

Abstract
Commensal non-toxigenic Bacteroides fragilis confers powerful health benefits to the host, and has recently been identified as a promising probiotic candidate. We previously isolated B. fragilis strain ZY-312 and identified it as a novel strain based on 16S rRNA sequencing and morphological analyses. We also determined that ZY-312 displayed desirable probiotic properties, including tolerance to simulated digestive fluid, adherence, and in vitro safety. In this study, we aim to investigate whether ZY-312 meets the safety criteria required for probiotic bacteria through comprehensive and systematic evaluation. Consequently, the fatty acid profile, metabolite production, and biochemical activity of strain ZY-312 were found to closely resemble descriptions of B. fragilis in Bergey’s manual. Taxonomic identification of strain ZY-312 based on whole genome sequencing indicated that ZY-312 and ATCC 25285 showed 99.99% similarity. The 33 putative virulence-associated factors identified in ZY-312 mainly encoded structural proteins and proteins with physiological activity, while the lack of bft indicated that ZY-312 was non-toxigenic. In vivo safety was proven in both normal and immune-deficient mice. The 11 identified antibiotic resistance genes were located on the chromosome rather than on a plasmid, ruling out the risk of plasmid-mediated transfer of antibiotic resistance. In vitro, ZY-312 showed resistance to cefepime, kanamycin, and streptomycin. Finally, and notably, ZY-312 exhibited high genetic stability after 100 passages in vitro. This study supplements the foundation work on the safety evaluation of ZY-312, and contributes to the development of the first probiotic representative from the dominant Bacteroidetes phylum.
Keywords: Bacteroides fragilis, safety evaluation, probiotic, whole genome sequencing, genetic stability
Introduction
Commensal gut microbiota are important for host health. They contribute to maturation of the immune system, building intestinal commensalism, resisting infectious microbes, digesting indigestible carbohydrates, and producing certain nutrients such as vitamins and short-chain fatty acids (Sekirov et al., 2010; Fijan, 2014). An increasing number of reports have confirmed the link between intestinal flora disorder and disease, especially autoimmune and metabolic diseases (Fijan, 2014; Althani et al., 2016). Although it is not clear who takes the leap and how it happens, dominant members of the intestinal microbiota are now being examined as potential probiotic candidates to assist in the treatment of disease, or become alternatives to antibiotics (McKenney and Pamer, 2015; Miquel et al., 2015).
Approved probiotic products based on intestinal microbiota species mainly include lactic acid bacteria, Bifidobacterium, and Escherichia coli (Foligne et al., 2013). However, there are currently no probiotic representatives of the phylum Bacteroidetes, the second-largest component of the intestinal flora. Recently, non-toxigenic Bacteroides fragilis (NTBF) was shown to have powerful health benefits to the host, and was recommended as a probiotic candidate (Troy and Kasper, 2010; Hsiao et al., 2013; Deng et al., 2016). It is therefore likely that B. fragilis will be the first probiotic species from the phylum Bacteroidetes.
We previously isolated a novel B. fragilis strain ZY-312, and have carried out basic safety assessments to test its probiotic properties (Deng et al., 2016). Results of 16S rRNA sequence analysis, tolerance to simulated digestive fluid, safety, and adhesion to HT-29 cells suggested B. fragilis ZY-312 possessed desirable probiotic properties. However, these results are not sufficient to definitively conclude that ZY-312 is safe for use as a probiotic. According to the framework of evaluations for probiotics from Europe (Miquel et al., 2015) and by the Food and Agriculture Organization of the United Nations and the World Health Organization (FAO/WHO, 2002), thorough characterization of a strain, including examination of the fatty acid profile, metabolite production, and biochemical activity, evaluation of the potential risk of transferable antibiotic resistance and genetic stability, and verification of safety in animals, especially immunodeficient animals, is an integral part of safety assessment (Miquel et al., 2015). Taking advantage of whole genome sequencing technology, we carried out a thorough characterization and systematic evaluation of novel B. fragilis strain ZY-312 to determine whether it meets the safety criteria required for probiotics.
Materials and Methods
Bacterial Strains and Culture Conditions
Strain ZY-312 was isolated from the feces of a healthy infant, and was previously identified using 16S rRNA gene sequence analysis (Deng et al., 2016). B. fragilis strain ATCC 25285 (also known as NCTC 9343) was purchased from the American Type Culture Collection (ATCC). The culture conditions and materials were as described previously for strains ZY-312 and ATCC 25285 (Deng et al., 2016). For the microbial contamination experiment, E. coli CBSLAM00087, Staphylococcus aureus (S. aureus) CMCC(B) 26058, Salmonella enterica (S. enterica) serotype Paratyphi B CBSLAM00994, Pseudomonas aeruginosa (P. aeruginosa) CBSLAM00818, Candida albicans (C. albicans) CMCC(F)98001, and Clostridium sporogenes (C. sporogenes) CMCC(B) 64941 were obtained from the Academy of Military Medical Science, Beijing, China. All strains were verified by PCR amplification of 16S rRNA gene using the universal primers 27 F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492 R (5′-GGTTACCTTGTTACGACTT-3′) (Lanes D–J). Amplified products were sequenced (Biomed).
Animals
For acute oral toxicity studies, 6- to 8-week-old female specific pathogen-free (SPF) BALB/c mice and nude mice were procured from the Animal Experiment Center of the Academy of Military Medical Sciences, Beijing, China. The animals had free access to tap water and standard rodent diet. All of the animal experiments were performed in accordance with the approval of the Animal Ethics Committee of the Beijing Institute of Microbiology and Epidemiology, Beijing, China.
Genome Sequencing, Assembly, and Analysis
Total DNA was extracted from all strains using a DNA extraction kit (Qiagen, USA) and the complete genome sequence of strain ZY-312 was determined using an Illumina Hiseq 2000 sequencing system (Xu et al., 2015). A genome sequencing library with an average insert size of 400 bp was generated, and raw short-read sequences were filtered using Seqprep1 and Sickle2 software. The genome was then assembled de novo using SOAP de novo software (Luo et al., 2012). Accuracy of the assembled genome sequence was evaluated by mapping all raw reads onto the scaffolds using SOAPaligner (Cui et al., 2013).
Gene prediction was carried out using Glimmer3.0 software (Xu et al., 2015). Putative antibiotic resistance genes and putative virulence factors were identified by BLAST analysis of the antibiotic resistance genes database (ARDB)3 (Liu and Pop, 2009) and the virulence factors database (VFDB)4 (Chen et al., 2005), respectively. A BLAST comparison was performed between the genome sequences of strain ZY-312 and B. fragilis strain NCTC 9343 (GenBank accession number NC_003228). A Perl script was written and the average nucleotide identity (ANI) was calculated using BLAST. A neighbor-joining phylogenetic tree was built using treebest based on the ZY-312 complete genome sequence, the complete shotgun sequences of other B. fragilis strains, and the complete genome sequences of B. fragilis strains NCTC 9343 (GenBank accession number NC_003228), YCH46 (GenBank accession number NC_006347), and B. fragilis 638R (GenBank accession number NC_016776).
General Characteristics of ZY-312
ZY-312 was cultured on Eosin methylene blue agar for 24 h at 37°C (E. coli as positive control), SS agar for 24 h at 37°C (S. enterica serovar Paratyphi B as positive control), Columbia agar containing blood and gentamicin for 48 h at 37°C (C. sporogenes as positive control), mannitol sodium chloride agar for 24 h at 37°C (S. aureus as positive control), NAC agar for 24 h at 37°C (P. aeruginosa as positive control), Sabouraud dextrose agar for 72 h at 28°C (C. albicans as positive control), rose bengal agar for 96 h at 28°C (C. albicans as positive control), and agar agar for 48 h at 37°C (S. aureus as positive control) for excluding microbial contamination. The major fatty acids were analyzed using an HP6890 gas chromatograph (ver. A 5.01), as previously described (Tan et al., 2010). Supernatant from late-logarithmic phase ZY-312 and ATCC 25285 cultures was collected by centrifugation for 10 min at 3000 ×g, and then analyzed by gas chromatography-mass spectrometry. Carbon-source utilization analyses were carried out using a Biolog AN microPlate test panel (Biolog, USA). Bacterial cells were collected from logarithmic phase ZY-312 and ATCC 25285 cultures and resuspended at a concentration of 1.5 × 108 colony forming units (cfu)/mL, and then inoculated into the test plate. After incubating anaerobically for 16–24 h, test results were read using a microplate reader (SpectraMax, M2), using deionized water as a negative control. To test catalase activity, 3–5 drops (about 100 μL) of 3% hydrogen peroxide (freshly prepared) were dropped onto the center of precipitated bacterial cells on microscope slides. To test gelatin liquefaction activity, bacteria were inoculated into gelatin medium, tryptone soy broth (TSB, OXIOD, UK) solidified with 15% gelatin, using a sterilized needle. Plates were incubated at 37°C for 48 h anaerobically, then placed at 4°C for 3–4 h. To determine hemolytic ability, bacteria were anaerobically cultured on tryptone soy agar (TSA, OXIOD, UK) supplemented with 5% (v/v) goat blood for 48 h at 37°C. To determine motility, bacteria were cultured on semi-solid medium (TSB solidified with 0.5% agar) at 37°C for 48 h anaerobically.
Antibiotic Resistance Testing
Antibiotic resistance testing was performed using the minimum inhibitory concentration (MIC) method as described previously (Fernandez et al., 2005). Test antibiotics were chosen based on the antibiotic resistance genes identified in the annotated genome of ZY-312, with the following antibiotics included in the analysis: cefoxitin, ceftriaxone, cefepime, trimethoprim, clarithromycin, chloromycetin, levofloxacin, streptomycin, kanamycin, tetracycline, vancomycin, and polymyxin B (NIFDC, China). Although bcrA was identified in the genome, bacitracin was not used because of high toxicity, while fosmidomycin (rosA) is still undergoing testing and was therefore also excluded. Quinupristin/dalfopristin, the targets of the vatB gene product are not used in China, and there is no corresponding antibiotic for ykkC. ZY-312 was cultivated anaerobically at 37°C for 48 h at a concentration of 107 cfu/mL with antibiotics at different concentrations. MIC was determined by measuring optical density at 600 nm (OD600) with a microplate reader (SpectraMax, M2).
Acute Toxicity to Normal Mice and Nude Mice
To assess the acute toxicity of ZY-312, SPF BALB/c mice were randomly assigned into five groups (n = 5–8). Each group was administered with ZY-312 in a suspension containing 1 × 109, 5 × 1010, or 5 × 1011 cfu/day, 0.5 mL/day culture supernatant, or 0.5 mL/day saline via oral gavage for 5 days. General condition and body weight were observed daily for 17–18 days. At the end of the experimental period, blood samples were obtained for hematological and serum biochemistry analyses. Stomach, colon, liver, and spleen were collected, weighed, and prepared for histopathological examination. Nude mice were orally administered with ZY-312 suspension at a dosage of 1 × 109 cfu/day for 3 days. General condition and bodyweight were observed for 7 days.
Genetic Stability
To explore whether genetic variation occurs in ZY-312 during in vitro passage, we continuously subcultured ZY-312 for 100 generations. Two parallel tubes were inoculated from an original culture of ZY-312 in TSB supplemented with 5% fetal bovine serum, and then incubated at 37°C in an anaerobic glove box (Bugbox, Ruskin) for 24 h. These initial cultures were designated A0 and B0. Subsequent passages were performed by inoculating 1% of each culture into fresh medium every 24 h, until A100 and B100 were obtained. Genetic variation was evaluated by complete genome sequencing of strains A10, A25, A50, and A100, and B10, B25, B50, and B100, and ANI was calculated for each generation. Morphological variations were observed by colony examination, Gram staining, and scanning electron microscopy (SEM) of A100 and B100, with wild-type ZY-312 used for comparison. Growth of ZY-312, A100, and B100 under anaerobic conditions was also examined over a 24-h period. Acute toxicity of A100 and B100 was detected in SPF mice at a dosage of 1 × 109 cfu/day for 3 days, with general condition and bodyweight observed for 7 days.
Experimental Replicates and Statistical Methods
All experiments were performed at least in triplicate using independent assays, and values were expressed as the mean ± standard error. An unpaired Student’s t-test was performed to determine statistically significant differences in the acute toxicity assays. A p-value of <0.05 was considered statistically significant.
Results
Morphological Characteristics
Strain ZY-312 only grew under anaerobic conditions (Supplementary Figure S1), with no growth observed under aerobic conditions or in 5% CO2, implying it was an obligate anaerobe. Microbial contamination was excluded by culturing ZY-312 in different culture media under different conditions (Supplementary Figure S2). ZY-312 formed circular, low convex, semi-opaque colonies following anaerobic cultivation on blood agar plates (Supplementary Figure S1). Cells were Gram-negative, and shown to be rod shaped with rounded ends by scanning electrochemical microscopy (Supplementary Figure S1). This morphology matched the descriptions of B. fragilis in Bergey’s manual (Krieg et al., 2001).
General Characteristics
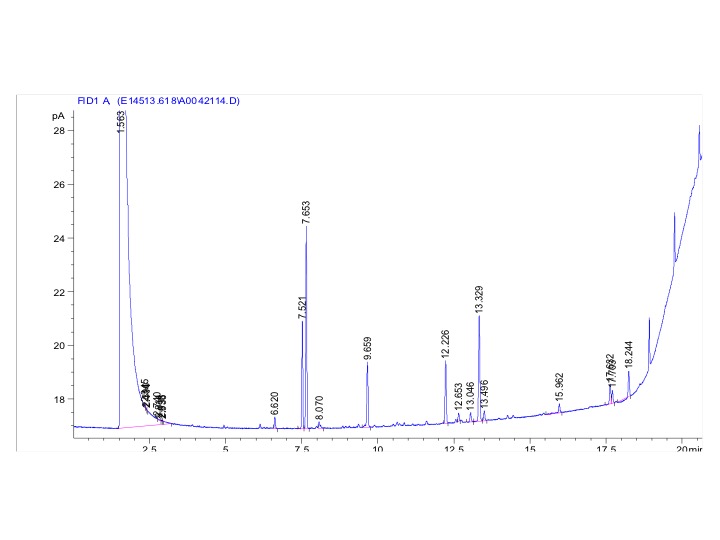
The major fatty acids of ZY-312 were C15:0 anteiso, C15:0 iso, C16:0, and C16:0 3-OH (Supplementary Figure S3 and Table 1A). The major products in the culture supernatant of ZY-312 were lactic acid, acetic acid, succinic acid, propionic acid, and phenylacetic acid (Table 1B). ZY-312 and ATCC 25285 differed in their ability to metabolize L-valine, salicin, L-alanine, L-alanyl-L-glutamine, and N-acetyl-D-glucosamine (Table 2A). Both strains were positive for catalase activity, weakly positive for gelatin liquefaction, and negative for hemolytic activity and motility (Table 2B). The physiological and biochemical characteristics of ZY-312 were in accordance with B. fragilis as described in Bergey’s manual (Krieg et al., 2001).
Table 1.
Major fatty acids (A) and major metabolites (B) of Bacteroides fragilis ZY-312.
| Peak name | Percentage (%) | |||
|---|---|---|---|---|
| (A) | ||||
| C15:0 anteiso | 32.21 | |||
| C15:0 iso | 17.29 | |||
| C16:0 | 11.66 | |||
| C16:0 3-OH | 11.08 | |||
| C14:0 | 1.82 | |||
| C17:0 ante 3-OH | 1.72 | |||
| C18:0 | 1.68 | |||
| C15:0 | 1.63 | |||
| C18:1 CIS 9 | 1.37 | |||
| Unable to distinguish | 19.55 | |||
|
ZY-312 |
ATCC 25285 |
|||
| Time (s) | Normalized area | Time (s) | Normalized area | |
| (B) | ||||
| Acetic acid | 1514 | 67.24 × 105 | 1513 | 27.65 × 105 |
| Butanedioic acid | 593 | 498.29 × 105 | 592 | 107.27 × 105 |
| propionic acid | 1353 | 2.65 × 105 | 1352 | 5.41 × 105 |
| Phenylacetic acid | 2404 | 1.33 × 105 | 2403 | 2.06 × 105 |
| lactic acid | 424 | 556.78 × 105 | 424 | 557.94 × 105 |
Table 2.
Physiological and biochemical properties of B. fragilis ZY-312.
| ZY-312 | ATCC | ZY-312 | ATCC | ||
|---|---|---|---|---|---|
| (A) | |||||
| Water | − | − | Dulcitol | − | − |
| N-Acetyl-D-galactosamine | + | + | D, L-α-Glycerol phosphate | − | − |
| N-Acetyl-D-glucosamine | / | + | N-Acetyl-β-D-mannosamine | − | − |
| D-Fructose | + | + | L-Fucose | − | − |
| Adonitol | − | − | D-Galactose | + | + |
| Amygdalin | + | + | D-Galacturonic acid | − | − |
| D-Arabitol | − | − | Gentiobiose | + | + |
| Arbutin | + | + | D-Gluconic acid | − | − |
| D-Cellobiose | + | + | D-Glucosaminic acid | − | − |
| α-Cyclodextrin | + | + | α-D-Glucose | + | + |
| β-Cyclodextrin | + | + | Glucose-1-phosphate | + | + |
| Dextrin | + | + | Glucose-6-phosphate | + | + |
| Glycerol | − | − | α-Methyl-D-galactoside | − | − |
| i-Erythritol | − | − | β-Methyl-D-galactoside | + | + |
| m-Inositol | − | − | α-Methyl-D-glucoside | − | − |
| α-D-Lactose | + | + | β-Methyl-D-glucoside | − | − |
| Lactulose | + | + | Palatinose | + | + |
| Maltose | + | + | D-Raffinose | + | + |
| Maltotriose | + | + | L-Rhamnose | − | − |
| D-Mannitol | − | − | Salicin | + | − |
| D-Mannose | + | + | D-Sorbitol | − | − |
| D-Melezitose | − | − | Stachyose | + | + |
| D-Melibiose | + | + | Sucrose | + | + |
| 3-Methyl-D-glucose | + | + | D-Trehalose | − | − |
| Pyruvic acid methyl ester | + | + | D-Lactic acid methyl ester | − | − |
| Acetic acid | − | − | D-Malic acid | − | − |
| Formic acid | − | − | L-Malic acid | − | − |
| Fumaric acid | − | − | Propionic acid | − | − |
| Glyoxylic acid | − | − | Pyruvic acid | + | + |
| α-Hydroxybutyric acid | / | / | Turanose | + | + |
| β-Hydroxybutyric acid | − | − | D-Saccharic acid | − | − |
| Itaconic acid | − | − | Succinamic acid | − | − |
| α-Ketobutyric acid | + | + | Succinic acid | − | − |
| α-Ketovaleric acid | + | + | L-Asparagine | − | − |
| D,L-Lactic Acid | − | − | m-Tartaric acid | − | − |
| L-Lactic acid | − | − | Urocanic acid | − | − |
| L-Alaninamide | − | − | L-Methionine | − | − |
| L-Alanine | / | + | L-Phenylalanine | − | − |
| L-Alanyl-L-glutamine | + | / | L-Serine | − | − |
| L-Alanyl-L-histidine | / | / | L-Threonine | − | − |
| L-Alanyl-L-threonine | + | + | L-Valine | / | + |
| Succinic acid monomethyl ester | − | − | L-Valine plus L-aspartic acid | − | − |
| L-Glutamic acid | − | − | 2′-Deoxy adenosine | + | + |
| L-Glutamine | − | − | Inosine | + | + |
| Glycyl-L-aspartic acid | + | + | Thymidine | + | + |
| Glycyl-L-glutamine | − | − | Uridine | + | + |
| Glycyl-L-methionine | − | − | Glycyl-L-proline | − | − |
| Thymidine-5′-monophosphate | − | − | Uridine-5′-monophosphate | − | / |
| ZY-312 | ATCC 25285 | ||||
| (B) | |||||
| Catalase assay | Positive | Positive | |||
| Gelatin liquefaction test | Weakly positive | Weakly positive | |||
| Hemolysis test | Negative | Negative | |||
| Dynamic test | Negative | Negative | |||
Genetic Characteristics
The phylogenetic trees generated from the whole genome sequence of ZY-312 and other B. fragilis strains are shown in Supplementary Figures S5, S6. ZY-312 and ATCC 25285 had an ANI of 99.99%. In total, 33 putative virulence factors (Table 3) and 11 antibiotic resistance genes (Table 4A) were annotated in the ZY-312 genome based on a minimum of 40% amino acid homology. The complete genome was 4,558,494 bp in length and contained on a single chromosome, with an average GC content of 43.08%, consistent with that of B. fragilis (41–44%) (Krieg et al., 2001). All drug-resistance genes were located on the chromosome rather than on plasmids.
Table 3.
Putative virulence-associated genes identified in the genome of B. fragilis ZY-312.
| Gene ID | VFDB_ID | Name | Function |
|---|---|---|---|
| YDGL001582 | VF0091 | Alginate algI | Alginate O-acetyltransferase AlgI |
| YDGL004163 | VF0003 | Capsule cap8E | Capsular polysaccharide synthesis enzyme Cap8E |
| YDGL004164 | VF0003 | Capsule cap8G | Capsular polysaccharide synthesis enzyme Cap8G |
| YDGL002547 | VF0003 | Capsule cap8M | Capsular polysaccharide synthesis enzyme Cap8M |
| YDGL002015 | VF0003 | Capsule cap8O | Capsular polysaccharide synthesis enzyme Cap8O |
| YDGL002539 | VF0003 | Capsule cap8O | Capsular polysaccharide synthesis enzyme Cap8O |
| YDGL002014 | VF0144 | Capsule cps4I | UDP-N-acetylglucosamine-2-epimerase |
| YDGL002540 | VF0144 | Capsule cps4I | UDP-N-acetylglucosamine-2-epimerase |
| YDGL001235 | VF0274 | Capsule cpsJ | Glycosyl transferase CpsJ(V) |
| YDGL004140 | VF0323 | Capsule fcl | Putative fucose syntheses |
| YDGL002177 | VF0323 | Capsule glf | UDP-galactopyranose mutase |
| YDGL001165 | VF0072 | ClpC clpC | Endopeptidase Clp |
| YDGL002866 | VF0072 | ClpC clpC | Endopeptidase Clp |
| YDGL002842 | VF0074 | ClpP clpP | ATP-dependent Clp protease |
| YDGL001133 | VF0215 | Dispersin aatC | AatC ATB binding protein of ABC transporter |
| YDGL003303 | VF0215 | Dispersin aatC | AatC ATB binding protein of ABC transporter |
| YDGL003305 | VF0215 | Dispersin aatC | AatC ATB binding protein of ABC transporter |
| YDGL003564 | VF0215 | Dispersin aatC | AatC ATB binding protein of ABC transporter |
| YDGL003567 | VF0215 | Dispersin aatC | AatC ATB binding protein of ABC transporter |
| YDGL004218 | VF0215 | Dispersin aatC | AatC ATB binding protein of ABC transporter |
| YDGL002299 | VF0273 | Flagella fleQ | Transcriptional regulator FleQ |
| YDGL002756 | VF0334 | HSI-I PA0073 | ATP-binding component of ABC transporter |
| YDGL002122 | VF0159 | Hsp60 htpB | Hsp60, 60K heat shock protein HtpB |
| YDGL004141 | VF0367 | LPS gmd | GDP-mannose 4,6-dehydratase |
| YDGL001251 | VF0106 | MgtBC mgtB | Mg2+transport protein |
| YDGL002018 | VF0153 | Mip mip | Macrophage infectivity potentiator (Mip) |
| YDGL003334 | VF0298 | MprAB mprA | MprB, sensor kinas. MprA, transcriptional factor |
| YDGL003892 | VF0392 | O-Antigen ddhA | Glucose-1-phosphate cytidylyltransferase |
| YDGL003891 | VF0392 | O-Antigen ddhB | CDP-glucose 4,6-dehydratase |
| YDGL003902 | VF0392 | O-Antigen galE | UDP-glucose 4-epimerase |
| YDGL003150 | VF0319 | PanC/PanD panD | panD |
| YDGL003842 | VF0169 | SodB sodB | Superoxide dismutase |
| YDGL003057 | VF0101 | Vi antigen tviB | Vi polysaccharide biosynthesis protein, UDP-glucose/GDP-mannose dehydrogenase |
Table 4.
Putative antibiotic resistance genes (A) identified in the genome of Bacteroides fragilis ZY-312, and minimum inhibitory concentration values (B) for each of the corresponding antibiotics.
| Gene ID | Name | Drug |
|---|---|---|
| (A) | ||
| YDGL002306 | BcrA | Bacitracin |
| YDGL000449 | BL2e_cepa | Cephalosporin |
| YDGL000258 | dfrA22 | Trimethoprim |
| YDGL003477 | MefA | Macrolide |
| YDGL001150 | MexF | Chloramphenicol, Fluoroquinolone |
| YDGL000604 | MexY | Aminoglycoside, Glycylcycline |
| YDGL003440 | RosA | Fosmidomycin |
| YDGL003171 | tet37 | Tetracycline |
| YDGL003704 | VanRA | Vancomycin, Teicoplanin |
| YDGL003646 | VatB | Streptogramin_A |
| YDGL000090 | YkkC | na_antimicrobials |
| Antibiotic | ZY-312 | ATCC 25285 |
| (B) | ||
| Cefoxitin | MIC = 2.0 | MIC = 1.5 |
| Ceftriaxone | MIC < 64 | MIC < 64 |
| Cefepime | MIC > 32 | MIC > 32 |
| Trimethoprim | MIC < 16 | MIC < 16 |
| Clarithromycin | MIC < 8 | MIC < 8 |
| Chloromycetin | MIC < 32 | MIC < 32 |
| Levofloxacin | MIC < 8 | MIC < 8 |
| Streptomycin | MIC > 1200 | MIC > 1200 |
| Kanamycin | MIC > 50 | MIC > 50 |
| Tetracycline | MIC < 16 | MIC < 16 |
| Vancomycin | 4 < MIC < 8 | 4 < MIC < 8 |
| Polymyxin B | MIC > 8 | MIC > 8 |
Bacitracin was eliminated because of high toxicity. Fosmidomycin is still being researched and was therefore excluded. Quinupristin/dalfopristin are not available in China and were also excluded. No corresponding antibiotic exists for the ykkC gene product.
Putative Virulence Factors
Through BLAST analysis of the VFDB, a total of 33 virulence factor homologs were identified in the genome of ZY-312 (Table 3). Most of these putative virulence genes encoded proteins involved in cellular structure or physiological activities, and none had previously been reported as being related to the pathogenesis of B. fragilis. Notably, bft was not present in the genome of ZY-312, indicating that it is a non-toxigenic strain.
Antibiotic Resistance
Antibiotic resistance tests were performed based on the antibiotic resistance genes identified in the genome of YZ-312 (Table 4A), and results are summarized in Table 4B. Based on guidelines for antibiotic resistance breakpoints (Russell and Sambrook, 2002; National Center for Clinical Laboratories, 2006), ZY-312 showed resistance to cefepime, kanamycin, and streptomycin, but was susceptible to ceftriaxone, trimethoprim, clarithromycin, chloramphenicol, tetracycline, and levofloxacin. Without available guidelines for B. fragilis, resistance of ZY-312 to vancomycin and polymyxin B could not be confirmed; however, we observed that 4–8 μg/mL vancomycin and >8 μg/mL polymyxin B were sufficient to inhibit ZY-312 growth in vitro.
ZY-312 Is Non-pathogenic in Both Normal and Immune-Deficient Mice
To confirm the in vivo safety of ZY-312, acute toxicity experiments were performed in both normal SPF BALB/c mice and nude mice. Lacking a thymus and an immune response, nude mice are an ideal animal model for evaluating probiotic safety. No death was observed in the BALB/c mice during the toxicity experiments, no treatment-related toxicity was observed, and no significant difference in body weight was noted between low, medium (Supplementary Figure S4), and high (Figure 1A-1) dose treatment groups and the control group, respectively. Similarly, no difference was found between the culture supernatant-treated group and the control group (Figure 1A-2). In addition, there was no obvious histopathological damage in the stomach, colon, liver, or spleen of BALB/c mice from the high dosage group (Figure 1B). There were also no significant differences in blood routine index or hepatorenal function between the high dose and control groups (data not shown). For the immune-deficient mouse toxicity experiments, ZY-312 again had no deleterious effects on body weight (Figure 1A-3).
FIGURE 1.

ZY-312 is non-pathogenic in mice. (A) Changes in body weight (%) per day were observed for the experimental and control groups. (A-1) Specific pathogen-free (SPF) BALB/c normal mice (n = 8) were treated with 5 × 1011 colony forming units (cfu)/day ZY-312 for 5 days and observed for 18 days. A control group was treated with saline. (A-2) SPF normal mice (n = 16) were treated with culture supernatant (0.5 mL/day) for 5 days and observed for 17 days. Tryptic soy broth was used for the control group. (A-3) Nude mice (n = 5) were treated with ZY-312 at a concentration of 1 × 109 cfu/day for 3 days and observed for 7 days. The control group were treated with saline. (B) Light micrograph images of the stomach, colon, liver, and spleen from mice belonging to the high dosage group (upper) and control group (lower). No significant lesions were observed (mean ± SE; NS, not significant, t-test).
ZY-312 Is Genetically and Phenotypically Stable
To explore whether ZY-312 undergoes major genomic rearrangements during passage, the stability of ZY-312 was examined after 100 generations in vitro. A100 and B100 were morphologically identical to the original ZY-312 strain (Figure 2A) based on visual inspection, optical microscopy, and SEM. The growth characteristics of A100 and B100 (Figure 2B) were not significantly different from those of ZY-312. Furthermore, following oral administration of A100 or B100 at a dose of 1 × 109 cfu/day for 3 days, mice did not show any clinical symptoms or weight loss during the observation period (Figure 2C). The calculated ANI values from each generation were at least 99.98% (Figure 2D).
FIGURE 2.

ZY-312 is genetically stable. (A) Gram-stained ZY-312, A100, and B100 cells following anaerobic culture on tryptic soy agar (5% sheep blood) for 48 h at 37°C. Observation were made using a light microscope (4000×) and a scanning electron microscope (30000×). (B) Growth curves of ZY-312, A100, and B100 cultured anaerobically for 24 h. (C) SPF BALB/c normal mice (n = 8) were treated with A100 or B100 at a concentration of 1 × 109 cfu/day for 3 days and observed for 7 days. A control group was treated with saline. (D) Average nucleotide identities were calculated between each generation.
Discussion
According the recent reports, non-toxic B. fragilis now is regarded as a commensal constituent with potential as a probiotic candidate (Hsiao et al., 2013), because of powerful immunoregulatory benefits and health-promoting effects it owns. Although B. fragilis makes up a very small part of the human intestinal flora, it plays a unique role in the maturation of the host immune system (Mazmanian et al., 2005; Troy and Kasper, 2010). Mono-colonization of mice with B. fragilis was sufficient to correct systemic immune defects in germ-free mice by stimulating maturation of splenic CD4+ T cells (Mazmanian et al., 2005) and rebalancing the Th1/Th2 response. Furthermore, B. fragilis can direct an anti-inflammatory response, conferring protection in experimental mouse models of colitis (Mazmanian et al., 2008; Round and Mazmanian, 2010) and of autoimmune encephalomyelitis (Ochoa-Reparaz et al., 2010). Those observed effects were absent in mice treated with a B. fragilis PSA-deficient mutant strain. PSA is a dominant member of capsular polysaccharide complex (CPC) at the surface of B. fragilis, and contains a zwitterionic motif. B. fragilis is associated with clinical anaerobic infection and the CPC, especially PSA plays a key role in arising those diseases (Lindberg et al., 1982; Mazmanian and Kasper, 2006). Nevertheless, the role of PSA plays in intra-abdominal abscess probably is beneficial rather than harmful, since the inflammation aroused by PSA helps limit the spread of other gut bacteria and prevent more serious infection (Mazmanian and Kasper, 2006; Deng et al., 2016). Instead of being recognized as virulence factor, recently, PSA has been reconsidered as a symbiosis factor with health-promoting effect (Mazmanian et al., 2008).
Therefore, under overall considerations about the pros and cons, we believe non-toxic B. fragilis is a good choice for probiotic candidate and we decided to isolate a new strain of B. fragilis named ZY-312 (Deng et al., 2016) and explore its safety whether satisfy the criteria required for probiotic bacteria. Although the health benefits of B. fragilis are generally recognized, much work still needs to be done to obtain certification of this species as a probiotic. Probiotics are defined as non-pathogenic live microorganisms that confer health benefits to the host when administered in adequate amounts (FAO/WHO, 2002). As living microorganisms with the potential for infection or in situ toxin production, probiotics should fulfill health and safety claims before entering the market. Safety assessment is the chief task for certifying a probiotic, as any adverse effects should be predicted in advance. Accordingly (FAO/WHO, 2002; Miquel et al., 2015), correct identification, sufficient characterization, and evaluation of potential risk and probiotic properties are integral for evaluation of a new probiotic.
Previously, we demonstrated that novel strain ZY-312, isolated from the feces of a healthy breast-fed infant, possessed similar morphological and growth characteristics to typical B. fragilis strains, as well as exhibiting desirable probiotic properties, including tolerance to air, simulated gastric fluid (pH 3.0), simulated intestinal fluid and ox bile (pH 6.8), adhesion, and in vitro safety in colon cells (Deng et al., 2016). Based on these results, we carried out a more thorough characterization and systemic evaluation of ZY-312 in the current study. As recommended (FAO/WHO, 2002), phenotypic testing, fatty acid analysis, metabolite production, biochemical activity, and in vivo toxicity testing, in combination with genetic analysis, taxonomic identification, and putative virulence and antibiotic resistance gene identification analysis, were performed to determine whether ZY-312 is safe for use as a probiotic. The major fatty acids and metabolic products of ZY-312 closely resemble descriptions of B. fragilis in Bergey’s manual, as do the results of biochemical activity testing. Our previous work showed that the 16S rRNA sequence of ZY-312 was 99% identical to that of B. fragilis strain ATCC 28285. However, 16S rRNA sequence analysis has potential drawbacks for separating closely related species because of low taxonomic resolution. Whole genome sequencing identifies taxa with higher taxonomic resolution, and provides more information about gene function, such as putative virulence factors and antibiotic resistance genes (Rijkers et al., 2011; Miquel et al., 2015). Taking advantage of whole genome sequencing technology in the current study, we showed that ZY-312 and ATCC 25285 shared 99.99% ANI, and were derived from the same origin (Supplementary Figure S5), consistent with previous results. Furthermore, a total of 33 putative virulence factors and 11 antibiotic resistance genes were annotated in the genome of ZY-312 through comparative analysis with the VFDB and the ARDB, respectively. The putative virulence factors consisted of structural proteins and proteins with physiological activity, and none had previously been reported in association with the pathogenesis of B. fragilis. Because of the absence of bft, ZY-312 was identified as a NTBF strain.
There are many reports claiming probiotic safety based on an assessed lack of infectivity in normal animals (Bernardeau et al., 2002; Shokryazdan et al., 2016). However, confidence would be increased if assessment could be performed in immunodeficient animals (FAO/WHO, 2002). Therefore, we examined the safety of ZY-312 in both normal and immunodeficient mice. The SPF normal mice treated with high doses or supernatant of ZY-312 did not display any significant strain-related toxigenic symptoms, based on the assessment of body weight changes, histopathological examination, blood routine index, and hepatorenal function. Notably, ZY-312 also proved safe in nude mice (Figure 1A-3).
The FAO/WHO recommends that probiotic strains should be fully characterized, including determination of antibiotic resistance patterns, and should have no risk of transferring antibiotic resistance (FAO/WHO, 2002). Theoretically, consumption of probiotics with transferrable antibiotic resistance genes might lead to refractory infections if multiple antibiotic resistance genes are transferred to a pathogen (Borchers et al., 2009; Sanders et al., 2010). We demonstrated that there is no risk of ZY-312 spreading antibiotic resistance because all identified drug-resistance genes (Table 4A) were located in the chromosome rather than on a plasmid. Moreover, we verified the antibiotic resistance phenotype of ZY-312, and discovered that it is resistant to cefepime, kanamycin, and streptomycin, but susceptible to ceftriaxone, trimethoprim, clarithromycin, chloramphenicol, tetracycline, and levofloxacin. The MICs of vancomycin and polymyxin B for ZY-312 were also identified (Table 4B). The inconsistencies between genotype and observed phenotype might stem from the fact that putative antibiotic resistance genes were annotated based on 40% amino acid homology, meaning that some genes were incorrectly identified in the genome of ZY-312, and are likely not present.
As potential genetic variation might lead to unpredictable risk, genetic stability should also be confirmed prior to starting large-scale production of ZY-312-based probiotics (Sanders et al., 2010). We confirmed that following in vitro passage of 100 generations from the original strain of ZY-312, there was no significant difference between A100, B100, and the parental strain with respect to morphological characteristics and growth features. No in vivo toxicity was observed for A100 or B100, and the ANI between each generation was at least 99.98%, demonstrating that ZY-312 has a high degree of genetic stability.
Conclusion
We confirmed that ZY-312 is a NTBF strain, without potential virulence factors or risk of spreading antibiotic resistance genes. As well as having desirable probiotic properties (Deng et al., 2016), ZY-312 has a high degree of genetic stability and is non-pathogenic, even to immune-deficient mice. Therefore, ZY-312 is most likely safe for use in future probiotic applications. This study supplements the initial safety assessment work already carried out for ZY-312, and contributes to the development of the first probiotic representative from the dominant Bacteroidetes phylum.
Author Contributions
YW did the experiments with DNA and bacteria, analyzed data, and contributed to revising the manuscript; HD did the experiments with bacteria and mice, analyzed data, and wrote the manuscript; ZL did the experiments with mice, analyzed data, and contributed to revising the manuscript; YT, YH, XW, and ZD analyzed data; YL and RY designed the experiments and contributed to revising the manuscript; YjB designed experiments, analyzed data, and provided overall direction, YB and FZ provided overall directions and contributed to revising the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The property of ZY-312 belongs to Guangzhou ZhiYi biotechnology Co. Ltd. Any use of ZY-312 without permission of Guangzhou ZhiYi biotechnology Co. Ltd. will be illegal.
Acknowledgments
This work was supported by National High Technology Research and Development Program 863 (No. 2015AA020702) and Science and Technology Program of Guangdong, China (Nos. 2015A010101345 & 2016B090918064 & 2016A020217010).
Footnotes
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00435/full#supplementary-material
(A) Colonies of ZY-312 on tryptone soy agar (5% sheep blood) following culture in 5% CO2 (A-A), air (A-B), or anaerobically (A-C) for 48 h at 37°C. (B) Cells observed under light microscope following Gram staining (4000×). (C) Cells observed under scanning electron microscope (5000×).
{kind=link}
Confirmation of no microbial contamination. (A) Escherichia coli and ZY-312 cultured on Eosin methylene blue agar for 24 h at 37°C. (B) Salmonella enterica serovar Paratyphi B and ZY-312 cultured on SS agar for 24 h at 37°C. (C) Clostridium sporogenes and ZY-312 cultured anaerobically on Columbia agar containing blood and gentamicin for 48 h at 37°C. (D) Staphylococcus aureus and ZY-312 cultured on mannitol sodium chloride agar for 24 h at 37°C. (E) Pseudomonas aeruginosa and ZY-312 cultured on NAC agar for 24 h at 37°C. (F) Candida albicans and ZY-312 cultured on Sabouraud dextrose agar for 72 h at 28°C. (G) S. aureus and ZY-312 cultured on agar agar for 48 h at 37°C. (H) C. albicans and ZY-312 cultured on rose bengal agar for 96 h at 28°C.
{kind=link}
The major fatty acids of ZY-312 as identified by gas chromatography.
{kind=link}
Acute toxicity of ZY-312 to mice. Changes in body weight (%) per day were observed for the experimental and control groups. (A) Specific pathogen-free (SPF) BALB/c normal mice (n = 5) were treated with ZY-312 at a concentration of 1 × 109 cfu/day for 5 days and observed for 15 days. The control group mice were treated with saline. (B) SPF normal mice (n = 5) were treated with ZY-312 at a concentration of 5 × 1010 cfu/day for 5 days and observed for 18 days. Tryptic soy broth was used to treat the control group. No significant weight loss was observed in any of the animals (mean ± SE; NS, not significant, t-test).
{kind=link}
Phylogenetic tree based on complete shotgun sequences showing the relationship between Bacteroides fragilis ZY-312 and closely related species. The tree was constructed using the neighbor-joining method.
Phylogenetic tree based on complete genome sequences showing the relationship between B. fragilis strain ZY-312, B. fragilis NCTC 9343, B. fragilis YCH46, and B. fragilis 638R. The tree was constructed using the neighbor-joining method.
References
- Althani A. A., Marei H. E., Hamdi W. S., Nasrallah G. K., El Zowalaty M. E., Al Khodor S., et al. (2016). Human microbiome and its association with health and diseases. J. Cell. Physiol. 231 1688–1694. 10.1002/jcp.25284 [DOI] [PubMed] [Google Scholar]
- Bernardeau M., Vernoux J. P., Gueguen M. (2002). Safety and efficacy of probiotic lactobacilli in promoting growth in post-weaning Swiss mice. Int. J. Food Microbiol. 77 19–27. [DOI] [PubMed] [Google Scholar]
- Borchers A. T., Selmi C., Meyers F. J., Keen C. L., Gershwin M. E. (2009). Probiotics and immunity. J. Gastroenterol. 44 26–46. 10.1007/s00535-008-2296-0 [DOI] [PubMed] [Google Scholar]
- Chen L., Yang J., Yu J., Yao Z., Sun L., Shen Y., et al. (2005). VFDB: a reference database for bacterial virulence factors. Nucleic Acids Res. 33 D325–D328. 10.1093/nar/gki008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cui Y., Yu C., Yan Y., Li D., Li Y., Jombart T., et al. (2013). Historical variations in mutation rate in an epidemic pathogen, Yersinia pestis. Proc. Natl. Acad. Sci. U.S.A. 110 577–582. 10.1073/pnas.1205750110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng H., Li Z., Tan Y., Guo Z., Liu Y., Wang Y., et al. (2016). A novel strain of Bacteroides fragilis enhances phagocytosis and polarises M1 macrophages. Sci. Rep. 6:29401 10.1038/srep29401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- FAO/WHO (2002). Guidelines for the Evaluation of Probiotics in Food: Joint FAO/WHO Working Group on Drafting Guidelines for the Evaluation of Probiotics in Food. Geneva: World Health Organization. [Google Scholar]
- Fernandez M. F., Boris S., Barbes C. (2005). Safety evaluation of Lactobacillus delbrueckii subsp. lactis UO 004, a probiotic bacterium. Res. Microbiol. 156 154–160. 10.1016/j.resmic.2004.09.006 [DOI] [PubMed] [Google Scholar]
- Fijan S. (2014). Microorganisms with claimed probiotic properties: an overview of recent literature. Int. J. Environ. Res. Public Health 11 4745–4767. 10.3390/ijerph110504745 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foligne B., Daniel C., Pot B. (2013). Probiotics from research to market: the possibilities, risks and challenges. Curr. Opin. Microbiol. 16 284–292. 10.1016/j.mib.2013.06.008 [DOI] [PubMed] [Google Scholar]
- Hsiao E. Y., McBride S. W., Hsien S., Sharon G., Hyde E. R., McCue T., et al. (2013). Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 155 1451–1463. 10.1016/j.cell.2013.11.024 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krieg N. R., Ludwig W., Euzéby J., Whitman W. B. (2001). Bergey’s Manual of Systematic Bacteriology, 2nd Edn, Vol. 1 New York, NY: Springer [Google Scholar]
- Lindberg A. A., Weintraub A., Kasper D. L., Lonngren J. (1982). Virulence factors in infections with Bacteroides fragilis: isolation and characterization of capsular polysaccharide and lipopolysaccharide. Scand. J. Infect. Dis. Suppl. 35 45–52. [PubMed] [Google Scholar]
- Liu B., Pop M. (2009). ARDB–antibiotic resistance genes database. Nucleic Acids Res. 37 D443–D447. 10.1093/nar/gkn656 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo R., Liu B., Xie Y., Li Z., Huang W., Yuan J., et al. (2012). SOAPdenovo2: an empirically improved memory-efficient short-read de novo assembler. Gigascience 1:18 10.1186/2047-217X-1-18 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazmanian S. K., Kasper D. L. (2006). The love-hate relationship between bacterial polysaccharides and the host immune system. Nat. Rev. Immunol. 6 849–858. 10.1038/nri1956 [DOI] [PubMed] [Google Scholar]
- Mazmanian S. K., Liu C. H., Tzianabos A. O., Kasper D. L. (2005). An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 122 107–118. 10.1016/j.cell.2005.05.007 [DOI] [PubMed] [Google Scholar]
- Mazmanian S. K., Round J. L., Kasper D. L. (2008). A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 453 620–625. 10.1038/nature07008 [DOI] [PubMed] [Google Scholar]
- McKenney P. T., Pamer E. G. (2015). From hype to hope: the gut microbiota in enteric infectious disease. Cell 163 1326–1332. 10.1016/j.cell.2015.11.032 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miquel S., Beaumont M., Martin R., Langella P., Braesco V., Thomas M. (2015). A proposed framework for an appropriate evaluation scheme for microorganisms as novel foods with a health claim in Europe. Microb. Cell Fact. 14:48 10.1186/s12934-015-0229-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- National Center for Clinical Laboratories (2006). The National Clinical Test Regulation of Operation, 3rd Edn. Nanjing: Southeast University Press. [Google Scholar]
- Ochoa-Reparaz J., Mielcarz D. W., Ditrio L. E., Burroughs A. R., Begum-Haque S., Dasgupta S., et al. (2010). Central nervous system demyelinating disease protection by the human commensal Bacteroides fragilis depends on polysaccharide A expression. J. Immunol. 185 4101–4108. 10.4049/jimmunol.1001443 [DOI] [PubMed] [Google Scholar]
- Rijkers G. T., de Vos W. M., Brummer R. J., Morelli L., Corthier G., Marteau P. (2011). Health benefits and health claims of probiotics: bridging science and marketing. Br. J. Nutr. 106 1291–1296. 10.1017/S000711451100287X [DOI] [PubMed] [Google Scholar]
- Round J. L., Mazmanian S. K. (2010). Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. U.S.A. 107 12204–12209. 10.1073/pnas.0909122107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russell D. W., Sambrook J. (2002). Molecular Cloning. Beijing: Science Press. [Google Scholar]
- Sanders M. E., Akkermans L. M., Haller D., Hammerman C., Heimbach J., Hormannsperger G., et al. (2010). Safety assessment of probiotics for human use. Gut Microbes 1 164–185. 10.4161/gmic.1.3.12127 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekirov I., Russell S. L., Antunes L. C., Finlay B. B. (2010). Gut microbiota in health and disease. Physiol. Rev. 90 859–904. 10.1152/physrev.00045.2009 [DOI] [PubMed] [Google Scholar]
- Shokryazdan P., Faseleh Jahromi M., Liang J. B., Kalavathy R., Sieo C. C., Ho Y. W. (2016). Safety assessment of two new Lactobacillus strains as probiotic for human using a rat model. PLoS ONE 11:e0159851 10.1371/journal.pone.0159851 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan Y., Wu M., Liu H., Dong X., Guo Z., Song Z., et al. (2010). Cellular fatty acids as chemical markers for differentiation of Yersinia pestis and Yersinia pseudotuberculosis. Lett. Appl. Microbiol. 50 104–111. 10.1111/j.1472-765X.2009.02762.x [DOI] [PubMed] [Google Scholar]
- Troy E. B., Kasper D. L. (2010). Beneficial effects of Bacteroides fragilis polysaccharides on the immune system. Front. Biosci. (Landmark Ed.) 15 25–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu T., Yu M., Lin H., Zhang Z., Liu J., Zhang X. H. (2015). Genomic insight into Aquimarina longa SW024T: its ultra-oligotrophic adapting mechanisms and biogeochemical functions. BMC Genomics 16:772 10.1186/s12864-015-2005-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(A) Colonies of ZY-312 on tryptone soy agar (5% sheep blood) following culture in 5% CO2 (A-A), air (A-B), or anaerobically (A-C) for 48 h at 37°C. (B) Cells observed under light microscope following Gram staining (4000×). (C) Cells observed under scanning electron microscope (5000×).
Confirmation of no microbial contamination. (A) Escherichia coli and ZY-312 cultured on Eosin methylene blue agar for 24 h at 37°C. (B) Salmonella enterica serovar Paratyphi B and ZY-312 cultured on SS agar for 24 h at 37°C. (C) Clostridium sporogenes and ZY-312 cultured anaerobically on Columbia agar containing blood and gentamicin for 48 h at 37°C. (D) Staphylococcus aureus and ZY-312 cultured on mannitol sodium chloride agar for 24 h at 37°C. (E) Pseudomonas aeruginosa and ZY-312 cultured on NAC agar for 24 h at 37°C. (F) Candida albicans and ZY-312 cultured on Sabouraud dextrose agar for 72 h at 28°C. (G) S. aureus and ZY-312 cultured on agar agar for 48 h at 37°C. (H) C. albicans and ZY-312 cultured on rose bengal agar for 96 h at 28°C.
The major fatty acids of ZY-312 as identified by gas chromatography.
Acute toxicity of ZY-312 to mice. Changes in body weight (%) per day were observed for the experimental and control groups. (A) Specific pathogen-free (SPF) BALB/c normal mice (n = 5) were treated with ZY-312 at a concentration of 1 × 109 cfu/day for 5 days and observed for 15 days. The control group mice were treated with saline. (B) SPF normal mice (n = 5) were treated with ZY-312 at a concentration of 5 × 1010 cfu/day for 5 days and observed for 18 days. Tryptic soy broth was used to treat the control group. No significant weight loss was observed in any of the animals (mean ± SE; NS, not significant, t-test).
Phylogenetic tree based on complete shotgun sequences showing the relationship between Bacteroides fragilis ZY-312 and closely related species. The tree was constructed using the neighbor-joining method.
Phylogenetic tree based on complete genome sequences showing the relationship between B. fragilis strain ZY-312, B. fragilis NCTC 9343, B. fragilis YCH46, and B. fragilis 638R. The tree was constructed using the neighbor-joining method.