain was genetically engineered for production and delivery of LTB, and its immune adjuvant activity was evaluated by oral vaccination of mice with the engineered strain and a Lpp20-based H. pylori vaccine candidate. The observations of this study demonstrate a novel efficient production and utilization mode of LTB, which forms a crucial basis for mucosal vaccine formulation.
Methods
Bacterial strains, plasmids and growth conditions
The bacteria and plasmids are shown in Additional file 1. The bacterial cultivation conditions were as described previously [12, 14]. SPF BALB/c mice, aged 6 weeks, were purchased from Henan Experimental Animal Center (Zhengzhou, China).
Polymerase chain reaction of ltB gene
The ltB gene was amplified by PCR from the plasmid pMAL-c2x-mlt63 using Pyrobest DNA polymerase (TaKaRa, China). A pair of oligonucleotide primers was designed based on the published sequence (GenBank EF057802). The sequences of the primers were 5’-CAGTCGGCATGCGCTCCCCAGTCTATTAC-3’ (Sense) and 5’-CGCTCTAGACTAGTTTTCCATACTGATTG-3’ (Antisense) with the endonuclease sites SphI and XbaI shown in bold letters, respectively. The PCR profile included 30 cycles of 94 °C for 1 min, 55 °C for 30 s and 72 °C for 2 min.
Construction of recombinant L. lactis strains
The ltB gene was ligated with TA cloning vector pMD19-T and used to transform E. coli DH5α by heat shock. The ltB gene fragment was collected by digestion of pMD19-T-ltB using SphI and XbaI (TaKaRa, China), ligated with plasmid pNZ8149-SP (GenBank KY385376), and used for transformation of L. lactis NZ3900 by electrophoration [12]. The recombinants were obtained by Elliker medium selection, and identified by restriction digestion and gene sequencing as reported before [12]. The recombinant NZ3900 strain carrying ltB gene was referred to as L. lactis NZ3900/pNZ8149-SP-ltB.
LTB expression and western blotting assays
The expression of LTB in NZ3900/pNZ8149-SP-ltB was induced using 25 ng/ml nisin (Sigma, USA) as inducer under the conditions as previously described [12].
Samples of the culture supernatant were prepared from 50 ml of culture. The supernatant was obtained by centrifugation at 10,000 rpm for 20 min at 4 °C, and filtered through 0.22 μm filter. The proteins in the filtrate were precipitated by adding trichloroacetic acid (10%, v/v), incubating at 4 °C for 16 h and centrifugation at 10,000 rpm for 30 min at 4 °C. The pellet was resuspended in 8 ml acetone, centrifugated at 10,000 rpm for 20 min at 4 °C, then kept in fume hood at room temperature until dry. The protein sample was added 360 μl PBS, kept at 4 °C for 3 h, and centrifugated at 10,000 rpm 4 °C for 10 min. The supernatant was collected and used as samples of the culture supernatant. Samples of bacterial cell lysates were processed as described before [12].
SDS-PAGE and western blotting assays were performed using mouse anti-LTB antibody (Abcam, USA) as the primary antibody as previously reported [12].
Oral vaccination of mice
The mice were assigned at random into three groups of 10 each. For Lpp20 group and Lpp20 + LTB group, the mice were treated by gavage with cell suspensions at a dose of 300 μl of NZ3900/pNZ8149-SP-lpp20 (1 × 1011 CFU/ml) and a mixture of NZ3900/pNZ8149-SP-lpp20 (1 × 1011 CFU/ml) and NZ3900/pNZ8149-SP-ltB (1 × 1011 CFU/ml), respectively, on day 0, 7, 14, 21, 28 and 35. For PBS group, mice were given equal volumes of PBS instead of the cell suspensions.
Blood and intestinal feces sampling
Seven days after the last vaccination, blood and intestinal feces were collected for half number of the mice from all the groups. The blood samples were fetched from orbital sinus and kept at 4 °C for 16–20 h, and then the sera were separated and stored in aliquots at −20 °C. For sampling intestinal feces, the mice were sacrificed by spinal dislocation, 100 mg of feces was fetched from the intestine for each mouse, and then 1 ml of PBS containing proteinase inhabitor (Phenylmethanesulfonyl fluoride, 0.1 mM) was injected by the duodenum to wash the intestinal wall. The eluate was recovered, mixed with the feces and kept at 4 °C for overnight (14–16 h). The supernatant of the mixture was separated via centrifugation at 12,000 rpm for 10 min, and stored at −20 °C as ELISA samples.
ELISA detection of IgG and SIgA antibodies
The H. pylori-specific serum IgG and fecal SIgA antibodies were quantified by ELISA as described before [12]. Briefly, 96-well microplates (Beijing Solarbio, China) were coated with soluble H. pylori somatic proteins. The ELISA signals were developed using biotinylated goat anti-mouse IgG (Abcam, USA), goat anti-mouse SIgA (Abcam, USA) and p-nitrophenyl phosphate (PNPP) substrate (Beijing Solarbio, China). The absorbances of the wells at 450 nm (OD450) were measured using a Microplate Reader (Tecan Sunrise, CH), and designed as indicators of the specific sIgA and IgG levels.
Statistical analysis
The measurement data were presented as means ± standard deviation The significance of the difference among the groups was tested using Kruskal-Wallis tests, while the pairwise comparisons of mean values were carried out using the Mann-Whitney U test with the aid of software SAS9.13. The difference was considered as significant at P <0.05.
Results
Genetic engineering of L. lactis
The PCR product of ltB gene was 334 bp in length, and 100% identical in nucleotide sequence to the published E. coli ltB gene (GenBank JX504011.1). The plasmids were isolated from the L. lactis transformants, and identified by restriction enzyme digestion. Gene sequencing confirmed that the ltB gene was exactly cloned downstream of the nisin controlled promoter (Pnis) within the expression vector pNZ8149-SP, generating pNZ8149-SP-ltB. The schematic diagram of pNZ8149-SP-ltB was shown in Fig. 1.
Fig. 1.
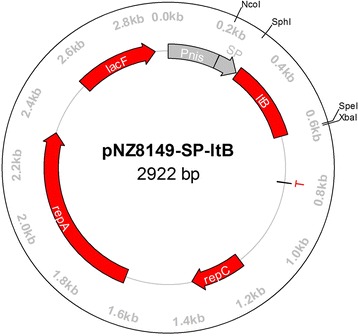
The schematic maps of pNZ8149-SP-ltB designed using software Omiga 2.0. Pnis, nisA promoter; SP, signal sequence of usp45 gene; ltB, ltB gene coding region; T, terminator; repA and repC, replication gene A and C; lacF, lacF gene as selection signature
Expression and immunoreactivity of Lpp20
To determine inducible expression of LTB in L. lactis, the cell lysates and culture supernatant were analyzed by SDS-PAGE and western blots. As results, SDS-PAGE analysis showed that a dense protein band was present at approximately 13 kDa position in the culture supernatant of NZ3900/pNZ8149-SP-ltB, while absent at the corresponding position in the controls (Fig. 2a). The percentage of LTB reached 10.3% in the culture supernatant proteins of the engineered stain.
Fig. 2.
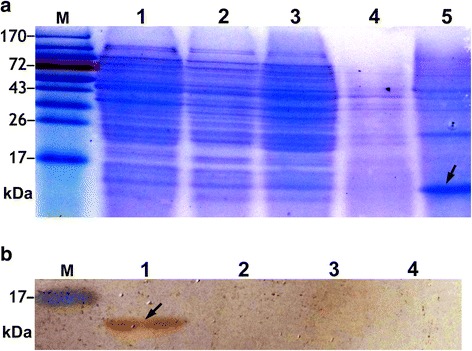
Analysis of cell lysate and culture supernatant proteins of the nisin-induced L. lactis strains. The arrows indicate the expressed UreB. a SDS-PAGE. Lane M, protein markers; lane 1, 2, 3, cell lysates of NZ3900, NZ3900/pNZ8149-SP and NZ3900/pNZ8149-SP-ltB, respectively; lane 4, 5, culture supernatant of NZ3900/pNZ8149-SP and NZ3900/pNZ8149-SP-ltB, respectively. These figures showed that the recombinant LTB was expressed as a secreted protein. b Western blotting assays. L. lactis NZ3900/pNZ8149-SP was used as the negative control, while mouse anti-LTB antibodies as the primary antibodies for detection of LTB. Lane M, protein markers; lane 1, 2, culture supernatant of NZ3900/pNZ8149-SP-ltB and NZ3900/pNZ8149-SP, respectively; lane 3, 4, cell lysates of NZ3900/pNZ8149-SP-ltB and NZ3900/pNZ8149-SP, respectively. These figures showed that the expressed LTB was detectable only in the culture supernatant of NZ3900/pNZ8149-SP-ltB, and possessed immunoreactivity with the commercial anti-LTB antibody
Western blotting analysis showed that the 13 kDa protein in the culture supernatant of NZ3900/pNZ8149-SP-ltB could be recognized by the mouse anti-LTB antibodies, while there was no positive band at the corresponding position in the lanes of NZ3900/pNZ8149-SP samples as negative controls (Fig. 2b).
Immunization of mice and antibody assays
The L. lactis NZ3900/pNZ8149-SP-lpp20 and NZ3900/pNZ8149-SP-ltB were induced with nisin as mentioned above. The mice of the immunized groups were orally inoculated with the induced NZ3900/pNZ8149-SP-lpp20 alone and its mixture with NZ3900/pNZ8149-SP-ltB, respectively, while the control group was treated with PBS. On day 7 after the final vaccination, the serum and intestinal feces were sampled, and tested for H. pylori-specific antibodies by ELISA assays. The results are shown in Additional file 2 and Fig. 3. The group treated with NZ3900/pNZ8149-SP-lpp20 alone had enhanced anti-H. pylori serum antibody responses, but no significantly elevated intestinal SIgA level, compared with the PBS group. The vaccination with the mixture of the two L. lactis strains evoked significantly elevated SIgA antibody level, and insignificantly lowered the serum IgG response, as compared with the group administered with NZ3900/pNZ8149-SP-lpp20 alone. The results demonstrated the adjuvant activity of the engineered L. lactis expressing LTB to promote mucosal immune responses.
Fig. 3.
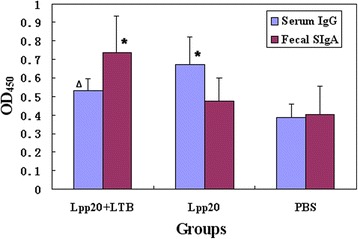
OD450 values of ELISA tests for H. pylori-specific serum IgG and intestinal SIgA levels. The Lpp20 group and Lpp20 + LTB group were vaccinated with L. lactis NZ3900/pNZ8149-SP-lpp20 and the mixture of NZ3900/pNZ8149-SP-lpp20 and NZ3900/pNZ8149-SP-ltB, respectively, while the PBS group, as the control, was treated with PBS. One week after oral immunization, blood sampling was performed for half number of the mice of all the groups. The serum (1:10 diluted) and fecal samples were tested for H. pylori-specific antibodies by ELISA. Bars, mean OD450 value; error bars, standard deviation; ∆ the value was significantly higher than that for the PBS group (P < 0.05); * the value was significantly different from those for the other two groups (P < 0.05)
Discussion
As proved, the species of bacteria as expression hosts might greatly affect the bioactivity of recombinant expression products [6]. Through screening of various microbial expression systems, it is possible to establish an efficient method for production and utilization of LTB. Nevertheless, although studies have demonstrated the potential of LTB as mucosal adjuvant and accumulated evidences proved L. lactis to be a safe vaccine vehicle, expression and delivery of LTB using a food-grade L. lactis strain have not been reported yet. Therefore, successfully engineering a food-grade L. lactis strain to secret LTB can be a crucial basis for mucosal vaccine development.
L. lactis NZ3900/pNZ8149-SP is a food-grade nisin-controlled expression system, which was constructed in our previous study by introducing a gene fragment encoding the signal peptide of L. lactis Usp45 protein into pNZ8149 for capacity of secretory expression of heterologous genes [12]. In this work, the ltB gene was inserted downstream of the nisin controlled promoter within pNZ8149-SP, and expression of LTB was achieved by inducement with food-grade subtoxic amount of nisin. All the components of L. lactis NZ3900/pNZ8149-SP-ltB, except for the target gene ltB, possess food-grade safety, suggesting the extremely high safety of the engineered strain as the LTB producer.
As reported, the L. lactis expression efficiency of heterologous protein can be rather low, and thus the expression products in certain studies were detectable by western blots, but not by SDS-PAGE [15, 16]. In this work, the expressed LTB was visual as the predominant band in the SDS-PAGE pattern, constituting 10.3% of the culture supernatant extracts of the engineered L. lactis. The expression efficacy was much higher than those reported [12, 15–17]. The mechanism underlying this phenomenon might include the diversity of the heterologous genes in genetic codon constitution. The LTB expressed in L. lactis had a molecular weight of approximately 13 kDa, corresponding to that deduced from the nucleotide sequence (GenBank JX504011.1). Western blotting assays showed that LTB produced by the engineered L. lactis strain retained potent antigenicity. These findings indicate that the engineered strain culture can be a novel source of LTB, from which LTB can be obtained and utilized without protein purification and the risk of pollution with toxic components from the commonly used expression hosts, such as E. coli and Salmonella spp..
A recurring issue on heterogeneous expression is that the recombinant products might lose bioactivity of the native proteins. Although the molecular basis of LTB adjuvant properties remains unclear, it might be included that LTB interacts with such receptors as GM1-ganglioside accelerating uptake of the toxic subunit A into the epithelial cells [6, 18]. In the present study, it was observed that vaccination of mice with L. lactis expressing Lpp20 was capable of arousing remarkable serum IgG antibody responses, but failed to induce significantly elevated intestinal specific SIgA antibody levels, as compared with the group given PBS, indicating that without the aid of suitable mucosal adjuvants, the employed L. lactis strain producing Lpp20 can hardly evoke notable mucosal immune responses. Similarly, a previous study showed that a L. lactis strain producing H. pylori UreB antigen was unable to elicit marked immune responses in mice [17]. The adjuvant activity of the engineered strain secreting LTB has been proved by the observations that the group received the mixture of the two engineered L. lactis strains expressing Lpp20 and LTB, respectively, had significantly elevated sIgA antibodies, in comparison with the group given the Lpp20-producing strain alone. Since mucosal SIgA antibodies have the key roles in immune protection against the pathogens invading human tissues by the mucosal surface, the findings of our study suggest that the engineered strain secreting LTB might markedly enhance the immune protective efficacy of the co-administered antigens [19].
Interestingly, accompanying with the enhanced mucosal SIgA, the serum IgG antibodies seemed to be suppressed in the mice immunized with the mixture of the two L. lactis strains. This result suggests that the LTB-secreting L. lactis might modulate the systemic and mucosal immune responses with opposite effects. The mechanism and its implication for vaccine designs need further investigation.
In recent decades, although in most countries the use of genetically modified organisms (GMOs) was prohibited or heavily regulated, it has been proposed to change the present regulations [20, 21]. In fact, the amount of commercially available GMOs has been continuously rising in recent years [22]. It is hopeful for the food-grade L. lactis engineered here to be used in pharmacy and food industry in future.
Conclusion
Taken together, this is the first report that E. coli LTB was efficiently produced and delivered via using a food-grade lactococcal expression system, which offers a novel production and utilization mode of this crucial mucosal adjuvant. The engineered L. lactis strain secreting LTB has considerable potential for oral vaccine formulation, owing to its outstanding safety, adjuvant activity and high-level production.
Acknowledgments
The authors are grateful to the funding suppliers mentioned above.
Funding
This study was funded by China Postdoctoral Science Foundation (200801273) and Henan Innovation Center of Molecular Diagnosis and Laboratory Medicine (XTCX-2015-ZD2).
Availability of data and materials
All data generated during this study are included in this published article and its Additional files 1 and 2.
Authors’ contributions
NS engineered the L. lactis strain, and performed the Western blot analysis and animal experimentation. RZ and GD took the responsibility of designing and executing the project, and writing the manuscript. RZ, XP, CW and SC performed the immunization of mice and evaluation of immune efficacy. QF and YX contributed to the animal experiments. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval
The animal experimentations were approved by the Institutional Review Board at Zhengzhou University (no reference number), and performed complying with the ARRIVE guidelines. This study does not involve human participants.
Abbreviations
- ELISA
Enzyme-linked immunosorbent assay
- ETEC
Enterotoxigenic E. coli
- LTB
Heat-labile enterotoxin B subunit
- PBS
Phosphate buffer saline
- PNPP
P-nitrophenyl phosphate
- SPF
Specific pathogens free
- UreB
Urease B subunit
Additional files
Bacteria and plasmids used in this work. (DOC 39 kb)
OD450 values of ELISA tests for H. pylori-specific IgG and SIgA levels. (DOC 60 kb)
Contributor Information
Rongguang Zhang, Email: zrg@zzu.edu.cn.
Guangcai Duan, Phone: +86 135 25 583039, Email: gcduan@zzu.edu.cn.
References
- 1.Daryani A, Hosseini-Teshnizi S, Hosseini SA, Ahmadpour E, Sarvi S, Amouei A, et al. Intestinal parasitic infections in Iranian preschool and school children: A systematic review and meta-analysis. Acta Trop. 2017;169:69–83. doi: 10.1016/j.actatropica.2017.01.019. [DOI] [PubMed] [Google Scholar]
- 2.Black RE, Cousens S, Johnson HL, Lawn JE, Rudan I, Bassani DG, et al. Global, regional, and national causes of child mortality in 2008: a systematic analysis. Lancet. 2010;375:1969–87. doi: 10.1016/S0140-6736(10)60549-1. [DOI] [PubMed] [Google Scholar]
- 3.Pérez O, Batista-Duharte A, González E, Zayas C, Balboa J, Cuello M, et al. Human prophylactic vaccine adjuvants and their determinant role in new vaccine formulations. Braz J Med Biol Res. 2012;45:681–92. doi: 10.1590/S0100-879X2012007500067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Holmgren J, Bourgeois L, Carlin N, Clements J, Gustafsson B, Lundgren A, et al. Development and preclinical evaluation of safety and immunogenicity of an oral ETEC vaccine containing inactivated E. coli bacteria overexpressing colonization factors CFA/I, CS3, CS5 and CS6 combined with a hybrid LT/CT B subunit antigen, administered alone and together with dmLT adjuvant. Vaccine. 2013;31:2457–64. doi: 10.1016/j.vaccine.2013.03.027. [DOI] [PubMed] [Google Scholar]
- 5.Lundgren A, Jertborn M, Svennerholm AM. Induction of long term mucosal immunological memory in humans by an oral inactivated multivalent enterotoxigenic Escherichia coli vaccine. Vaccine. 2016;34:3132–40. doi: 10.1016/j.vaccine.2016.04.055. [DOI] [PubMed] [Google Scholar]
- 6.Ma X, Yao B, Zheng W, Li L. Comparative study on characterization of recombinant B subunit of E. coli heat-labile enterotoxin (rLTB) prepared from E. coli and P. patoris. J Microbiol Biotechnol. 2010;20:550–7. [PubMed] [Google Scholar]
- 7.Norton EB, Branco LM, Clements JD. Evaluating the A-subunit of the heat-labile toxin (LT) as an immunogen and a protective antigen against enterotoxigenic Escherichia coli (ETEC) PLoS One. 2015;10:e0136302. doi: 10.1371/journal.pone.0136302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kataoka K, Fujihashi K. Dendritic cell-targeting DNA-based mucosal adjuvants for the development of mucosal vaccines. Expert Rev Vaccines. 2009;8:1183–93. doi: 10.1586/erv.09.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Jawale CV, Lee JH. Salmonella enterica serovar enteritidis ghosts carrying the Escherichia coli heat-labile enterotoxin B subunit are capable of inducing enhanced protective immune responses. Clin Vaccine Immunol. 2014;21:799–807. doi: 10.1128/CVI.00016-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yu Q, Zhu L, Kang H, Yang Q. Mucosal Lactobacillus vectored vaccines. Hum Vaccin Immunother. 2013;9:805–7. doi: 10.4161/hv.23302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Yu M, Qi R, Chen C, Yin J, Ma S, Shi W, et al. Immunogenicity of recombinant Lactobacillus casei-expressing F4 (K88) fimbrial adhesin FaeG in conjunction with a heat-labile enterotoxin A (LTAK63) and heat-labile enterotoxin B (LTB) of enterotoxigenic Escherichia coli as an oral adjuvant in mice. J Appl Microbiol. 2017;122:506–515. doi: 10.1111/jam.13352. [DOI] [PubMed] [Google Scholar]
- 12.Chen SY, Zhang RG, Duan GC, Shi JX. Food-grade expression of Helicobacter pylori ureB subunit in Lactococcus lactis and its immunoreactivity. Curr Microbiol. 2011;62:1726–31. doi: 10.1007/s00284-011-9920-6. [DOI] [PubMed] [Google Scholar]
- 13.Wyszyńska A, Kobierecka P, Bardowski J, Jagusztyn-Krynicka EK. Lactic acid bacteria--20 years exploring their potential as live vectors for mucosal vaccination. Appl Microbiol Biotechnol. 2015;99:2967–77. doi: 10.1007/s00253-015-6498-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ma YJ, Duan GC, Zhang RG, Fan QT, Zhang WD. Mutation of iceA in Helicobacter pylori compromised IL-8 induction from human gastric epithelial cells. J Basic Microbiol. 2010;50:S83–8. doi: 10.1002/jobm.200900410. [DOI] [PubMed] [Google Scholar]
- 15.Zhang XJ, Feng SY, Li ZT, Feng YM. Expression of Helicobacter pylori hspA gene in Lactococcus lactis NICE system and experimental study on its immunoreactivity. Gastroenterol Res Pract. 2015;2015:750932. doi: 10.1155/2015/750932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Gu Q, Song D, Zhu M. Oral vaccination of mice against Helicobacter pylori with recombinant Lactococcus lactis expressing urease subunit B. FEMS Immunol Med Microbiol. 2009;56:197–203. doi: 10.1111/j.1574-695X.2009.00566.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Lee MH, Roussel Y, Wilks M, Tabaqchali S. Expression of Helicobacter pylori urease subunit B gene in Lactococcus lactis MG1363 and its use as a vaccine delivery system against H. pylori infection in mice. Vaccine. 2001;19:3927–35. doi: 10.1016/S0264-410X(01)00119-0. [DOI] [PubMed] [Google Scholar]
- 18.Wang B, Pan X, Wang H, Zhou Y, Zhu J, Yang J, et al. Immunological response of recombinant H. pylori multi-epitope vaccine with different vaccination strategies. Int J Clin Exp Pathol. 2014;7:6559–66. [PMC free article] [PubMed] [Google Scholar]
- 19.Wang S, Liu H, Zhang X, Qian F. Intranasal and oral vaccination with protein-based antigens: advantages, challenges and formulation strategies. Protein Cell. 2015;6:480–503. doi: 10.1007/s13238-015-0164-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.McHughen A. A critical assessment of regulatory triggers for products of biotechnology: Product vs. process. GM Crops Food. 2016;7:125–58. doi: 10.1080/21645698.2016.1228516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Korobko IV, Georgiev PG, Skryabin KG, Kirpichnikov MP. GMOs in Russia: research, society and legislation. Acta Naturae. 2016;8:6–13. [PMC free article] [PubMed] [Google Scholar]
- 22.Peng C, Wang P, Xu X, Wang X, Wei W, Chen X, et al. Development of a qualitative real-time PCR method to detect 19 targets for identification of genetically modified organisms. Springerplus. 2016;5:889. doi: 10.1186/s40064-016-2395-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated during this study are included in this published article and its Additional files 1 and 2.