

Abstract
Background
The composition and structure of microbial communities that inhabit the mosquito midguts are poorly understood despite their well-documented potential to impede pathogen transmission.
Methodology/Principal findings
We used MiSeq sequencing of the 16S rRNA gene to characterize the bacterial communities of field-collected populations of 12 mosquito species. After quality filtering and rarefaction, the remaining sequences were assigned to 181 operational taxonomic units (OTUs). Approximately 58% of these OTUs occurred in at least two mosquito species but only three OTUs: Gluconobacter (OTU 1), Propionibacterium (OTU 9), and Staphylococcus (OTU 31) occurred in all 12 mosquito species. Individuals of different mosquito species shared similar gut microbiota and it was common for individuals of the same species from the same study site and collection date to harbor different gut microbiota. On average, the microbiota of Aedes albopictus was the least diverse and significantly less even compared to Anopheles crucians, An. quadrimaculatus, Ae. triseriatus, Ae. vexans, Ae. japonicus, Culex restuans, and Culiseta inornata. The microbial community of Cx. pipiens and Ae. albopictus differed significantly from all other mosquitoes species and was primarily driven by the dominance of Wolbachia.
Conclusion and significance
These findings expand the range of mosquito species whose gut microbiota has been characterized and sets the foundation for further studies to determine the influence of these microbiota on vector susceptibility to pathogens.
Author summary
The microbial communities that reside in mosquito midguts can impact transmission of mosquito-borne pathogens. We used high throughput next generation sequencing to characterize the midgut microbial communities of 12 mosquito species collected in urban residential areas in Champaign County, Illinois. A total of 181 OTUs from 11 phyla and 66 families were identified. Although several bacterial taxa were shared between two or more mosquito species, there was remarkable individual differences in gut microbiota and it was common for individuals of different mosquito species to harbor similar gut microbiota. The microbiota of Ae. albopictus was the least diverse and significantly less evenly distributed compared to 7 of 11 mosquito species. The microbial community of Cx. pipiens and Ae. albopictus differed significantly from other mosquito species and was primarily dominated by Wolbachia. These findings improve current knowledge on the composition and structure of mosquito gut microbiota and provide the framework for understanding their contribution to individual variation in vector competence and potential application in disease control.
Introduction
Mosquitoes transmit a wide range of pathogens that cause diseases in humans and other animals. The majority of mosquito-borne pathogens were previously confined to small geographic regions in the tropics but have recently emerged as a worldwide threat to human and animal health. Recent examples of mosquito-borne diseases that have caused major epidemics outside their native geographic range include West Nile virus [1], dengue virus [2], Chikungunya virus [3] and Zika virus [4, 5].
The transmission cycle of mosquito-borne pathogens involve interactions between at least three species: the pathogen, the vector, and the vertebrate host. When the mosquito takes a blood meal from an infected vertebrate host, the pathogen invades the midgut tissue where it undergoes further development and/or replication and then disseminates to secondary tissues such as nerve tissue, fat body, and finally the salivary glands [6]. At this point, the mosquito is considered infectious and is capable of transmitting the pathogen during a subsequent blood meal. However, the mosquito midgut is known to possess factors that may impede successful transmission of the pathogen [7–10]. These factors include the mosquito innate immune system and the digestive enzymes [6, 8, 11].
It is also well established that the mosquito midgut is colonized by a community of bacteria that can affect vector susceptibility to pathogens e.g. [12, 13]. For example, certain bacterial isolates from natural mosquito populations have been shown to reduce mosquito susceptibility to Plasmodium and dengue infection [12, 14, 15]. These effects are exerted through activation of the mosquito immune system [16], generation of reactive oxygen species by certain microbes [15], and formation of a physical barrier to infection [17]. Likewise, modification of midgut microbiota of Anopheles gambiae and Aedes aegypti through antibiotic treatment has been shown to enhance susceptibility to Plasmodium [16] and dengue infection [18], respectively. Other studies have shown that some midgut bacterial isolates can be genetically modified to express molecules that impair pathogen development within the mosquito [19, 20]. Collectively, these findings suggest that the composition of mosquito midgut microbiota likely contributes to within- and between-species variation in vector competence that is typical of many (if not all) mosquito-borne disease systems. Moreover, these studies demonstrate the potential for exploiting microbial functions for symbiotic control of mosquito-borne diseases [21].
Over the last few decades numerous studies have used culture-dependent and culture-independent approaches to characterize the microbial communities in the midguts of mosquito populations. These studies have revealed that the composition and diversity of gut microbiota can vary dramatically within [22] and between mosquito species [23] and are influenced by host diet [24], developmental stage [24], larval environment [25], and pathogen infection [26, 27]. As such, additional studies comparing the microbial communities of different mosquito species can further improve our understanding of mosquito microbiota and propel identification of specific microbes that may be harnessed for disease control.
In this study, we characterized the microbiota of 12 mosquito species collected from Champaign County, Illinois. The aim of this study was to determine how gut microbial diversity, composition and structure differs between mosquito species. Overall, we observed some remarkable similarities in gut microbiota between individuals of different mosquito species that were dominated by one or two bacterial taxa. These bacterial communities tended to vary markedly between individuals. We also found significant differences in bacterial community structure between some mosquito species. These findings advance current knowledge on the microbial communities that reside in mosquito midguts and provide the foundation for investigating their role in mosquito biology and potential application in mosquito-borne disease control.
Materials and methods
Mosquito collection
Mosquito samples for this study were collected once per week (July 2, 2015 to October 15, 2015) outside 19 urban residential houses in Champaign County, Illinois with permission from property owners (Fig 1). The sites were located within a 10 km radius of each other. The collections were done using standard CDC miniature light traps that were baited with dry ice as an attractant. The traps were tied to a tree outside the respective houses and operated between 1800 hours and 0900 hours. Mosquitoes from each trap were transported live in cool boxes, identified morphologically to species [28], and stored at -80°C until further processing.
Fig 1. Map of the study area showing the location of the traps.
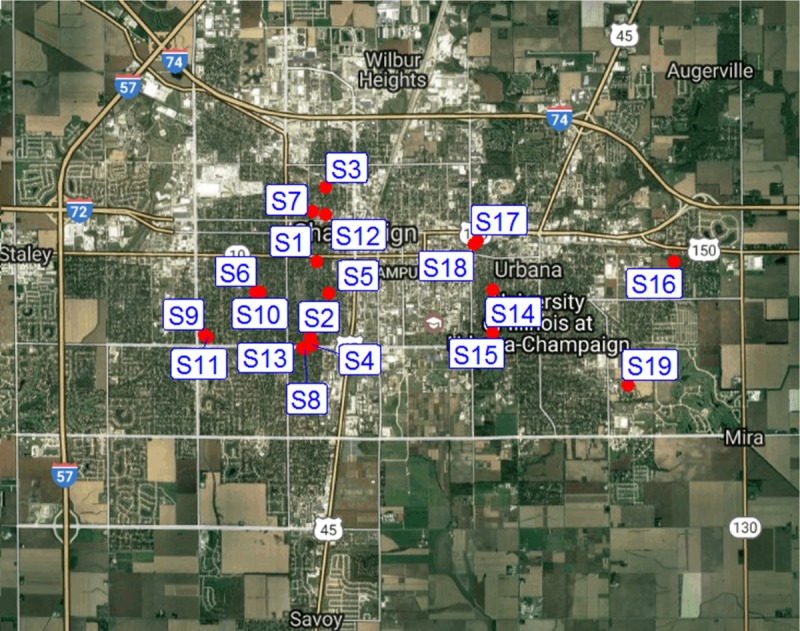
S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL.
Midgut dissection, DNA extraction and 16S rRNA gene library preparation
Individual female mosquitoes were surface sterilized as previously described [23] and dissected in 50 μl of Dulbecco’s phosphate buffered saline (DPBS) solution (Thermo Fisher Scientific, Waltham, MA). Total DNA was isolated from each midgut using QIAamp DNA mini kit (Qiagen, Valencia, CA). A portion of DNA from Culex mosquitoes was used for species identification using real-time polymerase chain reaction [29]. In total, 264 midguts from 12 mosquito species were processed (Table 1). The V3-V5 region of the 16S rRNA gene was amplified and sequenced using Illumina MiSeq Bulk v3 platform at the W. M. Keck Center for Comparative and Functional Genomics at the University of Illinois at Urbana-Champaign as previously described [23]. The following primer set was used: forward 5ʹ -CCTACGGGAGGCAGCAG-3`and reverse 5`-CCGTCAATTCMTTTRAGT-3ʹ.
Table 1. Number of midgdut samples that were processed for each of the 12 mosquito species collected in Urbana-Champaign, IL.
| Number of midgut samples | ||
|---|---|---|
| Mosquito species | Initial | Final |
| Aedes albopictus | 28 | 27 |
| Aedes japonicus | 28 | 21 |
| Aedes triseriatus | 28 | 23 |
| Aedes trivittatus | 28 | 18 |
| Aedes vexans | 27 | 27 |
| Anopheles crucians | 27 | 15 |
| Anopheles punctipennis | 27 | 14 |
| Anopheles quadrimaculatus | 27 | 15 |
| Culex pipiens | 16 | 15 |
| Culex restuans | 8 | 4 |
| Culiseta inornata | 11 | 10 |
| Psorophora ferox | 9 | 6 |
| 264 | 195 | |
In brief, all DNA samples were measured on a Qubit (Life Technologies) using High Sensitivity DNA Kit and diluted to 2 ng/μl. A master mix containing 0.5 μl -10X FastStart Reaction Buffer without MgCl2, 0.9 μl -25 mM MgCl2, 0.25 μl -DMSO, 0.1 μl -10 mM PCR grade Nucleotide Mix, 0.05 μl -5 U/μl FastStart High Fidelity Enzyme Blend, 0.25 μl -20X Access Array Loading Reagent, and 0.95 μl -water was prepared using the Roche High Fidelity Fast Start Kit and 20X Access Array loading reagent and aliquoted into 48 well PCR plates along with 1 μl DNA sample and 1 μl Fluidigm Illumina linkers (V3-V5-F357: ACACTGACGACATGGTTCTACA and V3-V5-R926:TACGGTAGCAGAGACTTGGTCT) with unique barcode. In a separate plate, primer pairs were prepared and aliquoted. 20X primer solutions were prepared by adding 2 μl of each forward and reverse primer, 5 μl of 20X Access Array Loading Reagent and water to a final volume of 100 μl.
Four μl of sample was loaded in the sample inlets and 4 μl of primer loaded in primer inlets of a previously primed Fluidigm 48.48 Access Array IFC. The IFC was placed in an AX controller (Fluidigm Corp.) for microfluidic loading of all primer/sample combinations. Following the loading stage, the IFC plate was loaded on the Fluidigm Biomark HD PCR machine and samples were amplified using the following Access Array cycling program without imaging: 50°C for 2 minutes (1 cycle), 70°C for 20 minutes (1 cycle), 95°C for 10 minutes (1 cycle), followed by 10 cycles at 95°C for 15 seconds, 60°C for 30 seconds, and 72°C for 1 minute, 2 cycles at 95°C for 15 seconds, 80°C for 30 seconds, 60°C for 30 seconds, and 72°C for 1 minute, 8 cycles at 95°C for 15 seconds, 60°C for 30 seconds, and 72° for 1 minute, 2 cycles at 95°C for 15 seconds, 80°C for 30 seconds, 60°C for 30 seconds, and 72°C for 1 minute, 8 cycles at 95°C for 15 seconds, 60°C for 30 seconds, and 72°C for 1 minute, and 5 cycles at 95°C for 15 seconds, 80°C for 30 seconds, 60°C for 30 seconds, and 72°C for 1 minute. The PCR product was transferred to a new 96 well plate, quantified on a Qubit fluorimeter (Thermo-Fisher) and stored at -20°C. All samples were run on a Fragment Analyzer (Advanced Analytics, Ames, IA) and amplicon regions and expected sizes confirmed. Samples were then pooled in equal amounts according to product concentration. The pooled products were size selected on a 2% agarose E-gel (Life Technologies) and extracted from the isolated gel slice with QIAquick gel extraction kit (QIAGEN). Cleaned size selected products were run on an Agilent Bioanalyzer to confirm appropriate profile and determination of average size. The final library pool was spiked with 10% non-indexed PhiX control library (Illumina) and sequenced using Illumina MiSeq V3 Bulk system. The libraries were sequenced from both ends of the molecules to a total read length of 300nt from each end. Cluster density was 964k/mm2 with 85.9% of clusters passing filter.
OTU picking and taxonomy assignment
IM-TORNADO 2.0.3.2 platform was used to process the de-multiplexed fasq-formatted files obtained from the sequencing facility. This platform is designed to process non-overlapping reads for analysis as a whole unit without sacrificing one of the reads in the pair and improves accuracy in read analysis compared to single-end read analysis [30]. The 5ʹ PCR primer for forward (R1) and reverse (R2) reads were trimmed using Trimmomatic program [31] with the parameter HEADCROP:17 for R1 read and HEADCROP: 18 for R2 read. The quality filtering process was performed using Trimmomatic program following previously described procedures with slight modifications [30]. Briefly, the sequences were subjected to a hard cutoff of PHRED score Q3 for 5 ʹ and 3ʹ ends of the reads (parameters LEADING: 3 and TRAILING: 3), trimming of the 3’ end with a moving average score of Q15, with a window size of four bases (parameter SLIDINGWINDOW: 4:15), and any reads with less than 150 base pairs removed with parameter R1_TRIM = 150 and R2_TRIM = 150. Reads with ambiguous base calls were discarded. To retain both reads while avoiding misinterpretation of the data, matching R1 and R2 reads were joined using an ambiguous nucleotide character “N” between R1 and R2 [30]. In a single run, IM-TORNADO generates outputs for R1 data only, R2 data only, and paired end data. Only output files related to paired end data were used for taxonomic assignment and downstream analysis. Reads were de-replicated building clusters of reads with 100% similarity and annotated with cluster size. Singletons and reads shorter than the cutoff length were discarded to ensure the use of high quality reads when assigning OTU representation. Reads were sorted by cluster size and processed in USEARCH using the UPARSE algorithm to find the OTU representatives using de novo OTU picking strategy. Chimeric reads are also removed during this step resulting in a set of OTU representatives of very high sequence quality [32]. Operational taxonomic units (OTUs) were assigned at 97% sequence similarity using the Ribosomal Database Project (RDP) version 10 as the reference set with a threshold of 80% bootstrap confidence [33].
PCR validation for Wolbachia surface antigen, wsp
Quantitative TaqMan real-time PCR (qPCR) was used to confirm the wsp gene of Wolbachia in mosquito midgut samples using the following primer set: forward: 5’-GSTTTTGCTKRTCAAGYAARAG-3’ and reverse: 5’-GYGCTGTAAAGAACKTTGWDY-3' respectively. Taqman probe sequence was 5’ FAM-TGTTAGTTATGATGTAACTCCRGAA-IABFQ 3’. The primers and probe were synthesized by Integrated DNA Technology, Inc. (IDT, Coralville, IA). Twenty microliter qPCR contained 1× SensiFAST Probe Hi Rox mastermix (BioLine, Taunton, MA), 0.5 μM of each primer, 0.25 μM Taqman probe and 2 μL of the mosquito midgut DNA isolate. The qPCR was run with 1 cycle of heat activation at 95°C for 15 minutes followed by 45 cycles of denaturation at 94°C for 1 minute, annealing at 50°C for 1 minute and elongation at 72°C for 1 minute.
Minigene was constructed using wsp gene segment flanked by the PCR primers and was synthesized by IDT (Coralville, IA). The gene sequences utilized for the minigene were downloaded from GenBank and the accession number was CP001391 for Wolbachia spp wRi. The minigene was used as a positive control for qPCR of Wolbachia wsp gene and as templates for building a standard curve to estimate the quantity of wsp gene in mosquito midgut samples. The copy number of minigene (2063 bp) containing wsp gene segment was calculated based on the DNA concentration determined by NanoDrop 1000 spectrophotometer (Thermo Scientific) and on the assumption that the average weight of a DNA base pair (bp) is 650 Daltons. The formula for copy number calculation is: copy numbers = ((minigene amounts in ng) × (6.022 × 1023)) / (2063 × 650 × 109). The concentration of minigene solution was adjusted to be 5 × 109 copies/μl and 10-fold serially diluted in nuclease free water (BioLine, Taunton, MA). Two microliter of the serially diluted minigene solution was utilized for qPCR. A standard curve was generated using the relationship between the cycle numbers at threshold (Ct values) and the minigene copy numbers in serially diluted minigene solution.
Statistical analysis
Unless otherwise stated statistical analysis were conducted using R 3.2.3 statistical software (https://cran.r-project.org/bin/windows/base/old/3.2.3/). OTUs accounting for < 0.005% of the total number of sequences were discarded before downstream analysis to reduce the problem of spurious OTUs [34]. The number of sequences varied markedly among individual mosquito midguts (mean ± SE = 6834.72 ± 460.75 per mosquito midgut; minimum = 0, maximum = 39,268). We rarefied the read depth to 1,036 reads per sample to standardize the sampling effort. Sixty nine samples that did not meet this criterion (i.e. had < 1,036 sequences) were excluded from subsequent analysis (Table 1). Alpha diversity metrics including Shannon diversity index, observed species, chao1, and evenness were generated in QIIME [35] and analysis of variance with Tukey adjustments was used to test whether there were any significant differences in these indices among mosquito species. Analysis of similarities (ANOSIM) using the “vegan” package in R was used to test whether microbial communities from samples of each mosquito species were more similar than those of different mosquito species [36]. The computed Bray-Curtis similarity matrix values were used for principal coordinate analysis (PCoA) to determine microbial community differences across mosquito species (“vegan” package in R). Hierarchical clusters based on Bray-Curtis dissimilarity measure were performed in PAST software to highlight the differences in mosquito samples based on the composition and abundance of their gut microbiota [37]. Similarity percentage (SIMPER) analysis was used to identify OTUs that were primarily responsible for observed differences between mosquito species (PAST version 3.14 software [37]).
Results
Bacterial species composition across mosquito species
MiSeq sequencing of the V3-V5 region of 16S rRNA gene amplicons from 264 mosquito samples generated a total of 1,804,366 sequences (Mean ± SE = 6834.72 ± 460.75 per mosquito midgut sample). After quality filtering and rarefying the reads to an even sampling depth of 1,036 sequences, a total of 202,020 sequences from 195 mosquito samples were retained. These sequences were clustered into 181 bacterial OTUs belonging to 11 phyla, 66 families and 111 genera. Only 16 of the 181 OTUs had an overall abundance equal to or greater than 1%. The majority of sequences were from Proteobacteria (81.1%) comprising of Alphaproteobacteria (47.4%), Gammaproteobacteria (29.2%), Betaproteobacteria (3.2%), Epsilonproteobacteria (1.1%), and Deltaproteobacteria (0.3%). Other observed phyla included, Actinobacteria (8.8%), Firmicutes (5.7%), Bacteroidetes (1.8%), Acidobacteria (0.8%), Cyanobacteria (0.6%), Tenericutes (0.5%), Spirochaetes (0.4%), Planctomycetes (0.3%), Parcubacteria (0.03%) and Fusobacteria (0.005%).
The most abundant OTUs were associated with the families Acetobacteraceae (25.7%), Enterobacteriaceae (20.6%), Rickettsiaceae (20.0%), Propionibacteriaceae (8.4%), and Orbaceae (4.2%) (S1 Fig). Acetobacteraceae occurred in high abundance among some individuals of all mosquito species except An. crucians. However, they were found in fewer individuals of Ae. albopictus, An. punctipennis, An. quadrimaculatus, Cx. pipiens and Cx. restuans compared to the remaining mosquito species. Enterobacteriaceae was more common among Ae. triseriatus, Ae. trivittatus, and Ae. vexans and also occurred in high abundance in the guts of a few individuals of the remaining mosquito species. Rickettsiaceae was more abundant and widespread in Ae. albopictus and Cx. pipiens and was also present in high abundance in a few samples of An. crucians, An. punctipennis, and An. quadrimaculatus. Propionibacteriaceae were mostly associated with An. crucians and An. punctipennis and occurred in high abundance in a few individuals of Ae. triseriatus, Ae. vexans, An. quadrimaculatus, Cx. restuans, and Cs. inornata. Orbaceae occurred in high abundance in a few individuals of An. crucians, An. punctipennis, An. quadrimaculatus, Cs. inornata, Ps. ferox, Ae. japonicus and Ae. triseriatus. Overall, only 1–3 major families of bacteria tended to dominate the guts of the 12 mosquito species (S1 Fig). It was also common for some individuals of a given mosquito species from the same study site and collection date to harbor different gut microbiota.
The top 9 OTUs accounted for 69.2% of the total sequences and their relative abundance varied markedly between mosquito species (Fig 2). OTU 1 (Gluconobacter) accounted for 23.1% of the total sequences and was more abundant in all Aedes mosquito species (except Ae. albopictus) as well as Cs. inornata and Ps. ferox. This OTU also occurred in high abundance in a few samples of Cx. pipiens, Cx. restuans, An. punctipennis, and An. quadrimaculatus. OTU 2 (Wolbachia) was more prevalent and abundant in the guts of Ae. albopictus and Cx. pipiens and also occurred in three Ae. japonicus samples and one sample each of An. crucians, An. punctipennis and An. quadrimaculatus. OTU 9 (Propionibacterium) was mostly associated with An. crucians and An. punctipennis but it also occurred in higher abundance in a few samples of other mosquito species. OTU 8 (Morganella) was mostly associated with Ae. triseriatus, Ae. trivittatus, and Ae. vexans and OTU 5 (Providencia) was mostly associated with Ae. vexans. OTU 182 (Gluconobacter) was mostly associated with Ae. japonicus but was also present in high abundance in the guts of some individuals of other mosquito species. OTU 6 (Orbus), OTU 86 (Pantoea), and OTU 12 (Tatumella) occurred in high abundance in one or a few individuals of different mosquitoes (Fig 2). Some individuals of a given mosquito species also tended to differ in their microbial composition despite being collected from the same study sites and collection dates. The majority of mosquito samples were dominated by 1–2 OTUs.
Fig 2. Mean relative abundances of bacterial OTUs associated with 12 species of mosquitoes at different sites and collection dates.

OTUs with abundance of less than 1.5% were pooled together as “Other”. S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL. T = date of collection; T1 = July, 2, 2015; T2 = July, 7, 2015; T3 = July, 21, 2015; T4 = July, 28, 2015; T5 = August, 3, 2015; T6 = August, 11, 2015; T7 = August, 19, 2015; T8 = August, 28, 2015; T9 = September, 4, 2015; and T10 = October, 15, 2015.
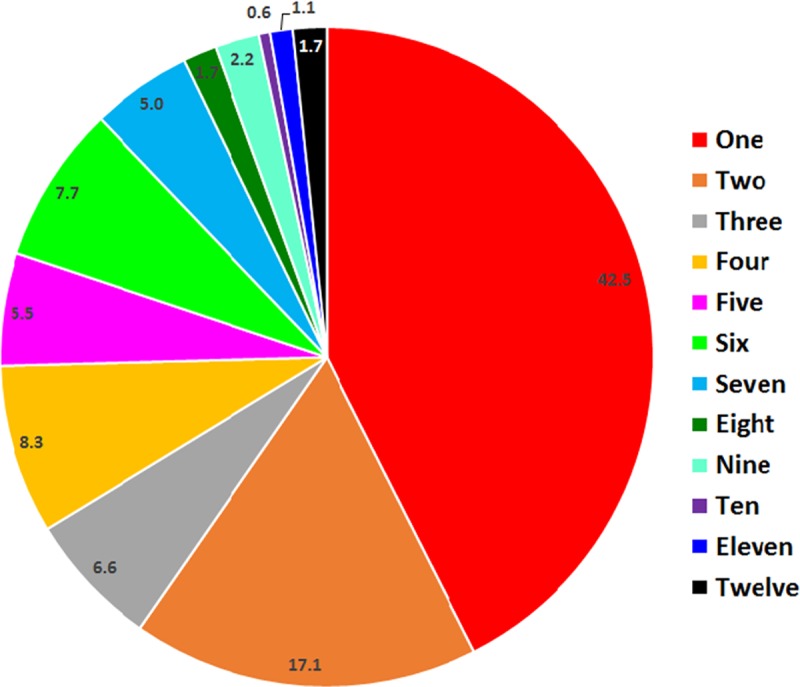
Overall, 57.5% of bacterial OTUs were shared between at least two mosquito species (Fig 3). However, only three bacterial OTUs occurred in all 12 mosquito species. These were OTU 1 (Gluconobacter), OTU 9 (Propionibacterium), and OTU 31 (Staphylococcus).
Fig 3. Number of OTUs that were unique to one mosquito species or shared between two or more mosquito species. n = 181.
Diversity of mosquito microbiota
Shannon diversity indices revealed that on average, the gut microbiota of Aedes albopictus was the least diverse and significantly less even compared to gut microbiota of An. crucians, An. quadrimaculatus, Ae. triseriatus, Ae. vexans, Ae. japonicus, Cx. restuans, and Cs. inornata (Shannon: F = 6.4, df = 11, 179, P < 0.001; Evenness: F = 6.4, df = 11, 179, P < 0.001; Table 2). The gut microbiota of An. crucians was also significantly more diverse and more evenly distributed compared to that of Ae. trivittatus, Cx. pipiens, and Ps. ferox (Table 2). We also calculated Chao1 estimator based on OTUs abundance to determine the expected richness in each sample (Table 2). We were able to detect more than 93% ± 1.3% (mean ± SE) of the expected number of OTUs suggesting that most OTUs were recovered. On average, our results revealed that a mosquito midgut contains 5–10 bacterial OTUs (Table 2). The observed and predicted (Chao1) number of OTUs were significantly lower in Ae. albopictus compared to Ae. vexans (Observed OTUs: F = 3.2, 11, 179, P = 0.0005; Chao 1: F = 2.6, df = 11, 179, P = 0.005; Table 2). Significantly more bacterial OTUs were also observed in An. crucians and Ae. triseriatus guts compared to Ae. albopictus guts.
Table 2. Bacterial diversity and richness (mean ± SE) in the guts of 12 mosquito species.
| Species | Shannon diversity | OTU evenness | Observed OTUS | Chao1 |
|---|---|---|---|---|
| Anopheles crucians | 1.87 ± 0.23 | 0.62 ± 0.06 | 8.00 ± 0.89 | 8.00 ± 0.89 |
| Anopheles punctipennis | 1.05 ± 0.15 | 0.39 ± 0.05 | 6.29 ± 0.57 | 7.04 ± 0.80 |
| Anopheles quadrimaculatus | 1.23 ± 0.20 | 0.45 ± 0.06 | 6.29 ± 0.53 | 6.82 ± 0.67 |
| Aedes albopictus | 0.36 ± 0.11 | 0.14 ± 0.03 | 4.50 ± 0.51 | 5.13 ± 0.67 |
| Aedes japonicus | 1.18 ± 0.17 | 0.43 ± 0.06 | 6.38 ± 0.47 | 7.14 ± 0.54 |
| Aedes triseriatus | 1.31 ± 0.15 | 0.45 ± 0.05 | 7.43 ± 0.68 | 8.41 ± 0.98 |
| Aedes vexans | 1.25 ± 0.11 | 0.41 ± 0.03 | 8.78 ± 0.88 | 9.82 ± 1.04 |
| Aedes trivittatus | 0.76 ± 0.12 | 0.31 ± 0.05 | 6.11 ± 0.50 | 6.58 ± 0.63 |
| Culex pipiens | 0.78 ± 0.15 | 0.29 ± 0.05 | 5.79 ± 0.59 | 6.18 ± 0.76 |
| Culex restuans | 1.61 ± 0.26 | 0.60 ± 0.09 | 6.50 ± 0.29 | 6.50 ± 0.29 |
| Culiseta inornata | 1.59 ± 0.33 | 0.54 ± 0.10 | 6.90 ± 0.99 | 7.15 ± 0.95 |
| Psorophora ferox | 0.65 ± 0.34 | 0.25 ± 0.10 | 5.83 ± 1.30 | 6.08 ± 1.37 |
Variation in midgut bacterial communities across mosquito species
The ANOSIM analysis based on Bray-Curtis distances revealed a significant difference in microbial communities among the 12 mosquito species (ANOSIM, R = 0.59, P = 0.001). To better visualize the results, a principal coordinates analysis (PCoA) plot was generated based on Bray-Curtis distances (Fig 4). Ordination based on this metric demonstrated a clear separation of Ae. albopictus and Cx. pipiens samples from the other mosquito species indicating that the microbial communities of the two mosquito species differed from those of the other mosquito species (Fig 4). Cluster analysis based on Bray-Curtis distances confirmed that the majority of Ae. albopictus and Cx. pipiens samples tended to cluster together and that it was common for individuals of different mosquito species from different sites and collection dates to harbor similar gut microbiota (S2 Fig).
Fig 4. Principal coordinates analysis (PCoA) comparing the bacterial communities across 12 mosquito species.

PCoA was based on Bray-Curtis distance values computed for mosquito communities at the 97% OTU level. CRU = An. crucians, PUN = An. punctipennis, QUA = An. quadrimaculatus, ALB = Ae. albopictus, JAP = Ae. japonicus, TRIS = Ae. triseriatus, VEX = Ae. vexans, TRIV = Ae. trivittatus, PIP = Cx. pipiens, RES = Cx. restuans, INO = Cs. inornata, FER = Ps. ferox.
The SIMPER analysis was used to identify the bacterial OTUs primarily responsible for the observed separation of gut communities between mosquito species, using the relative abundances of bacterial OTUs (S1 Table; S3 Fig). Twelve OTUs accounted for 69.8% of observed differences between mosquito species with OTU 1 (19%), OTU 2 (18%) and OTU 9 (8%) accounting for the largest variation (S1 Table). OTU 1 (Gluconobacter), was found in all mosquito species but was more abundant in Ae. japonicus, Ps. ferox, Ae. trivittatus, Ae. triseriatus, and Cs. inornata (S3 Fig). OTU 2 (Wolbachia) was mainly associated with Ae. albopictus and Cx. pipiens and OTU 9 (Propionibacterium) was mainly associated Cx. restuans, Ae. triseriatus and the three Anopheles species (An. crucians, An. quadrimaculatus, An. punctipennis, S3 Fig).
Wolbachia prevalence and wsp gene copy numbers
Real-time qPCR results confirmed the presence of Wolbachia in all three Ae. japonicus samples, 25 of 27 Ae. albopictus samples, 12 of 15 Cx. pipiens samples, and the 1 An. punctipennis sample (S4 Fig). None of the other mosquito species had Wolbachia. Wolbachia wsp gene copy numbers ranged from 0 to 10151 and were relatively higher in Ae. albopictus compared to the other mosquito species (S4 Fig).
Discussion
In this study we characterized and compared the midgut bacterial communities of 12 mosquito species encompassing four mosquito genera, many of them important vectors of medical, veterinary and wildlife significance. Overall, we found a low diversity of gut microbiota that was characterized by large individual variability and the dominance of one or two bacterial OTUs. Analysis of microbial composition revealed that the bacterial community in mosquito midguts was dominated by a few phyla with only three phyla (Proteobacteria (81.1%), Actinobacteria (8.8%) and Firmicutes (5.7%) accounting for 97% of the total sequences. These bacterial phyla are commonly reported in the guts of mosquitoes and other insects [22, 24, 25, 38, 39]. The Phylum Proteobacteria is highly diverse and contains a wide variety of species that are adapted to a wide range of environments; thus it is no surprise that its dominance in mosquito midguts is well established [22, 24, 25, 40, 41].
Individual variability in gut microbiota was not only restricted to mosquito samples collected from different sites and different dates but was also common among individual mosquitoes collected at the same sites and collection dates. Similar individual variability in gut microbiota and the dominance of a few bacterial taxa in mosquito guts has been reported before [22]. These variations may result from individual variations in external and internal factors such as the gut physiological conditions, larval and adult diet, infection with parasites and pathogens, host aging [24, 26, 27, 38, 42], and host genetic background [43]. Our experimental design cannot decipher the contribution of these factors to the observed pattern of gut microbiota since adult mosquito samples were collected using the CDC light traps and we had no prior knowledge of the factors these mosquitoes were exposed to before collection. Individual variation in gut microbiota may be epidemiologically relevant since some bacterial species are known to enhance [13, 44, 45] or reduce mosquito susceptibility to Plasmodium parasites and dengue viruses [14, 46, 47]. Thus it is possible that differences in gut microbiota observed in this study may be one of the primary factors contributing to individual variation in vector competence that is commonly observed in nature. Future studies targeting the role of specific members of this bacterial community on vector competence and other aspects of mosquito biology may provide important insights into their epidemiological significance.
Ae. albopictus and Cx. pipiens harbored distinct bacterial communities that was primarily dominated by OTU 2 (Wolbachia). We also found Wolbachia sequences in three samples of Ae. japonicus and one sample of An. crucians, An. quadrimaculatus, and An. punctipennis. Real-time qPCR results confirmed the widespread occurrence of Wolbachia in Ae. albopictus and Cx. pipiens samples as well as its presence in the 1 and 3 An. punctipennis and Ae. japonicus samples that had Wolbachia sequences, respectively. We processed only intact mosquitoes and sterilized their surfaces before dissecting their midguts to minimize the potential for contamination. This process is expected to remove bacteria from the body surface but it is still possible these mosquitoes were contaminated with Wolbachia from damaged Ae. albopictus and Cx. pipiens samples either in the traps or during sorting and sample identification. However, the dominance of Wolbachia sequences in one of An. punctipennis samples and three Ae. japonicus samples is unlikely due to cross contamination and may imply that a few individuals of Ae. japonicus and An. punctipennis may harbor Wolbachia endosymbionts. Wolbachia are a genus of maternally-inherited bacterial endosymbionts that are estimated to occur in approximately 65% of insect species [48]. This bacterium acts as a reproductive parasite in arthropods; it induces male killing, feminization, and cytoplasmic incompatibility which facilitate its spread throughout the arthropod population [49]. Both Ae. albopictus and Cx. pipiens are known to harbor Wolbachia endosymbionts [23, 38, 50–52] and our study suggest the need for detailed investigations of Wolbachia infection to ascertain that its absence in other mosquito species as reported in the past is not due to lack of adequate sampling effort. The mechanism underlying the high Wolbachia infection and low diversity of midgut bacteria in Ae. albopictus is unclear but could be due to methodological bias where the rarefaction depth of 1,036 employed in this study may not have been sufficient to detect low abundance OTUs or due to Wolbachia interacting negatively with other bacterial species. Additional studies are needed to develop a better understanding of how Wolbachia interacts with other microbiota. Wolbachia has been shown to inhibit transmission of mosquito-borne pathogens [53–55] and is currently under investigation for potential application in biological control of mosquitoes and associated pathogens [56–58]. Unfortunately, Wolbachia can also enhance transmission of other pathogens such as malaria and West Nile Virus [44, 45, 59]. These effects are dependent on Wolbachia strain and the mosquito-borne pathogen under investigation as it is possible for some Wolbachia strains to inhibit transmission of some pathogens while enhancing transmission of others [60, 61]. These findings reinforce the need to understand the potential impact of Wolbachia on different mosquito-borne pathogens before large scale application of Wolbachia-based disease control strategies.
SIMPER analyses indicated that OTU 1 (Gluconobacter), OTU 2 (Wolbachia), and OTU 9 (Propionibacterium) contributed most to the average dissimilarity between mosquito species. OTU 1 (Gluconobacter) was found in all mosquito species but was strongly associated with Ae. japonicus, Ae. triseriatus, Ae. vexans, Ae. trivittatus, Cs. inornata, and Ps. ferox. Gluconobacter are acetic acid bacteria that are adapted to various sugar- and ethanol-rich environments [62]. These bacteria have been found in association with insects that rely on sugar-based diets including mosquitoes [63, 64]. As an example, the genus Asaia (a member of Acetobacteraceae), are frequently found in the nectar of flowers e.g. [65–67] and have been shown to establish symbiotic associations with mosquitoes [63, 64, 68, 69]. Propionibacterium was mostly associated with Anopheles mosquitoes and Cx. restuans. Propionibacterium is a common bacteria of human skin and other animals [70–72] and has also been isolated in mosquitoes [73]. These bacteria may have been acquired from vertebrate hosts during a blood meal [73]. Another notable OTU accounting for observed differences was OTU 5 (Providencia) which was strongly associated with Ae. vexans. This bacterium is a common gastrointestinal pathogen of humans and animals and also occurs in human and animal wastes [74]. It may have been acquired through contact with blood meal hosts or during larval development. Further studies are needed to investigate the potential role of these bacteria on mosquito biology including susceptibility to pathogens.
In general, there were small differences in bacterial diversity and evenness between most species of mosquitoes. However, the bacterial communities of Ae. albopictus were significantly less diverse and less evenly distributed compared to those of An. crucians, An. quadrimaculatus, Ae. japonicus, Ae. triseriatus, Ae. vexans, Cx. restuans, or Cs. inornata. Similar bacterial diversity and evenness between mosquito species across the four mosquito genera suggest that the mosquito midgut likely plays an active role in regulating the colonization and assembly of bacterial communities. Lower microbial diversity in Ae. albopictus relative to the seven mosquito species may be due to inability of some bacterial taxa to proliferate in the guts of Ae. albopictus either due to species differences in gut physiological conditions [75] and/or modulation of microbial communities by the mosquito innate immune system [12]. The physical presence of some bacterial taxa or other microbes (e.g. fungi) also may render the mosquito midgut uninhabitable to other bacterial taxa due to interspecific competition for resources and/or production of toxins and inhibitory factors. Differences in food sources also may partly account for the observed differences because although all mosquito species tend to feed on microbes as larvae and blood and nectar as adults, different mosquito species portray marked variations in their preferred larval habitats and sugar and blood meal hosts which may pre-expose them to different microorganisms. In addition, sugar feeding and blood feeding can reduce the diversity of gut bacteria in mosquitoes [24]. Although we purposefully selected individuals that were not engorged with blood for microbiome analysis, we could not establish whether our mosquito samples had prior access to a blood meal or a sugar meal. It is possible that the majority of Ae. albopictus that were analyzed in this study had acquired a blood meal and/or a sugar meal leading to major reductions in bacterial diversity.
In summary, our study has characterized the midgut bacterial communities of 12 of the most common mosquito species in the United States, expanding current knowledge on mosquito species whose gut microbes have been studied. We found significant differences in gut microbial composition between some mosquito species and documented marked variation in gut microbiota between individuals of the same mosquito species. The 12 mosquito species included the known vectors of arboviruses of global public health significance such as dengue, chikungunya, Zika, West Nile virus, and La Crosse virus encephalitis. Given the well-documented ability of midgut microbiota to influence vector susceptibility to pathogens [12, 14–16, 25, 46], our results provide critical knowledge that can inspire further studies to determine which of the identified microbial communities could be exploited for disease control.
Supporting information
Families with abundance of less than 1.2% were pooled together as “Other”. S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL. T = date of collection; T1 = July, 2, 2015; T2 = July, 7, 2015; T3 = July, 21, 2015; T4 = July, 28, 2015; T5 = August, 3, 2015; T6 = August, 11, 2015; T7 = August, 19, 2015; T8 = August, 28, 2015; T9 = September, 4, 2015; and T10 = October, 15, 2015.
(TIFF)
CRU = An. crucians, PUN = An. punctipennis, QUA = An. quadrimaculatus, ALB = Ae. albopictus, JAP = Ae. japonicus, TRIS = Ae. triseriatus, VEX = Ae. vexans, TRIV = Ae. trivittatus, PIP = Cx. pipiens, RES = Cx. restuans, INO = Cs. inornata, FER = Ps. ferox. S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL. T = date of collection; T1 = July, 2, 2015; T2 = July, 7, 2015; T3 = July, 21, 2015; T4 = July, 28, 2015; T5 = August, 3, 2015; T6 = August, 11, 2015; T7 = August, 19, 2015; T8 = August, 28, 2015; T9 = September, 4, 2015; and T10 = October, 15, 2015.
(TIF)
CRU = An. crucians, PUN = An. punctipennis, QUA = An. quadrimaculatus, ALB = Ae. albopictus, JAP = Ae. japonicus, TRIS = Ae. triseriatus, VEX = Ae. vexans, TRIV = Ae. trivittatus, PIP = Cx. pipiens, RES = Cx. restuans, INO = Cs. inornata, FER = Ps. ferox. Number at the end of the genus name is the OTU number. Values in the heatmap cells represent the relative abundance of respective OTUs in different mosquito species.
(TIF)
S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL. T = date of collection; T1 = July, 2, 2015; T2 = July, 7, 2015; T3 = July, 21, 2015; T4 = July, 28, 2015; T5 = August, 3, 2015; T6 = August, 11, 2015; T7 = August, 19, 2015; T8 = August, 28, 2015; T9 = September, 4, 2015; and T10 = October, 15, 2015.
(TIF)
(DOCX)
Acknowledgments
We thank Andrew Donelson, Katherine Micek, Millon Blackshear, Therese Adgie, Therin Bradshaw, Dionysios Mikros, and Nathan Stables for their technical support. Any opinions, findings, conclusions, or recommendations expressed in this publication are those of the author(s) and do not necessarily reflect the view of the U.S. Department of Agriculture. The mention of firm names or trade products does not imply that they are endorsed or recommended by the U.S. Department of Agriculture over other firms or similar products not mentioned; the USDA is an equal opportunity employer.
Data Availability
All relevant data are within the paper and its Supporting Information files except for the 16S rRNA gene bacterial sequences which are available from Genbank under the accession number PRJNA374733.
Funding Statement
This study was partly supported by Waste Tire Fund from the State of Illinois. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Gubler DJ. The continuing spread of West Nile virus in the western hemisphere. Clin Infect Dis. 2007; 45(8):1039–46. 10.1086/521911 [DOI] [PubMed] [Google Scholar]
- 2.Gubler D, editor. Dengue and dengue hemorrhagic fever: its history and resurgence as a global public health problem. London (United Kingdom): CAB International; 1997. [Google Scholar]
- 3.Charrel RN, de Lamballerie X, Raoult D. Chikungunya outbreaks: the globalization of vectorborne diseases. N Engl J Med. 2007; 356(8):769–71. 10.1056/NEJMp078013 [DOI] [PubMed] [Google Scholar]
- 4.Duffy MR, Chen TH, Hancock WT, Powers AM, Kool JL, Lanciotti RS, et al. Zika Virus Outbreak on Yap Island, Federated States of Micronesia. New England J Med. 2009; 360(24):2536–43. [DOI] [PubMed] [Google Scholar]
- 5.Fauci AS, Morens DM. Zika Virus in the Americas—Yet Another Arbovirus Threat. N Engl J Med. 2016; 374(7):601–4. 10.1056/NEJMp1600297 [DOI] [PubMed] [Google Scholar]
- 6.Sim S, Jupatanakul N, Dimopoulos G. Mosquito immunity against arboviruses. Viruses. 2014; 6(11):4479–504. 10.3390/v6114479 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bennett KE, Olson KE, Munoz Mde L, Fernandez-Salas I, Farfan-Ale JA, Higgs S, et al. Variation in vector competence for dengue 2 virus among 24 collections of Aedes aegypti from Mexico and the United States. Am J Trop Med Hyg. 2002; 67(1):85–92. [DOI] [PubMed] [Google Scholar]
- 8.Khoo CC, Piper J, Sanchez-Vargas I, Olson KE, Franz AW. The RNA interference pathway affects midgut infection- and escape barriers for Sindbis virus in Aedes aegypti. BMC Microbiol. 2010; 10:130 10.1186/1471-2180-10-130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Paulson SL, Grimstad PR, Craig GB Jr. Midgut and salivary gland barriers to La Crosse virus dissemination in mosquitoes of the Aedes triseriatus group. Med Vet Entomol. 1989; 3(2):113–23. [DOI] [PubMed] [Google Scholar]
- 10.Sinden RE, Billingsley PF. Plasmodium invasion of mosquito cells: hawk or dove? Trends Parasitol. 2001; 17(5):209–11. [DOI] [PubMed] [Google Scholar]
- 11.Smith RC, Vega-Rodriguez J, Jacobs-Lorena M. The Plasmodium bottleneck: malaria parasite losses in the mosquito vector. Memorias Do Instituto Oswaldo Cruz. 2014; 109(5):644–61. 10.1590/0074-0276130597 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ramirez JL, Souza-Neto J, Torres Cosme R, Rovira J, Ortiz A, Pascale JM, et al. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl Trop Dis. 2012; 6(3):e1561 10.1371/journal.pntd.0001561 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Apte-Deshpande A, Paingankar M, Gokhale MD, Deobagkar DN. Serratia odorifera a midgut inhabitant of Aedes aegypti mosquito enhances its susceptibility to Dengue-2 virus. PLoS ONE. 2012; 7(7):e40401 10.1371/journal.pone.0040401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ramirez JL, Short SM, Bahia AC, Saraiva RG, Dong Y, Kang S, et al. Chromobacterium Csp_P reduces malaria and dengue infection in vector mosquitoes and has entomopathogenic and in vitro anti-pathogen activities. PLoS Pathog. 2014; 10(10):e1004398 Epub 2014/10/24. 10.1371/journal.ppat.1004398 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Cirimotich CM, Dong Y, Clayton AM, Sandiford SL, Souza-Neto JA, Mulenga M, et al. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science. 2011; 332(6031):855–858. 10.1126/science.1201618 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dong Y, Manfredini F, Dimopoulos G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009; 5(5):e1000423 10.1371/journal.ppat.1000423 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Bando H, Okado K, Guelbeogo WM, Badolo A, Aonuma H, Nelson B, et al. Intra-specific diversity of Serratia marcescens in Anopheles mosquito midgut defines Plasmodium transmission capacity. Scientific Rep. 2013; 3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Xi Z, Ramirez JL, Dimopoulos G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008; 4(7):e1000098 10.1371/journal.ppat.1000098 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Wang S, Ghosh AK, Bongio N, Stebbings KA, Lampe DJ, Jacobs-Lorena M. Fighting malaria with engineered symbiotic bacteria from vector mosquitoes. Proc Natl Acad Sci U S A. 2012; 109(31):12734–9. 10.1073/pnas.1204158109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Riehle MA, Moreira CK, Lampe D, Lauzon C, Jacobs-Lorena M. Using bacteria to express and display anti-Plasmodium molecules in the mosquito midgut. Int J Parasitol. 2007; 37(6):595–603. 10.1016/j.ijpara.2006.12.002 [DOI] [PubMed] [Google Scholar]
- 21.Ricci I, Valzano M, Ulissi U, Epis S, Cappelli A, Favia G. Symbiotic control of mosquito borne disease. Path Global Health. 2012; 106(7):380–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Osei-Poku J, Mbogo CM, Palmer WJ, Jiggins FM. Deep sequencing reveals extensive variation in the gut microbiota of wild mosquitoes from Kenya. Mol Ecol. 2012; 21(20):5138–50. 10.1111/j.1365-294X.2012.05759.x [DOI] [PubMed] [Google Scholar]
- 23.Muturi EJ, Kim CH, Bara J, Bach EM, Siddappaji MH. Culex pipiens and Culex restuans mosquitoes harbor distinct microbiota dominated by few bacterial taxa. Parasit Vectors. 2016; 9(1):18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Wang Y, Gilbreath TM 3rd, Kukutla P, Yan G, Xu J. Dynamic gut microbiome across life history of the malaria mosquito Anopheles gambiae in Kenya. PLoS ONE. 2011; 6(9):e24767 10.1371/journal.pone.0024767 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Boissiere A, Tchioffo MT, Bachar D, Abate L, Marie A, Nsango SE, et al. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012; 8(5):e1002742 10.1371/journal.ppat.1002742 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zouache K, Michelland RJ, Failloux AB, Grundmann GL, Mavingui P. Chikungunya virus impacts the diversity of symbiotic bacteria in mosquito vector. Mol Ecol. 2012; 21(9):2297–309. 10.1111/j.1365-294X.2012.05526.x [DOI] [PubMed] [Google Scholar]
- 27.Muturi EJ, Bara JJ, Rooney AP, Hansen AK. Midgut fungal and bacterial microbiota of Aedes triseriatus and Aedes japonicus shift in response to La Crosse virus infection. Mol Ecol. 2016; 25(16):4075–90. 10.1111/mec.13741 [DOI] [PubMed] [Google Scholar]
- 28.Darsie RFJ, Ward RA. Identification and geographical distribution of the mosquitoes of North America, north of Mexico. Mosq Syst Suppl. 1981; 1:1–313. [Google Scholar]
- 29.Sanogo YO, Kim CH, Lampman R, Novak RJ. A real-time TaqMan polymerase chain reaction for the identification of Culex vectors of West Nile and Saint Louis encephalitis viruses in North America. Am J Trop Med Hyg. 2007; 77(1):58–66. [PubMed] [Google Scholar]
- 30.Jeraldo P, Kalari K, Chen X, Bhavsar J, Mangalam A, White B, et al. IM-TORNADO: a tool for comparison of 16S reads from paired-end libraries. PLoS One. 2014; 9(12):e114804 10.1371/journal.pone.0114804 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Lohse M, Bolger AM, Nagel A, Fernie AR, Lunn JE, Stitt M, et al. RobiNA: a user-friendly, integrated software solution for RNA-Seq-based transcriptomics. Nucleic Acids Res. 2012; 40(Web Server issue):W622–7. 10.1093/nar/gks540 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Edgar RC. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat Methods. 2013; 10(10):996–998. 10.1038/nmeth.2604 [DOI] [PubMed] [Google Scholar]
- 33.Cole JR, Wang Q, Fish JA, Chai B, McGarrell DM, Sun Y, et al. Ribosomal Database Project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 2014; 42(Database issue):D633–42. 10.1093/nar/gkt1244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bokulich NA, Subramanian S, Faith JJ, Gevers D, Gordon JI, Knight R, et al. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat Methods. 2013; 10(1):57–9. 10.1038/nmeth.2276 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, et al. QIIME allows analysis of high-throughput community sequencing data. Nat Methods. 2010; 7(5):335–6. Epub 2010/04/13. 10.1038/nmeth.f.303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Oksanen J, Blanchet FG, Kindt R, Legandre P, Minchin PR, O'Hara RB. vegan: Community ecology package. https://CRANR-projectorg/package=vegan. 2013.
- 37.Hammer O, Harper DAT, Ryan PD. PAST: Paleontological statistics software package for education and data analysis. Paleontologia Electronica. 2001; 4:4–9. [Google Scholar]
- 38.Zouache K, Raharimalala FN, Raquin V, Tran-Van V, Raveloson LH, Ravelonandro P, et al. Bacterial diversity of field-caught mosquitoes, Aedes albopictus and Aedes aegypti, from different geographic regions of Madagascar. FEMS Microbiol Ecol. 2011; 75(3):377–89. 10.1111/j.1574-6941.2010.01012.x [DOI] [PubMed] [Google Scholar]
- 39.Engel P, Moran NA. The gut microbiota of insects—diversity in structure and function. FEMS Microbiol Rev. 2013; 37(5):699–735. 10.1111/1574-6976.12025 [DOI] [PubMed] [Google Scholar]
- 40.Chandel K, Mendki MJ, Parikh RY, Kulkarni G, Tikar SN, Sukumaran D, et al. Midgut microbial community of Culex quinquefasciatus mosquito populations from India. PLoS ONE. 2013; 8(11). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Demaio J, Pumpuni CB, Kent M, Beier JC. The midgut bacterial flora of wild Aedes triseriatus, Culex pipiens, and Psorophora columbiae mosquitoes. Am J Trop Med Hyg. 1996; 54(2):219–23. [DOI] [PubMed] [Google Scholar]
- 42.Dillon RJ, Webster G, Weightman AJ, Charnley AK. Diversity of gut microbiota increases with aging and starvation in the desert locust. Antonie Van Leeuwenhoek Inter J Gen Mol Microbiol. 2010; 97(1):69–77. [DOI] [PubMed] [Google Scholar]
- 43.Stathopoulos S, Neafsey DE, Lawniczak MK, Muskavitch MA, Christophides GK. Genetic dissection of Anopheles gambiae gut epithelial responses to Serratia marcescens. PLoS Pathog. 2014; 10(3):e1003897 10.1371/journal.ppat.1003897 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Dodson BL, Hughes GL, Paul O, Matacchiero AC, Kramer LD, Rasgon JL. Wolbachia enhances West Nile Virus (WNV) infection in the mosquito Culex tarsalis. PLOSs Negl Trop Dis. 2014; 8(7). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hughes GL, Rivero A, Rasgon JL. Wolbachia can enhance Plasmodium infection in mosquitoes: implications for malaria control? PLoS Pathog. 2014; 10(9). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Cirimotich CM, Ramirez JL, Dimopoulos G. Native microbiota shape insect vector competence for human pathogens. Cell Host Microbe. 2011; 10(4):307–10. 10.1016/j.chom.2011.09.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Pumpuni CB, Beier MS, Nataro JP, Guers LD, Davis JR. Plasmodium falciparum: inhibition of sporogonic development in Anopheles stephensi by gram-negative bacteria. Exp parasitol. 1993; 77(2):195–9. 10.1006/expr.1993.1076 [DOI] [PubMed] [Google Scholar]
- 48.Hilgenboecker K, Hammerstein P, Schlattmann P, Telschow A, Werren JH. How many species are infected with Wolbachia?—a statistical analysis of current data. FEMS Microbiol Lett. 2008; 281(2):215–20. 10.1111/j.1574-6968.2008.01110.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Werren JH, Baldo L, Clark ME. Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol. 2008; 6(10):741–51. 10.1038/nrmicro1969 [DOI] [PubMed] [Google Scholar]
- 50.Sanogo YO, Dobson SL, Bordenstein SR, Novak RJ. Disruption of the Wolbachia surface protein gene wspB by a transposable element in mosquitoes of the Culex pipiens complex (Diptera, Culicidae). Insect Mol Biol. 2007; 16(2):143–54. 10.1111/j.1365-2583.2006.00707.x [DOI] [PubMed] [Google Scholar]
- 51.Atyame CM, Delsuc F, Pasteur N, Weill M, Duron O. Diversification of Wolbachia endosymbiont in the Culex pipiens mosquito. Mol Biol Evol. 2011; 28(10):2761–72. 10.1093/molbev/msr083 [DOI] [PubMed] [Google Scholar]
- 52.Atyame CM, Duron O, Tortosa P, Pasteur N, Fort P, Weill M. Multiple Wolbachia determinants control the evolution of cytoplasmic incompatibilities in Culex pipiens mosquito populations. Mol Ecol. 2011; 20(2):286–98. 10.1111/j.1365-294X.2010.04937.x [DOI] [PubMed] [Google Scholar]
- 53.Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu GJ, Pyke AT, Hedges LM, et al. A Wolbachia symbiont in Aedes aegypti limits infection with Dengue, Chikungunya, and Plasmodium. Cell. 2009; 139(7):1268–78. 10.1016/j.cell.2009.11.042 [DOI] [PubMed] [Google Scholar]
- 54.Mousson L, Zouache K, Arias-Goeta C, Raquin V, Mavingui P, Failloux AB. The native Wolbachia symbionts limit transmission of dengue virus in Aedes albopictus. PLoS Negl Trop Dis. 2012; 6(12). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Walker T, Johnson PH, Moreira LA, Iturbe-Ormaetxe I, Frentiu FD, McMeniman CJ, et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature. 2011; 476(7361):450–3. 10.1038/nature10355 [DOI] [PubMed] [Google Scholar]
- 56.Iturbe-Ormaetxe I, Walker T, SL ON. Wolbachia and the biological control of mosquito-borne disease. EMBO Rep. 2011; 12(6):508–18. 10.1038/embor.2011.84 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Rasgon JL, Styer LM, Scott TW. Wolbachia-induced mortality as a mechanism to modulate pathogen transmission by vector arthropods. J Med Entomol. 2003; 40(2):125–32. [DOI] [PubMed] [Google Scholar]
- 58.Atyame CM, Pasteur N, Dumas E, Tortosa P, Tantely ML, Pocquet N, et al. Cytoplasmic incompatibility as a means of controlling Culex pipiens quinquefasciatus mosquito in the islands of the South-Western Indian Ocean. PLoS Negl Trop Dis. 2011; 5(12). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zele F, Nicot A, Berthomieu A, Weill M, Duron O, Rivero A. Wolbachia increases susceptibility to Plasmodium infection in a natural system. Proc Roy Soc B-Biol Scies. 2014; 281(1779). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Glaser RL, Meola MA. The Native Wolbachia endosymbionts of Drosophila melanogaster and Culex quinquefasciatus increase host resistance to West Nile Virus infection. PLoS One. 2010; 5(8). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hughes GL, Vega-Rodriguez J, Xue P, Rasgon JL. Wolbachia Strain wAlbB enhances infection by the rodent malaria parasite Plasmodium berghei in Anopheles gambiae mosquitoes. App Environ Microbiol. 2012; 78(5):1491–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kersters K, Lisdiyanti P, Komagata K, Swings J. The Family Acetobacteraceae: the Genera Acetobacter, Acidomonas, Asaia, Gluconacetobacter, Gluconobacter, and Kozakia In: Dworkin M, Falkow S, Rosenberg E, Schleifer K-H, Stackebrandt E, editors. The Prokaryotes: Volume 5: Proteobacteria: Alpha and Beta Subclasses. New York, NY: Springer New York; 2006. p. 163–200. [Google Scholar]
- 63.Crotti E, Damiani C, Pajoro M, Gonella E, Rizzi A, Ricci I, et al. Asaia, a versatile acetic acid bacterial symbiont, capable of cross-colonizing insects of phylogenetically distant genera and orders. Environ Microbiol. 2009; 11(12):3252–64. 10.1111/j.1462-2920.2009.02048.x [DOI] [PubMed] [Google Scholar]
- 64.Crotti E, Rizzi A, Chouaia B, Ricci I, Favia G, Alma A, et al. Acetic acid bacteria, newly emerging symbionts of insects. Appl Environ Microbiol. 2010; 76(21):6963–70. 10.1128/AEM.01336-10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Suzuki R, Zhang Y, Iino T, Kosako Y, Komagata K, Uchimura T. Asaia astilbes sp. nov., Asaia platycodi sp. nov., and Asaia prunellae sp. nov., novel acetic acid bacteria isolated from flowers in Japan. J Gen Appl Microbiol. 2010; 56(4):339–46. [DOI] [PubMed] [Google Scholar]
- 66.Malimas T, Yukphan P, Takahashi M, Kaneyasu M, Potacharoen W, Tanasupawat S, et al. Asaia lannaensis sp nov., a new acetic acid bacterium in the Alphaproteobacteria. Bioscie Biotech Biochem. 2008; 72(3):666–71. [DOI] [PubMed] [Google Scholar]
- 67.Yamada Y, Katsura K, Kawasaki H, Widyastuti Y, Saono S, Seki T, et al. Asaia bogorensis gen. nov., sp. nov., an unusual acetic acid bacterium in the alpha-Proteobacteria. Int J Syst Evol Microbiol. 2000; 50 Pt 2:823–9. [DOI] [PubMed] [Google Scholar]
- 68.Favia G, Ricci I, Damiani C, Raddadi N, Crotti E, Marzorati M, et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. Proc Natl Acad Sci U S A. 2007; 104(21):9047–51. 10.1073/pnas.0610451104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chouaia B, Rossi P, Epis S, Mosca M, Ricci I, Damiani C, et al. Delayed larval development in Anopheles mosquitoes deprived of Asaia bacterial symbionts. BMC Microbiology. 2012; 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Grice EA, Segre JA. The skin microbiome. Nat Rev Microbiol. 2011; 9(4):244–53. 10.1038/nrmicro2537 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Vyas D, McGeough EJ, McGinn SM, McAllister TA, Beauchemin KA. Effect of Propionibacterium spp. on ruminal fermentation, nutrient digestibility, and methane emissions in beef heifers fed a high-forage diet. J Animal Sci. 2014; 92(5):2192–201. [DOI] [PubMed] [Google Scholar]
- 72.Vyas D, McGeough EJ, Mohammed R, McGinn SM, McAllister TA, Beauchemin KA. Effects of Propionibacterium strains on ruminal fermentation, nutrient digestibility and methane emissions in beef cattle fed a corn grain finishing diet. Animal. 2014; 8(11):1807–15. 10.1017/S1751731114001657 [DOI] [PubMed] [Google Scholar]
- 73.Chandler JA, Liu RM, Bennett SN. RNA shotgun metagenomic sequencing of northern California (USA) mosquitoes uncovers viruses, bacteria, and fungi. Frontiers in Microbiol. 2015; 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.O'Hara CM, Brenner FW, Miller JM. Classification, identification, and clinical significance of Proteus, Providencia, and Morganella. Clinical Microbiol Rev. 2000; 13(4):534-+. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Klowden MJ. Making generalizations about vectors: is there a physiology of "the mosquito"? Entomol Res. 2007; 37:1–13. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Families with abundance of less than 1.2% were pooled together as “Other”. S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL. T = date of collection; T1 = July, 2, 2015; T2 = July, 7, 2015; T3 = July, 21, 2015; T4 = July, 28, 2015; T5 = August, 3, 2015; T6 = August, 11, 2015; T7 = August, 19, 2015; T8 = August, 28, 2015; T9 = September, 4, 2015; and T10 = October, 15, 2015.
(TIFF)
CRU = An. crucians, PUN = An. punctipennis, QUA = An. quadrimaculatus, ALB = Ae. albopictus, JAP = Ae. japonicus, TRIS = Ae. triseriatus, VEX = Ae. vexans, TRIV = Ae. trivittatus, PIP = Cx. pipiens, RES = Cx. restuans, INO = Cs. inornata, FER = Ps. ferox. S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL. T = date of collection; T1 = July, 2, 2015; T2 = July, 7, 2015; T3 = July, 21, 2015; T4 = July, 28, 2015; T5 = August, 3, 2015; T6 = August, 11, 2015; T7 = August, 19, 2015; T8 = August, 28, 2015; T9 = September, 4, 2015; and T10 = October, 15, 2015.
(TIF)
CRU = An. crucians, PUN = An. punctipennis, QUA = An. quadrimaculatus, ALB = Ae. albopictus, JAP = Ae. japonicus, TRIS = Ae. triseriatus, VEX = Ae. vexans, TRIV = Ae. trivittatus, PIP = Cx. pipiens, RES = Cx. restuans, INO = Cs. inornata, FER = Ps. ferox. Number at the end of the genus name is the OTU number. Values in the heatmap cells represent the relative abundance of respective OTUs in different mosquito species.
(TIF)
S = trap site, S1 = 504 S Lynn St, Champaign IL; S2 = 1509 Grandview Dr, Champaign IL; S3 = 408 W Maple St, Champaign IL; S4 = 1809 Grandview Dr, Champaign IL; S5 = 805 S. Prairie St, Champaign IL; S6 = 1116 Charles St, Champaign IL; S7 = 604 W Hill St, Champaign IL; S8 = 1605 Coronado Dr, Champaign IL; S9 = 1413 S Western Ave, Champaign IL; S10 = 909 S McKinley, Champaign IL; S11 = 1418 S Western Ave, Champaign IL; S12 = 409 W Hill St, Champaign IL; S13 = 705 W Kirby Ave, Champaign IL; S14 = 602 Nevada St, Urbana IL; S15 = 602 Vermont Ave, Urbana IL; S16 = 2402 E Elm St, Urbana IL; S17 = 804 W Main St, Urbana IL; S18 = 807 W Main St, Urbana IL; S 19 = 2010 Morrow Ct, Urbana IL. T = date of collection; T1 = July, 2, 2015; T2 = July, 7, 2015; T3 = July, 21, 2015; T4 = July, 28, 2015; T5 = August, 3, 2015; T6 = August, 11, 2015; T7 = August, 19, 2015; T8 = August, 28, 2015; T9 = September, 4, 2015; and T10 = October, 15, 2015.
(TIF)
(DOCX)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files except for the 16S rRNA gene bacterial sequences which are available from Genbank under the accession number PRJNA374733.