

Abstract
Background
Toll-interacting protein is a negative regulator in the TLR signaling cascade, particularly by impeding the TLR2 and, TLR4 pathway. Recently, TOLLIP was shown to regulate human TLR signaling pathways. Two common TOLLIP polymorphisms (rs5743899 and rs3750920) were reported to be influencing IL-6, TNF and IL-10 expression. In this study, TOLLIP variants were investigated to their relation to Plasmodium vivax malaria in the Brazilian Amazon.
Methods
This cohort study was performed in the municipalities of Careiro and, Manaus, in Western Brazilian Amazon. A total of 319 patients with P. vivax malaria and, 263 healthy controls with no previous history of malaria were included in the study. Genomic DNA was extracted from blood collected on filter paper, using the QIAamp® DNA Mini Kit, according to the manufacturer’s suggested protocol. The rs5743899 and rs3750920 polymorphisms of the TOLLIP gene were typed by PCR–RFLP.
Results
Homozygous individuals for the rs3750920 T allele gene had twice the risk of developing malaria when compared to individuals homozygous for the C allele (OR 2.0 [95% CI 1.23–3.07]; p = 0.004). In the dominant model, carriers the C allele indicates protection to malaria, carriers of the C allele were compared to individuals with the T allele, and the difference is highly significant (OR 0.52 [95% CI 0.37–0.76]; p = 0.0006). The linkage disequilibrium between the two polymorphisms was weak (r2 = 0.037; D′ = 0.27).
Conclusions
These findings suggest that genes involved in the TLRs-pathway may be involved in malaria susceptibility. The association of the TOLLIP rs3750920 T allele with susceptibility to malaria further provides evidence that genetic variations in immune response genes may predispose individuals to malaria.
Keywords: Malaria, Plasmodium vivax, TOLLIP gene, Amazon
Background
Malaria, a health burden in the Amazon region of Brazil, is responsible for 99.9% of all reported malarial cases in Brazil and nearly 140,000 cases were registered the region in 2015 [1]. The socioeconomic and, environmental conditions in the Amazon favor the proliferation of the vector, the mosquito Anopheles darlingi. In the State of Amazonas, 75,575 cases were reported in 2015. Plasmodium vivax accounts for 90% and, Plasmodium falciparum is responsible for the majority of the remaining cases [2]. Plasmodium vivax, in contrast to P. falciparum, has a dormant form in the liver, the hypnozoite, which subsequently causes new infections in the blood known as relapses, and this represents a real challenge for vivax malaria eradication [3].
Several studies have linked high levels of TNF, IL-1β, IL-2, IL-4, IL-6, IL-8, IL-10 and IL-12 cytokines with P. vivax infections [4–7]. Severe malaria patients display increase levels of pro-inflammatory cytokines, TNF, IL-6 and IFN-γ [8–11] while acute malaria individuals show high levels of the anti-inflammatory cytokines IL10 [12–14].
Innate immunity is defined as the first line of host defense to invading pathogens. Toll-like receptors recognize invading pathogens and, TLR-2 and, TLR-4 sense the glycosylphosphatidylinositol (GPI) of Plasmodium [15]. Mice immunized with synthesized GPI and challenged with Plasmodium berghei are protected against malaria-related acidosis and, pulmonary oedema, suggesting that GPI may contribute to malaria mortality and, pathogenesis [16]. In vitro, GPI influences the expression of adhesion molecules and, the pro-inflammatory cytokines IL-1 and TNF [17].
The above studies show the importance of the TLR pathway in Plasmodium-infection. Overexpression of Toll-interacting protein (TOLLIP) leads to the impairment of the activation of the transcription factor NF-κB and, thus limits the production of pro-inflammatory mediators [18]. TOLLIP is also suggested to inhibit TLR-mediated cellular responses by associating directly with TLR2 and, TLR4 to suppress the phosphorylation and kinase activity of IL-1 receptor associated Kinase [19]. TOLLIP skews the pro-inflammatory cytokines response of TLR-2 and, TLR-4 into an increased IL-10 and, decreased IL-6 expression in human peripheral blood monocytes [20]. IL-6 and TNF are key mediators associated with malarial symptoms and their levels are proportional to the severity of the disease [21]. Several variants of the TOLLIP gene have been shown to be associated with tuberculosis and its transcription levels [20], cutaneous leishmaniasis [22] and, leprosy [23].
The role played by the TLR pathway in keeping in check parasite multiplication in animal models and the suggestion that TOLLIP gene may downregulate TLR-2 and, TLR-4 lead us to hypothesize that variants of the TOLLIP gene may be associated with malaria caused by P. vivax in the state of Amazonas, Brazil. To this end, a case–control study was conducted by investigating two common variants rs5743899 and rs3750920 that were previously reported to probably influence the expression of TNF, IL-6 and, IL-10 in patients with malaria and healthy controls from the same endemic areas followed for a period of 12 months. The authors report that the TOLLIP rs3750920C allele is associated with protection to malaria while the T allele with susceptibility.
Methods
Site of study and patients
The present study was conducted from a cohort studies in the municipality of Careiro (in the central region of the Amazon State) and, in the peri-urban region of Manaus (Brasileirinho, Ipiranga and Puraquequara communities), the capital city of the Amazonas state, Brazil. These regions have been invaded over the years by destroying the forest for settlements, agriculture and, farming.
Manaus has a population of approximately 2,020,301 inhabitants. Most of them live in urban/peri-urban area where there has been an intense migration process, combined with precarious epidemiological and, entomological surveillance resulting in the active transmission of malaria. The IPA in the city ranges from low to medium risk in rural areas and, between the peri-urban areas varies from no risk to high risk.
The municipality has an average of 75% of malaria caused by P. vivax reported in the last 5 years, which is very similar to the percentage of infections by P. vivax reported in other areas of the Brazilian Amazon. In the last 5 years there has been some seasonality to malaria despite the stable transmission, with most reported cases during the period from May to September.
The communities of Brasileirinho, Ipiranga and, some extension Puraquequara, are located in the peri-urban area, east of the city of Manaus. The city’s boundary is surrounded with forest and has suffered deforestation and, gradually becomes endemic areas of malaria. In the city of Manaus, these areas have become vulnerable to malaria transmission, despite the decline in IPA observed in recent years. The subsistence activities of the population are concentrated in the agricultural sector and, extraction. Many of the residents of these areas, however, work in the city of Manaus and, thus travel daily to Manaus. It is noteworthy that in this location there are several farms used for recreation and, religious retreat, where many residents of Manaus go to spend their weekends and, holidays. These fluxes can increase the risk of malaria transmission. The occurrence of malaria in this population is common. Basic sanitary is lacking in these communities. There is no garbage collection. Drinking water is from wells or streams.
The Municipality of Careiro currently has an estimated population of 30,000 inhabitants, most of them living in rural areas, supported by federal programs that encourage the practice of agriculture. This population has been previously described elsewhere [24].
These areas have become endemic particularly for P. vivax. These populations were chosen for study based on low migration of its inhabitants (intra-regional) and, similar profile of rural malaria transmission. All of the case patients included in the study are patients with symptomatic malaria, diagnosed in the health post of municipalities and, are provided with malarial treatments following detection of the parasites.
A total of 319 patients with malaria confirmed by direct microscope examination of Giemsa-stained specimens for the presence of P. vivax parasites and, 263 healthy controls with no history of malaria are included in the study. Basic characteristics of the study population are shown in Table 2.
Table 2.
Genotype and allele frequencies for the single nucleotide polymorphisms rs5743899 and, rs3750920 in patients with and, without P. vivax malaria
| Polymorphism, genotypes and, alleles | P. vivax malaria patients | Controls |
|---|---|---|
| n = 301 (%) | n = 255 (%) | |
| TOLLIP rs5743899 | ||
| GG | 32 (11) | 24 (10) |
| GA | 124 (41) | 108 (42) |
| AA | 145 (48) | 123 (48) |
| G | 188 (31) | 156 (31) |
| A | 414 (69) | 354 (69) |
| Polymorphism, genotypes and, alleles | P. vivax malaria patients | Controls |
|---|---|---|
| n = 319 (%) | n = 263 (%) | |
| TOLLIP rs3750920 | ||
| CC | 66 (21) | 68 (26) |
| CT | 134 (42) | 132 (50) |
| TT | 119 (37) | 63 (24) |
| C | 133 (42) | 134 (51) |
| T | 186 (58) | 129 (49) |
| Polymorphism, genotypes and, alleles | P. vivax malaria patients | Controls |
|---|---|---|
| p value | OR [95% CI] | |
| Genotypes and alleles comparisons TOLLIP rs3750920 | ||
| CC vs. TT | 0.004 | 1.95 [1.23–3.07] |
| CC + CT vs. TT | 0.0005 | 0.52 [0.37–0.76] |
| TT vs. CT | 0.002 | 0.53 [0.36–0.79] |
| T vs. C | 0.002 | 0.69 [0.55–0.87] |
Molecular characterization
Genomic DNA was extracted from blood collected on filter paper, using the QIAamp® DNA Mini Kit (QIAGEN®, Germany), according to the manufacturer’s suggested protocol.
Genotypic determinations of the rs5743899 and rs3750920 located in the intron and exon 4 respectively of the TOLLIP gene were performed by PCR–RFLP as described elsewhere [22]. Briefly, the following pairs of primers: rs5743899F: 5′-GGC AAT GGC AGT GGC CAC CAG TGA-3′ and rs5743899R: 5′-CCG ATGCCC GCA CAC CTG TGT GAT-3′ for (rs5743899) and rs3750920F: 5′-AGG CGT GCA GCTCAC CGC GTA GGA-3′ and rs3750920R: 5′-GAG AGC CTT CTC CAT GGA CGA CCG C-3′ for (rs3750920) flanking the polymorphisms, were used to amplify separately a stretch of DNA of 279 and 169 bp respectively and, digested by the corresponding restriction enzymes HhaI and, MspI, with the fragment-size separated by electrophoresis in 3% agarose gel (Fig. 1).
Fig. 1.
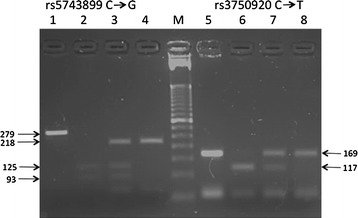
The rs5743899 and, rs3750920 polymorphisms of the TOLLIP gene as detected by restriction enzymes HhaI and, MspI respectively. The gel depicts the PCR product (lane 1), restriction patterns related to homozygozity GG (lane 2), heterozygozity GA (lane 3) and, homozygosity AA (lane 4) for the rs5743899 and, similarly for the rs3750920, PCR product (lane 5), homozygote CC (lane 6), heterozygote CT (lane 7) and, homozygote TT (lane 6). Lane M, DNA molecular-weight marker ladder 50 base pairs
Data analysis
Statistical analysis was performed using the website http://ihg.gsf.de/cgibin/hw/hwa1.pl for logistic regression analysis. Two-tailed χ2 test along with the odds ratio (OR) and, 95% confidence interval [CI] was applied for comparison of patients with malaria to healthy controls. Allelic and genotypic frequencies were calculated by gene counting directly. Hardy–Weinberg equilibrium (HWE) was calculated by χ2 test by comparing the observed number of different genotypes frequency with those expected under HW expectation. The software Haploview 4.2 was used for calculating Linkage disequilibrium between the rs5743899 and, rs3750920.
Results
Previous study of surveillance in the Careiro, Brasileirinho, Ipiranga and, Puraquequara communities showed that a significant proportion of the population reported previous infection with P. vivax and/or P. falciparum, indicating that most individuals were exposed to the malaria parasite along the years. The baseline characteristics of the individuals participating in this study are shown in Table 1. A total of 319 had malaria and, 263 healthy controls were recruited from the same endemic area. Approximately 60% were male with a mean age of 38 years [Careiro: 35.9 (SD 17.8); Brasileirinho + Ipiranga + Puraquequara: 39.5 (SD 21.3)]. Among individuals with malaria, male has a higher frequency of malaria. Most of them reported previous history of the disease (Careiro: 65%; Brasileirinho + Ipiranga + Puraquequara: 84%), with an average of five episodes in Careiro and, ten episodes in Brasileirinho + Ipiranga + Puraquequara.
Table 1.
Clinical and demographic characteristics of patients with and without P. vivax malaria
| Clinical and demographic characteristics | P. vivax malaria patients | Controls |
|---|---|---|
| n = 111 | N = 97 | |
| Careiro | ||
| Gender | ||
| Male n (%) | 63 (57) | 61 (63) |
| Female n (%) | 48 (43) | 36 (37) |
| Age/years (mean ± SD) | 35.9 ± 17.8 | 34.2 ± 16 |
| Previous history of malaria n (%) | 72 (65) | – |
| Episodes of malaria (mean) | 05 | – |
| Plasmodium vivax (%) | 81 | – |
| Plasmodium falciparum (%) | 19 | – |
| Clinical and demographic characteristics | P. vivax malaria patients | Controls |
|---|---|---|
| n = 208 | n = 166 | |
| Brasileirinho + Ipiranga + Puraquequara | ||
| Gender | ||
| Male n (%) | 131 (63) | 84 (51) |
| Female n (%) | 77 (37) | 82 (49) |
| Age/years (mean ± SD) | 39.5 ± 21.3 | 39.5 ± 21.3 |
| Previous history of malaria n (%) | 174 (84) | – |
| Episodes of malaria (mean) | 10 | – |
| Plasmodium vivax (%) | 89 | – |
| Plasmodium falciparum (%) | 11 | – |
These individuals were genotyped for the rs5743899 and, rs3750920 of TOLLIP gene. The distribution of genotypes and, allele frequencies of both polymorphisms are shown in Table 2. Both polymorphisms were in Hardy–Weinberg equilibrium among the healthy controls while the rs3750920 deviate slightly among patients with malaria (p = 0.02). No difference was observed for the rs5743899 genotypes and, alleles distributions between patients with malaria and, healthy controls. In contrast, the genotypes distributions of the rs3750920 differed significantly (p = 0.0024). Among the patients with malaria, homozygosity for the T allele was prevalent 37 vs. 24% and, had twice the risk of developing malaria when compared to individuals homozygous for the C allele (p = 0.004; OR 2.0 [95% CI 1.23–3.07]). The C allele indicates protection to malaria. In the dominant model, carriers of the C allele were compared to individuals with the T allele and, the difference is highly significant (p = 0.0006; OR 0.52 [95% CI0.37–0.76]). Comparison of heterozygous individuals (CT) to homozygous individuals for the T allele also showed a similar trend (p = 0.002; OR: 0.54 [95% CI 0.36–0.80]). The linkage disequilibrium between the two polymorphisms is weak as calculated by the Haploview 4.2 program. The r2 and, D′ are 0.037 and, 0.27 respectively.
The study examined whether the genotypes of both SNPs were correlated with parasitaemia. Parasitaemia was determined as elsewhere by quantitative Real-Time PCR [25]. Parasites loads were available for only 205 patients with P. vivax-infection and ranges from 1 to 158,777 copies/µL. Low parasitaemia was considered to be equal or below to the median (183 copies/µL). None of the genotypes correlated to either low or high parasitaemia as shown in Table 3.
Table 3.
Analysis of association of the genotype of polymorphism in TOLLIP and studied with variables in Plasmodium vivax infection
| Polymorphism and, genotypes | P. vivax malaria patients | OR (IC 95%) | χ2 | (p) value | ||
|---|---|---|---|---|---|---|
| Low parasite load | High parasite load | |||||
| n = 103 (%) | n = 102 (%) | |||||
| TOLLIP 899 (rs3750920) | ||||||
| G/G | 8 (8) | 11 (11) | 0.738 (0.423–1.287) | 1.15 | 0.283 | G/G + G/A vs. A/A |
| G/A | 39 (38) | 28 (27) | 0.522 (0.186–1.466) | 1.55 | 0.213 | G/G vs. G/A |
| A/A | 56 (54) | 63 (62) | 0.638 (0.349–1.168) | 2.13 | 0.144 | G/A vs. A/A |
| TOLLIP rs3750920 | ||||||
| C/C | 35 (34) | 37 (36) | 0.808 (0.434–1.506) | 0.45 | 0.501 | C/C vs. C/T |
| C/T | 48 (47) | 41 (40) | 0.712 (0.345–1.470) | 0.85 | 0.357 | C/T vs. T/T |
| T/T | 20 (19) | 24 (24) | 1.135 (0.535–2.408) | 0.11 | 0.741 | C/C vs. T/T |
Discussion
Despite an increasing amount of research on malaria, the molecular mechanism that influences the clinical outcome of the disease is poorly understood. In endemic areas, only a fraction of Plasmodium-infected individuals progress to clinical manifestations while the rest remains asymptomatic. Many may also have recurrent infections or malarial episodes in contrast to their neighbors. Altogether, these suggest that the host genetics may play an important role in the clinical outcome and, the identification of genes involved in either susceptibility or resistance to Plasmodium-infection is of utmost interest to the molecular understanding of the disease. Vaccine design may take into account the host genetics.
TLRs upon recognizing invading pathogens orchestrate innate immune responses through the induction of chemokines and inflammatory cytokines to check the parasite. Several polymorphisms present in TLRs genes and, genes involved in its pathways are associated with various infectious diseases [26, 27]. Plasmodium GPI is recognized by various cell surface TLRs [15] and, provokes severe malaria symptoms in mice [16]. A perfect control of Plasmodium infection by the host requires a well tune immune response sufficient to restrict the multiplication of the parasite and also to avoid an excess activation of the TLRs intracellular signaling that may lead to an exacerbation of pro-inflammatory cytokines such as TNF, IL-6 and, IFN-γ injuring the host tissue.
Two polymorphisms, rs5743899 and rs3750920, in the TOLLIP gene that is a negative regulator of TLRs signaling, are associated with tuberculosis in a Vietnamese population [20] and with cutaneous leishmaniasis in the Amazonas state, Brazil [22]. The current study shows that rs3750920 but not rs5743899 is associated with malaria. The rs3750920 T allele is here related to susceptibility to malaria and, is in line with the association with cutaneous leishmaniasis [22] and, with leprosy [23, 28] but in contrast to tuberculosis [20].
TOLLIP diverts the pro-inflammatory cytokine response after TLR-2 and, TLR-4 signaling into an anti-inflammatory response that is characterized by an increased IL-10 and, a decreased IL-6 and TNF expression in peripheral blood monocytes [22]. One study showed that the rs3750920 TT genotype is associated with higher levels of TOLLIP RNA compared to the CC genotype but not with levels of IL-10 or IL-6 [23]. Here the study shows that individuals homozygous for the rs3750920 T allele have twice the chance of developing malaria. It is highly plausible that the diversion of the pro-inflammatory to anti-inflammatory response in carriers of the rs3750920 TT genotype may expose the individual to a higher risk as TNF and, IL-6 cytokines are important early in infection to keep the parasite in check. The genotypes of both SNPs did not correlate to parasitaemia. The rs3750920 may probably do not have an influence on the parasites but mostly on cytokines expressions.
This study has several limitations. Firstly, the sample size of the study participants is small and although the level of the association with malaria is high, it needs validation with a larger sample size. Furthermore, it can be argued that the association to malaria may be spurious as this Amazonian population is a miscegenation of African, European and, Amerindian origin. The controls were properly selected to match the patients’ ethnicity and, are from the same area of endemicity as the patients. Moreover, the frequency of the genotypes of rs5743899 which is very close to rs3750920 was similar in both patients and controls ruling out a spurious association. Secondly, there were no patients with different clinical manifestations, such as severe, mild and asymptomatic malaria, to perform intra-comparison to denote which allele is related to severe malaria. Lastly, plasma cytokines levels were not assayed to correlate with the genotypes of the SNPs.
Conclusion
The present findings suggest that genes involved in the TLRs-pathway may be involved in the pathogenesis of malaria. TOLLIP interacts with TLR-2 and, TLR-4 to skew the pro-inflammatory to anti-inflammatory response. The association of the TOLLIP rs3750920 T allele with susceptibility to malaria further provides evidence that genetic variations in immune response genes may predispose individuals to malaria. Further studies are needed in other vivax regions to confirm whether this allele can be used as a prospective genetic marker for vivax malaria in endemic areas in the future.
Authors’ contributions
LWB performed laboratory work and drafted the first version of the manuscript. LRAB and FJA helped in laboratory work. AGC, LDO and SKP contributed in data analysis. ACGA, AK, SVS and GCM were responsible for the collection and organization of field samples. WMM, MVGL and RR participated in study design, coordination and elaborated the final version of manuscript. All authors read and approved the final manuscript.
Acknowledgements
We acknowledge the collaboration of the local microscopists Juscelino Torres dos Santos, Eliana Lima and Irailton dos Santos. We thank Nelida Sanchez for technical support and the study subjects for their participation.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data generated or analysed during this study are included in this published article and its can be made available files.
Consent for publication
All authors declare to consent of the manuscript and agree to the proposed authorship order.
Ethics approval and consent to participate
The studies were approved by the Research Ethics Committee in the Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (CEP/FMT-HVD process #51536/2012) and by the National Commission for Ethics in Brazilian Research (CONEP process #15197/2008, #349211/2013). All participants read and signed the written informed consent form.
Funding
This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (Grant number 575788/2008-9), Fundació Cellex (P. vivax Consortium) and Bill & Mellinda Gates Foundation (TransEPI Consortium). MVGL is a level 1 research fellow from CNPq. RR is a research fellow from FAPEAM (PVS Program—PECTI-AM/PG#019/2013). AGC, FJA, LRAB, LWB and SKP have fellowship from CAPES, CNPq and FAPEAM (PhD, Master and SI students). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Contributor Information
Larissa W. Brasil, Email: larissa_brasil@hotmail.com
Laila R. A. Barbosa, Email: laila_rowena@hotmail.com
Felipe J. de Araujo, Email: felipe_jules@hotmail.com
Allyson G. da Costa, Email: allyson.gui.costa@gmail.com
Luan D. O. da Silva, Email: luandiego.oliveira@gmail.com
Suzana K. Pinheiro, Email: skanawatip@gmail.com
Anne C. G. de Almeida, Email: anne.almeida.gb@gmail.com
Andrea Kuhn, Email: a-kuehn@gmx.eu.
Sheila Vitor-Silva, Email: sheilavitorr@gmail.com.
Gisely C. de Melo, Email: cardosogisely@gmail.com
Wuelton M. Monteiro, Email: wueltonmm@gmail.com
Marcus V. G. de Lacerda, Email: marcuslacerda.br@gmail.com
Rajendranath Ramasawmy, Phone: +55 92 981357425, Email: ramasawm@gmail.com.
References
- 1.Oliveira-Ferreira J, Brasil P, Ladislau JL, Tauil PL, Daniel-Ribeiro CT. Malaria in Brasil: an overview. Malar J. 2010;9:115. doi: 10.1186/1475-2875-9-115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Secretaria de Vigilância em Saúde. The epidemiological situation of malaria in Brazil. 1998. http://portalsaude.saude.gov.br. Accessed 15 Dec 2016.
- 3.Mueller I, Galinski MR, Baird JK, Carlton JM, Kochar DK, Alonso PL, et al. Key gaps in the knowledge of Plasmodium vivax, a neglected human malaria parasite. Lancet Infect Dis. 2009;9:555–566. doi: 10.1016/S1473-3099(09)70177-X. [DOI] [PubMed] [Google Scholar]
- 4.Medina TS, Costa SP, Oliveira MD, Ventura AM, Souza JM, Gomes TF, et al. Increased interleukin-10 and interferon-gamma levels in Plasmodium vivax malaria suggest a reciprocal regulation which is not altered by IL-10 gene promoter polymorphism. Malar J. 2011;10:264. doi: 10.1186/1475-2875-10-264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Karunaweera ND, Carter R, Grau GE, Kwiatkowski D, Del Giudice G, Mendis KN. Tumour necrosis factor-dependent parasite-killing effects during paroxysms in non-immune Plasmodium vivax malaria patients. Clin Exp Immunol. 1992;88:499–505. doi: 10.1111/j.1365-2249.1992.tb06478.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Karunaweera ND, Wijesekera SK, Wanasekera D, Mendis KN, Carter R. The paroxysm of Plasmodium vivax malaria. Trends Parasitol. 2003;19:188–193. doi: 10.1016/S1471-4922(03)00036-9. [DOI] [PubMed] [Google Scholar]
- 7.Zeyrek FY, Kurcer MA, Zeyrek D, Simsek Z. Parasite density and serum cytokine levels in Plasmodium vivax malaria in Turkey. Parasite Immunol. 2006;28:201–207. doi: 10.1111/j.1365-3024.2006.00822.x. [DOI] [PubMed] [Google Scholar]
- 8.Day NP, Hien TT, Schollaardt T, Loc PP, Chuong LV, Chau TT, et al. The prognostic and pathophysiologic role of pro- and antiinflammatory cytokines in severe malaria. J Infect Dis. 1999;180:1288–1297. doi: 10.1086/315016. [DOI] [PubMed] [Google Scholar]
- 9.Lyke KE, Burges R, Cissoko Y, Sangare L, Dao M, Diarra I, et al. Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect Immun. 2004;72:5630–5637. doi: 10.1128/IAI.72.10.5630-5637.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Clark IA, Budd AC, Alleva LM, Cowden WB. Human malarial disease: a consequence of inflammatory cytokine release. Malar J. 2006;5:85. doi: 10.1186/1475-2875-5-85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Andrade BB, Reis-Filho A, Souza-Neto SM, Clarêncio J, Camargo LM, Barral A, et al. Severe Plasmodium vivax malaria exhibits marked inflammatory imbalance. Malar J. 2010;9:13. doi: 10.1186/1475-2875-9-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Peyron F, Burdin N, Ringwald P, Vuillez JP, Rousset F, Banchereau J. High levels of circulating IL-10 in human malaria. Clin Exp Immunol. 1994;95:300–303. doi: 10.1111/j.1365-2249.1994.tb06527.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Couper KN, Blount DG, Riley EM. IL-10: the master regulator of immunity to infection. J Immunol. 2008;180:5771–5777. doi: 10.4049/jimmunol.180.9.5771. [DOI] [PubMed] [Google Scholar]
- 14.Hansen DS, Schofield L. Natural regulatory T cells in malaria: host or parasite allies? PLoS Pathog. 2010;6:1000771. doi: 10.1371/journal.ppat.1000771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Krishnegowda G, Hajjar AM, Zhu J, Douglass EJ, Uematsu S, Akira S, et al. Induction of proinflammatory responses in macrophages by the glycosylphosphatidylinositols of Plasmodium falciparum: cell signaling receptors, glycosylphosphatidylinositol (GPI) structural requirement, and regulation of GPI activity. J Biol Chem. 2005;280:8606–8616. doi: 10.1074/jbc.M413541200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schofield L, Hewitt MC, Evans K, Siomos MA, Seeberger PH. Synthetic GPI as a candidate anti-toxic vaccine in a model of malaria. Nature. 2002;418:785–789. doi: 10.1038/nature00937. [DOI] [PubMed] [Google Scholar]
- 17.Schofield L, Novakovic S, Gerold P, Schwarz RT, McConville MJ, Tachado SD. Glycosylphosphatidylinositol toxin of Plasmodium up-regulates intercellular adhesion molecule-1, vascular cell adhesion molecule-1, and E-selectin expression in vascular endothelial cells and increases leukocyte and parasite cytoadherence via tyrosine kinase-dependent signal transduction. J Immunol. 1996;156:1886–1896. [PubMed] [Google Scholar]
- 18.Burns K, Clatworthy J, Martin L, Martinon F, Plumpton C, Maschera B, et al. Tollip, a new component of the IL-1RI pathway, links IRAK to the IL-1 receptor. Nat Cell Biol. 2000;2:346–351. doi: 10.1038/35014038. [DOI] [PubMed] [Google Scholar]
- 19.Zhang G, Ghosh S. Negative regulation of toll-like receptor-mediated signaling by Tollip. J Biol Chem. 2002;277:7059–7065. doi: 10.1074/jbc.M109537200. [DOI] [PubMed] [Google Scholar]
- 20.Shah JA, Vary JC, Chau TT, Bang ND, Yen NT, Farrar JJ, et al. Human TOLLIP regulates TLR2 and TLR4 signaling and its polymorphisms are associated with susceptibility to tuberculosis. J Immunol. 2012;189:1737–1746. doi: 10.4049/jimmunol.1103541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Goncalves RM, Scopel KK, Bastos MS, Ferreira MU. Cytokine balance in human malaria: does Plasmodium vivax elicit more inflammatory responses than Plasmodium falciparum? PLoS ONE. 2012;7:44394. doi: 10.1371/journal.pone.0044394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.de Araujo FJ, da Silva LD, Mesquita TG, Pinheiro SK, Vital WdS, Chrusciak-Talhari A, et al. Polymorphisms in the TOLLIP gene influence susceptibility to cutaneous leishmaniasis caused by Leishmania guyanensis in the Amazonas State of Brazil. PLoS Negl Trop Dis. 2015;9:000387. doi: 10.1371/journal.pntd.0003875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Shah JA, Berrington WR, Vary JC, Jr, Wells RD, Peterson GJ, Kunwar CB, et al. Genetic variation in toll-interacting protein is associated with leprosy susceptibility and cutaneous expression of interleukin 1 receptor antagonist. J Infect Dis. 2016;213:1189–1197. doi: 10.1093/infdis/jiv570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Vitor-Silva S, Siqueira AM, de Souza Sampaio V, Guinovart C, Reyes-Lecca RC, de Melo GC, et al. Declining malaria transmission in rural Amazon: changing epidemiology and challenges to achieve elimination. Malar J. 2016;15:266. doi: 10.1186/s12936-016-1326-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Perandin F, Manca N, Calderaro A, Piccolo G, Galati L, Ricci L, et al. Development of a real-time PCR assay for detection of Plasmodium falciparum, Plasmodium vivax, and Plasmodium ovale for routine clinical diagnosis. J Clin Microbiol. 2004;42(1214–9):26. doi: 10.1128/JCM.42.3.1214-1219.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schroder NW, Schumann RR. Single nucleotide polymorphisms of Toll-like receptors and susceptibility to infectious disease. Lancet Infect Dis. 2005;5:156–164. doi: 10.1016/S1473-3099(05)70023-2. [DOI] [PubMed] [Google Scholar]
- 27.Turvey SE, Hawn TR. Towards subtlety: understanding the role of Toll-like receptor signaling in susceptibility to human infections. Clin Immunol. 2006;120:1–9. doi: 10.1016/j.clim.2006.02.003. [DOI] [PubMed] [Google Scholar]
- 28.Montoya-Buelna M, Fafutis-Morris M, Tovar-Cuevas AJ, Alvarado-Navarro A, Valle Y, Padilla-Gutierrez JR, et al. Role of toll-interacting protein gene polymorphisms in leprosy Mexican patients. Biomed Res Int. 2013;2013:459169. doi: 10.1155/2013/459169. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated or analysed during this study are included in this published article and its can be made available files.