

Abstract
In vitro culture of ovarian granulosa cells and theca cells has been very important for our understanding of their function and regulation. One of the most eagerly sought attributes of cell culture is the use of chemically-defined conditions. However, even under such in vitro conditions cell behaviour could differ from the in vivo situation because of differences in oxygen tension, nutrients, adhesion matrix and other factors. To examine this further we compared the transcriptomes of both granulosa cells and cells from the theca interna that were cultured in what are arguably the best in vitro conditions for maintaining the ‘follicular’ phenotypes of both tissue types, as displayed by their respective freshly-isolated counterparts. The array data analysed are from recently published data and use the same sizes of bovine follicles (small antral 3–6 mm) and the same Affymetrix arrays. We conducted analysis using Partek, Ingenuity Pathway Analysis and GOEAST. Principal Component Analysis (PCA) and hierarchical clustering clearly separated the in vivo from the in vitro groups for both cells types and transcriptomes were more homogeneous upon culture. In both cell cultures behaviours associated with cell adhesion, migration and interaction with matrix or substrate were more abundant. However, the pathways involved generally differed between the two cell types. With the thecal cultures a gene expression signature of an immune response was more abundant, probably by leukocytes amongst the cells cultured from the theca interna. These results indicate differences between in vivo and in vitro that should be considered when interpreting in vitro data.
Background
In ovaries oocytes develop within follicles which at the initial primordial stage are composed of an inactive oocyte surrounded by a quiescent population of epithelial granulosa cells. A number of primordial follicles are activated daily and subsequently the granulosa cells begin to divide and, over a period of months in bovine ovaries, increase to about 50 million cells [1]. At the same time as the follicle expands it develops an antral cavity, filled with follicular fluid. Growth of follicles is important for expanding the number of granulosa cells to ensure that enough of the steroid hormone oestradiol is secreted to control and regulate the reproductive cycle. At about the time an antrum forms, the stroma surrounding the membrana granulosa specialises into the theca interna and externa layers. Specialised cells in the vascularised theca interna are steroidogenic and secrete androgens, such as androstenedione and testosterone, which are converted by granulosa cells into oestradiol. Differentiation of the theca interna is therefore integral for oestrogen synthesis.
During early stages of growth, the granulosa cells express receptors for follicle-stimulating hormone (FSH), which is secreted from the anterior pituitary gland and stimulates follicle growth. Very late in follicular development the granulosa cells also express receptors for luteinising hormone (LH) and large follicles which have granulosa cells expressing these receptors are capable of ovulating in response to a surge release of LH from the anterior pituitary. The steroidogenic cells of the theca, on the other hand, express LH receptors from an early stage and respond to LH by synthesising androgens. Both the granulosa cells and the theca interna cells are key somatic cell types whose function and regulation are pivotal to follicle development, steroidogenesis and female fertility.
The development of in vitro culture systems for both granulosa cells and theca cells has been very important for our understanding of their function and regulation within the context of follicle development and ovarian steroidogenesis. Earlier studies utilising culture media supplemented with fetal calf serum [2] showed that granulosa cells have a propensity to differentiate terminally into luteal cells, as occurs upon ovulation in vivo. Anchorage-independent culture was used to examine the stem cell population of granulosa cells [3,4] and this was followed by a serum free system [5], in which granulosa cells retained the ability to produce oestradiol. In vivo, this feature of granulosa cells is seen during the final stages of antral follicle maturation and particularly in large follicles approaching ovulatory size [6]. An important property of thecal cells in culture is their ability to produce androgens and retain responsiveness to LH [7]. These properties are lost when cells are cultured in serum-containing media and the cells undergo changes characteristic of luteinisation.
One of the most eagerly sought attributes of cell culture is the use of chemically-defined conditions where the use of fetal calf serum or other biological reagents that might vary from batch to batch is avoided. However, even such in vitro conditions could differ markedly from in vivo conditions because of differences in oxygen tension, nutrients and a myriad of other factors. In vitro, cell behaviour could also be affected by the sporadic changes of cell culture media that is likely to produce acute temporal fluctuations in the concentrations of nutrients and metabolites during culture. Additionally, interactions with cell culture plastic and altered cell-cell interactions could occur in vitro. Even just the process of harvesting cells and transferring them into culture could alter their phenotype in vitro. Therefore, it would be expected that differences exist between the in vivo and in vitro conditions. However, most in vitro studies on ovarian granulosa and theca interna cells have been confined to examination of a limited set of targeted parameters like steroidogenesis and expression of known receptors and growth factors.
To examine the effects of culture further we compared the transcriptomes of both granulosa cells and cells from the theca interna that were cultured in what are arguably regarded as the most physiologically relevant culture conditions for maintaining ‘follicular’ (i.e. non-luteinized) phenotypes in vitro of both cell types [5,8–10] comparable with those of their respective freshly-isolated (in vivo) counterparts. The microarray data are from recently published reports from our groups in which comparisons were made within in vivo [granulosa cells [11], thecal cells [12]] and within in vitro [granulosa cells [13], theca interna [14]] conditions but not between in vivo and in vitro conditions. These previous studies whilst conducted in different locations used the same sizes of bovine follicles (small antral 3–6 mm) and the same Affymetrix arrays. During culture, steroid hormone production was monitored and confirmed that both the granulosa and thecal cells maintained their follicular phenotypes [13,14].
Materials and methods
Tissues and cells, RNA extraction and array hybridisation
In vivo studies: Granulosa cells and intact theca interna were isolated from two different groups of individual small healthy follicles (n = 10 each group, 3–5 mm in diameter) obtained from an abattoir according to previously described methods [15,16]. The health status of the follicles was confirmed by histological examination of a portion of the follicle wall based on the number of apoptotic figures observed in the membrana granulosa [17,18]. RNA was extracted from granulosa cells and theca interna of each follicle preparation and two μg of RNA per sample was processed for hybridisation to a Bovine Affymetrix Genome Array (Australian Genomics Research Facility, Parkville and ACRF Cancer Genomics facility, Adelaide) whereby ten arrays each of either granulosa cells or theca interna were available for analysis as was previously published by our groups [11,12]. The CEL files can be obtained at GSE39589 and GSE49505 for granulosa and theca data, respectively.
In vitro studies: Two separate experiments utilised granulosa cells and cells of the theca interna each isolated from bovine ovarian follicles 4–6 mm in diameter obtained from an abattoir as previously described [8–10]. For granulosa cell isolation [10] medium-sized follicles (4- to 6-mm diameter) lacking obvious signs of atresia were removed, follicular fluid was aspirated, a slit was made in the follicle wall and the granulosa cell layer was gently disrupted with the aid of a plastic inoculation loop. Double-distilled water (10 ml) was added and the cells were agitated for 10 secs to lyse any red blood cells present; isotonicity was quickly restored by the addition of 10 ml of 3× concentrated PBS. For thecal cell isolation [9] follicles were isolated and granulosa cells removed. Follicle halves were then shaken vigorously to remove any remaining granulosa cells, and the medium was changed three times. Follicle halves were examined under a dissecting microscope, and the theca interna layer was gently peeled away from the basement membrane. Pooled theca layers were incubated with collagenase (type IV, 1 mg/ml; Sigma Ltd., Poole, UK) and trypsin inhibitor (100 μg/ml; Sigma). After 30 min, the cell layers were triturated with a Pasteur pipette and returned to the water bath for another 15 min. Finally, any remaining undigested material was allowed to settle, and the resulting theca-cell-rich supernatant decanted. In each of 4 independent replicate cultures, pooled granulosa or theca cells from approximately 50 follicles were seeded at 5 x 105/ml in 24 well plates and incubated for six days with hormone /growth factor treatments on days 4 to 6 of culture in McCoys 5A medium supplemented with antibiotics, 0.1% bovine serum albumin and other growth supplements as detailed previously [13,14]. The granulosa cells were cultured under four conditions: FSH only (0.33 ng/ml), tumour necrosis factor α (TNFα) only (10 ng/ml), FSH plus TNFα or no treatment (control) [13], but only control and FSH treated cells were analysed here. The theca cells were also subjected to four treatments: LH only (160 pg/ml), bone morphogenic protein-6 (BMP-6) only (10 ng/ml), LH plus BMP-6 or control [14], but only control and LH treated cells were analysed here. In both experiments the production of steroid hormones was measured to ensure the cells retained their follicular phenotype [13,14]. Each independent experiment used n = 4 wells for each condition and two μg of RNA from pooled extracts was used to hybridise to a Bovine Affymetrix Genome Array for analysis of global gene expression (Almac Diagnostics Limited, Craigavon, UK) forming the basis of recently published studies [13,14]. The CEL files can be obtained at GSE42535 and GSE44704 for granulosa and theca data, respectively.
Array analyses
Affymetrix CEL file data were pre-processed in Partek Genomics Suite (version beta 6.6; St Louis, Missouri, USA) using RMA background summarisation, with quantile normalisation and log base 2 transformation and mean probe set summarisation with adjustment for GC content. All arrays passed the spike-in hybridisation quality controls as conducted previously [11].
Clustering and statistical analyses
Partek Genomics Suite was used for clustering and statistical analysis of the array data. Initially, PCA and hierarchical clustering were conducted separately on granulosa (n = 17) and thecal cell arrays (n = 18) according to parameters previously published [11,15]. The hormone-treated (either FSH or LH, for granulosa and thecal cell, respectively) and control cultured arrays were analysed collectively as the single group in vitro due to their similar transcriptional profiles [13,14]. Comparison of the in vitro and in vivo groups for both the granulosa cell and theca interna arrays was performed by one-way ANOVA with followed by Benjamini-Hochberg false discovery rate corrections for multiple testing.
Ingenuity Pathway Analyses (IPA) and GOEAST snalyses
Canonical pathway analysis was conducted in IPA and Gene Ontology enrichment analysis in GOEAST (Gene Ontology Enrichment Analysis Software Toolkit) as previously described and using statistically significantly differentially-expressed genes identified by ANOVA with followed by Benjamini-Hochberg false discovery rate corrections for multiple testing [11]. Regulator effect network analysis was also conducted in IPA whereby possible upstream regulators determined from the Ingenuity knowledge base from our differentially expressed data sets were matched with predicted downstream processes using Fishers exact t-test and an algorithm which computes the consistency of these interactions [19]. Only the highest ranked networks are presented for each analysed data set.
Results
Behaviour of granulosa cells and theca interna cells during in vitro culture
Dispersed granulosa cells and theca interna cells re-aggregated into multicellular clumps during the course of the 6-day culture periods (Fig 1) indicating cell-cell or cell-extracellular matrix interactions as seen previously [20].
Fig 1. Appearance of Granulosa Cells (GC) and Thecal Cells (TC) cultured under serum-free conditions for 48, 96 and 144 hours.

Comparisons of in vivo and in vitro granulosa cell arrays
The expression of the housekeeping genes was similar between the in vivo and in vitro situations for granulosa cells with the mean probe intensity of the housekeeping genes GAPDH, ACTB, PPIA and MRPL32 on the in vitro chips being 1.229, 1.041, 0.875 and 1.385-fold of that of the in vivo chips, respectively.
Principal component analysis (PCA) (Fig 2A) and hierarchical clustering (S1 Fig) were applied to in vitro and in vivo granulosa cell array data. These two methods separated the two groups from each other with a greater spread evident in the in vivo group. The differentially expressed genes are listed in Tables 1 and 2. Genes that are known to be specifically highly expressed in granulosa cells from antral follicles such as CYP19A1, FSHR and AMH were found in lower abundance in vitro (Table 1). In contrast, many of the genes which more abundant in vitro (Table 2), including CTGF, COL1A2, COL5A2, DCN and BMP2 are related to extracellular matrix production and hence tend to fit a mesenchymal tissue expression profile.
Fig 2.
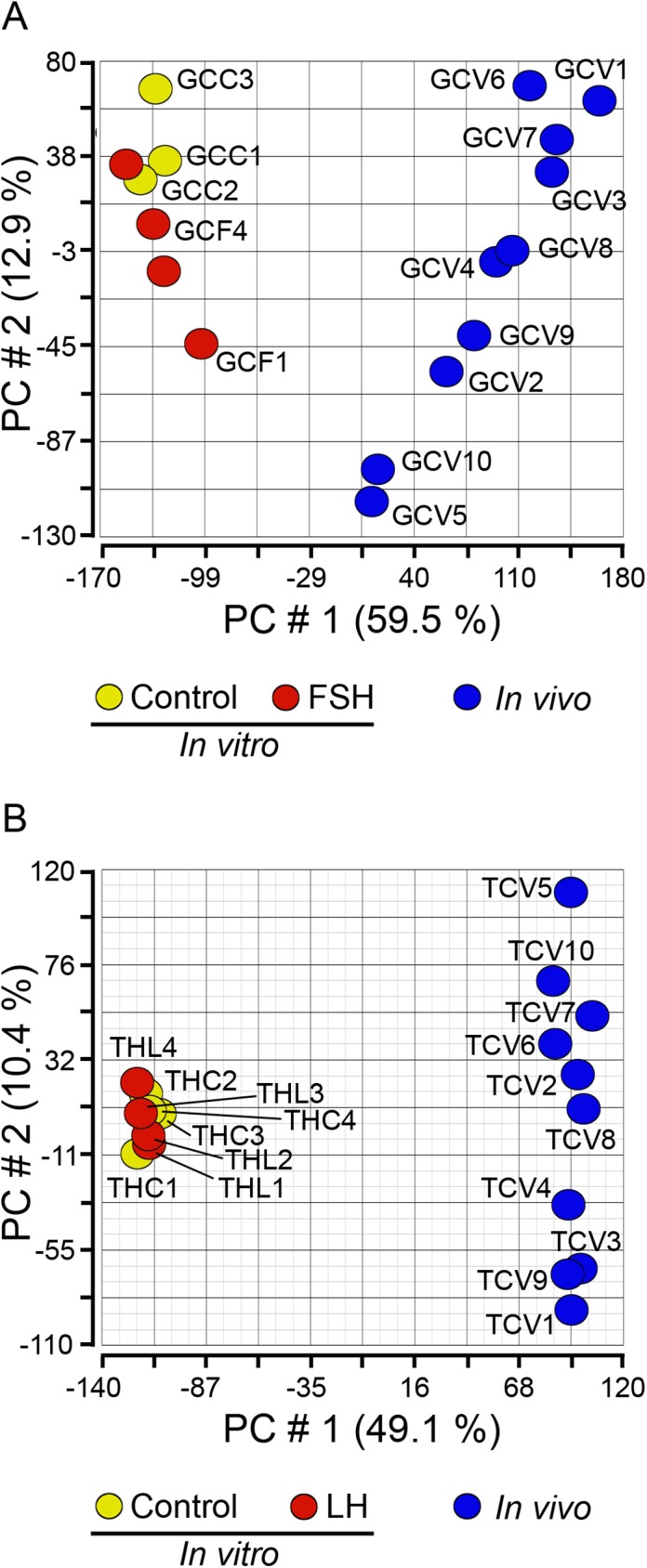
Unsupervised PCA of granulosa cell (A) and theca interna (B) arrays. The graphs are scatter plots of the values for the first (X) and second (Y) principal components based on the correlation matrix of the total normalised array intensity data. The numbering of each sample enables the samples in the figure to be identified in S1 and S2 Figs. Abbreviations are GC (granulosa cell), TH (theca interna), CC (cultured under control conditions), CF (cultured and FSH treated), CL (cultured and LH treated) and V (in vivo freshly isolated cells).
Table 1. Top 50 genes that were most differentially up regulated in vitro in granulosa cells.
| Gene Symbol | Fold-Change | Gene Symbol | Fold-Change |
|---|---|---|---|
| TNFAIP6 | 316.1 | S100A12 | 29.8 |
| XCL1 | 145.2 | NUAK1 | 28.3 |
| STC1 | 102.7 | COL5A2 | 27.5 |
| ANKRD1 | 102.1 | BMP2 | 26.2 |
| SFRP2 | 97.1 | CLCA3 | 26.1 |
| PLAT | 79.4 | SGK1 | 24.9 |
| CXCL6 | 67.1 | CTSH | 24.7 |
| DCN | 66.7 | GFPT2 | 24.7 |
| STAR | 62.8 | PLK2 | 24.3 |
| PDGFRA | 57.8 | TIMP1 | 23.5 |
| PTX3 | 55.1 | CXCL2 | 22.8 |
| IL18 | 52.6 | NCAM1 | 22.5 |
| DKK3 | 50.1 | AXL | 22.4 |
| SCG2 | 49.0 | GIMAP8 /// LOC100847526 | 21.0 |
| NID2 | 49.0 | FN1 | 20.9 |
| CLDN11 | 43.4 | CAV1 | 20.8 |
| RND3 | 39.1 | ANXA1 | 19.3 |
| THBS2 | 38.6 | OLR1 | 19.3 |
| TNFRSF12A | 38.2 | COL3A1 | 19.2 |
| LGALS3 | 35.6 | TRIB1 | 18.5 |
| MMP9 | 34.9 | IGFBP5 | 18.5 |
| CTGF | 33.2 | MAP1LC3C | 18.4 |
| COL1A2 | 32.8 | NDP | 17.8 |
| PRKAG2 | 31.0 | DCLK1 | 17.6 |
| LUM | 30.5 | POSTN | 17.4 |
Table 2. Top 50 genes that were most differentially down regulated in vitro in granulosa cells.
| Gene Symbol | Fold-Change | Gene Symbol | Fold-Change |
|---|---|---|---|
| IHH | 26.8 | LAMC2 | 6.8 |
| CARTPT | 20.0 | NALCN | 6.7 |
| MEST | 18.9 | CHRDL1 | 6.5 |
| HSD17B1 | 15.5 | LTF | 6.4 |
| ETNK2 | 13.0 | PLEK | 6.3 |
| PTI | 12.8 | SVOPL | 6.2 |
| GUCA1A | 12.6 | CRISPLD2 | 6.0 |
| PLA2G1B | 12.6 | CA14 | 6.0 |
| GPC3 | 12.2 | KIFC1 | 6.0 |
| SLC27A3 | 10.0 | STAC3 | 6.0 |
| SEP4 | 10.0 | SLC10A2 | 5.8 |
| CYP19A1 | 9.3 | NOS2 | 5.8 |
| FSHR | 9.1 | HAUS4 | 5.7 |
| HSPA1A | 9.0 | JAKMIP1 | 5.7 |
| MYH11 | 8.8 | PABPN1 | 5.5 |
| ANGPT2 | 8.8 | RAC3 | 5.5 |
| SLC22A3 | 8.7 | MOB3B | 5.4 |
| GANAB | 8.6 | AQP1 | 5.4 |
| GYLTL1B | 8.0 | PGF | 5.2 |
| ABCC8 | 7.8 | AOAH | 5.2 |
| AMH | 7.7 | TOP2A | 5.2 |
| ACSS1 | 7.3 | PKDCC | 5.1 |
| GPT | 7.2 | IQGAP2 | 5.1 |
| NUP210 | 7.2 | EFHD1 | 5.1 |
| CCBL1 | 7.0 | SORL1 | 5.1 |
A subset of differentially regulated genes (4-fold change, FDR P < 0.05) was uploaded to the Ingenuity database and GOEAST for network, pathway and function analyses (Fig 3). This subset (n = 604 genes) is listed in S1 Table. Ingenuity Pathway Analyses and GOEAST analyses identified that cell adhesion and extracellular matrix related pathways were up regulated in vitro. Regulator effect network analysis (Fig 4) identified many effects on cell adhesion, movement, binding, migration and chemo-attraction and macrophage functions including phagocytosis. The upstream regulators that were predicted to be switched on included SREB1, HMGB1, FOS, IRF2, IRF5, JUNB, ID3 and IFI16 those predicted to be switched off included NR1H2, SIRT1, NR3C1 and MYC.
Fig 3.

Top canonical pathways in granulosa cells mapped in IPA (A) and GO terms (B) classified under biological process. In (A) the bar chart on the left represents the percentage of genes from the data set that map to each canonical pathway showing those which are up regulated (in red) and down regulated (in blue) in vitro with respect to in vivo. The line chart on the right ranks these pathways derived for the same data set, from the highest to lowest degree of association based on multiple correction testing for the Benjamini-Hochberg False Discovery Rate. In (B) the bar chart on the left represents the percentage of genes from the data set that map to each GO term showing those which are differentially regulated (in blue) in vitro with respect to in vivo. The line chart on the right ranks these pathways derived for the same data set, from the highest to lowest degree of association using the Benjamini-Yuketeli test for multiple corrections (bottom to top in graphs on right).
Fig 4. Regulator effect network analysis in IPA identifying altered regulators and networks in granulosa cells cultured in vitro.

Orange and blue are predicted up and down regulated regulators on top, and the pink and green represent up and down regulated genes from the array data with the intensity of colour reflecting the degree of change. The pathways altered are listed at the bottom and orange indicates they are up regulated in vitro.
To compare transcriptional processes of granulosa cells in vitro under specific conditions with those known to occur naturally in vivo such as atresia or maturation prior to ovulation, we decided to compare the differentially regulated dataset between in vitro and in vivo granulosa cells with the small atretic versus healthy follicle granulosa gene set [11], and with the small healthy follicle versus large healthy follicle granulosa gene set [15], from previous studies. S2A Fig shows that for granulosa cells 39.8% and 13.9% of genes differentially expressed (4-fold different between groups with FDR P <0.05) between in vivo and in vitro conditions were also differentially expressed in atresia and upon growth to large follicle sizes, respectively, indicating the majority of changes upon culture were not associated changes that would be observed with growth or atresia of follicles. We additionally checked to see if there was any potential thecal contamination that could have artefactually contributed to these results (S2B Fig). As only 17.5% of differentially expressed genes between the in vivo and in vitro (4-fold different between groups with FDR P < 0.05) were higher in the theca versus granulosa cells in the in vivo arrays and the thecal-specific markers CYP17 and INSL3 were not present in this group, any potential thecal contamination of granulosa cells is unlikely to have contributed to the results.
Comparisons of in vivo and in vitro theca cell arrays
The expression of the housekeeping genes was similar between the in vivo and in vitro situations for thecal cells with the mean probe intensity of the housekeeping genes GAPDH, ACTB, PPIA and MRPL32 on the in vitro chips being 1.254, 0.997, 0.714 and 0.881-fold of that of the in vivo chips, respectively.
PCA (Fig 2B) and hierarchical clustering (S3 Fig) were applied to the in vitro and in vivo thecal cell array data. These two methods separated the two groups from each other with a far greater spread amongst the in vivo group indicating greater heterogeneity in gene expression amongst the in vivo group. The differentially expressed genes are listed in Tables 3 and 4. There was a high proportion of genes related to immune function in the group up regulated in vitro including; those which encode the major histocompatibility complex (classes I, BOLA-N JSP.1, BOLA-A; class II; BOLA-DQA2, BOLA-DQA5, CD74); interleukins IL18, IL6 and IL8 and the inflammatory chemokines CXCL6, CXCL2, CCL5 and CCL3. Many other genes associated with immune responses were also more abundant, such as interferon gamma (S2 Table). Those genes which constitute the less abundant group in vitro were heterogeneous in function ranging from developmental differentiation for example SHISA2, ALDH1A2 and FRZB to extracellular matrix such as ASPN, COL16A1 and COL13A1.
Table 3. Top 50 genes that were most differentially up regulated in vitro in thecal cells.
| Gene Symbol | Fold-Change | Gene Symbol | Fold-Change |
|---|---|---|---|
| BOLA /// BOLA | 213.7 | IL6 | 23.6 |
| CFB | 123.4 | IL8 | 23.1 |
| NT5E | 59.4 | BoLA /// BOLA /// BOLA-A | 23.1 |
| IDO1 | 58.3 | TNC | 22.8 |
| SPP1 | 55.2 | TNFAIP6 | 21.6 |
| CXCL6 | 50.8 | ATF3 | 21.5 |
| CXCL2 | 49.9 | CHI3L1 /// LOC788983 | 21.4 |
| GBP5 | 45.3 | IRF1 | 21.0 |
| ANKRD1 | 43.2 | SERPINB2 | 20.3 |
| BoLA /// BOLA-A | 43.0 | MMP9 | 20.2 |
| IL18 | 40.8 | GRO1 | 19.5 |
| NUPR1 | 40.8 | C1R | 19.4 |
| C1S | 36.0 | CLDN1 | 18.6 |
| SAA3 | 34.6 | SRRM2 | 18.1 |
| CXCL10 | 32.6 | CD74 | 17.5 |
| GPNMB | 30.1 | CCL5 | 17.4 |
| OLR1 | 29.2 | POSTN | 17.1 |
| CCL2 | 27.9 | ARHGAP29 | 16.8 |
| BOLA-N /// JSP.1 | 27.8 | EFEMP1 | 16.8 |
| GBP4 /// LOC507055 | 27.2 | CCL3 | 16.2 |
| ERAP2 | 27.2 | ISG20 | 15.6 |
| MX2 | 27.1 | BOLA-DQA5 | 15.5 |
| CXCL11 | 25.1 | AP2B1 | 15.2 |
| WARS | 24.3 | BOLA-DQA2 | 14.7 |
| RASSF4 | 24.1 | EFR3A | 14.5 |
Table 4. Top 50 genes that were most differentially down regulated in vitro in thecal cells.
| Gene Symbol | Fold-Change | Gene Symbol | Fold-Change |
|---|---|---|---|
| CXCL14 | 76.5 | APLNR | 11.0 |
| SHISA2 | 57.2 | LMCD1 | 10.7 |
| DUSP12 | 26.0 | PRMT1 | 10.5 |
| COLEC11 | 20.3 | PTH1R | 10.4 |
| AQP1 | 17.9 | CCL14 | 10.1 |
| HMGB3 | 17.7 | PABPN1 | 10.0 |
| DHRS3 | 17.7 | C1QA | 9.9 |
| BRB | 17.4 | NALCN | 9.9 |
| GPC3 | 16.6 | CD34 | 9.8 |
| HPGD | 16.0 | TCF21 | 9.8 |
| HSD17B1 | 15.5 | COL13A1 | 9.6 |
| SDPR | 13.7 | HEYL | 9.1 |
| ASPN | 13.6 | AKAP8L | 8.6 |
| ALDH1A2 | 13.2 | ISLR | 8.6 |
| PLIN5 | 12.5 | AFAP1L1 | 8.5 |
| CLEC14A | 12.4 | GANAB | 8.4 |
| TMEM88 | 12.3 | MEOX2 | 8.4 |
| RGS5 | 12.2 | TM4SF18 | 8.4 |
| LRRC70 | 12.0 | PLXND1 | 8.2 |
| LAMA2 /// LOC100848461 | 11.9 | FAM64A | 8.2 |
| FRZB | 11.8 | HFM1 | 8.0 |
| COL16A1 /// LOC100849968 | 11.8 | RBM3 | 7.9 |
| AMH | 11.5 | CDKN1C | 7.6 |
| RNASE6 | 11.4 | TFF2 | 7.5 |
| EGFLAM /// LOC100847583 | 11.4 | C1QC | 7.5 |
A subset of differentially regulated genes (> 4-fold change, FDR P < 0.05) was uploaded to the Ingenuity database and GOEAST for network, pathway and function analyses (Fig 5). This subset (n = 610 genes) is listed in S2 Table. IPA and GOEAST analyses identified pathways related to immune response to be up regulated in vitro. Regulator effect network analysis (Fig 6) identified many effects on cell movement, adhesion, migration, homing, filopodia formation, extension of cell processes and chemo-attraction. The upstream regulators that were predicted to be switched on included STAT4, HIF1A, SNAI1, IRF3, SMAD3, EGR1, CREBP, NFKB2, FOXO4 and MYB.
Fig 5.

Top canonical pathways in thecal cells mapped in IPA (A) and GO terms (B) classified under biological process. In (A) on the left, the percentage of genes from the data set that map to each canonical pathway which are up regulated (in red) and down regulated (in blue) in vitro with respect to in vivo are shown. On the right, these pathways are then ranked from the highest to lowest degree of association based on multiple correction testing for the Benjamini-Hochberg False Discovery Rate (A). In (B) on the left, the percentage of genes from the data set that map to each GO term which are differentially regulated (in blue) in vitro with respect to in vivo are shown. These pathways are ranked from the highest to lowest degree of association using the Benjamini-Yuketeli test for multiple corrections (bottom to top in graphs on right).
Fig 6. Regulator effect network analysis in IPA identifying altered regulators and networks in thecal cells cultured in vitro.

Orange and blue are predicted up and down regulated regulators on top, and the pink and green represent up and down regulated genes from the array data with the intensity of colour reflecting the degree of change. The pathways altered are listed at the bottom and orange indicates they are up regulated in vitro.
Similar to the comparison described for granulosa cells above, thecal transcriptional profiles associated with cultured cells and atretic follicles revealed that there was only 3.6% overlap in the genes differentially expressed between these conditions and healthy follicles in vivo. Furthermore, only one gene showed altered expression during antral follicle development, out of the total number of 604 due to the effect of in vitro culture.
Discussion
In vitro culture models utilising isolated granulosa cells and cells from the thecal layers of ovarian follicles have been used extensively to produce data to create and test hypotheses about ovarian follicular development [21–25]. However, it is generally well recognised that in vitro conditions will likely alter the behaviour of the cells once removed from their normal follicular environment where they are exposed to a myriad of factors derived from neighbouring somatic cells, the oocyte and from the circulation. To identify which behaviours and genes are altered upon culture we therefore conducted analyses of microarrays of RNA expression profiles of both bovine granulosa cells and cells from theca interna layer to compare the transcriptomes of cultured cells with those of their in vivo counterparts.
Both granulosa cell and cells from the thecal layers behaved similarly in some respects. Both hierarchical clustering and PCA indicated that the in vitro transcriptomes were different to the in vivo transcriptomes, and that culture produced a more uniform expression pattern across the whole mRNA pool in both cell types. This might have been expected as the cultures were conducted for a 6-day period and the composition of the media with respect to added hormones and factors were constant. However, it could reflect that the in vivo data were derived from individual follicles whereas the in vitro data were from pools of approximately 50 follicles.
Ingenuity Pathway Analyses and GOEAST analyses of granulosa cells identified that cell adhesion and extracellular matrix related pathways were up regulated in vitro. Regulator effect network analysis identified many effects on cell adhesion, movement, binding, migration and chemo-attraction. Clearly the culture conditions in two dimensions on plastic, activated granulosa cell movement and cell matrix interactions. Indeed this would be consistent with the observation that the mono-dispersed cells granulosa cells plated out in the culture wells re-aggregate into 3-dimensional clumps over the course of the culture period [5,10]. Regulator effect network analysis of thecal cell cultures also identified many effects on cell movement, adhesion, migration, homing, filopodia, extension of cell processes and chemo-attraction, probably indicating interactions associated with attachment and interaction with the culture dish and cellular clumping [7,9]. The upstream regulators in granulosa cells that were predicted to be switched on included SREB1, HMGB1, FOS, IRF3, IRF5, JUNB, ID3 and IFI16 and those predicted to be switched off included NR1H2, SIRT1, NR3C1 and MYC. Notably, only one of these, IRF3, is specifically common with the upstream regulators in thecal cell cultures that were predicted to be switched on and that also included STAT4, HIF1A, SNAI1, IRF3, SMAD3, EGR1, CREBP, NFKB2, FOXO4 and MYB. Members of the interferon regulatory family (IRF) that regulate transcription of interferon genes were identified in the upstream regulator pathways (IRF3 and IRF5) and also identified in the list of differentially up regulated genes in both cell types (IRF1).
This type of effect has been reported for other cell types where adherence to plastic initiates cytoskeletal changes in epithelial cells and immune signalling in macrophages [26–28]. Granulosa cells, like macrophages, express Toll-Like Receptors (TLRs) and are capable of responding to bacterial endotoxins and inflammatory cytokines [29]. It is possible that the culture system used in our experiments produced these kinds of phenotypic changes in the granulosa cells. This has possible implications for the design of experiments which investigate granulosa cell behaviour, whereby the surface substrate composition and rigidity become important due to the effect on specific signalling pathways as shown here.
Our transcriptome analysis also indicated that phagocytosis was activated in cultures of granulosa cells. Macrophages can undertake phagocytosis for immunity (M1) or tissue repair (M2). However, the bovine membrana granulosa is free of immune cells until follicle atresia or ovulation commences [17,30] and granulosa cells have the ability to carry out phagocytosis [31], as can other epithelial cells [32]. Additionally there did not appear to be an activation of an innate immune response in the cultured granulosa cells despite these cells having this capability [29,33] as there was no increase in abundance of IL-1β or IL-6. Hence it is likely that phagocytosis is carried out by the granulosa cells in vitro as they can do in vivo to remove dead and dying neighbouring granulosa cells [31].
Ingenuity Pathway Analyses and GOEAST analyses of thecal cells identified substantial up regulation of immune responses and the major histocompatibility complex molecules (MHC Class 1: BOLA-N JSP.1, BOLA-A; MHC class 2: BOLA-DQA2, BOLA-DQA5, CD74); interleukins IL18, IL6 and IL8. The inflammatory chemokines CXCL6, CXCL2, CCL5, CCL3 and interferon gamma were also up regulated upon culture. These responses appear to be driving an innate immune response but possibly also initiating an adaptive immune response. MHC class II antigens are found in antigen presenting cells such as dendritic cells and macrophages. The vast majority of the immune cells in the theca interna layer are macrophages and neutrophils with relatively few T lymphocytes present [34–36] and these would have undoubtedly been present in the thecal interna cell cultures. The question arises: why would an immune response be triggered at all? The culture medium was serum free, but with additives of insulin or serum albumin which were bovine in origin and highly unlikely to induce expression of these genes involved in an immune response. The media also contained penicillin, streptomycin and amphotericin B but these agents would not be predicted to initiate an immune response. Another possibility is that microbial contamination during the culture period up regulated immune response genes but no overt signs of microbial contamination were ever observed. There is the possibility that the response is to dead cells or to non-self antigens, as the theca preparations were from pools of follicles from different animals. Thus, whilst we do not know the type of immune response or its cause it is clear that, in vitro, increased expression of inflammatory cytokines occurs and this could possibly affect the outcome and/or interpretation of any in vitro experiment using this culture system.
In conclusion, in vitro culture of both granulosa cells and cells from the theca interna layer alters the behaviour and transcriptomes of these cells, making the transcriptomes more uniform but with up regulation of cell behaviour associated with cell matrix interactions and with adhesion. Additionally, with cultured thecal interna cells there was clear evidence of an immune response likely mediated by leukocytes present within those cultures. These changes are important to be aware of when interpreting data derived from such cell culture systems. The approach we have taken could also be applied more widely to design and validate cell culture systems that more faithfully mimic in vivo situations.
Supporting information
The unsupervised hierarchical clustering across all probe sets (n = 24,182) for 17 arrays from granulosa cells was performed using the Euclidian dissimilarity algorithm with the average linkage method in Partek Genomics Suite. The heatmap represents the distribution of normalised signal intensity, grouping by pattern similarity for both probe set and array. Abbreviations are as for Fig 2A.
(JPG)
{kind=link}
All statistical cut-offs for differential expression were > 4-fold change with FDR P < 0.05. In A, the upper diagram shows numbers of genes with altered expression due to culture or atresia, the lower diagram shows numbers of genes with differences in expression due to culture or maturation. Intersected regions indicate those genes which are shared between different conditions and the proportion of the total numbers with changed expression resulting from culture alone. B shows those genes which are differentially regulated during atresia and culture and also those genes up regulated in the theca interna in vivo, indicating the proportion of genes with altered expression common to the different cell types of the follicle.
(JPG)
{kind=link}
The unsupervised hierarchical clustering occurred across all probe sets (n = 24,182) for 18 arrays from thecal cells was performed using the Euclidian dissimilarity algorithm with the average linkage method in Partek Genomics Suite. The heatmap represents the distribution of normalised signal intensity, grouping by pattern similarity for both probe set and array. Abbreviations are as for Fig 2B.
(JPG)
{kind=link}
The complete gene list is showing full names of the genes, fold changes, signal intensities and FDR values (> 4-fold change, FDR P < 0.05).
(PDF)
The complete gene list showing full names of the genes, fold changes, signal intensities and FDR values (> 4-fold change, FDR P < 0.05).
(PDF)
Acknowledgments
This work was supported by the United Kingdom Biotechnology and Biological Sciences Research Council (grant no. BB/G017174/1 to P.G.K. and C.G. and Underwood Fellowship award to R.J.R.) and by the National Health and Medical Research Council of Australia and the Australian Research Council and the University of Adelaide.
Data Availability
The CEL files can be obtained at NCBI with accession numbers GSE39589 and GSE49505 for granulosa and theca data, respectively.
Funding Statement
This work was supported by the United Kingdom Biotechnology and Biological Sciences Research Council (grant no. BB/G017174/1 to PGK and CG and Underwood Fellowship award to RJR) and by the National Health and Medical Research Council of Australia and the Australian Research Council and the University of Adelaide.
References
- 1.van Wezel IL, Rodgers RJ Morphological characterization of bovine primordial follicles and their environment in vivo. Biol Reprod 1996; 55: 1003–1011. [DOI] [PubMed] [Google Scholar]
- 2.Channing CP, Bae IH, Stone SL, Anderson LD, Edelson S, Fowler SC Porcine granulosa and cumulus cell properties. LH/hCG receptors, ability to secrete progesterone and ability to respond to LH. Mol Cell Endocrinol 1981; 22: 359–370. [DOI] [PubMed] [Google Scholar]
- 3.Lavranos TC, Rodgers HF, Bertoncello I, Rodgers RJ Anchorage-independent culture of bovine granulosa cells: the effects of basic fibroblast growth factor and dibutyryl cAMP on cell division and differentiation. Exp Cell Res 1994; 211: 245–251. 10.1006/excr.1994.1084 [DOI] [PubMed] [Google Scholar]
- 4.Rodgers HF, Lavranos TC, Vella CA, Rodgers RJ Basal lamina and other extracellular matrix produced by bovine granulosa cells in anchorage-independent culture. Cell Tissue Res 1995; 282: 463–471. [DOI] [PubMed] [Google Scholar]
- 5.Gutierrez CG, Campbell BK, Webb R Development of a long-term bovine granulosa cell culture system: induction and maintenance of estradiol production, response to follicle-stimulating hormone, and morphological characteristics. Biol Reprod 1997; 56: 608–616. [DOI] [PubMed] [Google Scholar]
- 6.Alila HW, Hansel W Origin of different cell types in the bovine corpus luteum as characterized by specific monoclonal antibodies. Biol Reprod 1984; 31: 1015–1025. [DOI] [PubMed] [Google Scholar]
- 7.Campbell BK, Baird DT, Webb R Effects of dose of LH on androgen production and luteinization of ovine theca cells cultured in a serum-free system. J Reprod Fertil 1998; 112: 69–77. [DOI] [PubMed] [Google Scholar]
- 8.Glister C, Kemp CF, Knight PG Bone morphogenetic protein (BMP) ligands and receptors in bovine ovarian follicle cells: actions of BMP-4, -6 and -7 on granulosa cells and differential modulation of Smad-1 phosphorylation by follistatin. Reproduction 2004; 127: 239–254. 10.1530/rep.1.00090 [DOI] [PubMed] [Google Scholar]
- 9.Glister C, Richards SL, Knight PG Bone morphogenetic proteins (BMP) -4, -6, and -7 potently suppress basal and luteinizing hormone-induced androgen production by bovine theca interna cells in primary culture: could ovarian hyperandrogenic dysfunction be caused by a defect in thecal BMP signaling? Endocrinology 2005; 146: 1883–1892. 10.1210/en.2004-1303 [DOI] [PubMed] [Google Scholar]
- 10.Glister C, Tannetta DS, Groome NP, Knight PG Interactions between follicle-stimulating hormone and growth factors in modulating secretion of steroids and inhibin-related peptides by nonluteinized bovine granulosa cells. Biol Reprod 2001; 65: 1020–1028. [DOI] [PubMed] [Google Scholar]
- 11.Hatzirodos N, Hummitzsch K, Irving-Rodgers HF, Harland ML, Morris SE, Rodgers RJ Transcriptome profiling of granulosa cells from bovine ovarian follicles during atresia. BMC Genomics 2014; 15: 40 10.1186/1471-2164-15-40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hatzirodos N, Irving-Rodgers HF, Hummitzsch K, Rodgers RJ Transcriptome profiling of the theca interna from bovine ovarian follicles during atresia. PLoS One 2014; 9: e99706 10.1371/journal.pone.0099706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Glister C, Hatzirodos N, Hummitzsch K, Knight PG, Rodgers RJ The global effect of follicle-stimulating hormone and tumour necrosis factor α on gene expression in cultured bovine ovarian granulosa cells. BMC Genomics 2014; 15: 72 10.1186/1471-2164-15-72 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Glister C, Satchell L, Bathgate RA, Wade JD, Dai Y, Ivell R, et al. Functional link between bone morphogenetic proteins and insulin-like peptide 3 signaling in modulating ovarian androgen production. Proc Natl Acad Sci U S A 2013; 110: E1426–1435. 10.1073/pnas.1222216110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hatzirodos N, Irving-Rodgers HF, Hummitzsch K, Harland ML, Morris SE, Rodgers RJ Transcriptome profiling of granulosa cells of bovine ovarian follicles during growth from small to large antral sizes. BMC Genomics 2014; 15: 24 10.1186/1471-2164-15-24 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Hatzirodos N, Hummitzsch K, Irving-Rodgers HF, Rodgers RJ Transcriptome profiling of the theca interna in transition from small to large antral ovarian follicles. PLoS One 2014; 9: e97489 10.1371/journal.pone.0097489 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Irving-Rodgers HF, van Wezel IL, Mussard ML, Kinder JE, Rodgers RJ Atresia revisited: two basic patterns of atresia of bovine antral follicles. Reproduction 2001; 122: 761–775. [PubMed] [Google Scholar]
- 18.Rodgers RJ, Irving-Rodgers HF Morphological classification of bovine ovarian follicles. Reproduction 2010; 139: 309–318. 10.1530/REP-09-0177 [DOI] [PubMed] [Google Scholar]
- 19.Kramer A, Green J, Pollard J Jr., Tugendreich S Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014; 30: 523–530. 10.1093/bioinformatics/btt703 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Yamada S, Fujiwara H, Honda T, Higuchi T, Nakayama T, Inoue T, et al. Human granulosa cells express integrin alpha2 and collagen type IV: possible involvement of collagen type IV in granulosa cell luteinization. Mol Hum Reprod 1999; 5: 607–617. [DOI] [PubMed] [Google Scholar]
- 21.Woodruff TK, Shea LD The role of the extracellular matrix in ovarian follicle development. Reprod Sci 2007; 14: 6–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Desai N, Alex A, AbdelHafez F, Calabro A, Goldfarb J, Fleischman A, et al. Three-dimensional in vitro follicle growth: overview of culture models, biomaterials, design parameters and future directions. Reprod Biol Endocrinol 2010; 8: 119 10.1186/1477-7827-8-119 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Havelock JC, Rainey WE, Carr BR Ovarian granulosa cell lines. Mol Cell Endocrinol 2004; 228: 67–78. 10.1016/j.mce.2004.04.018 [DOI] [PubMed] [Google Scholar]
- 24.Erickson GF Primary cultures of ovarian cells in serum-free medium as models of hormone-dependent differentiation. Mol Cell Endocrinol 1983; 29: 21–49. [DOI] [PubMed] [Google Scholar]
- 25.Wickenheisser JK, Nelson-DeGrave VL, McAllister JM Human ovarian theca cells in culture. Trends Endocrinol Metab 2006; 17: 65–71. 10.1016/j.tem.2006.01.003 [DOI] [PubMed] [Google Scholar]
- 26.Eierman DF, Johnson CE, Haskill JS Human monocyte inflammatory mediator gene expression is selectively regulated by adherence substrates. J Immunol 1989; 142: 1970–1976. [PubMed] [Google Scholar]
- 27.Liscia DS, Bernardi A, Griselli B, Cappa AP Different substrates influence the expression of intermediate filaments and the deposition of basement membrane proteins. In Vitro Cell Dev Biol 1988; 24: 183–187. [DOI] [PubMed] [Google Scholar]
- 28.Xing Z, Jordana M, Gauldie J IL-1 beta and IL-6 gene expression in alveolar macrophages: modulation by extracellular matrices. Am J Physiol 1992; 262: L600–605. [DOI] [PubMed] [Google Scholar]
- 29.Price JC, Sheldon IM Granulosa cells from emerged antral follicles of the bovine ovary initiate inflammation in response to bacterial pathogen-associated molecular patterns via Toll-like receptor pathways. Biol Reprod 2013; 89: 119 10.1095/biolreprod.113.110965 [DOI] [PubMed] [Google Scholar]
- 30.Spanel-Borowski K, Rahner P, Ricken AM Immunolocalization of CD18-positive cells in the bovine ovary. J Reprod Fertil 1997; 111: 197–205. [DOI] [PubMed] [Google Scholar]
- 31.van Wezel IL, Dharmarajan AM, Lavranos TC, Rodgers RJ Evidence for alternative pathways of granulosa cell death in healthy and slightly atretic bovine antral follicles. Endocrinology 1999; 140: 2602–2612. 10.1210/endo.140.6.6758 [DOI] [PubMed] [Google Scholar]
- 32.Kerr JF, Gobe GC, Winterford CM, Harmon BV Anatomical methods in cell death. Methods Cell Biol 1995; 46: 1–27. [DOI] [PubMed] [Google Scholar]
- 33.Price JC, Bromfield JJ, Sheldon IM Pathogen-associated molecular patterns initiate inflammation and perturb the endocrine function of bovine granulosa cells from ovarian dominant follicles via TLR2 and TLR4 pathways. Endocrinology 2013; 154: 3377–3386. 10.1210/en.2013-1102 [DOI] [PubMed] [Google Scholar]
- 34.Brannstrom M, Mayrhofer G, Robertson SA Distribution of leukocyte subsets in the rat ovary during ovulation. Biology of Reproduction 1992; suppl 1. [DOI] [PubMed] [Google Scholar]
- 35.Brannstrom M, Pascoe V, Norman RJ, McClure N Localization of leukocyte subsets in the follicle wall and in the corpus luteum throughout the human menstrual cycle. Fertil Steril 1994; 61: 488–495. [PubMed] [Google Scholar]
- 36.Best CL, Pudney J, Welch WR, Burger N, Hill JA Localization and characterization of white blood cell populations within the human ovary throughout the menstrual cycle and menopause. Hum Reprod 1996; 11: 790–797. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The unsupervised hierarchical clustering across all probe sets (n = 24,182) for 17 arrays from granulosa cells was performed using the Euclidian dissimilarity algorithm with the average linkage method in Partek Genomics Suite. The heatmap represents the distribution of normalised signal intensity, grouping by pattern similarity for both probe set and array. Abbreviations are as for Fig 2A.
(JPG)
All statistical cut-offs for differential expression were > 4-fold change with FDR P < 0.05. In A, the upper diagram shows numbers of genes with altered expression due to culture or atresia, the lower diagram shows numbers of genes with differences in expression due to culture or maturation. Intersected regions indicate those genes which are shared between different conditions and the proportion of the total numbers with changed expression resulting from culture alone. B shows those genes which are differentially regulated during atresia and culture and also those genes up regulated in the theca interna in vivo, indicating the proportion of genes with altered expression common to the different cell types of the follicle.
(JPG)
The unsupervised hierarchical clustering occurred across all probe sets (n = 24,182) for 18 arrays from thecal cells was performed using the Euclidian dissimilarity algorithm with the average linkage method in Partek Genomics Suite. The heatmap represents the distribution of normalised signal intensity, grouping by pattern similarity for both probe set and array. Abbreviations are as for Fig 2B.
(JPG)
The complete gene list is showing full names of the genes, fold changes, signal intensities and FDR values (> 4-fold change, FDR P < 0.05).
(PDF)
The complete gene list showing full names of the genes, fold changes, signal intensities and FDR values (> 4-fold change, FDR P < 0.05).
(PDF)
Data Availability Statement
The CEL files can be obtained at NCBI with accession numbers GSE39589 and GSE49505 for granulosa and theca data, respectively.