

ABSTRACT
Discovered decades ago, extracellular vesicles (EVs) emerge as dedicated organelles, able to deliver protected, specific cellular cues throughout the organism. While virtually every cell can release EVs, cancer cells co-opted this feature and efficiently unleashed them both in the tumor microenvironment and toward healthy tissues. This might contribute to tumor aggressiveness and spreading. Cancer-derived EVs that contain DNA, mRNA, miRNA, and packed and transmembrane proteins can operate locally or at distance. This review will focus on the high-grade brain tumor (i.e. glioblastoma)-derived EVs, discussing recent reports on i) their phenotype and content, ii) their putative functions, and iii) their clinical potential for improving diagnosis and therapeutics.
KEYWORDS: angiogenesis, brain tumors, exosomes, glioma, permeability, signaling, tumor microenvironment
Introduction
Although rare, glioblastoma (GBM) represents the most malignant and deadly brain tumor in adults, with a poor prognosis despite intensive researches and clinical efforts.1 GBM exhibit high proliferation rate, invasion, angiogenesis and necrosis because of the diffuse and aggressive nature of the tumor cells. Relapse is almost inevitable and fatal within a short window of 7–10 months, while the average survival does not exceed 18 months following diagnosis. Current standard treatments are essentially palliative, typically involving surgery followed by radiotherapy and DNA-alkylating chemotherapeutic agents to eliminate the remaining cells.1 Meanwhile, a subpopulation of cells with tumor-initiating properties bears resistance to conventional therapies, and as such has been implicated in tumor recurrence.2
Numerous recent studies unveil that tumor cells can efficiently release selected intracellular content in the milieu, embedded and protected in lipid layer-based structures, named extracellular vesicles (EVs). Such tumor-derived EVs may contribute to intercellular communication, tumor progression and resistance mechanisms in the context of GBM.3
Extracellular vesicles in glioblastoma
General definition of the tumor-derived EVs
Extracellular vesicles (EVs) were discovered over 30 years ago and defined as extracellular lipid bilayer spherical structures from 30 to 1000 nm that are secreted in the cellular environment.4,5 An international consortium adopted the generic term of extracellular vesicles (EVs) to clarify between the different names used in the literature.6 The main subcategories of EVs are classified according to their size and their subcellular origin, while their content appears rather versatile. Classically, exosomes exhibit diameters of 30–100 nm and mostly originate from intra-cytoplasmic vesicular bodies; microvesicles (or ectosomes) range between 100 nm to 1 μm diameter and are derived from the plasma membrane; apoptotic blebs are the largest population of EVs, with a diameter of approximately 1–2 μm and emanating from dying cells.7,8 Recently, the term of oncosomes was coined to define large EVs (1 to 2 μm) arising from the budding of the plasma membrane in cancer cells.7,8
Studies in the 1970's reported cell membrane undulations and peripheral vacuoles bordering the extracellular space of GBM giant-cells,9 suggesting that tumor cells could be a non-physiological source of extracellular lipid layer-based structures. More recently, abundant EVs were observed on the surface of primary human GBM cells in culture.10 Interestingly, it has been estimated that one primary GBM cell releases about 10.000 EVs in vitro over a period of 48 hours.10,11 Molecularly, the fact that tumor cells exhibit enhanced EVs release as opposed to normal cells could be intrinsically caused by oncogenic mutations. For instance, the EGF receptor variant III (EGFRvIII), a feature of nearly 50% of adult GBM was shown to augment GBM cell-derived vesiculation.12
The molecular composition of EVs in glioblastoma
Tumor-based EVs can carry a wide range of molecules from nucleic acids (including genomic and mitochondrial DNA, mRNA, miRNA) to proteins and lipids that mirror tumor cells and state.13
In 2008, Skog et al were among the first to report that GBM-derived EVs contain mRNA and miRNA.10 They showed that the EV content is selectively enriched, as compared to the parental cells. Likewise, miRNA can be conveyed within GBM-released EVs.15,16 In that view, miR-21, known for its protective action on GBM cells17 is enriched in GBM-shed EVs.10 Conversely, genomic or mitochondrial DNA were rarely reported in GBM-derived EVs.11,18 Proteins are also specifically sorted and embedded in EVs, although the packaging mechanism is not yet elucidated (for a review, please see ref. 8 and 19). Mass spectrometry analysis of GBM-derived EVs identified more than 100 proteins that are also usually found in exosomes, based on the ExoCarta database.15,20 Interestingly, cytokines are enriched in GBM-produced EVs, such as pro-angiogenic agents, among which VEGF-A,10 the pro-permeability guidance molecule Semaphorin 3A,21 and the immunosuppressive cytokine TGF-β.22 Additionally, one of the best-characterized GBM-based EV-harbored proteins is the tumor-specific EGF mutated receptor EGFRvIII.10,12 Finally, whereas lipids are structural components of EVs, there is a lack of data on lipid composition23 and rare information as for GBM.
An important layer of complexity arose form the heterogeneity of EVs content produced by GBM tumors. Briefly, 4 different GBM subtypes, namely proneural, neural, classical and mesenchymal, have been described according to their molecular signatures.24 Importantly, GBM-derived EV protein content remain clustered within the same subtype of their origin (i.e., with proneural or mesenchymal signatures),25 suggesting that hallmarks of each subtype is conserved and transferred throughout GBM-derived EVs even if the exact cargoes remain unknown. As a consequence, GBM-produced EVs may exhibit the ability to disseminate oncogenic materials.
EV uptake and diffusion
Among the communication network that operates between cancer cells and their environs (i.e secretion, cell-cell junction, mechanical forces, tunneling nanotubes, etc), EVs represent a powerful tool. EVs are released locally into the extracellular tumor microenvironment and are able to spread throughout the organism, as GBM-derived EVs can be found circulating into the bloodstream, urine and cerebrospinal fluid.26,27 The molecular and cellular mechanisms of uptake, including endocytic mechanisms, ligand/receptor interaction and fusion with plasma membranes will not be discussed here (for a review, please see ref. 7).
In GBM, neighboring cells such as healthy neural cells, endothelial cells, microglia, monocytes/macrophages and tumor cells have been documented to uptake tumor-derived EVs (Table 1 and please see our next section).10,21 Indeed, mRNA materials identified in GBM-derived EVs could be transferred to surrounding recipient cells.10,14,15 Moreover, specific mRNA mutations (i.e. IDH1) were detected in EVs isolated from glioma patient sera and cerebrospinal fluid.29 As IDH1 mutations are well described in a subset of GBM patients,30,31 we can hypothesize that these circulating EVs, which contain specific mutated mRNA, may arise directly from GBM cells. Recently, intravital imaging unveiled that GBM-derived EVs are ingested by the surrounding microglia and monocyte/macrophages in vivo.32
Table 1.
GBM-derived EV cargoes and their functional impacts.
| Recipient cells | Cargo | Biological effects | |
|---|---|---|---|
| Intra-Tumoral | Tumor cells | mtDNA | nd18 |
| miR-1 | Tumorgenicity, invasion and growth34 | ||
| miR-21 | Anti-apoptosis, proliferation10,17 | ||
| hypoxia signatureEGFRvIIIclic1proteinsTrkB | Cell migration14Tumor growth12Tumor growth65GBM subtype tumorigenesis25Aggressiveness50 | ||
| Endothelial cells | miR-9mRNAshypoxia signatureangiogenic factorssemaphorin 3A | Migration, angiogenesis66Angiogenesis10Angiogenesis14Angiogenesis10Permeability21 | |
| Microglia Monocyte/macrophage | miR-451, miR-21 | Cytokine profile32 | |
| Extra-Tumoral | Endothelial cells | Pro-coagulation39, 40 | |
| Endothelial cells Monocyte/macrophage | semaphorin 3A | nd21 | |
| TrkB | nd50 | ||
| proteins | M2 phenotype37 | ||
| Lymphocyte | proteins | Immunotolerance67 |
nd: not determined
Interestingly, electronic microscopy analysis unveiled the presence of nanofilaments on exosomes derived from U87 and U251 GBM cell lines, but not on normal astrocytes.28 As such, it is tempting to speculate that this phenotypic observation could have functional consequences. For instance, such nanofilaments may create a diffusive network around cancer cells and participate in the direct fixation of secreted exosomes to their cellular targets.28
Thus, GBM-released EVs might ultimately contribute to tumor progression and heterogeneity, since they retain their ability to be taken up by multiple cellular targets and to disseminate throughout the body.
Functions in the brain tumor microenvironment and beyond
EVs action within brain tumors
In the central nervous system, the main postulated physiological role for EVs corresponds to their ability to rapidly and robustly exchange signals between glia and neurons, thus promoting neuronal survival, microglia-mediated immune responses, and synapse assembly and plasticity.33 This system is most likely corrupted in brain tumors to the tumor's own benefit. Indeed, EVs represent important means of communication within the tumor microenvironment and GBM-derived EVs have been suggested to promote cell proliferation, invasion, immunotolerance, angiogenesis and endothelium defects (Fig. 1).
Figure 1.
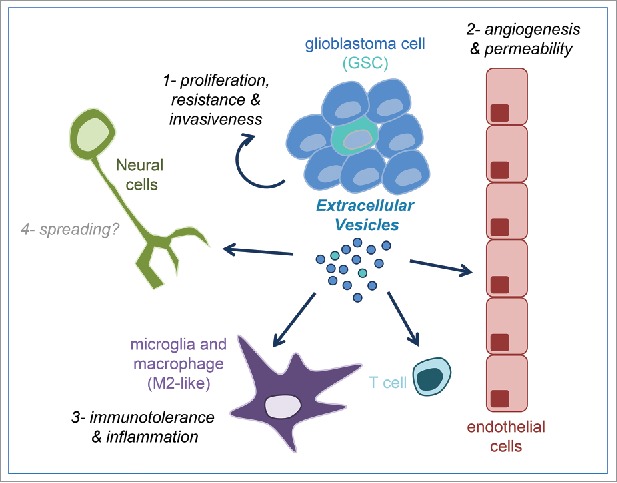
Glioblastoma-derived extracellular vesicles actions in the tumor microenvironment. Glioblastoma and Glioblastoma Stem-like (GSC) cells composing the tumor mass release extracellular vesicles that can be taken up and affect tumors cells themselves or neighboring cells such as endothelial cells and immune system cells to corrupt their functions (labeled as #1–3). They could also potentially affect healthy surrounding neural cells, such as neurons and astrocytes, and therefore spread oncogenic materials (indicated as #4).
It has been shown, for example, that hypoxic conditions can increase the hypoxic-related mRNA and protein content of tumor-released EVs, which in turn impacts on tumor growth both in vitro and in vivo.14 The analysis of these patient-derived exosome-like fractions revealed that the enriched mRNA and proteins were associated with poor prognosis,14 suggesting that cancer cells can adapt to the hostile hypoxic microenvironment. Conversely, EVs also convey messages to hack their environment. MiR-1 overexpression in glioblastoma stem-like cells (GSC) was able to modify EVs protein cargoes and to ultimately reduce tumorgenicity, invasiveness and angiogenesis in vivo in xenografted mice.34 From a molecular standpoint, miR-1 targets the mRNA of Annexin A2, one of the most abundant proteins in GBM-derived EVs, in addition to other pro-oncogenic signals.34 In keeping with this idea, GBM-produced EVs were shown to operate directly on endothelial cells to promote tubulogenesis and permeability, 2 hallmarks of tumor angiogenesis.25,35 Likewise, EVs collected from GBM cell lines can tRNA to normal brain endothelial cells.10 Moreover, our team has demonstrated that GSC-liberated EVs convey the pro-permeability guidance molecule semaphorin 3A toward neighboring brain endothelial cells and ultimately contribute to enhance vascular permeability in orthotopic GBM xenograft models,21 reinforcing the concept that tumor-derived EVs can pervert the vasculature. In line with the hypothesis that EVs play a role in the sabotage of their environment, GBM-produced EVs can be taken up by innate immune cells, microglia and monocytes/macrophages within the brain, and shift their cytokine expression profile and mRNA content toward a pro-tumoral phenotype (Fig. 1).32
Although these recent compelling data highlight the likely involvement of GBM-derived EVs to tumor properties, we have to keep in mind that most of these observations occurred following administration of EVs collected after several steps of purification/concentration and/or from in vitro experiments. Interestingly, intravital imaging was recently deployed to directly image GBM-derived EVs transfer to microglia and monocytes/macrophages within the brain in living animals. As a consequence, recipient cells were denatured, as exemplified by changes in the levels of miR-21 and c-MYC RNA.32
Is there a role for circulating EVs?
EVs emanating from the tumor mass have been detected outside the tumor microenvironment, in particular circulating freely in the plasma.21,26,27 While this feature could be employed as a diagnostic tool (please see our next section), the functional and biological consequences remain unclear in GBM (Fig. 1). In the context of melanoma, endogenous tumor-derived EVs were tracked in vivo by the means of multiple reporter mouse strains, and their further dissemination through lymph nodes was visualized and established to precede tumor cell detection.36 This study established that circulating tumor-derived EVs could also reshape at distance systemic responses, and affect in turn tumor progression.
In GBM, 2 recent studies suggested that circulating EVs can modify the immune system and contribute to immunomodulation in systemic and in the tumors.37 First, GBM-derived EVs impact on the monocytic lineage, which acquires a de novo tumor-supportive phenotype.37 Additionally, EVs derived from serum-purified GBM patients were shown to promote the M2-like phenotype, with the serum cytokine profile typical of Th2 bias, suggesting an action of EVs throughout the organism.38
Paralleling the action on the immune system, GBM-derived EVs can affect the biology of the vasculature. For instance, thromboembolic diseases observed in GBM patients might rely on the pro-coagulant activity detected in circulating microparticles.39,40 Circulating EVs were shown to transport pro-permeability and pro-angiogenic factors,10,21 including GSC-derived semaphorin 3A that could reach the endothelium of virtually all healthy organs.21 Indeed, GBM patient serum-purified EVs are loaded with semaphorin 3A, which further orchestrated the loss of barrier integrity.21 One important challenge will be to track in vivo specifically GBM-derived EVs and explore both their cargoes and their biological functions, in the course of tumor initiation, progression and relapse.
Translational outcomes and clinical potential
Could EVs serve as prognosis/theranostic marker?
Lack of therapeutically convenient monitoring tools remains a major cause of failure in the management of GBM patients.41 For example, MRI only detects already established tumor of several hundreds or thousands of tumor cells.42 According to the recent review by Westphal and Lamszus,43 there are still only very few clinically relevant markers for GBM, among them MGMT methylation status, 1p/19q codeletion in oligodendroglial tumors, the EGFRvIII variant, and IDH1 and BRAF mutations.43 Interestingly, analysis of EVs derived from biofluids emerged as a promising source of biomarkers because: i) they are easy to collect and implied non-invasive procedures as opposed to intracranial tissue biopsies; ii) their content mirrors the genetic and cellular status of mother tumor cells;13 and iii) the half-life of labeled EVs seems quite short in the blood circulation, suggesting that they could be a suitable biomarker reflecting rapid changes in tumor cell state (Fig. 2).44 This raises the possibility that several biomarkers could be combined to recapitulate the tumor heterogeneity at a given time point and in the course of tumor progression, from multiple exploitable clinical samples (blood, urine, cerebrospinal fluid, saliva, lymph, sexual secretion etc).
Figure 2.
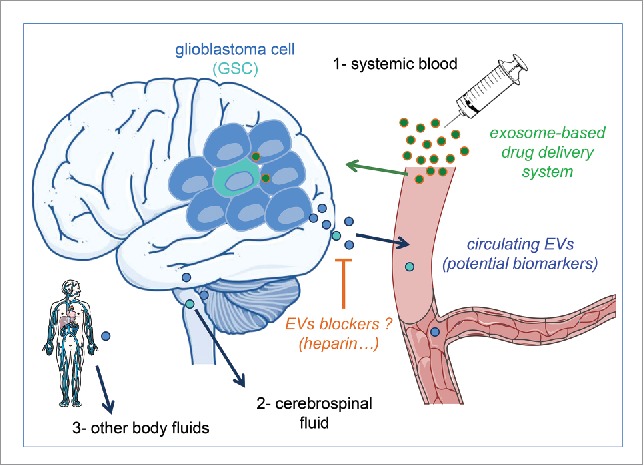
Potential translational applications of extracellular vesicles in glioblastoma. Extracellular Vesicles (EVs) are not yet used in clinic for glioblastoma patients but recent researches highlight them as promising circulating biomarkers, potentiate new tools for drug delivery and new target to block intercellular communication.
The translational interest for GBM diagnosis was firstly highlighted with the pioneer work of Skog et al.10 In this study, they demonstrated that EVs purified from patient sera, as opposed to healthy donors, are enriched in a specific subset of RNA (i.e., EGFRvIII and miR-21) that could be further used to monitor oncogenic mutations and gene expression.10 In keeping with this idea, Akers et al.45 characterized the presence of miR-21 in EVs prepared from cerebrospinal fluid of GBM patients. However, the in-depth determination of the relative abundance of miRNA in plasma- or cerebrospinal fluid-isolated EVs reveals the heterogeneity of such biomarkers.46 Indeed, this study reported that the amount of detectable miRNA was rather low and heterogeneous in plasma exosomes or microvesicles, with an average of less than one miRNA molecule monitored per 200 to 85.000 EVs. In contrast, this average was higher in cerebrospinal fluid and increased to around one molecule of miRNA for 150–25.000 EVs.46 This emphasizes that extraction protocols and detection sensitivity are critical steps for robust clinical applications.47,48 Optimizing preservation of EVs content and improving quantification methods are currently under investigation to determine new informative markers.46,47,49 Furthermore, a number of recent studies established that, in addition to mRNA and miRNA, proteins can also be examined from plasma-isolated EVs.21,50
Besides diagnosis purposes, the fact that EVs can somehow mirror tumor progression (please see our previous section) raises the possibility that EVs are suitable for theranostic application, personalized medicine and evaluation of response to treatment. In this regard, the levels of exosomal mRNA collected from plasma correlate with the immunological responses of patients enrolled in a vaccination trial.51 In addition, as mentioned above, MGMT methylation status in circulating EVs was also found to correspond with the identity of the parental tumor and be predictive of the current response to treatment.52
Altogether, this recent series of studies hold promise that EVs content may provide a novel tool for early diagnosis and companion biomarkers in combination with the current methods.10,48
Could EVs be functionally targeted?
Blocking EV emission, diffusion and transmission may be envisioned as a novel therapeutic strategy. First, an early study demonstrated that the secretory mechanism involved in tumor-based EV release could be halted.53 Indeed, this was achieved by interfering with the Rab27 small GTPase. Although blocking Rab27 in GBM efficiently reduced migration and invasiveness, this involves slightly different mechanisms, such as lysosomal action.54 Thus, it is still unclear whether Rab27 contribution to GBM disease relies on EV secretion, as shown in other cancers.55 On the other hand, exosome uptake can be impaired by heparin.56 Heparin and α-difluoromethylornithine (DFMO) target heparan sulfate proteoglycans, impacting the general mechanisms of vesicle endocytosis, including in GBM cells.57 Cholesterol of lipid rafts on EVs can also be manipulated by methyl-ß-cyclodextrin (MßCD) in the U87 GBM cell line, in order to block lipid raft-mediated endocytosis.58 Applied to the tumor microenvironment, these approaches could hypothetically affect tumor growth and invasion. However, further efforts must elucidate the signaling mechanisms involved prior to clinical application.
Could EVs be used as drug delivery agents?
Because EVs are prone to protect their content in order to cross membrane/biological barriers, to travel throughout the body and reach specific cellular targets, they emerge as attractive strategy to deliver drugs into the tumor mass, with the idea to target defined cellular components within the tumor microenvironment (Fig. 2). Paralleling drug delivery systems such as synthetic vectors, adeno-associated virus and lipidic and carbone-based nanocarriers, EVs could be engineered to transport and release therapeutic compounds into the brain.59 The proof-of-concept has been elegantly established in healthy and Alzheimer-like mouse brains, where systemically injected exosomes were shown to convey siRNA to brain tissues and to specifically and efficiently target RNA expression in neurons, microglia and oligodendrocytes.60 Dendritic cells were manipulated to release exosomes expressing at their surface Lamp2 fused to the neuron peptide RVG, in order to specifically target neural cells. Meanwhile, siRNA were prior introduced in donor cells.60 Likewise, exosomes were manipulated to express EGFR ligands and therefore designed to deliver siRNA to EGFR-expressing breast cancer cells.61 Similarly, intranasal administration of exosomes was proven to efficiently reach brain tissues in mice.62 In this search for novel therapeutic approaches, systemic injection of brain endothelial cell-derived EVs loaded with cytotoxic drugs (doxorubicin and paclitaxel) were demonstrated to cross the blood brain barrier in a fish model for GBM.63 In addition, the expression of GBM-specific multidrug transporter could be reduced by anti-miR strategy introduced into exosome-producing donor cells.64
Thus, cellular exosomes can be manipulated to specifically reach brain tissues, normally protected against xenobiotic invasion by the blood brain barrier (Fig. 2). Their content can be specifically enriched with siRNA, miRNA to be delivered to the tumor and even loaded with cytotoxic drugs via direct electroporation onto exosome preparation.56
Although promising, such data were obtained from in vitro approaches and in vivo mouse models. To our knowledge, there is no translational application yet in GBM. Preclinical studies should define whether risk for patients can be reduced and therapeutics well tolerated when using non-viable biological structures. To this end, it will be essential to identify safety donor cells (i.e. autologous to the patient, cultured in safety and sterile conditions without derivations…) that can be engineered and loaded with specifics cargoes to cross the blood brain barrier and reach the recipient cells.
Conclusions and perspectives
Over the past 5 years, researchers have collected information on glioblastoma-derived extracellular vesicles in terms of their content and their fundamental properties. Although their genesis and the triage process of cellular component specifically addressed into EVs are rather elusive, it is now well established that circulating EVs reflect tumor state and therefore can serve as diagnostic and theranostic tools toward personalized medicine. Additionally, EVs offer useful features to improve drug delivery systems as they can pass through biological barriers, are immunotolerated, can freely circulate, and their content is protected from degradation. Better knowledge is still however required to clarify their biological functions, their mode of action both within and outside the tumor microenvironment, and their privileged cellular targets.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
The authors are thankful to the present and past members of SOAP laboratory.
Funding
Research in SOAP team was funded by: Fondation ARC pour la recherche contre le Cancer, Ligue Nationale contre le Cancer comite Pays-de-la-Loire, comite Maine-et-Loire and comite Sarthe, and Connect Talent grant from Region Pays-de-la-Loire and Nantes Metropole.
References
- [1].Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJB, Belanger K, Brandes AA, Marosi C, Bogdahn U, et al.. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med 2005; 352:987-96; PMID:15758009; http://dx.doi.org/ 10.1056/NEJMoa043330 [DOI] [PubMed] [Google Scholar]
- [2].Bao S, Wu Q, McLendon RE, Hao Y, Shi Q, Hjelmeland AB, Dewhirst MW, Bigner DD, Rich JN. Glioma stem cells promote radioresistance by preferential activation of the DNA damage response. Nature 2006; 444:756-60; PMID:17051156; http://dx.doi.org/ 10.1038/nature05236 [DOI] [PubMed] [Google Scholar]
- [3].Zappulli V, Friis KP, Fitzpatrick Z, Maguire CA, Breakefield XO. Extracellular vesicles and intercellular communication within the nervous system. J Clin Invest 2016; 126:1198-207; PMID:27035811; http://dx.doi.org/ 10.1172/JCI81134 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [4].Trams EG, Lauter CJ, Salem N, Heine U. Exfoliation of membrane ecto-enzymes in the form of micro-vesicles. Biochim Biophys Acta 1981; 645:63-70; PMID:6266476; http://dx.doi.org/ 10.1016/0005-2736(81)90512-5 [DOI] [PubMed] [Google Scholar]
- [5].Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem 1987; 262:9412-20; PMID:3597417 [PubMed] [Google Scholar]
- [6].Gould SJ, Raposo G. As we wait: coping with an imperfect nomenclature for extracellular vesicles. J Extracell Vesicles 2013; 2:20389; PMID:24009890; http://dx.doi.org/ 10.3402/jev.v2i0.20389 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [7].Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol 2013; 200:373-83; PMID:23420871; http://dx.doi.org/ 10.1083/jcb.201211138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Cocucci E, Meldolesi J. Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trends Cell Biol 2015; 25:364-72; PMID:25683921; http://dx.doi.org/ 10.1016/j.tcb.2015.01.004 [DOI] [PubMed] [Google Scholar]
- [9].Tani E, Nakano M, Itagaki T, Fukumori T. Cell membrane structure of human giant-celled glioblastoma. Acta Neuropathol (Berl) 1978; 41:61-5; http://dx.doi.org/ 10.1007/BF00689558 [DOI] [PubMed] [Google Scholar]
- [10].Skog J, Würdinger T, van Rijn S, Meijer DH, Gainche L, Sena-Esteves M, Curry WT, Carter BS, Krichevsky AM, Breakefield XO. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol 2008; 10:1470-6; PMID:19011622; http://dx.doi.org/ 10.1038/ncb1800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [11].Balaj L, Lessard R, Dai L, Cho Y-J, Pomeroy SL, Breakefield XO, Skog J. Tumour microvesicles contain retrotransposon elements and amplified oncogene sequences. Nat Commun 2011; 2:180; PMID:21285958; http://dx.doi.org/ 10.1038/ncomms1180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Al-Nedawi K, Meehan B, Micallef J, Lhotak V, May L, Guha A, Rak J. Intercellular transfer of the oncogenic receptor EGFRvIII by microvesicles derived from tumour cells. Nat Cell Biol 2008; 10:619-24; PMID:18425114; http://dx.doi.org/ 10.1038/ncb1725 [DOI] [PubMed] [Google Scholar]
- [13].Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol 2007; 9:654-9; PMID:17486113; http://dx.doi.org/ 10.1038/ncb1596 [DOI] [PubMed] [Google Scholar]
- [14].Kucharzewska P, Christianson HC, Welch JE, Svensson KJ, Fredlund E, Ringnér M, Mörgelin M, Bourseau-Guilmain E, Bengzon J, Belting M. Exosomes reflect the hypoxic status of glioma cells and mediate hypoxia-dependent activation of vascular cells during tumor development. Proc Natl Acad Sci U S A 2013; 110:7312-7; PMID:23589885; http://dx.doi.org/ 10.1073/pnas.1220998110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [15].Li CCY, Eaton SA, Young PE, Lee M, Shuttleworth R, Humphreys DT, Grau GE, Combes V, Bebawy M, Gong J, et al.. Glioma microvesicles carry selectively packaged coding and non-coding RNAs which alter gene expression in recipient cells. RNA Biol 2013; 10:1333-44; PMID:23807490; http://dx.doi.org/ 10.4161/rna.25281 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [16].Godlewski J, Krichevsky AM, Johnson MD, Chiocca EA, Bronisz A. Belonging to a network–microRNAs, extracellular vesicles, and the glioblastoma microenvironment. Neuro-Oncol 2015; 17:652-62; PMID:25301812; http://dx.doi.org/ 10.1093/neuonc/nou292 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Chan JA, Krichevsky AM, Kosik KS. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res 2005; 65:6029-33; PMID:16024602; http://dx.doi.org/ 10.1158/0008-5472.CAN-05-0137 [DOI] [PubMed] [Google Scholar]
- [18].Guescini M, Genedani S, Stocchi V, Agnati LF. Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. J Neural Transm Vienna Austria 1996 2010; 117:1-4 [DOI] [PubMed] [Google Scholar]
- [19].Redzic JS, Ung TH, Graner MW. Glioblastoma extracellular vesicles: reservoirs of potential biomarkers. Pharmacogenomics Pers Med 2014; 7:65-77 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [20].Mathivanan S, Fahner CJ, Reid GE, Simpson RJ. ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic Acids Res 2012; 40:D1241-1244; PMID:21989406; http://dx.doi.org/ 10.1093/nar/gkr828 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Treps L, Edmond S, Harford-Wright E, Galan-Moya EM, Schmitt A, Azzi S, Citerne A, Bidère N, Ricard D, Gavard J. Extracellular vesicle-transported Semaphorin3A promotes vascular permeability in glioblastoma. Oncogene 2016; 35:2615-23; PMID:26364614; http://dx.doi.org/ 10.1038/onc.2015.317 [DOI] [PubMed] [Google Scholar]
- [22].Graner MW, Alzate O, Dechkovskaia AM, Keene JD, Sampson JH, Mitchell DA, Bigner DD. Proteomic and immunologic analyses of brain tumor exosomes. FASEB J Off Publ Fed Am Soc Exp Biol 2009; 23:1541-57 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Llorente A, Skotland T, Sylvänne T, Kauhanen D, Róg T, Orłowski A, Vattulainen I, Ekroos K, Sandvig K. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim Biophys Acta 2013; 1831:1302-9; PMID:24046871; http://dx.doi.org/ 10.1016/j.bbalip.2013.04.011 [DOI] [PubMed] [Google Scholar]
- [24].Verhaak RGW, Hoadley KA, Purdom E, Wang V, Qi Y, Wilkerson MD, Miller CR, Ding L, Golub T, Mesirov JP, et al.. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell 2010; 17:98-110; PMID:20129251; http://dx.doi.org/ 10.1016/j.ccr.2009.12.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [25].Ricklefs F, Mineo M, Rooj AK, Nakano I, Charest A, Weissleder R, Breakefield XO, Chiocca EA, Godlewski J, Bronisz A. Extracellular Vesicles from High-Grade Glioma Exchange Diverse Pro-oncogenic Signals That Maintain Intratumoral Heterogeneity. Cancer Res 2016; 76:2876-81; PMID:27013191; http://dx.doi.org/ 10.1158/0008-5472.CAN-15-3432 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [26].Noerholm M, Balaj L, Limperg T, Salehi A, Zhu LD, Hochberg FH, Breakefield XO, Carter BS, Skog J. RNA expression patterns in serum microvesicles from patients with glioblastoma multiforme and controls. BMC Cancer 2012; 12:22; PMID:22251860; http://dx.doi.org/ 10.1186/1471-2407-12-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Street JM, Barran PE, Mackay CL, Weidt S, Balmforth C, Walsh TS, Chalmers RTA, Webb DJ, Dear JW. Identification and proteomic profiling of exosomes in human cerebrospinal fluid. J Transl Med 2012; 10:5; PMID:22221959; http://dx.doi.org/ 10.1186/1479-5876-10-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Sharma S, Das K, Woo J, Gimzewski JK. Nanofilaments on glioblastoma exosomes revealed by peak force microscopy. J R Soc Interface R Soc 2014; 11:20131150; http://dx.doi.org/ 10.1098/rsif.2013.1150 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [29].Chen WW, Balaj L, Liau LM, Samuels ML, Kotsopoulos SK, Maguire CA, Loguidice L, Soto H, Garrett M, Zhu LD, et al.. BEAMing and droplet digital PCR analysis of mutant IDH1 mRNA in glioma patient serum and cerebrospinal fluid extracellular vesicles. Mol Ther Nucleic Acids 2013; 2:e109; PMID:23881452; http://dx.doi.org/ 10.1038/mtna.2013.28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Parsons DW, Jones S, Zhang X, Lin JC-H, Leary RJ, Angenendt P, Mankoo P, Carter H, Siu I-M, Gallia GL, et al.. An integrated genomic analysis of human glioblastoma multiforme. Science 2008; 321:1807-12; PMID:18772396; http://dx.doi.org/ 10.1126/science.1164382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Cancer Genome Atlas Research Network Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature 2008; 455:1061-8; PMID:18772890; http://dx.doi.org/ 10.1038/nature07385 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].van der Vos KE, Abels ER, Zhang X, Lai C, Carrizosa E, Oakley D, Prabhakar S, Mardini O, Crommentuijn MHW, Skog J, et al.. Directly visualized glioblastoma-derived extracellular vesicles transfer RNA to microglia/macrophages in the brain. Neuro-Oncol 2016; 18:58-69; PMID:26433199; http://dx.doi.org/ 10.1093/neuonc/nov244 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Budnik V, Ruiz-Cañada C, Wendler F. Extracellular vesicles round off communication in the nervous system. Nat Rev Neurosci 2016; 17:160-72; PMID:26891626; http://dx.doi.org/ 10.1038/nrn.2015.29 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].Bronisz A, Wang Y, Nowicki MO, Peruzzi P, Ansari KI, Ogawa D, Balaj L, Rienzo GD, Mineo M, Nakano I, et al.. Extracellular Vesicles Modulate the Glioblastoma Microenvironment via a Tumor Suppression Signaling Network Directed by miR-1. Cancer Res 2014; 74:738-50; PMID:24310399; http://dx.doi.org/ 10.1158/0008-5472.CAN-13-2650 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [35].Giusti I, Monache SD, Di Francesco M, Sanità P, D'Ascenzo S, Gravina GL, Festuccia C, Dolo V. From glioblastoma to endothelial cells through extracellular vesicles: messages for angiogenesis. Tumor Biol 2016; 37:12743; http://dx.doi.org/ 10.1007/s13277-016-5165-0 [DOI] [PubMed] [Google Scholar]
- [36].Pucci F, Garris C, Lai CP, Newton A, Pfirschke C, Engblom C, Alvarez D, Sprachman M, Evavold C, Magnuson A, et al.. SCS macrophages suppress melanoma by restricting tumor-derived vesicle-B cell interactions. Science 2016; 352:242-6; PMID:26989197; http://dx.doi.org/ 10.1126/science.aaf1328 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].de Vrij J, Maas SLN, Kwappenberg KMC, Schnoor R, Kleijn A, Dekker L, Luider TM, de Witte LD, Litjens M, van Strien ME, et al.. Glioblastoma-derived extracellular vesicles modify the phenotype of monocytic cells. Int J Cancer 2015; 137:1630-42; PMID:25802036; http://dx.doi.org/ 10.1002/ijc.29521 [DOI] [PubMed] [Google Scholar]
- [38].Harshyne LA, Nasca BJ, Kenyon LC, Andrews DW, Hooper DC. Serum exosomes and cytokines promote a T-helper cell type 2 environment in the peripheral blood of glioblastoma patients. Neuro-Oncol 2016; 18:206-15; PMID:26180083; http://dx.doi.org/ 10.1093/neuonc/nov107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [39].Perry JR. Thromboembolic disease in patients with high-grade glioma. Neuro-Oncol 2012; 14 Suppl 4:iv73-80; PMID:23095833; http://dx.doi.org/ 10.1093/neuonc/nos197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Sartori MT, Della Puppa A, Ballin A, Saggiorato G, Bernardi D, Padoan A, Scienza R, d'Avella D, Cella G. Prothrombotic state in glioblastoma multiforme: an evaluation of the procoagulant activity of circulating microparticles. J Neurooncol 2011; 104:225-31; PMID:21104297; http://dx.doi.org/ 10.1007/s11060-010-0462-8 [DOI] [PubMed] [Google Scholar]
- [41].Clarke JL, Chang SM. Neuroimaging: diagnosis and response assessment in glioblastoma. Cancer J Sudbury Mass 2012; 18:26-31; http://dx.doi.org/ 10.1097/PPO.0b013e318244d7c8 [DOI] [PubMed] [Google Scholar]
- [42].Sorensen AG, Batchelor TT, Wen PY, Zhang W-T, Jain RK. Response criteria for glioma. Nat Clin Pract Oncol 2008; 5:634-44; PMID:18711427; http://dx.doi.org/ 10.1038/ncponc1204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [43].Westphal M, Lamszus K. Circulating biomarkers for gliomas. Nat Rev Neurol 2015; 11:556-66; PMID:26369507; http://dx.doi.org/ 10.1038/nrneurol.2015.171 [DOI] [PubMed] [Google Scholar]
- [44].Morishita M, Takahashi Y, Nishikawa M, Sano K, Kato K, Yamashita T, Imai T, Saji H, Takakura Y. Quantitative analysis of tissue distribution of the B16BL6-derived exosomes using a streptavidin-lactadherin fusion protein and iodine-125-labeled biotin derivative after intravenous injection in mice. J Pharm Sci 2015; 104:705-13; PMID:25393546; http://dx.doi.org/ 10.1002/jps.24251 [DOI] [PubMed] [Google Scholar]
- [45].Akers JC, Ramakrishnan V, Kim R, Skog J, Nakano I, Pingle S, Kalinina J, Hua W, Kesari S, Mao Y, et al.. MiR-21 in the extracellular vesicles (EVs) of cerebrospinal fluid (CSF): a platform for glioblastoma biomarker development. PloS One 2013; 8:e78115; PMID:24205116; http://dx.doi.org/ 10.1371/journal.pone.0078115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [46].Akers JC, Ramakrishnan V, Kim R, Phillips S, Kaimal V, Mao Y, Hua W, Yang I, Fu C-C, Nolan J, et al.. miRNA contents of cerebrospinal fluid extracellular vesicles in glioblastoma patients. J Neurooncol 2015; 123:205-16; PMID:25903655; http://dx.doi.org/ 10.1007/s11060-015-1784-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [47].Akers JC, Ramakrishnan V, Nolan JP, Duggan E, Fu C-C, Hochberg FH, Chen CC, Carter BS. Comparative Analysis of Technologies for Quantifying Extracellular Vesicles (EVs) in Clinical Cerebrospinal Fluids (CSF). PloS One 2016; 11:e0149866; PMID:26901428; http://dx.doi.org/ 10.1371/journal.pone.0149866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [48].Shao H, Chung J, Balaj L, Charest A, Bigner DD, Carter BS, Hochberg FH, Breakefield XO, Weissleder R, Lee H. Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat Med 2012; 18:1835-40; PMID:23142818; http://dx.doi.org/ 10.1038/nm.2994 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Akers JC, Ramakrishnan V, Yang I, Hua W, Mao Y, Carter BS, Chen CC. Optimizing preservation of extracellular vesicular miRNAs derived from clinical cerebrospinal fluid. Cancer Biomark 2016; 17(2):125-132; http://dx.doi.org/ 10.3233/CBM-160609 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].Pinet S, Bessette B, Vedrenne N, Lacroix A, Richard L, Jauberteau M-O, Battu S, Lalloué F. TrkB-containing exosomes promote the transfer of glioblastoma aggressiveness to YKL-40-inactivated glioblastoma cells. Oncotarget 2016; 7(31):50349; PMID:27385098; http://dx.doi.org/ 10.18632/oncotarget.10387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [51].Muller L, Muller-Haegele S, Mitsuhashi M, Gooding W, Okada H, Whiteside TL. Exosomes isolated from plasma of glioma patients enrolled in a vaccination trial reflect antitumor immune activity and might predict survival. Oncoimmunology 2015; 4:e1008347; PMID:26155415; http://dx.doi.org/ 10.1080/2162402X.2015.1008347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [52].Shao H, Chung J, Lee K, Balaj L, Min C, Carter BS, Hochberg FH, Breakefield XO, Lee H, Weissleder R. Chip-based analysis of exosomal mRNA mediating drug resistance in glioblastoma. Nat Commun 2015; 6:6999; PMID:25959588; http://dx.doi.org/ 10.1038/ncomms7999 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [53].Ostrowski M, Carmo NB, Krumeich S, Fanget I, Raposo G, Savina A, Moita CF, Schauer K, Hume AN, Freitas RP, et al.. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat Cell Biol 2010; 12:19-30-13; PMID:19966785; http://dx.doi.org/ 10.1038/ncb2000 [DOI] [PubMed] [Google Scholar]
- [54].Liu Y, Zhou Y, Zhu K. Inhibition of glioma cell lysosome exocytosis inhibits glioma invasion. PLOS ONE 2012; 7:e45910; PMID:23029308; http://dx.doi.org/ 10.1371/journal.pone.0045910 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [55].Hendrix A, De Wever O. Rab27 GTPases distribute extracellular nanomaps for invasive growth and metastasis: implications for prognosis and treatment. Int J Mol Sci 2013; 14:9883-92; PMID:23665896; http://dx.doi.org/ 10.3390/ijms14059883 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [56].Atai NA, Balaj L, van Veen H, Breakefield XO, Jarzyna PA, Van Noorden CJF, Skog J, Maguire CA. Heparin blocks transfer of extracellular vesicles between donor and recipient cells. J Neurooncol 2013; 115:343-51; PMID:24002181; http://dx.doi.org/ 10.1007/s11060-013-1235-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- [57].Christianson HC, Svensson KJ, van Kuppevelt TH, Li J-P, Belting M. Cancer cell exosomes depend on cell-surface heparan sulfate proteoglycans for their internalization and functional activity. Proc Natl Acad Sci U S A 2013; 110:17380-5; PMID:24101524; http://dx.doi.org/ 10.1073/pnas.1304266110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [58].Svensson KJ, Christianson HC, Wittrup A, Bourseau-Guilmain E, Lindqvist E, Svensson LM, Mörgelin M, Belting M. Exosome uptake depends on ERK1/2-heat shock protein 27 signaling and lipid Raft-mediated endocytosis negatively regulated by caveolin-1. J Biol Chem 2013; 288:17713-24; PMID:23653359; http://dx.doi.org/ 10.1074/jbc.M112.445403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [59].Aryani A, Denecke B. Exosomes as a Nanodelivery System: a Key to the Future of Neuromedicine? Mol Neurobiol 2014; 53:818-34; PMID:25502465; http://dx.doi.org/ 10.1007/s12035-014-9054-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [60].Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJA. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol 2011; 29:341-5; PMID:21423189; http://dx.doi.org/ 10.1038/nbt.1807 [DOI] [PubMed] [Google Scholar]
- [61].Ohno S, Takanashi M, Sudo K, Ueda S, Ishikawa A, Matsuyama N, Fujita K, Mizutani T, Ohgi T, Ochiya T, et al.. Systemically Injected Exosomes Targeted to EGFR Deliver Antitumor MicroRNA to Breast Cancer Cells. Mol Ther 2013; 21:185-91; PMID:23032975; http://dx.doi.org/ 10.1038/mt.2012.180 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [62].Zhuang X, Xiang X, Grizzle W, Sun D, Zhang S, Axtell RC, Ju S, Mu J, Zhang L, Steinman L, et al.. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol Ther 2011; 19:1769-79; PMID:21915101; http://dx.doi.org/ 10.1038/mt.2011.164 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [63].Yang T, Martin P, Fogarty B, Brown A, Schurman K, Phipps R, Yin VP, Lockman P, Bai S. Exosome delivered anticancer drugs across the blood-brain barrier for brain cancer therapy in Danio rerio. Pharm Res 2015; 32:2003-14; PMID:25609010; http://dx.doi.org/ 10.1007/s11095-014-1593-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- [64].Munoz JL, Bliss SA, Greco SJ, Ramkissoon SH, Ligon KL, Rameshwar P. Delivery of functional anti-miR-9 by mesenchymal stem cell–derived exosomes to glioblastoma multiforme cells conferred chemosensitivity. Mol Ther — Nucleic Acids 2013; 2:e126; http://dx.doi.org/ 10.1038/mtna.2013.60 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [65].Setti M, Osti D, Richichi C, Ortensi B, Del Bene M, Fornasari L, Beznoussenko G, Mironov A, Rappa G, Cuomo A, et al.. Extracellular vesicle-mediated transfer of CLIC1 protein is a novel mechanism for the regulation of glioblastoma growth. Oncotarget 2015; 6:31413-27; PMID:26429879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [66].Zhuang G, Wu X, Jiang Z, Kasman I, Yao J, Guan Y, Oeh J, Modrusan Z, Bais C, Sampath D, et al.. Tumour-secreted miR-9 promotes endothelial cell migration and angiogenesis by activating the JAK-STAT pathway. EMBO J 2012; 31:3513-23; PMID:22773185; http://dx.doi.org/ 10.1038/emboj.2012.183 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Hellwinkel JE, Redzic JS, Harland TA, Gunaydin D, Anchordoquy TJ, Graner MW. Glioma-derived extracellular vesicles selectively suppress immune responses. Neuro-Oncol 2016; 18:497-506; PMID:26385614; http://dx.doi.org/ 10.1093/neuonc/nov170 [DOI] [PMC free article] [PubMed] [Google Scholar]