

Abstract
Most patients with epithelial ovarian cancer eventually die due to recurrence. However, little is known about the microRNA (miRNA) expression pattern and its involvement in recurrent ovarian cancer. We analyzed miRNA expression profiles related to the recurrence of advanced ovarian high-grade serous carcinoma (HGSC) using miRNA microarrays. Between May 2006 and December 2012, 37 ovarian HGSC patients underwent secondary cyto-debulking surgery at recurrence. Among them, only 8 pairs of primary and recurrent tumor samples were deemed to be adequate for analysis. The expression profiles of primary ovarian HGSC compared with normal ovarian tissue were significantly consistent with those of recurrent ovarian HGSC compared with normal ovarian tissue (correlation coefficient = 0.81, P = 0.0078). Among 31 miRNAs that increased by more than 4-fold in primary tumors compared with normal ovarian tissue, 27 were also significantly increased in recurrent tumor samples. Likewise, among 35 miRNAs that decreased by more than 4-fold in primary tumors compared with normal ovarian tissue, 34 were also significantly decreased in recurrent tumor samples. We identified 60 miRNAs that were significantly increased in recurrent serous ovarian carcinoma compared with primary tumor tissue, including miR-630, miR-370, and miR-575. Additionally, 52 miRNAs were significantly decreased in recurrent samples, including miR-509-3p, miR-514a-3p, and miR-506-3p. Our results demonstrate that primary and recurrent ovarian HGSC displayed similar miRNA expression patterns. Nevertheless, altered miRNA expression could be implicated in the recurrence of ovarian HGSC, and further study is needed to validate these data in independent cases using a homogeneous methodology.
Keywords: recurrence, ovarian cancer, microRNA, chemoresistant
INTRODUCTION
Epithelial ovarian cancer is the most lethal malignancy among gynecological cancers, reflecting early peritoneal dissemination with advanced-stage disease at diagnosis [1]. The incidence of ovarian carcinoma exceeded 22,000 new cases in the United States in 2012 and was responsible for approximately 14,000 cancer-related deaths [2]. Although radical surgery and chemotherapy frequently afford complete remission, most patients eventually succumb to chemoresistant disease [1]. Therefore, a greater understanding of the molecular mechanisms related to recurrence is needed for the development of enhanced therapeutic approaches to treat ovarian cancer.
MicroRNAs (miRNAs) are non-coding, single-stranded RNAs of approximately 22 nucleotides in length that constitute a novel class of gene regulators [3]. miRNAs are involved in various biological processes, including cell differentiation, proliferation, apoptosis, carcinogenesis, and metabolism [4]. Although previous studies have characterized gene expression profiles, including those of miRNAs, in primary ovarian cancers [5–7], only limited data are available regarding miRNA gene expression profiles in recurrent ovarian cancers [8–13]. This is largely due to a lack of available tumor specimens from recurrent ovarian cancer patients, since secondary surgeries or biopsies are rarely performed in these patients. In addition, there are no suitable in vitro or in vivo pre-clinical models to study recurrent cancers.
In this study, we analyzed miRNA expression profiles associated with the recurrence of advanced high-grade serous ovarian carcinoma (HGSC). To minimize the effect of genetic differences among individual patients, we evaluated paired primary ovarian tumor tissue and corresponding tumor tissue from the same patients at recurrence.
RESULTS
Comparison of miRNA expression profiles between fresh-frozen and paraffin-embedded ovarian cancer tissue
To determine whether miRNA expression profiles are preserved in FFPE specimens compared with those in SF specimens, expression of 1205 human miRNAs in SF and FFPE specimens were compared using miRNA microarrays. All patients had primary advanced ovarian HGSC (Supplementary Table S1). Five samples in SF tissues and five corresponding FFPE samples from the same patients were compared using a Pearson correlation test (Figure 1). Three SF normal ovarian tissue samples from patients treated for benign uterine disease were used as a control. There was a significant correlation between the expression of each miRNA in all samples (P < 0.001).
Figure 1. Comparison of miRNA expression between fresh snap-frozen (SF) and paraffin-embedded (FFPE) ovarian cancer tissue.
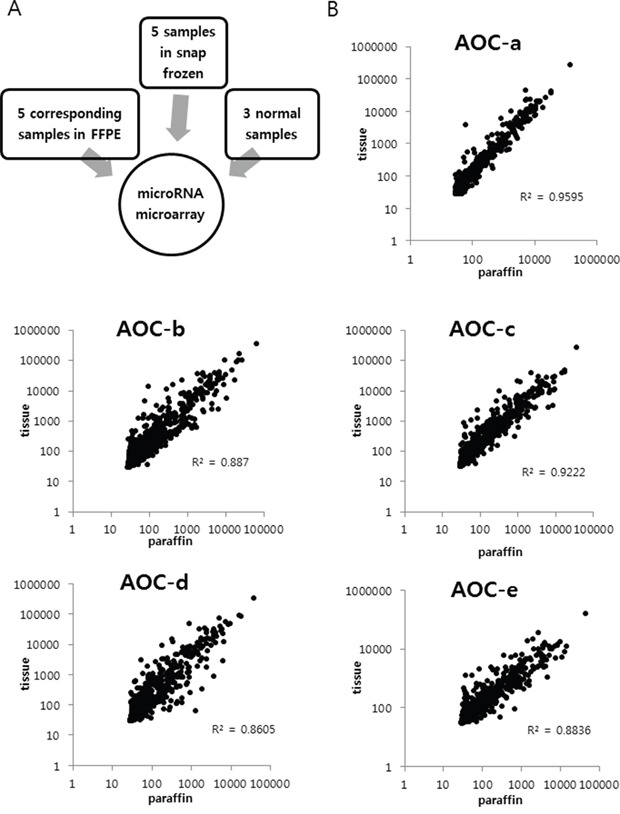
All patients had primary advanced serous ovarian cancer. A. Five samples in SF tissues and five corresponding FFPE samples from the same patients were compared. B. A significant correlation was observed between the miRNA expression profiles of snap-frozen tissue and FFPE tissue in five patients (AOC-a, b, c, d, and e). (P < 0.001).
Patient characteristics
miRNA expression profiles associated with recurrent ovarian HGSC were analyzed in eight patients. All patients were in an advanced disease stage and primary cyto-debulking operations were performed in all cases (Table 1). All patients received post-operative adjuvant chemotherapy (POAC) consisting of six courses of paclitaxel and carboplatin. Secondary cytoreductive operations were performed at recurrence in seven patients. One patient (P3) had palliative surgery immediately following POAC due to intestinal obstruction; a tumor sample was obtained during this surgery. Two patients (P1 and P4) had platinum-sensitive recurrence and five patients (P2, P5, P6, P7, and P8) had marginally platinum-sensitive recurrence (platinum-free interval: 6–12 months). The median number of chemotherapy lines before secondary surgery was 2, with a range of 1 to 4 chemotherapy lines. In four patients (P1, P2, P3, and P4), specimens from both primary cyto-debulking operations and secondary surgeries were stored as SF tissue, and specimens from the other patients (P5, P6, P7, and P8) were stored as FFPE tissue.
Table 1. Patient characteristics.
| Patient number | Tissue type | Stage | Grade | Date of primary surgery | Number of chemotherapy lines | Date of secondary surgery | Site of recurrent tumor | PFS | Recurrence type |
|---|---|---|---|---|---|---|---|---|---|
| P1 | SF | IIIc | G3 | 2010-02-08 | 1 | 2011-12-01 | Pelvic cavity | 16 | PS |
| P2 | SF | IIIc | G2 | 2010-12-09 | 1 | 2011-12-16 | Lymph node | 8 | PS |
| P3 | SF | IIIc | G2 | 2011-11-23 | 1 | 2012-07-10 | Abdominal wall | 0 | PR |
| P4 | SF | IIIa | G3 | 2009-10-05 | 1 | 2012-05-16 | Pelvic cavity | 27 | PS |
| P5 | FFPE | IIIc | G3 | 2006-05-30 | 3 | 2009-12-02 | Left psoas muscle | 6 | PS |
| P6 | FFPE | IV | G3 | 2010-12-24 | 1 | 2011-12-06 | Sub-hepatic mass | 6 | PS |
| P7 | FFPE | IV | G2 | 2011-08-18 | 1 | 2012-08-23 | Liver | 7 | PS |
| P8 | FFPE | IIIc | G3 | 2010-04-01 | 1 | 2011-03-23 | Pelvic cavity | 8 | PS |
PFS, progression-free survival (between the last chemotherapy and the date of secondary surgery); SF, snap-frozen; FFPE, formalin-fixed, paraffin-embedded; PS, platinum-sensitive recurrence; PS, platinum-resistant recurrence.
miRNA expression profiles associated with recurrence in advanced HGSC
Eight pairs of primary-recurrent samples from eight patients were analyzed, with four normal ovarian tissue samples used as controls (Figure 2). miRNA expression profiles were compared between primary ovarian cancer tissue and normal ovarian tissue, recurrent ovarian cancer tissue and normal ovarian tissue, and recurrent ovarian cancer tissue and primary ovarian cancer tissue. In the comparison of primary ovarian HGSC with normal ovarian tissue, 57 miRNAs were significantly increased and 73 miRNAs were significantly decreased. These altered miRNA profiles are in agreement with previous findings [5]. In recurrent ovarian HGSC compared with normal ovarian tissue, 80 miRNAs were increased and 119 miRNAs were decreased. In recurrent ovarian HGSC compared with primary ovarian cancer tissue, 60 miRNAs, including miR-630, miR-370, and miR-575, were significantly increased and 52 miRNAs, including miR-509-3p, miR-514a-3p, and miR-506-3p, were decreased (Table 2). The top differentially expressed miRNAs (>2-fold change) in recurrent ovarian HGSC compared with primary HGSC are listed in Table 3. In addition, SF samples tended to cluster separately from FFPE samples, except for samples P3 and R3, in the heatmap (Figure 2).
Figure 2. MicroRNA microarray analysis.
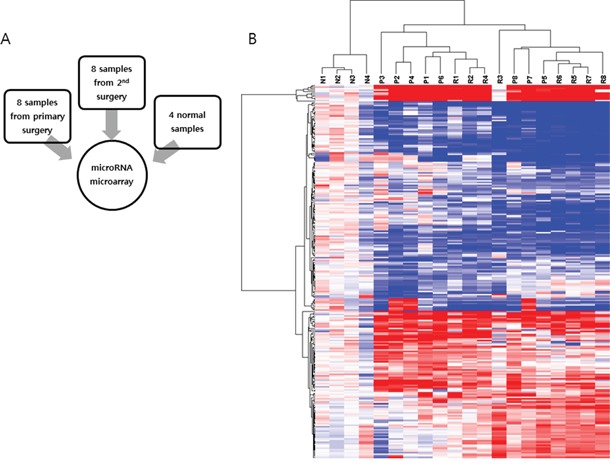
A. Eight pairs of primary-recurrent samples from eight patients were analyzed, and four normal ovarian tissue samples were used as a control. B. Significantly altered miRNAs are shown on a heatmap.
Table 2. Numbers of significantly increased and decreased miRNAs identified using miRNA microarrays.
| Significantly increased miRNAs | Significantly decreased miRNAs | |
|---|---|---|
| Primary ovarian HGSC vs. normal ovarian tissue | 57 | 73 |
| Recurrent ovarian HGSC vs. normal ovarian tissue | 80 | 119 |
| Recurrent ovarian HGSC vs. primary ovarian HGSC | 60 | 52 |
HGSC, high-grade serous carcinoma.
Table 3. Top differentially expressed miRNAs with >2-fold change in recurrent ovarian HGSC compared with primary ovarian tumor HGSC.
| Increased miRNAs | Fold change (log2 scale) |
Decreased miRNAs | Fold change (log2 scale) |
|---|---|---|---|
| hsa-miR-630 | 2.48039 | hsa-miR-509-3p | −3.6413 |
| hsa-miR-370 | 2.09488 | hsa-miR-514a-3p | −3.127 |
| hsa-miR-575 | 2.08748 | hsa-miR-506-3p | −2.4918 |
| hsa-miR-3692-5p | 1.72906 | hsa-miR-508-3p | −2.3845 |
| hsa-miR-4270 | 1.67729 | hsa-miR-509-3-5p | −2.3436 |
| hsa-miR-548q | 1.67019 | hsa-miR-95 | −2.3274 |
| hsa-miR-135a-3p | 1.63426 | hsa-miR-30a-3p | −2.2598 |
| hsa-miR-1207-5p | 1.62239 | hsa-miR-362-5p | −2.0035 |
| hsa-miR-4281 | 1.60895 | hsa-miR-30b-5p | −2.0014 |
| hsa-miR-671-5p | 1.59902 | hsa-miR-30e-3p | −1.9538 |
Similar profiles of differentially expressed miRNAs in primary and recurrent ovarian HGSC compared with normal ovarian tissue
Among the 31 miRNAs that were increased by more than 4-fold in primary tumors, 27 were also significantly increased in recurrent tumor samples. Likewise, among the 35 miRNAs that were decreased by more than 4-fold in primary tumors, 34 were also significantly decreased in recurrent tumor samples (Figure 3, Supplementary Tables S2 and S3). Overall, the differentially expressed miRNAs identified in primary HGSC and recurrent HGSC compared with normal ovarian tissue were largely consistent. Specifically, miR-200a, b, c, miR-141, and miR-429 were significantly increased in both primary and recurrent ovarian HGSC, and miR-125, miR-99a, and miR-145 were significantly decreased in both primary and recurrent ovarian HGSC. The expression profiles of miRNAs in primary ovarian HGSC compared with normal ovarian tissue significantly correlated with those in recurrent ovarian HGSC compared with normal ovarian tissue (correlation coefficient = 0.81, P = 0.0078). In addition, primary and recurrent ovarian HGSC samples were not separated in three-dimensional principal component analysis, although the normal ovarian specimens clustered together.
Figure 3. Characterization of miRNA expression.
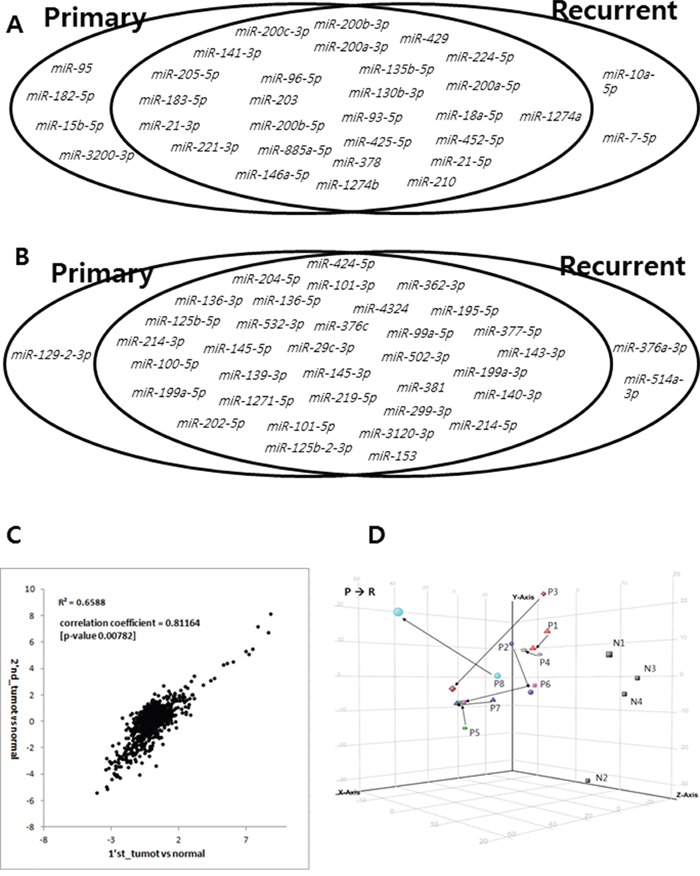
A. Among 31 microRNAs upregulated by more than 4-fold in primary tumors, 27 were also significantly upregulated in recurrent tumor samples. B. Likewise, among 35 microRNAs downregulated by more than 4-fold in primary tumors, 34 were also significantly downregulated in recurrent tumor samples. C. Expression profiles of miRNAs in primary ovarian HGSC compared with those of normal ovarian tissue were similar to those in recurrent ovarian HGSC compared with normal ovarian tissue (correlation coefficient = 0.811, P = 0.0078). D. Primary and recurrent ovarian HGSC samples were not separated in three-dimensional principal components analysis, although normal ovarian specimens clustered together.
Validation of miRNA results
Among the differentially expressed miRNAs, 5–6 were subjected to qRT-PCR analysis to validate the miRNA microarray results. We selected miR-141, miR-200c, miR-205, miR-429, miR-204, miR-506, and miR-509-3p, which were differentially expressed between normal ovarian tissue and ovarian HGSC (Figure 4). In agreement with the microarray results, the expression levels of miR-141, miR-200c, miR-205, and miR-429 were increased, whereas those of miR-204, miR-506, and miR-509-3p were decreased in HGSC. We selected miR-630, miR-370, miR-575, miR-506, and miR-509-3p, which were differentially expressed between recurrent and primary ovarian HGSC. For the selected miRNAs, the P-value and FDR were calculated and were found to be significant (<0.05, Supplementary Table S4). In agreement with the microarray results, the expression levels of miR-630, miR-370, and miR-575 were increased, whereas those of miR-506 and miR-509-3p were decreased in the recurrent samples. Thus, the microarray data were validated, warranting further miRNA analyses in a clinical setting.
Figure 4. Real-time RT-PCR verification of the miRNA microarray results.
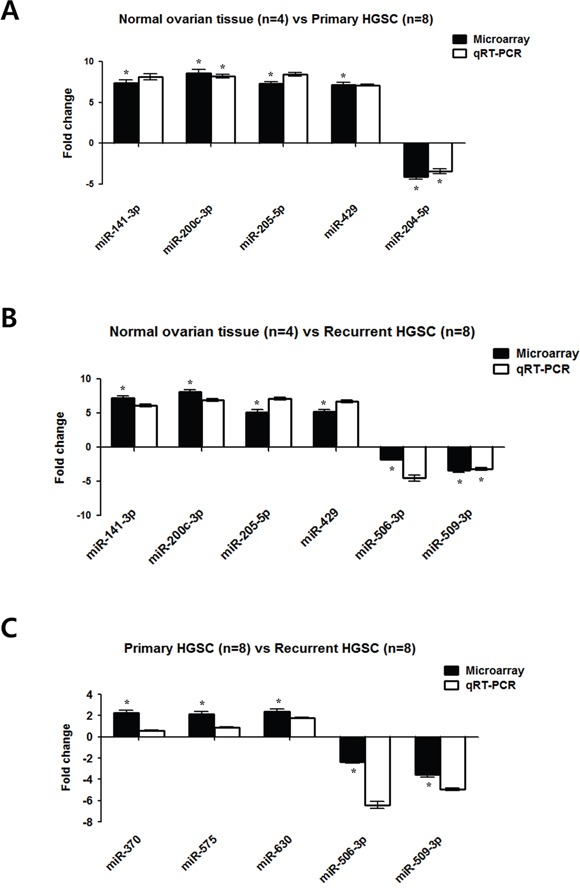
A. Differentially expressed miRNAs in primary ovarian HGSC (n = 8) compared with normal ovarian tissue (n = 4) from miRNA microarray experiments were consistent with data obtained using qRT-PCR. B. Differentially expressed miRNAs in recurrent ovarian HGSC (n = 8) compared with normal ovarian tissue (n = 4). C. Differentially expressed miRNAs in recurrent ovarian HGSC (n = 8) compared with primary HGSC (n = 8). Results are the mean ± standard deviation of three independent experiments performed in triplicate. (*P < 0.05).
DISCUSSION
In this study, we determined that the miRNA expression profiles in primary and recurrent ovarian HGSC were consistent (correlation coefficient = 0.81, P = 0.0078). Although some miRNAs were significantly increased or decreased in recurrent ovarian HGSC, most of the miRNAs matched with those of primary HGSC samples. To our knowledge, this is the first study to show a similarity of miRNA expression profiles between primary and recurrent ovarian HGSC.
A few previous studies explored miRNA expression profiles in recurrent ovarian HGSC compared with primary HGSC [10–13]. Laios et al. reported that miR-9 and miR-223 are potential biomarkers of recurrent epithelial ovarian cancer [11]. Chong et al. compared miRNA expression profiles in primary and recurrent ovarian serous carcinoma tissue [12]. However, primary and recurrent tumor specimens were not taken from the same patient in that study. Therefore, their resulting list of differentially expressed miRNAs was not consistent with our present data.
Recently, a whole-genome characterization of chemoresistant ovarian cancer tissue was published [13]. The authors showed that relapse samples had a higher mutational burden (single nucleotide variants and indels) at both the coding and non-coding levels than matched primary samples, suggesting that HGSC continues to evolve during treatment. In concordance with these findings, we found that recurrent tumors had a higher number of significantly increased or decreased miRNAs (Table 2).
The reason for the miRNA expression similarity between primary and recurrent HGSC has not yet been elucidated. We speculate that miRNAs are highly conserved throughout the species [14], and that miRNA profiles are not largely changed under the selective pressure of chemotherapy. Otherwise, HGSC may undergo clonal evolution under treatment, such that miRNA expression profiles of primary tumors could be conserved at recurrence.
Nevertheless, some miRNAs were significantly increased or decreased in recurrent HGSC compared with primary HGSC. The most significantly increased miRNA in recurrent HGSC observed was miR-630. Increased expression of miR-630 in gastric cancer and colorectal cancer has been reported to be associated with poor survival [15, 16]. A proposed mechanism is that miR-630 inhibits p53 activation by limiting the early DNA damage response [17]. miR-630 was found to be increased in non-small cell lung cancer cells and head and neck squamous cell carcinoma cells in response to cisplatin treatment [18]. Upregulation of miR-630 was also observed in epithelial ovarian cancer, and downregulation of miR-630 using an inhibitor improved the chemosensitivity of ovarian cancer cells to cisplatin [19]. Thus, further functional validation of miR-630 should be performed.
In the present study, we used different tissue types (SF and FFPE) for miRNA expression analyses. Previous miRNA expression studies have typically used FFPE specimens, and have shown that the miRNAs remain largely intact in routinely processed FFPE clinical specimens [20–22]. In agreement with previous studies, we also demonstrated strong correlations between expression profiles in matched SF and FFPE specimens. In addition, miRNA expression profiles obtained using SF + FFPE specimens in primary ovarian HGSC compared with normal ovarian tissue were consistent with previous results obtained using SF specimens used for miRNA analysis of primary ovarian HGSC [5]. However, the SF samples tended to cluster separately from FFPE samples, except for P3 and R3, in the heatmap (Figure 3). These findings indicate that pooling different tissue types together could affect the miRNA expression profiling results.
A limitation of our study is that we did not perform correlation analyses between miRNA expression profiles and clinicopathologic factors owing to the limited number of cases. With the exception of one patient, the patients included in this study had chemosensitive recurrence at secondary surgery. Secondary cyto-debulking operations are usually considered in only platinum-sensitive recurrent cases. In addition, recurrent HGSC specimens usually have extensive fibrosis. Although we used samples with a high tumor percentage, the presence of fibrosis may have led to confounding miRNA expression results.
In conclusion, primary and recurrent ovarian HGSC displayed similar miRNA expression patterns. Nevertheless, some altered miRNAs could be implicated in the recurrence of ovarian HGSC, and therefore further studies are required to determine the functional role of differentially expressed miRNAs in recurrent ovarian HGSC.
MATERIALS AND METHODS
Patients and study design
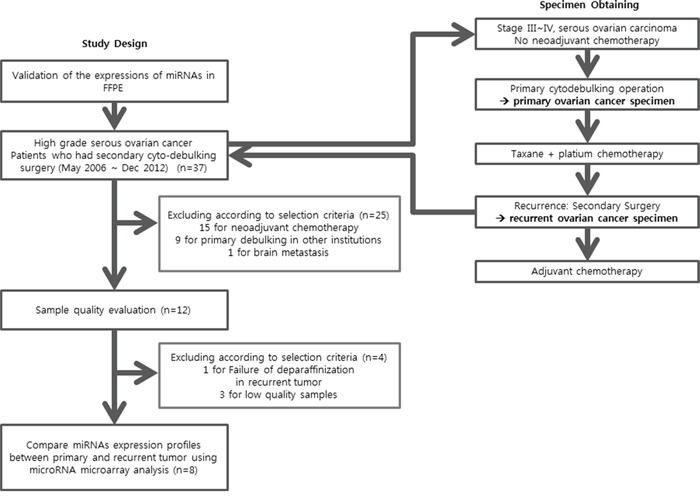
The experimental protocol was approved by the Institutional Review Board of the Yonsei University College of Medicine (4-2012-0741). Between May 2006 and December 2012, 37 ovarian HGSC patients underwent secondary debulking surgery at recurrence (Figure 5). Among them, 15 patients received neo-adjuvant chemotherapy before primary surgery was excluded. Nine patients that underwent primary surgery at another institution were excluded because primary tumor tissues were not available. Finally, three patients were excluded because of the low quality of samples, and one patient was excluded owing to failure of deparaffinization. Therefore, for the final analysis, eight pairs of primary-recurrent tumor tissues from eight patients were used for miRNA microarray analysis. Snap-frozen fresh (SF) tissues stored in liquid nitrogen and archival formalin-fixed, paraffin-embedded (FFPE) tissues were used for miRNA expression analyses. This strategy was used because ovarian tumor samples with both primary and corresponding recurrent samples preserved as SF tissues were limited. Therefore, we initially validated the miRNA expression profiles of FFPE-preserved tissues compared with those of SF-preserved tissues. Four SF normal ovarian tissue samples from patients treated for benign uterine disease were used as a control; the age of the control patients ranged from 49 to 60 years. For primary HGSC, FFPE tissues were sampled at the point where the tumor percentage was above 90% based on hematoxylin and eosin (H&E) staining, and 10-μm serial sectioning was performed. In recurrent HGSC, the tumor density is relatively low and tumors are surrounded by stromal cells. Therefore, core tissue (2 mm) was obtained using a hollow needle at the point where the tumor percentage was above 90% based on H&E staining, as performed for the tissue microarrays.
Figure 5. Study design and patient selection.
RNA preparation
Frozen tissue samples were homogenized in TRIzol® reagent (Invitrogen, Glasgow, UK) using an Omni-Mixer Homogenizer (Omni International, Kennesaw, GA, USA). Total RNA was then isolated according to the manufacturer's instructions. FFPE specimens were deparaffinized in xylene (tissue slides, 3 min; tissue core, overnight), and total RNA isolation was performed using a QIAamp RNA Mini Kit (Qiagen, Hilden, Germany). RNA quantity was assessed using a Nanodrop 1000A spectrophotometer (Nanodrop Technologies, Wilmington, DE, USA), and RNA quality was assessed using an Agilent 2100 Bioanalyzer and RNA 6000 Nano Chip (Agilent Technologies, Amstelveen, the Netherlands).
miRNA microarray analysis
Samples of total RNA (100 ng) containing miRNA were labeled with cyanine 3-pGp (Cy3) using an Agilent miRNA Complete Labeling and Hyb Kit (Agilent Technologies, Santa Clara, CA, USA). Samples were placed on an Agilent Human miRNA v14 chip (AMDID 029297) and covered with a Gasket slide (Agilent Technologies, Santa Clara, CA, USA). Slides were hybridized for 16 h at 42°C using an Agilent hybridization system. miRNA arrays were analyzed using GeneSpring GX v11.5 (Agilent Technologies, Santa Clara, CA, USA) with standard normalization methods for one-channel microarrays with percentile median normalization methods. Fold-change values were calculated for unpaired comparisons between normal and test samples, and then averaged to generate a mean fold-change value. Welch's t-tests were used to determine statistical significance. Target predictions of significantly modified miRNAs were analyzed using the TargetScan 5.1 and miRBase v16 databases. miRNA microarray data from this study have been submitted to the Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/info/geo_agil.html) under accession number GSE83693.
Reverse transcription and quantitative real-time reverse transcription-polymerase chain reaction (qRT-PCR)
To validate results of the miRNA microarray experiments, the five most differentially expressed miRNAs were further evaluated using qRT-PCR. Total miRNA was isolated using a mirVANA microRNA Isolation Kit (Ambion, Austin, TX, USA) according to the manufacturer's protocol. Synthesis of cDNA was performed using TaqMan MicroRNA Assays (Applied Biosystems, Foster City, CA, USA). qRT-PCR was performed using an Applied Biosystems Prism 7500 Fast Sequence Detection System (Applied Biosystems, Warrington, UK) according to the manufacturer's protocol. Relative gene expression was analyzed using the 2−ΔΔCT method, and results are expressed as the fold-change relative to control values. All experiments were repeated three times.
Statistical analysis
To ensure the accuracy of microarray data analysis, false discovery rate (FDR)-adjusted P-values (q-values) were calculated using the methods reported by Storey [23]. The qRT-PCR data are expressed as the mean ± standard deviation of at least three independent experiments. Significance was determined using the Student's t-test. Comparison of miRNA expression profiles was performed using Pearson correlation. Statistical analyses were performed using SPSS (Version 18.0, SPSS, Inc., USA).
SUPPLEMENTARY TABLES
Acknowledgments
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (2014R1A1A1A05002926 and 2015R1A2A2A01008162).
Footnotes
CONFLICTS OF INTEREST
The authors declare that there are no conflicts of interest.
REFERENCES
- 1.Bookman MA. Should studies of maintenance therapy be maintained in women with ovarian cancer? J Gynecol Oncol. 2013;24:105–107. doi: 10.3802/jgo.2013.24.2.105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Jemal A, Siegel R, Xu J, Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010;60:277–300. doi: 10.3322/caac.20073. [DOI] [PubMed] [Google Scholar]
- 3.Hannon GJ. RNA interference. Nature. 2002;418:244–251. doi: 10.1038/418244a. [DOI] [PubMed] [Google Scholar]
- 4.Ambros V. MicroRNA pathways in flies and worms: growth, death, fat, stress, and timing. Cell. 2003;113:673–676. doi: 10.1016/s0092-8674(03)00428-8. [DOI] [PubMed] [Google Scholar]
- 5.Nam EJ, Yoon H, Kim SW, Kim H, Kim YT, Kim JH, Kim JW, Kim S. MicroRNA expression profiles in serous ovarian carcinoma. Clin Cancer Res. 2008;14:2690–2695. doi: 10.1158/1078-0432.CCR-07-1731. [DOI] [PubMed] [Google Scholar]
- 6.Iorio MV, Visone R, Di Leva G, Donati V, Petrocca F, Casalini P, Taccioli C, Volinia S, Liu CG, Alder H, Calin GA, Menard S, Croce CM. MicroRNA signatures in human ovarian cancer. Cancer Res. 2007;67:8699–8707. doi: 10.1158/0008-5472.CAN-07-1936. [DOI] [PubMed] [Google Scholar]
- 7.Cancer Genome Atlas Research N. Integrated genomic analyses of ovarian carcinoma. Nature. 2011;474:609–615. doi: 10.1038/nature10166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ross JS, Ali SM, Wang K, Palmer G, Yelensky R, Lipson D, Miller VA, Zajchowski D, Shawver LK, Stephens PJ. Comprehensive genomic profiling of epithelial ovarian cancer by next generation sequencing-based diagnostic assay reveals new routes to targeted therapies. Gynecol Oncol. 2013 doi: 10.1016/j.ygyno.2013.06.019. [DOI] [PubMed] [Google Scholar]
- 9.Boren T, Xiong Y, Hakam A, Wenham R, Apte S, Chan G, Kamath SG, Chen DT, Dressman H, Lancaster JM. MicroRNAs and their target messenger RNAs associated with ovarian cancer response to chemotherapy. Gynecol Oncol. 2009;113:249–255. doi: 10.1016/j.ygyno.2009.01.014. [DOI] [PubMed] [Google Scholar]
- 10.Gallagher MF, Heffron CC, Laios A, O'Toole SA, Ffrench B, Smyth PC, Flavin RJ, Elbaruni SA, Spillane CD, Martin CM, Sheils OM, O'Leary JJ. Suppression of cancer stemness p21-regulating mRNA and microRNA signatures in recurrent ovarian cancer patient samples. J Ovarian Res. 2012;5:2. doi: 10.1186/1757-2215-5-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Laios A, O'Toole S, Flavin R, Martin C, Kelly L, Ring M, Finn SP, Barrett C, Loda M, Gleeson N, D'Arcy T, McGuinness E, Sheils O, Sheppard B, J OL. Potential role of miR-9 and miR-223 in recurrent ovarian cancer. Mol Cancer. 2008;7:35. doi: 10.1186/1476-4598-7-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chong GO, Jeon HS, Han HS, Son JW, Lee YH, Hong DG, Lee YS, Cho YL. Differential MicroRNA Expression Profiles in Primary and Recurrent Epithelial Ovarian Cancer. Anticancer Res. 2015;35:2611–2617. [PubMed] [Google Scholar]
- 13.Patch AM, Christie EL, Etemadmoghadam D, Garsed DW, George J, Fereday S, Nones K, Cowin P, Alsop K, Bailey PJ, Kassahn KS, Newell F, Quinn MC, et al. Whole-genome characterization of chemoresistant ovarian cancer. Nature. 2015;521:489–494. doi: 10.1038/nature14410. [DOI] [PubMed] [Google Scholar]
- 14.Voinnet O. Origin, biogenesis, and activity of plant microRNAs. Cell. 2009;136:669–687. doi: 10.1016/j.cell.2009.01.046. [DOI] [PubMed] [Google Scholar]
- 15.Chu D, Zhao Z, Li Y, Li J, Zheng J, Wang W, Zhao Q, Ji G. Increased microRNA-630 expression in gastric cancer is associated with poor overall survival. PLoS One. 2014;9:e90526. doi: 10.1371/journal.pone.0090526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chu D, Zheng J, Li J, Li Y, Zhang J, Zhao Q, Wang W, Ji G. MicroRNA-630 is a prognostic marker for patients with colorectal cancer. Tumour Biol. 2014;35:9787–9792. doi: 10.1007/s13277-014-2223-3. [DOI] [PubMed] [Google Scholar]
- 17.Galluzzi L, Morselli E, Vitale I, Kepp O, Senovilla L, Criollo A, Servant N, Paccard C, Hupe P, Robert T, Ripoche H, Lazar V, Harel-Bellan A, et al. miR-181a and miR-630 regulate cisplatin-induced cancer cell death. Cancer Res. 2010;70:1793–1803. doi: 10.1158/0008-5472.CAN-09-3112. [DOI] [PubMed] [Google Scholar]
- 18.Huang Y, Chuang A, Hao H, Talbot C, Sen T, Trink B, Sidransky D, Ratovitski E. Phospho-DeltaNp63alpha is a key regulator of the cisplatin-induced microRNAome in cancer cells. Cell Death Differ. 2011;18:1220–1230. doi: 10.1038/cdd.2010.188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zou YT, Gao JY, Wang HL, Wang Y, Wang H, Li PL. Downregulation of microRNA-630 inhibits cell proliferation and invasion and enhances chemosensitivity in human ovarian carcinoma. Genet Mol Res. 2015;14:8766–8777. doi: 10.4238/2015.July.31.25. [DOI] [PubMed] [Google Scholar]
- 20.Hasemeier B, Christgen M, Kreipe H, Lehmann U. Reliable microRNA profiling in routinely processed formalin-fixed paraffin-embedded breast cancer specimens using fluorescence labelled bead technology. BMC Biotechnol. 2008;8:90. doi: 10.1186/1472-6750-8-90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Xi Y, Nakajima G, Gavin E, Morris CG, Kudo K, Hayashi K, Ju J. Systematic analysis of microRNA expression of RNA extracted from fresh frozen and formalin-fixed paraffin-embedded samples. RNA. 2007;13:1668–1674. doi: 10.1261/rna.642907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zhang X, Chen J, Radcliffe T, Lebrun DP, Tron VA, Feilotter H. An array-based analysis of microRNA expression comparing matched frozen and formalin-fixed paraffin-embedded human tissue samples. J Mol Diagn. 2008;10:513–519. doi: 10.2353/jmoldx.2008.080077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Owzar K, Barry WT, Jung SH. Statistical considerations for analysis of microarray experiments. Clin Transl Sci. 2011;4:466–477. doi: 10.1111/j.1752-8062.2011.00309.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.