

Abstract
MicroRNAs are small noncoding RNAs that work posttranscriptionally to negatively regulate protein levels. They influence neuronal and glial structure and function, neuroinflammatory signaling, cell death, neurogenesis, and other processes relevant to epileptogenesis. Functional studies using oligonucleotide inhibitors (antagomirs) and mimics (agomirs) to modulate microRNAs in rat and mouse models of epilepsy show effects on evoked and spontaneous seizures and attendant neuropathology. The present review summarizes recent findings and points to gaps in our knowledge of the underlying mechanisms and directions for the future.
Commentary
There has been substantial progress in understanding the cellular and molecular mechanisms of epileptogenesis, and this has led to new ideas about how to develop novel disease-modifying treatments (1–3). A compelling idea is to find control points—nodes—that coordinate the expression of one or more networks of genes (4, 5). Targeting these could bring about larger-scale interference in the pathways that drive the development or maintenance of the epileptic state. MicroRNAs offer potential opportunities in this regard. MicroRNAs are small noncoding RNAs that control gene expression at a post-transcriptional level, promoting either degradation of their target mRNAs or translational repression (6). MicroRNAs exert network-level effects on gene expression because each can target multiple transcripts that encode proteins in the same pathway or single targets in multiple pathways. Their effects are particularly important for reducing “noise” and conferring precision to cellular protein levels, particularly for low-abundance targets (7, 8). Levels of numerous microRNAs have been found to be dysregulated in experimental and human epilepsy, and modulation of their biogenesis or targeting of individual microRNAs can produce potent effects on evoked and spontaneous seizures in animals (9). The present review aims to capture the pace of discovery by summarizing recent in vivo studies that manipulated microRNAs in rat and mouse models of epilepsy. The review also points to gaps in our knowledge and suggestions for future directions.
On the Mechanics of MicroRNAs
MicroRNAs function at a posttranscriptional level to fine-tune protein levels in cells. They do this by binding to short complementary sequences in mRNAs, often within the 3′ untranslated region of the transcript. This has the effect of promoting degradation or inhibiting translational efficiency, leading to overall lower protein levels in cells (6). MicroRNAs originate from inter-and intragenic sites including introns of protein-coding genes and their own specific gene loci. Transcription generates a primary transcript (pri-microRNA). which goes through a nuclear RNase-mediated processing event mediated by the Drosha microprocessor followed by cytoplasmic cleavage of the pre-microRNA by Dicer, resulting in the mature, double-stranded microRNA (~22 nucleotides) (10, 11). One strand of the mature microRNA is selected by the protein Argonaute-2, forming the so-called RNA-induced silencing complex (RISC), which then begins to search mRNAs for a region of sufficient complementarity upon which stable base-pair binding occurs (12). It requires only a 7 to 8 nucleotide “seed” region match for microRNA to mRNA targeting effects to occur; this is why microRNAs have so many potential targets and is the source of their multi-targeting property. After stable binding, other proteins may be recruited to the RISC, which promotes degradation or translational inhibition (13). MicroRNAs are currently named using a number-based system in order of their discovery (e.g., miR-1, miR-2). The number of microRNAs encoded in the human or rodent genome likely ranges from ~500 to more than 2,000 (14). Figure 1A provides an overview of microRNA biogenesis and function.
FIGURE 1.
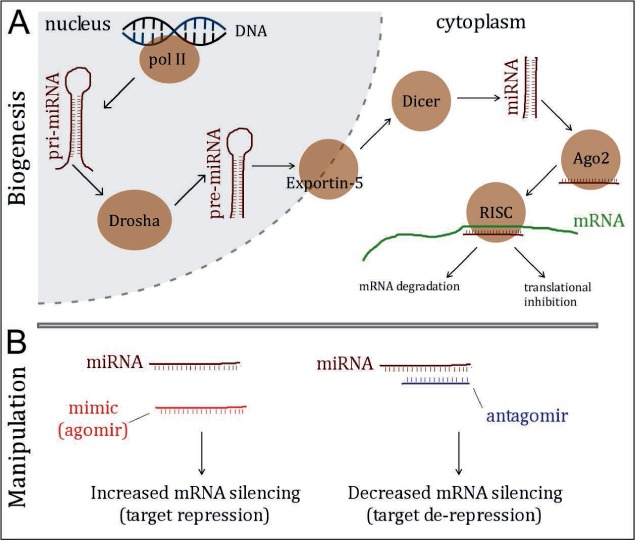
Overview of microRNA biogenesis and approaches used to manipulate microRNAs in recent functional studies in epilepsy. (A) Simplified microRNA (miRNA) biogenesis pathway. Polymerase II (pol II) transcribes a pri-miRNA transcript, which is then shortened by the actions of the Drosha microprocessor complex to form a pre-miRNA. This is then exported from the nucleus to the cytoplasm and acted on by another RNase complex containing Dicer, resulting in the mature double-stranded microRNA. One strand is selected by Argonaute-2 (Ago2) forming the RNA-induced silencing complex (RISC), which is then guided to mRNA targets with complementary pairing, often within the 3′ untranslated region of the mRNA. Stable binding results in destabilization and degradation of the mRNA or translational inhibition. (B) Overview of the common techniques used to manipulate miRNAs in in vivo models of epilepsy. Overexpressing microRNA involves introducing a mimic (agomir), whereas introducing an oligonucleotide (antagomir) with complementary base-pairing to the microRNA blocks it from functioning.
MicroRNAs Are Important for Brain Function
Studies in genetically modified mice lacking key biogenesis genes have revealed that microRNAs are essential for normal organism development, including brain maturation (15). More recent work in conditional models has determined that loss of microRNA biogenesis components from the mature brain results in progressive tissue dysmorphogenesis, neurodegeneration, and seizures (16–18). The role of a number of specific microRNAs in brain excitability is understood. MicroRNAs including miR-132 and miR-134 target transcripts encoding proteins crucial to growth and remodeling of dendrites, thereby regulating synaptic strength and plasticity (19–21). MicroRNAs miR-9 and miR-124 target mRNAs involved in neurogenesis and integration of newborn neurons into the circuitry of the mature brain (22, 23). MicroRNAs that control the regenerative capacity of axons during aging have also been identified in a model organism (24). There are also a number of glial-expressed microRNAs with potential relevance to epilepsy. For example, miR-155 regulates a network of genes coordinating microglial responses (25). Together, these studies reveal an important influence of microRNAs on brain function that spans most of the processes implicated in epileptogenesis, from controlling synaptic structure and function, glial morphology, inflammatory responses, and beyond. Thus, microRNAs may be relevant to the pathogenesis and treatment of epilepsy.
Targeting MicroRNAs in Models of Epilepsy
The first evidence that seizures could be altered by targeting an individual microRNA was reported for miR-134 (26). Intra-cerebroventricular injection of oligonucleotides designed to bind the microRNA, termed antagomirs (Figure 1B), resulted in suppression of evoked and spontaneous seizures in a mouse kainate model (26). Later, knockout of miR-128 in mice was shown to result in fatal epilepsy, whereas overexpression of miR-128 had acute seizure-suppressive effects (27). The hyperexcitability effect of reduced miR-128 was recently confirmed by an independent team using primary neurons transfected with a “sponge” construct to deplete miR-128 (28). Additional studies found that antagomirs targeting miR-34a (29) and miR-132 (30) could reduce seizure-induced neuronal death in models of status epilepticus. Inhibiting miR-132 after status epilepticus has also been reported to reduce the frequency of spontaneous seizures (31). The number of microRNAs for which functional data are available in in vivo models of epilepsy has been expanding rapidly. Recent examples are reviewed below.
New MicroRNAs in Epilepsy Models
The recent studies on microRNA manipulations in in vivo rat and mouse models have invariably used oligonucleotide-based approaches in two types of experiments: 1) by injecting an antagomir to bind and block the microRNA, or 2) by injecting a microRNA mimic (agomir) to artificially increase the microRNA level (Figure 1B). These may be delivered before inducing status epilepticus, looking at the effects on acute seizures or damage, or injected after status epilepticus, and then following the effects on the emergence or frequency of spontaneous seizures. The first type of study screens for potential acute seizure-suppressive effects that would work much like current anticonvulsive drugs. The second approach has the potential to identify disease-modifying (antiepileptogenic) effects of microRNAs.
MicroRNAs With Potential Seizure-Suppressive (Anticonvulsant) Effects
Four microRNAs have recently been shown to have potential direct or indirect seizure-suppressive effects. Intracerebroventricular injection of mimics of miR-22 (a microRNA identified as upregulated in nondamaged brain after status epilepticus) into mice during the first few days after status epilepticus, induced by intra-amygdala kainic acid, was shown to transiently reduce spontaneous seizures (32). Conversely, antagomirs targeting miR-22 caused more spontaneous seizures in the model and exacerbated astrogliosis within the hippocampus (32). A key target of this microRNA was shown to be the ATP-gated P2X7 receptor, which is known to promote release of interleukin-1β. Thus, miR-22 may function to suppress neuroinflammatory signaling in epilepsy.
Three other microRNAs with putative seizure-suppressive effects are miR-23b, miR-124, and miR-219. Levels of miR-23b were reported to be downregulated in a mouse kainate model, and delivery of miR-23b mimics into the mouse ventricle reduced post-kainate spiking seen the next day (33). The mechanisms and targets of miR-23b were not explored. Another microRNA downregulated in experimental epilepsy was miR-124. Pretreating rats with miR-124 mimics delayed seizure onset and resulted in less severe seizures in the pilocarpine model (34). Tests in the pentylenetetrazol model produced similar results (34). Potential mechanisms were suggested including targeting of glutamate-related signaling components and the cyclic-AMP response element binding protein (34). Lower levels of miR-219 were reported in experimental and human epilepsy and injection of an miR-219 mimic protected against kainate seizures in mice. Inhibiting miR-219 with antagomirs was, on its own, sufficient to produce epileptiform spiking in mice. The effects of miR-219 were suggested to be mediated by targeting of glutamatergic signaling components (35).
MicroRNAs With Potential Seizure-Promoting (Proconvulsive) Effects
Three new microRNAs have been identified, which appear to promote seizures or epilepsy. A recent study showed that delivery of antagomirs targeting microRNA-155 prior to status epilepticus induced by pilocarpine in mice resulted in a trend toward lower mortality and lower scores in a Racine-type behavioral assessment and suggested brain-derived neurotrophic factor as a possible target. The effects of the antagomir on post–status epilepticus epilepsy or neuropathology were not reported (36).
Levels of miR-199a were found to be increased following pilocarpine-induced status epilepticus in rats (37). Pretreating rats with an antagomir targeting miR-199a prior to status epilepticus reduced acute seizure severity and damage. The authors identified the mechanism of the antagomir effect as partly the result of de-repression of Sirtuin1, a transcriptional silencer (37).
In a profiling study using a mouse model of pilocarpine-induced status epilepticus, miR-203 was identified to be upregulated during the epileptic phase and increased in hippocampi from patients with epilepsy (38). Using intranasal delivery, the authors showed that antagomirs targeting miR-203 reduced the frequency of spontaneous seizures in epileptic mice over a two-week analysis period and identified the glycine receptor B as a potential target (38).
MicroRNAs With Effects Only on Seizure-Related Pathology
While Brennan and colleagues found no improvement in epilepsy in animals given miR-124 mimics, they observed that injection of the mimics alone evoked inflammatory responses. This may have obviated any potential seizure-suppressive effects of elevating miR-124 in neurons. The finding appears to conflict with the anticonvulsant effects reported for miR-124 mimics given 3 days prior to pilocarpine-induced seizures in another study (34). However, the differences may relate to whether pathology is present in the brain at the time of miR-124 manipulation, such that anticonvulsant effects of miR-124 are only observed when delivered to a normal brain. Antagomirs targeting miR-181a were reported to reduce neuronal death after status epilepticus in rats (39).
Limitations of Recent Studies and Future Directions
The recent findings are encouraging in identifying several new microRNAs that offer opportunities for seizure manipulation and neuroprotection. Despite this progress, a number of questions and concerns remain. Most of the studies tested with only a single model. Do the microRNA manipulations work in other models? Validation by an independent group and tests in more than one species would increase confidence in the value of specific microRNAs as targets. So far, microRNA manipulations have only been explored in chemoconvulsant models. These suffer certain confounders, and tests in models such as kindling or electrical stimulation of the perforant path are warranted. By what mechanism does the microRNA manipulation work? The targets of the microRNAs have often not been directly demonstrated or even investigated. Studies will need to prove colocalization and direct interaction of the microRNA and proposed target(s).
Another question is whether effects of microRNA manipulations are a combination of small effects on multiple targets or strong effects on just a few. Many of the recent functional studies on microRNAs have limitations in experimental design or data reporting. While acute anticonvulsant effects of microRNA manipulations are interesting, it is unlikely that a microRNA-based treatment would be used for this purpose unless it worked through markedly different mechanisms to current anticonvulsants. Rather, their therapeutic utility lies more in an ability to produce sustained disruption or stabilization of gene networks during development or maintenance of the epileptic state. Most of the recent studies, however, do not allow a distinction to be made between 1) an acute seizure-suppressive effect, 2) a chronic, long-lasting seizure-suppressive effect, and 3) a true antiepileptogenic/disease-modifying effect. Distinguishing between possibilities 2 and 3 may primarily depend on the duration of the microRNA manipulation's effects on the target. If the effect on seizures only persists as long as the direct microRNA manipulation effect is present but then wears off, it would be 2; if the effect on seizures persists longer or permanently, it would be 3. With perhaps one exception (31), the recent studies did not track animals long enough to conclude a disease-modifying effect occurred. Last, while perhaps a coincidence, many of the new microRNAs with antiseizure properties were downregulated after status epilepticus, whereas those with proseizure effects were upregulated. This was the case previously for miR-134 and miR-128. This finding may shape decision making around selecting microRNAs to target in the future.
Even if a microRNA manipulation passes the hurdles above, there remain important challenges. First, how will a microRNA-targeting treatment be given to a patient? Oligonucleotide-based molecules are too large to cross an intact blood-brain barrier. Delivery could be timed with injury-induced opening of the blood-brain barrier or via a route such as intrathecal or intranasal (26, 38). Alternatively, therapeutic application might use some kind of cell-penetrating adaptor or lipid-based carrier. The benefits of microRNAs owing to their multi-targeting actions may be offset by difficult-to-predict off-target effects. These could be limited, however, if the number of relevant targets is restricted or where the microRNA is expressed only within the brain region triggering seizures. Biotechnology companies are active in developing RNA-based therapeutics, and both an antagomir (40) and a mimic (41) have now been tested in clinical trials.
Highlights
MicroRNAs are important regulators of gene expression in the brain.
Studies on several new microRNAs have appeared in the past year that functionally interrogated the role.
Seven of these microRNAs, when blocked or overexpressed, were found to affect evoked or spontaneous seizures.
MicroRNA-based treatment may offer novel approaches to understanding the pathogenesis of epilepsy and may represent novel therapeutic targets.
In summary, the pace of recent functional studies interrogating microRNAs in animal models is encouraging. This must be accompanied, however, by a period of consolidation in which the reported effects are validated by others, the mechanisms better understood, and delivery options explored. If successful, we may see microRNA-based treatments enter preclinical development and, perhaps one day, reach patients.
Acknowledgments
The author would like to acknowledge funding from the European Union Seventh Framework Programme (FP7/2007-2013) under grant agreement No. 602130, Science Foundation Ireland (13/IA/1891, 14/ADV/RC2721, 11/TIDA/B1988, 08/IN1/B1875), and the Health Research Board Ireland (HRA-POR-2013-325).
Editor's Note: Authors have a Conflict of Interest disclosure which is posted under the Supplemental Materials (209.8KB, docx) link.
References
- 1. Pitkanen A, Lukasiuk K.. Mechanisms of epileptogenesis and potential treatment targets. Lancet Neurol 2011; 10: 173– 186. [DOI] [PubMed] [Google Scholar]
- 2. Goldberg EM, Coulter DA.. Mechanisms of epileptogenesis: A convergence on neural circuit dysfunction. Nat Rev Neurosci 2013; 14: 337– 349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Devinsky O, Vezzani A, Najjar S, De Lanerolle NC, Rogawski MA.. Glia and epilepsy: Excitability and inflammation. Trends Neurosci 2013; 36: 174– 184. [DOI] [PubMed] [Google Scholar]
- 4. Henshall DC, Kobow K.. Epigenetics and epilepsy. Cold Spring Harb Perspect Med 2015; 5: a022731 doi:10.1101/cshperspect.a022731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Johnson MR, Behmoaras J, Bottolo L, Krishnan ML, Pernhorst K, Santoscoy PL, Rossetti T, Speed D, Srivastava PK, Chadeau-Hyam M, Hajji N, Dabrowska A, Rotival M, Razzaghi B, Kovac S, Wanisch K, Grillo FW, Slaviero A, Langley SR, Shkura K, Roncon P, De T, Mattheisen M, Niehusmann P, O'Brien TJ, Petrovski S, von Lehe M, Hoffmann P, Eriksson J, Coffey AJ, Cichon S, Walker M, Simonato M, Danis B, Mazzuferi M, Foerch P, Schoch S, De Paola V, Kaminski RM, Cunliffe VT, Becker AJ, Petretto E.. Systems genetics identifies Sestrin 3 as a regulator of a proconvulsant gene network in human epileptic hippocampus. Nat Commun 2015; 6: 6031 doi:10.1038/ncomms7031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Fabian MR, Sonenberg N, Filipowicz W.. Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem 2010; 79: 351– 179. [DOI] [PubMed] [Google Scholar]
- 7. Ebert MS, Sharp PA.. Roles for microRNAs in conferring robustness to biological processes. Cell 2012; 149: 515– 524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Schmiedel JM, Klemm SL, Zheng Y, Sahay A, Blüthgen N, Marks DS, van Oudenaarden A.. Gene expression: MicroRNA control of protein expression noise. Science 2015; 348: 128– 132. [DOI] [PubMed] [Google Scholar]
- 9. Cattani AA, Allene C, Seifert V, Rosenow F, Henshall DC, Freiman TM.. Involvement of microRNAs in epileptogenesis. Epilepsia 2016; 57: 1015– 1026. [DOI] [PubMed] [Google Scholar]
- 10. O'Carroll D, Schaefer A.. General principals of miRNA biogenesis and regulation in the brain. Neuropsychopharmacology 2013; 38: 39– 54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Lin S, Gregory RI.. MicroRNA biogenesis pathways in cancer. Nat Rev Cancer 2015; 15: 321– 333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Chandradoss SD, Schirle NT, Szczepaniak M, MacRae IJ, Joo C.. A dynamic search process underlies microRNA targeting. Cell 2015; 162: 96– 107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Czech B, Hannon GJ.. Small RNA sorting: Matchmaking for Argonautes. Nat Rev Genet 2011; 12: 19– 31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Fromm B, Billipp T, Peck LE, Johansen M, Tarver JE, King BL, Newcomb JM, Sempere LF, Flatmark K, Hovig E, Peterson KJ.. A uniform system for the annotation of vertebrate microRNA genes and the evolution of the human microRNAome. Annu Rev Genet 2015; 49: 213– 242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Bernstein E, Kim SY, Carmell MA, Murchison EP, Alcorn H, Li MZ, Mills AA, Elledge SJ, Anderson KV, Hannon GJ.. Dicer is essential for mouse development. Nat Genet 2003; 35: 215– 217. [DOI] [PubMed] [Google Scholar]
- 16. Davis TH, Cuellar TL, Koch SM, Barker AJ, Harfe BD, McManus MT, Ullian EM.. Conditional loss of Dicer disrupts cellular and tissue morphogenesis in the cortex and hippocampus. J Neurosci 2008; 28: 4322– 4330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Tao J, Wu H, Lin Q, Wei W, Lu XH, Cantle JP, Ao Y, Olsen RW, Yang XW, Mody I, Sofroniew MV, Sun YE.. Deletion of astroglial dicer causes non-cell-autonomous neuronal dysfunction and degeneration. J Neurosci 2011; 31: 8306– 8319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Fiorenza A, Lopez-Atalaya JP, Rovira V, Scandaglia M, Geijo-Barrientos E, Barco A.. Blocking miRNA biogenesis in adult forebrain neurons enhances seizure susceptibility, fear memory, and food intake by increasing neuronal responsiveness. Cereb Cortex 2016; 26: 1619– 1633. [DOI] [PubMed] [Google Scholar]
- 19. Schratt GM, Tuebing F, Nigh EA, Kane CG, Sabatini ME, Kiebler M, Greenberg ME.. A brain-specific microRNA regulates dendritic spine development. Nature 2006; 439: 283– 239. [DOI] [PubMed] [Google Scholar]
- 20. Siegel G, Obernosterer G, Fiore R, Oehmen M, Bicker S, Christensen M, Khudayberdiev S, Leuschner PF, Busch CJ, Kane C, Hubel K, Dekker F, Hedberg C, Rengarajan B, Drepper C, Waldmann H, Kauppinen S, Greenberg ME, Draguhn A, Rehmsmeier M, Martinez, Schratt GM.. A functional screen implicates microRNA-138-dependent regulation of the depalmitoylation enzyme APT1 in dendritic spine morphogenesis. Nat Cell Biol 2009; 11: 705– 716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Magill ST, Cambronne XA, Luikart BW, Lioy DT, Leighton BH, Westbrook GL, Mandel G, Goodman RH.. MicroRNA-132 regulates dendritic growth and arborization of newborn neurons in the adult hippocampus. Proc Natl Acad Sci U S A 2010; 107: 20382– 20387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Zhao C, Sun G, Li S, Shi Y.. A feedback regulatory loop involving microRNA-9 and nuclear receptor TLX in neural stem cell fate determination. Nat Struct Mol Biol 2009; 16: 365– 371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Cheng LC, Pastrana E, Tavazoie M, Doetsch F.. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat Neurosci 2009; 12: 399– 408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Zou Y, Chiu H, Zinovyeva A, Ambros V, Chuang CF, Chang C.. Developmental decline in neuronal regeneration by the progressive change of two intrinsic timers. Science 2013; 340: 372– 376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Butovsky O, Jedrychowski MP, Cialic R, Krasemann S, Murugaiyan G, Fanek Z, Greco DJ, Wu PM, Doykan CE, Kiner O, Lawson RJ, Frosch MP, Pochet N, Fatimy RE, Krichevsky AM, Gygi SP, Lassmann H, Berry J, Cudkowicz ME, Weiner HL.. Targeting miR-155 restores abnormal microglia and attenuates disease in SOD1 mice. Ann Neurol 2015; 77: 75– 99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Jimenez-Mateos EM, Engel T, Merino-Serrais P, McKiernan RC, Tanaka K, Mouri G, Sano T, O'Tuathaigh C, Waddington JL, Prenter S, Delanty N, Farrell MA, O'Brien DF, Conroy RM, Stallings RL, Defelipe J, Henshall DC.. Silencing microRNA-134 produces neuroprotective and prolonged seizure-suppressive effects. Nat Med 2012; 18: 1087– 1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Tan CL, Plotkin JL, Veno MT, von Schimmelmann M, Feinberg P, Mann S, Handler A, Kjems J, Surmeier DJ, O'Carroll D, Greengard P, Schaefer A.. MicroRNA-128 governs neuronal excitability and motor behavior in mice. Science 2013; 342: 1254– 1258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. McSweeney KM, Gussow AB, Bradrick SS, Dugger SA, Gelfman S, Wang Q, Petrovski S, Frankel WN, Boland MB, Goldstein DB.. Inhibition of microRNA 128 promotes excitability of cultured cortical neuronal networks. Genome Res 2016; 26: 1411– 1416. doi:10.1101/gr.199828.115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Hu K, Xie YY, Zhang C, Ouyang DS, Long HY, Sun DN, Long LL, Feng L, Li Y, Xiao B.. MicroRNA expression profile of the hippocampus in a rat model of temporal lobe epilepsy and miR-34a-targeted neuroprotection against hippocampal neurone cell apoptosis post-status epilepticus. BMC Neurosci 2012; 13: 115 doi: 10.1186/1471-2202-13-115. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 30. Jimenez-Mateos EM, Bray I, Sanz-Rodriguez A, Engel T, McKiernan RC, Mouri G, Tanaka K, Sano T, Saugstad JA, Simon RP, Stallings RL, Henshall DC.. miRNA expression profile after status epilepticus and hippocampal neuroprotection by targeting miR-132. Am J Pathol 2011; 179: 2519– 2532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Yuan J, Huang H, Zhou X, Liu X, Ou S, Xu T, Li R, Ma L, Chen Y.. MicroRNA-132 interact with p250GAP/Cdc42 pathway in the hippocampal neuronal culture model of acquired epilepsy and associated with epileptogenesis process. Neural Plast 2016; 2016: 5108489 doi:10.1155/2016/5108489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Jimenez-Mateos EM, Arribas-Blazquez M, Sanz-Rodriguez A, Concannon C, Olivos-Ore LA, Reshcke CR, Mooney CM, Mooney C, Lugara E, Morgan J, Langa E, Jimenez-Pacheco A, Almeida-Silva L-F, Mesuret GB, Boison D, Miras-Portugal MT, Letavic M, Artalejo AR, Bhattacharya A, Diaz-Hernandez M, Henshall DC, Engel T.. MicroRNA targeting of the P2X7 purinoceptor opposes a contralateral epileptogenic focus in the hippocampus. Sci Rep 2015; 5: e17486 doi:10.1038/srep17486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Zhan L, Yao Y, Fu H, Li Z, Wang F, Zhang X, He W, Zheng W, Zhang Y, Zheng H.. Protective role of miR-23b-3p in kainic acid-induced seizure. Neuroreport 2016; 27: 764– 768. [DOI] [PubMed] [Google Scholar]
- 34. Wang W, Wang X, Chen L, Zhang Y, Xu Z, Liu J, Jiang G, Li J, Zhang X, Wang K, Wang J, Chen G, Luo J.. The microRNA miR-124 suppresses seizure activity and regulates CREB1 activity. Expert Rev Mol Med 2016; 18: e4 doi:10.1017/erm.2016.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Zheng H, Tang R, Yao Y, Ji Z, Cao Y, Liu Z, Peng F, Wang W, Can D, Xing H, Bu G, Xu H, Zhang YW, Zheng W.. MiR-219 protects against seizure in the kainic acid model of epilepsy. Mol Neurobiol 2016; 53: 1– 7. [DOI] [PubMed] [Google Scholar]
- 36. Cai Z, Li S, Li S, Song F, Zhang Z, Qi G, Li T, Qiu J, Wan J, Sui H, Guo H.. Antagonist targeting microRNA-155 protects against lithium-pilocarpine-induced status epilepticus in C57BL/6 mice by activating brain-derived neurotrophic factor. Front Pharmacol 2016; 7: 129 doi:10.3389/fphar.2016.00129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Wang D, Li Z, Zhang Y, Wang G, Wei M, Hu Y, Ma S, Jiang Y, Che N, Wang X, Yao J, Yin J.. Targeting of microRNA-199a-5p protects against pilocarpine-induced status epilepticus and seizure damage via SIRT1-p53 cascade. Epilepsia 2016; 57: 706– 716. [DOI] [PubMed] [Google Scholar]
- 38. Lee ST, Jeon D, Chu K, Jung KH, Moon J, Sunwoo J, Park DK, Yang H, Park JH, Kim M, Roh JK, Lee SK.. Inhibition of miR-203 reduces spontaneous recurrent seizures in mice [published online ahead of print May 10, 2016]. Mol Neurobiol. In press. [DOI] [PubMed]
- 39. Ren L, Zhu R, Li X. . Silencing miR-181a produces neuroprotection against hippocampus neuron cell apoptosis post-status epilepticus in a rat model and in children with temporal lobe epilepsy. Genet Mol Res 2016; 15 doi:10.4238/gmr.15017798. [DOI] [PubMed] [Google Scholar]
- 40. Janssen HL, Reesink HW, Lawitz EJ, Zeuzem S, Rodriguez-Torres M, Patel K, van der Meer AJ, Patick AK, Chen A, Zhou Y, Persson R, King BD, Kauppinen S, Levin AA, Hodges MR.. Treatment of HCV infection by targeting microRNA. New Engl J Med 2013; 368: 1685– 1694. [DOI] [PubMed] [Google Scholar]
- 41. Bouchie A. First microRNA mimic enters clinic. Nat Biotechnol 2013; 31: 577 doi:10.1038/nbt0713-577. [DOI] [PubMed] [Google Scholar]