

Abstract
Inositol polyphosphate-5-phosphatase (INPP5D) was reported to be associated with Alzheimer's disease (AD) through modulating the inflammatory process and immune response. A recent genome-wide association study discovered a new locus single nucleotide polymorphism (SNP, rs35349669) of INPP5D which was significantly associated with susceptibility to late-onset Alzheimer's disease (LOAD) in Caucasians. In this study, we investigated the relations between the INPP5D polymorphism rs35349669 and LOAD in Han Chinese population comprising 984 LOAD cases and 1352 healthy controls being matched for age and gender. Our results showed no obvious differences in the genotypic or allelic distributions of rs35349669 polymorphism between LOAD cases and healthy controls (genotype: p = 0.167; allele: p = 0.094). Additionally, when these data were stratified by APOEε4 status, there are still no evident differences in the genotypic or allelic distributions in APOEε4 carriers (p > 0.05). Furthermore, meta-analysis of 81964 individuals confirmed that rs35349669 was significantly associated with the risk for LOAD (OR=1.08, 95%CI=1.06-1.11), but the results remained negative in Chinese subgroup (OR=0.77, 95%CI=0.53-1.13). Overall, the current evidence did not indicate that INPP5D rs35349669 polymorphism play a role in the genetic predisposition to LOAD in Chinese population.
Keywords: Alzheimer's disease, INPP5D, rs35349669, association study, meta-analysis, Gerotarget
INTRODUCTION
Alzheimer's disease (AD), the most common form of dementia in the elderly [1], is a neurodegenerative disorder that is pathologically characterized by deposition of extracellular amyloid-beta (Aβ) plaques, intracellular neurofibrillary tangle composed of hyperphosphorylated tau and massive neuronal loss [2, 3]. It is well acknowledged that genetic variations play an important role in the progression of AD [4]. Mutations in APP, PSEN1 and PSEN2 are associated with early-onset familial AD, which is responsible for part of AD [5]. The genetic background of the more common late-onset Alzheimer's disease (LOAD) is more complicated, and only the variant of the apolipoprotein E (APOE) gene has been consistently related to the risk of this disease [6]. In addition to the established association of APOE, recent genome-wide association studies (GWAS) have identified other 20 genes/loci that affected the susceptibility of LOAD [7].
Given the significant part of inflammation in AD pathogenesis, inflammation-relevant genes including Inositol polyphosphate-5-phosphatase (INPP5D) are increasingly investigated in AD genetic association studies. INPP5D, mainly expressed in hematopoietic cells, plays an important role in a series of inflammatory responses [8].It has also reported that INPP5D was implicated in the pathogenesis of LOAD through the regulation of microglial cell function [8, 9].Recently, INPP5D rs35349669 polymorphism was recognized to be strongly associated with the development of LOAD in 74,046 Caucasians [10]. Since variants and their frequencies of INPP5D in diverse ethnic groups might be different, replication is necessary to validate the potential effects of INPP5D in non-Caucasian population including Asians. To address this question, the present study was conducted to evaluate the association of rs35349669 SNP with LOAD in Northern Han Chinese population.
RESULTS
The demographic and clinical characteristics of the subjects are detailed in Table 1. LOAD patients were well-matched with controls in terms of age and gender. No statistically significant differences were detected for age (age at onset for LOAD and age at examination for controls) and gender (P > 0.05). As expected, MMSE scores were obviously less in LOAD patients than that in control subjects (P < 0.001). The APOE4 allele frequency was also obviously different between patients and control subjects (P < 0.001). Distributions of the APOE and rs35349669 genotypes in cases and controls were all in the Hardy–Weinberg equilibrium (P > 0.05).
Table 1. The characteristics of the study population.
| AD (n = 984) | HC (n = 1352) | P value | |
|---|---|---|---|
| Age, years; mean±SD | 75.15±6.08 | 75.50±6.49 | 0.185* |
| Gender, n (%) | 0.074 | ||
| Male | 406 (41.3) | 608 (45.0) | |
| Female | 578 (58.7) | 744 (55.0) | |
| MMSE score, mean±SD | 11.99±6.20 | 28.49±1.09 | <0.001 |
| APOE ε4 status, n (%) | <0.001 | ||
| APOE ε4 (+) | 280 (28.5) | 191 (14.1) | |
| APOE ε4 (-) | 704 (71.5) | 1161 (85.9) |
Abbreviations: AD, Alzheimer's disease; HC, healthy controls; MMSE, Mini-Mental State Examination; ApoE; apolipoprotein E; SD, standard deviation.
P value was calculated with the age of onset for late-onset AD and age at examination for control. Differences in the characteristics of age and MMSE score between the two groups were examined using Student's t test. Differences in gender and ApoE-ε4 frequency between AD patients and HC were assessed using the Pearson χ2 test.
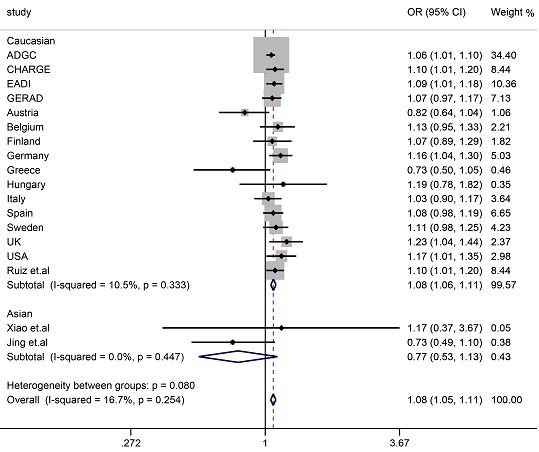
The allele and genotype distributions of rs35349669 in two groups are presented in Table 2. The genotype and allele frequencies did not differ between LOAD and controls (genotype P = 0.167, allele P = 0.094). To rule out blending effects in our initial association analyses, we reassessed rs35349669 effects under various genetic models in logistic regression adjusting for age at onset in LOAD patients (age at examination in control group), gender, and APOE ε4 status. Disappointedly, the result also failed to reveal any significant difference between LOAD and controls. Thus, our study observed no significant differences in the genotypic or allelic distributions of rs35349669 polymorphism between LOAD cases and healthy controls in a Northern Han Chinese population. Meanwhile, we carried out a meta-analysis about the association of rs35349669 with LOAD and found rs35349669 was strongly associated with LOAD (OR = 1.08, 95%CI = 1.05-1.11) (Figure 1) without evident analysis heterogeneity (I2 = 16.7%). In subgroup analysis, the rs35349669 polymorphism was also significantly associated with the risk for LOAD in Caucasian, however, the results showed that there was no association of rs35349669 with AD risk in Chinese population (OR = 0.77, 95%CI = 0.53~1.13).
Table 2. Distribution of the rs35349669 genotypes and alleles in the entire group and subgroup stratified by APOE ε4.
| rs35349669 | n | Genotypesn (%) | Allelen (%) | |||||
|---|---|---|---|---|---|---|---|---|
| TT | CT | CC | P | T | C | P | ||
| AD | 984 | 0 (0) | 54 (5.5) | 930 (94.5) | 0.167 | 54(2.7) | 1914 (97.3) | 0.094 |
| Controls | 1352 | 2 (0.1) | 94 (7.0) | 1256 (92.9) | 98(3.6) | 2606 (96.4) | ||
| APOE ε4 (+) | ||||||||
| AD | 280 | 0 (0) | 18 (6.4) | 262 (93.6) | 0.713 | 18 (3.2) | 542 (96.8) | 0.708 |
| Controls | 191 | 0 (0) | 14 (7.3) | 177 (92.7) | 14 (3.7) | 368 (96.3) | ||
| APOE ε4 (-) | ||||||||
| AD | 704 | 0 (0) | 36 (5.1) | 668 (94.9) | 0.176 | 36 (2.6) | 1372 (97.4) | 0.075 |
| Controls | 1161 | 2 (0.2) | 80 (6.9) | 1079 (92.9) | 84 (3.6) | 2238 (96.4) | ||
Figure 1. Forest plot for rs35349669 in LOAD and healthy controls in 81964 individuals.
In addition, we assessed the influence of the interaction between rs35349669 and APOE genotype on the risk for LOAD in logistic regression models (Table 3), no interaction on AD risk was detected here (dominant, P = 0.151; additive, P = 0.127). In order to further investigate whether the presence of the APOE ε4 allele modified the association of rs35349669 with LOAD, the total group was stratified in by APOE ε4 carriers and APOE ε4 noncarriers. We observed no significant differences between AD and controls in genotype and allele distributions (P > 0.05) (Table 2), and in the multivariate analysis (Table 3). To exclude the effects of gender on our initial association analyses, the total group was also stratified into male and female subgroup by gender (Table 4). Likewise, the frequency of genotypes and allele for rs35349669 did not show any obvious differences between AD and controls in male or female subgroup (P > 0.05) (Table 4).
Table 3. Logistic regression analysis of INPP5D rs35349669 and AD.
| SNP | Total samplea | APOE ε4 (+)b | APOEε4 (-)b | ||||
|---|---|---|---|---|---|---|---|
| rs35349669 | OR(95%CI) | P | P for APOE interaction | OR(95%CI) | P | OR(95%CI) | P |
| Dom | 0.773 (0.543-1.099) | 0.151 | 1.167 | 0.846 (0.407-1.758) | 0.654 | 0.744 (0.495-1.117) | 0.154 |
| Add | 0.763 (0.539-1.080) | 0.127 | 0.689 | 0.846 (0.407-1.758) | 0.654 | 0.733 (0.492-1.093) | 0.128 |
| Rec | - | - | - | - | - | - | - |
APOE, apolipoprotein E; ApoEε4 (+), APOE ε4 allele carrier; ApoEε4 (−), APOE ε4 allele noncarrier; SNP, single nucleotide polymorphism; Dom, dominant; Rec, recessive; Add, additive model; OR, odd ratio; CI, confidence interval
, adjusted for age, gender, and APOE ε4 allele status;
, adjusted for age, gender.
, the recessive genetic model could not be analysed when minor allele homozygote counts <14.
Table 4. INPP5D genotype and allele frequencies in AD cases and controls stratified by sex.
| N | Genotypes, n (%) | P | Alleles, n (%) | P | ||||
|---|---|---|---|---|---|---|---|---|
| CC | TC | TT | C | T | ||||
| AD | 984 | 930 (94.5) | 54 (5.5) | 0(0) | 0.167 | 1914 (97.3) | 54 (2.7) | 0.094 |
| Controls | 1352 | 1256 (92.9) | 94 (7.0) | 2 (0.1) | 2606 (96.4) | 98 (3.6) | ||
| Male | 1014 | 934 (92.11) | 78 (7.69) | 2 (0.20) | 1946 (95.96) | 82 (4.04) | ||
| AD | 406 | 378 (93.10) | 28 (6.90) | 0 | 0.464a | 784 (40.29) | 28 (34.15) | 0.266 |
| Controls | 608 | 556 (91.45) | 50 (8.22) | 2 (0.33) | 1162 (59.71) | 54 (65.85) | ||
| Female | 1322 | 1252 (94.70) | 70 (5.30) | 0 | 2574 (97.35) | 70 (2.65) | ||
| AD | 578 | 552 (95.5) | 26 (4.50) | 0 | 0.268a | 1130 (43.90) | 26 (37.14) | 0.261 |
| Controls | 744 | 700 (94.10) | 44 (5.9) | 0 | 1444 (56.10) | 44 (62.86) | ||
Fisher's exact test was performed
DISCUSSION
INPP5D, as a member of the inositol polyphosphate-5-phosphatase (INPP5) family, is located on chromosome 2q37.1 [8, 11], it was implicated in AD pathogenesis through microglia-mediated inflammatory process and immune response [8, 12, 13]. Our study replicated the association between the rs35349669 polymorphism within INPP5D and LOAD risk in Northern Han Chinese. Finally, we failed to find significant differences in the genotypic or allelic distributions of rs35349669 polymorphism between LOAD cases and healthy controls in a Han Chinese population, even after adjustment for age, gender, and APOE ε4 status, these findings might have resulted from the relatively small sample size of our experimental study, in order to reduce the possibility, we explored the association of rs35349669 with LOAD risk in the meta-analysis in 81 964 individuals including Caucasian and Chinese, rs35349669 polymorphism was strongly associated with AD risk in the entire population (OR = 1.08, 95%CI = 1.06-1.11)and in Caucasian (OR = 1.06, 95%CI = 1.01-1.10), while rs35349669 polymorphism did not relate to LOAD in Chinese. Therefore, the current evidence did not support the correlation between rs35349669 and LOAD in Chinese.
Recently, INPP5D rs35349669 polymorphism was identified to be strongly associated with the development of LOAD (P < 0.05) in two-stage meta-analysis of GWAS in 74,046 Caucasians [10]. Subsequently, Ruiz et al. confirmed the significant association in Spanish, however, our study and Xiao et al. failed to replicate the association in Caucasian population. Several factors may be responsible for the inconsistency: Firstly, the genetic heterogeneity which is inherent in different ethnic populations could be the main reason. The minor allele frequency (MAF) of rs35349669 in Caucasians is significantly different from the MAF of rs35349669 in Chinese based on the dbSNP database (http://www.ncbi.nlm.nih.gov/projects/SNP). Alternatively, the effects of rs35349669 may be population-specific, possibly due to specific interactions between gene and environment. The differentiated effects of INPP5D rs35349669 polymorphism on AD probably attribute to a brain structural mechanism for such population specific genetic effects. In addition to the population specification and special genetic backgrounds, environments, educational background, sample size, etc. may be the source of different results between Caucasian and Chinese. On the other hand, although our study failed to replicate any association of the examined SNP with LOAD in Chinese, we could not rule out the possibility that other SNPs of INPP5D associated with LOAD.
The function of INPP5D in immune response and inflammation in the central nervous system(CNS) is still poorly understood. Recent studies showed that the human SHIP protein, encoded by the INPP5D gene, was supposed to restrain the release of diverse inflammatory cytokines from microglia,astrocytes or even neurons, such as IL-6, IL-8,TNF-α [11]. In addition, a previous study revealed that the binding of INPP5D with the product of CD2AP, another LOAD risk gene, could control degradation of IgE receptor FceRIc, some IgE receptors are members of the MS4A gene superfamily that has been correlated with LOAD risk [14]. Besides, according to a clinical-pathologic correlation study about AD, the minor allele (T) from rs35349669 of the INPP5D gene was associated with other neuropathologies coexist with AD pathology, such as microinfarcts and LB disease [15]. With renewed genetic sequencing about INPP5D in the near future, it may be helpful to find new loci that related to LOAD.
In conclusion, we were not able to detect the significant association of INPP5D rs35349669 polymorphism with LOAD in Chinese. It is likely that the implication of INPP5D variation in AD risk may be specific to particular ethnic groups, besides, the implication is too small to be detected responsibly by a cohort of our size. It warranted investigators to clarify the role of INPP5D polymorphisms in LOAD in larger cohorts and in other ethnic populations.
MATERIALS AND METHODS
Subjects
We investigated 984 sporadic LOAD patients (mean age at onset: 75.15 ± 6.08 years; 578women) and 1352 healthy control subjects (mean age at examination: 75.50 ± 6.49 years; 744 women) matched for sex and age. All the subjects were unrelated Northern Han Chinese originated from Shandong Province. The patients were recruited from the Department of Neurology at Qingdao Municipal Hospital and some other hospitals in Shandong province. The diagnose of probable AD met with the criteria of the National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer's disease and Related Disorders Association [16]. No family history of neurodegenerative disorders or other dementias were reported among AD patients. The control subjects were enrolled from the Health Examination Center of the Qingdao Municipal Hospital according to the principles described elsewhere [2].They were identified healthy and neurologically normal according to medical history, general examinations, laboratory examinations and acquired ≥28 points on the Mini-Mental State Examination (MMSE). Informed consent was obtained from all individuals or from the subject's guardian, and the protocol of this study was approved by the Ethical Committee of Qingdao Municipal Hospital.
Genotyping
Genomic DNA was extracted from peripheral blood leukocytes of AD patients and healthy individuals using the Wizard genomic DNA purification kit (Cat. #A1125, Promega, USA).Genotyping of INPP5D (rs35349669) and APOE polymorphisms was performed by a custom-by-design 48-PlexSNP scan TM Kit (Genesky Biotechnologies, Inc., Shanghai, China), as previously described [17]. This was high-throughput and cost-saving SNP genotyping methods based on double ligation and multiplex fluorescence PCR. The genotyping of APOE were carried out by Shanghai Genesky Bio-Tech Co., Ltd. (http://biotech.geneskies.com/index.html) using the improved multiplex ligase detection reaction (iMLDR) method [18]. Data analysis was accomplished using GeneMapper Software v4.0 (Applied Biosystems). Randomly selected DNA samples from each genotype were analyzed in duplicate using ligation detection reaction and sequence analysis method. Consistent results were achieved by these two methods.
Statistical analysis
Statistical analysis was performed using SPSS16.0 software. Genotype and allele frequencies were calculated by counting. The Hardy–Weinberg equilibrium (HWE) was tested for each SNP with genotype data from both AD patients and controls. Differences in the characteristics between the two groups were tested using Student's t-test or the chi-square test. Genotypes and alleles frequencies between LOAD and healthy control group were compared using Pearson χ2 test or Fisher's exact test. Differences between cases and controls after stratification for APOE4 status were also examined by the chi-square test. The association of rs35349669 with LOAD risk was further analyzed using logistic regression adjusting for age, gender, and APOE 4 status under various genetic models that were defined as 1 (TT + TC) versus 0 (CC) for dominant, 1 (TT) versus 0 (TC + CC) for recessive, and 0 (CC) versus 1 (TC) versus 2 (TT) for additive. The P value, odds ratios (ORs) and 95% confidence intervals (CIs) were calculated. Estimation of the statistical power was performed with the STPLAN 4.3 software. Values of P < 0.05 were considered statistically significant.
Additionally, we also combined our data with the results from other studies about rs35349669 and LOAD risk [10, 19, 20] through fixed-effects inverse variance-weighted methods. Meanwhile, we generated I2 estimates with evaluate the possible effect of study heterogeneity on the results. Stata V.12.0 was used to perform all these analyses.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (81471309, 81371406, 81571245, and 81501103), the Shandong Provincial Outstanding Medical Academic Professional Program, Qingdao Key Health Discipline Development Fund, Qingdao Outstanding Health Professional Development Fund, and Shandong Provincial Collaborative Innovation Center for Neurodegenerative Disorders.
Footnotes
CONFLICTS OF INTEREST
The authors disclose no conflicts of interest.
REFERENCES
- 1.Tan L, Yu JT, Zhang W, Wu ZC, Zhang Q, Liu QY, Wang W, Wang HF, Ma XY, Cui WZ. Association of GWAS-linked loci with late-onset Alzheimer's disease in a northern Han Chinese population. Alzheimer's & dementia. 2013;9:546–553. doi: 10.1016/j.jalz.2012.08.007. [DOI] [PubMed] [Google Scholar]
- 2.Wang J, Yu JT, Jiang T, Tan MS, Wang HF, Tan L, Hu N, Sun L, Zhang W, Tan L. Association of LRRTM3 polymorphisms with late-onset Alzheimer's disease in Han Chinese. Experimental gerontology. 2014;52:18–22. doi: 10.1016/j.exger.2014.01.013. [DOI] [PubMed] [Google Scholar]
- 3.Tan MS, Yu JT, Jiang T, Zhu XC, Wang HF, Zhang W, Wang YL, Jiang W, Tan L. NLRP3 polymorphisms are associated with late-onset Alzheimer's disease in Han Chinese. Journal of neuroimmunology. 2013;265:91–95. doi: 10.1016/j.jneuroim.2013.10.002. [DOI] [PubMed] [Google Scholar]
- 4.Yu JT, Song JH, Ma T, Zhang W, Yu NN, Xuan SY, Tan L. Genetic association of PICALM polymorphisms with Alzheimer's disease in Han Chinese. Journal of the neurological sciences. 2011;300:78–80. doi: 10.1016/j.jns.2010.09.027. [DOI] [PubMed] [Google Scholar]
- 5.Miyashita A, Wen Y, Kitamura N, Matsubara E, Kawarabayashi T, Shoji M, Tomita N, Furukawa K, Arai H, Asada T, Harigaya Y, Ikeda M, Amari M, Hanyu H, Higuchi S, Nishizawa M, et al. Lack of genetic association between TREM2 and late-onset Alzheimer's disease in a Japanese population. Journal of Alzheimer's disease. 2014;41:1031–1038. doi: 10.3233/JAD-140225. [DOI] [PubMed] [Google Scholar]
- 6.Liu QY, Miao D, Yu JT, Xing YY, Zhong XL, Wu ZC, Zhang Q, Tan L. Lack of association between rs597668 polymorphism near EXOC3L2 and late-onset Alzheimer's disease in Han Chinese. Neuroscience letters. 2012;513:174–177. doi: 10.1016/j.neulet.2012.02.030. [DOI] [PubMed] [Google Scholar]
- 7.Wang X, Lopez OL, Sweet RA, Becker JT, DeKosky ST, Barmada MM, Demirci FY, Kamboh MI. Genetic determinants of disease progression in Alzheimer's disease. Journal of Alzheimer's disease. 2015;43:649–655. doi: 10.3233/JAD-140729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Rosenthal SL, Kamboh MI. Late-Onset Alzheimer's Disease Genes and the Potentially Implicated Pathways. Current genetic medicine reports. 2014;2:85–101. doi: 10.1007/s40142-014-0034-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wilkins HM, Carl SM, Greenlief AC, Festoff BW, Swerdlow RH. Bioenergetic dysfunction and inflammation in Alzheimer's disease: a possible connection. Frontiers in aging neuroscience. 2014;6:311. doi: 10.3389/fnagi.2014.00311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lambert JC, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R, Bellenguez C, DeStafano AL, Bis JC, Beecham GW, Grenier-Boley B, Russo G, Thorton-Wells TA, Jones N, Smith AV, Chouraki V, Thomas C, et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer's disease. Nature genetics. 2013;45:1452–1458. doi: 10.1038/ng.2802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Zhang ZG, Li Y, Ng CT, Song YQ. Inflammation in Alzheimer's Disease and Molecular Genetics: Recent Update. Archivum immunologiae et therapiae experimentalis. 2015;63:333–344. doi: 10.1007/s00005-015-0351-0. [DOI] [PubMed] [Google Scholar]
- 12.Karch CM, Goate AM. Alzheimer's disease risk genes and mechanisms of disease pathogenesis. Biological psychiatry. 2015;77:43–51. doi: 10.1016/j.biopsych.2014.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schott JM, Crutch SJ, Carrasquillo MM, Uphill J, Shakespeare TJ, Ryan NS, Yong KX, Lehmann M, Ertekin-Taner N, Graff-Radford NR, Boeve BF, Murray ME, Khan QU, Petersen RC, Dickson DW, Knopman DS, et al. Genetic risk factors for the posterior cortical atrophy variant of Alzheimer's disease. Alzheimer's & dementia. 2016;12:862–71. doi: 10.1016/j.jalz.2016.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Bao M, Hanabuchi S, Facchinetti V, Du Q, Bover L, Plumas J, Chaperot L, Cao W, Qin J, Sun SC, Liu YJ. CD2AP/SHIP1 complex positively regulates plasmacytoid dendritic cell receptor signaling by inhibiting the E3 ubiquitin ligase Cbl. Journal of immunology. 2012;189:786–792. doi: 10.4049/jimmunol.1200887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Farfel JM, Yu L, Buchman AS, Schneider JA, De Jager PL, Bennett DA. Relation of genomic variants for Alzheimer disease dementia to common neuropathologies. Neurology. 2016;87:489–496. doi: 10.1212/WNL.0000000000002909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McKhann G, Drachman D, Folstein M, Katzman R, Price D, Stadlan EM. Clinical diagnosis of Alzheimer's disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer's Disease. Neurology. 1984;34:939–944. doi: 10.1212/wnl.34.7.939. [DOI] [PubMed] [Google Scholar]
- 17.Zhou X, Zhu J, Zhang H, Zhou G, Huang Y, Liu R. Is the microRNA-146a (rs2910164) polymorphism associated with rheumatoid arthritis? Association of microRNA-146a (rs2910164) polymorphism and rheumatoid arthritis could depend on gender. Joint, bone, spine. 2015;82:166–171. doi: 10.1016/j.jbspin.2014.12.009. [DOI] [PubMed] [Google Scholar]
- 18.Zhang L, Fan D, Liu L, Yang T, Ding N, Hu Y, Cai G, Wang L, Xin L, Xia Q, Li X, Xu S, Xu J, Yang X, Zou Y, Pan F. Association Study of IL-12B Polymorphisms Susceptibility with Ankylosing Spondylitis in Mainland Han Population. PloS one. 2015;10:e0130982. doi: 10.1371/journal.pone.0130982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Xiao Q, Liu ZJ, Tao S, Sun YM, Jiang D, Li HL, Chen H, Liu X, Lapin B, Wang CH, Zheng SL, Xu J, Wu ZY. Risk prediction for sporadic Alzheimer's disease using genetic risk score in the Han Chinese population. Oncotarget. 2015;6:36955–36964. doi: 10.18632/oncotarget.6271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ruiz A, Heilmann S, Becker T, Hernandez I, Wagner H, Thelen M, Mauleon A, Rosende-Roca M, Bellenguez C, Bis JC, Harold D, Gerrish A, Sims R, Sotolongo-Grau O, Espinosa A, Alegret M, et al. Follow-up of loci from the International Genomics of Alzheimer's Disease Project identifies TRIP4 as a novel susceptibility gene. Translational psychiatry. 2014;4:e358. doi: 10.1038/tp.2014.2. [DOI] [PMC free article] [PubMed] [Google Scholar]