

Summary
Familial hypercholesterolemia (FH) causes elevation of low-density lipoprotein cholesterol (LDL-C) in blood and carries an increased risk of early-onset cardiovascular disease. A caveat for exploration of new therapies for FH is the lack of adequate experimental models. We have created a comprehensive FH stem cell model with differentiated hepatocytes (iHeps) from human induced pluripotent stem cells (iPSCs), including genetically engineered iPSCs, for testing therapies for FH. We used FH iHeps to assess the effect of simvastatin and proprotein convertase subtilisin/kexin type 9 (PCSK9) antibodies on LDL-C uptake and cholesterol lowering in vitro. In addition, we engrafted FH iHeps into the liver of Ldlr−/−/Rag2−/−/Il2rg−/− mice, and assessed the effect of these same medications on LDL-C clearance and endothelium-dependent vasodilation in vivo. Our iHep models recapitulate clinical observations of higher potency of PCSK9 antibodies compared with statins for reversing the consequences of FH, demonstrating the utility for preclinical testing of new therapies for FH patients.
Keywords: familial hypercholesterolemia, hepatocytes, human liver chimeric mice, induced pluripotent stem cells, LDL receptor, PCSK9 antibodies, statins, genetic engineering, endothelium-dependent vasodilation
Graphical Abstract
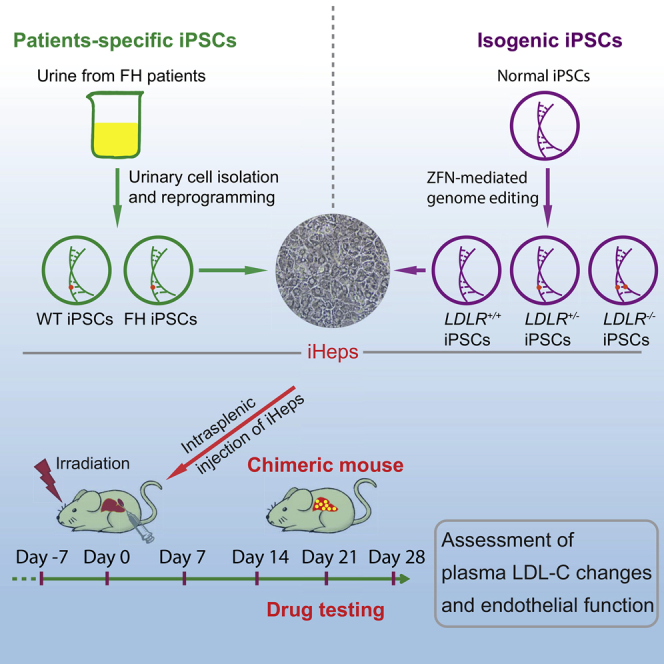
Highlights
-
•
Generation of a comprehensive stem cell model for familial hypercholesterolemia
-
•
Generation of chimeric mice with engrafted human iPSC-derived iHeps
-
•
FH iHeps respond to LDL-C-lowering medications in vitro and in vivo
-
•
Our model can be used for preclinical testing of novel LDL-C-lowering medications
In this paper, Tse, Esteban and colleagues have generated chimeric mice with human hepatocytes derived from familial hypercholesterolemia (FH) induced pluripotent stem cells. These chimeric mice resemble FH patients in the response to clinical-grade PCSK9 antibodies and statins. These results indicate the utility of this stem cell model for preclinical testing of new LDL-C-lowering therapies for FH patients.
Introduction
Mutations in LDLR (encoding LDL receptor, LDLR), often heterozygous, underlie most cases of familial hypercholesterolemia (FH), which predisposes to premature cardiovascular disease due to marked elevation of plasma levels of lipids, in particular low-density lipoprotein cholesterol (LDL-C) (Brown and Goldstein, 1986). Besides diet control and physical activity, FH patients are treated with statins, a class of drugs that inhibit 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase and hence reduce cholesterol synthesis in the liver (Endo, 1992). Statins also increase LDLR protein levels in hepatocytes and LDL-C clearance from plasma. Because of these properties, statins are used to treat FH patients and also patients with non-familial hypercholesterolemia. However, statins fail to reduce plasma LDL-C adequately in the majority of these patients for prevention of cardiovascular events (Cannon et al., 2015, Reiner, 2015), and a proportion of patients suffers from significant adverse effects (Dormuth et al., 2014, Stroes et al., 2015).
Importantly, FH can be caused by mutations in other genes besides LDLR, including gain-of-function mutations in proprotein convertase subtilisin/kexin type 9 (PCSK9) (Sniderman et al., 2014). This observation led to the subsequent identification of PCSK9 as an extracellular protein responsible for the internalization and degradation of surface LDLR in hepatocytes (Maxwell et al., 2005). Interestingly, PCSK9 is induced by statins, and activates a negative feedback loop that controls LDLR expression and consequently restrains the efficacy of statins (Dubuc et al., 2004). The latter may account for the limited efficacy or even loss of efficacy of statins in some patients, and has suggested that PCSK9 modulation could be of therapeutic relevance. In this regard, clinical trials with anti-PCSK9 therapy (in the form of monoclonal antibodies) have shown potent lowering effects on LDL-C and good tolerance, creating a tremendous interest in this approach as an alternative medication for hypercholesterolemia (Robinson et al., 2015, Roth et al., 2012). Besides the potential paradigm shift in FH treatment, these findings imply that interfering with additional components of the LDLR degradation pathway (besides PCSK9) may provide alternative therapeutic avenues for hypercholesterolemia.
Despite the exciting developments with PCSK9 antibodies, a relevant caveat for further exploration of new therapies (new drugs or new formulations) for FH (and also other genetic liver diseases) is the lack of easily accessible and bona fide in vitro and in vivo models for preclinical testing. In vitro models for FH are mostly based on patient-derived hepatocytes from liver biopsy (Cayo et al., 2012), which have limited accessibility and cannot be expanded in culture. In vivo models including the hyperlipidemic rabbit, the rhesus macaque, and the Ldlr knockout mouse, have the limitation of not fully recapitulating human hepatocyte function (Bissig-Choisat et al., 2015). Patient-specific induced pluripotent stem cells (iPSCs) can provide an unlimited source of differentiated cell types including hepatocytes (iHeps) that can be used for in vitro and in vivo studies (Grskovic et al., 2011, Takahashi et al., 2007). This approach combined with the transplantation into immunodeficient mice may help overcome existing problems in modeling FH in vitro and in vivo (Carpentier et al., 2014, Chen et al., 2012, Liu et al., 2011). Several groups have generated FH iPSCs that harbor mutations in LDLR (Cayo et al., 2012, Ramakrishnan et al., 2015, Rashid et al., 2010) or PCSK9 (Si-Tayeb et al., 2016) and have tested the ability of the derived iHeps to mimic the disease phenotype and respond to statins in vitro. However, there are no reports so far testing the effect of anti-PCSK9 therapies on FH iPSC-derived iHeps in vitro, or in vivo disease modeling and drug testing with FH iHeps transplanted into appropriate animal models.
Here, we report that FH iHeps derived from patient-specific and genetically engineered FH iPSCs can be used to test the efficacy of two well-known medications for lowering LDL-C, statins and PCSK9 antibodies, not only in vitro but also in vivo, by engrafting FH iHeps into the liver of immunodeficient mice knockout for Ldlr. With these assays, we have confirmed clinical observations of higher potency of PCSK9 antibodies compared with statins for treating FH (Robinson et al., 2015), indicating the potential of our model for further drug discovery and preclinical testing of experimental therapies.
Results
Generation of a Panel of Patient-Specific and Genetically Engineered FH iPSCs for In Vitro and In Vivo Studies
We selected a family with clinical FH due to heterozygous duplication of “TGCTGGC” (c.2108_2114dup, p.Ala705fsX14) in exon 14 of LDLR (Khoo et al., 2000) (Figures 1A and 1B), which results in a premature stop codon. Using urinary cells as a donor cell source (Benda et al., 2013, Zhou et al., 2011) and episomal vectors as the reprogramming method (Yu et al., 2009), we generated integration-free iPSCs from the two affected sisters (FH-1 and FH-2) and the healthy sister (wild-type, WT) (Figures 1A and S1A); individual clones for each person were selected for further study. The resulting cell lines were fully pluripotent as shown by immunofluorescence (SSEA-4 and NANOG), RT-qPCR (OCT4, SOX2, NANOG, and TERT), DNA demethylation of the NANOG proximal promoter, and the formation of teratomas in immunocompromised mice (Figures S1B–S1E). Moreover, their karyotypes were normal (Figure S1C) and after serial passaging there was no remnant of the episomal vectors employed for reprogramming, as tested by PCR (Figure S1F). We also confirmed the LDLR mutation and the reduced expression of LDLR protein in the two FH iPSC clones by sequencing and western blotting, respectively (Figures 1B and 1C).
Figure 1.

Generation of a Panel of FH iPSCs
(A) Family tree of WT and FH patients. Asterisk indicates patient-specific iPSCs generated in this study.
(B) Schematic depicting the genomic region of LDLR mutations in FH iPSCs. The boxed area indicates the position of heterozygous duplicate of “TGCTGGC” in FH-1 iPSCs.
(C) Western blotting shows LDLR levels in HepG2 cells (control) and iPSCs. ACTIN was used as loading control.
(D) Genotype of a panel of FH isogenic knockout iPSC clones. Red labels the interval of both ZFN-recognized fragments; black in lowercase among the red indicates insertion that resulted in frameshift.
(E) Phase contrast and immunofluorescence for ASGPR and A1AT of iHeps at day 17 of differentiation. Scale bars represent 50 μm.
(F) Bar graph shows the percentage of ASGPR+ iHeps obtained with our differentiation protocol as measured by flow cytometry. A representative experiment with samples measured in triplicate is shown; error bars indicate SD.
Next, because variations in the genetic background among iPSCs can be a confounding factor for in vitro disease modeling (Soldner et al., 2011), we used zinc-finger nucleases (ZFNs) (Moehle et al., 2007) targeting exon 4 of LDLR (Figure S2A) to create a panel of isogenic LDLR knockout iPSCs in a previously generated and well-characterized urinary iPSC clone from an unaffected individual (Zhou et al., 2011). This produced two biallelic knockout clones and 27 monoallelic knockout clones out of 214 clones screened by sequencing (Figure S2B). Of the resulting cell lines, we selected one monoallelic knockout (+/−) clone, one biallelic knockout (−/−) clone, and the unmodified control (+/+) as a panel for further study (Figure 1D). These isogenic iPSCs retained pluripotency, displayed normal karyotypes, and expressed LDLR levels according to their genotypes (Figures 1C, S2C, and S2D). Therefore, we have generated two complementary panels of iPSCs for studying FH mechanisms and performing drug screening.
PCSK9 Antibodies Show Stronger LDL-Lowering Ability than Statins in FH iHeps
We differentiated our panel of patient-specific and isogenic FH iPSCs into functional iHeps with a previously reported three-step protocol (Kajiwara et al., 2012). After 17 days of differentiation, iHeps with equivalent morphology to human hepatocytes appeared (Figure 1E). These iHeps displayed markers of primary hepatocytes including asialoglycoprotein receptor (ASGPR) and α1-antitrypsin (A1AT) (Figure 1E), stored glycogen and lipids, secreted albumin and urea, and had the ability to metabolize rifampicin via the cytochrome P450 pathway (Figures S3A–S3E). Importantly, the efficiency of iHep differentiation between patient-specific and isogenic FH iPSCs was comparable, as measured by analyzing the population of ASGPR+ cells by flow cytometry (Figure 1F).
To reveal the main phenotypic feature of FH (inability to uptake plasma LDL-C by hepatocytes) in vitro, we incubated iHeps from the various iPSC lines with 5 μg/mL fluorescently labeled LDL for 3.5 hr, followed by co-staining with hepatocyte nuclear factor 4α (HNF4A) to confirm the identity of the iHeps. As expected, WT and +/+ iHeps showed high—and rather comparable—LDL uptake while this was significantly impaired in FH-1, FH-2, and +/− iHeps, and completely abolished in −/− iHeps (Figure 2A). Similar results were obtained when LDL capture was measured using flow cytometry and normalized by the number of ASGPR+ cells (Figure 2B, left panel). With the same assay, we also observed that 10 μM simvastatin elevated LDL uptake in FH-1 iHeps and +/− iHeps (Figure 2B, left panel), and more modestly in WT and +/+ iHeps. However, there was only marginal basal LDL uptake, without elevation with simvastatin, in −/− iHeps (Figure 2B, left panel). In addition, we measured total cholesterol (TC) and apolipoprotein B (APOB) secretion (an indirect measure of LDL secretion) (Cayo et al., 2012) in our panel of iHeps. TC and APOB were increased in FH-1 iHeps compared with WT, and in +/− and −/− iHeps compared with +/+ iHeps (Figure 2B, middle and right panels). On the other hand, 10 μM simvastatin reduced TC and APOB levels in iHeps from all tested iHeps but in particular in +/−, −/−, and FH-1 iHeps (Figure 2B, middle and right panels).
Figure 2.

PCSK9 Antibodies Have a Stronger Effect than Statins on LDL Uptake by FH iHeps In Vitro
(A) Micrographs show co-localization of fluorescently labeled LDL (green) and HNF4A (red). Scale bars represent 50 μm.
(B) Left panel: bar graph shows percentage of DiI-LDL capture by flow cytometry in ASGPR+ iHeps with or without simvastatin. Statistical tests were performed with three independent experiments; p values are indicated on the graph and were obtained using two-way ANOVA adjusted with Tukey's multiple comparison. Error bars indicate SEM. Middle and right panels: bar graphs show concentration of TC and APOB secretion in iHeps culture as determined by ELISA; values were normalized by the concentration of secreted hALB. Statistical tests were performed on three independent experiments; p values are indicated on the graphs and were obtained using a Kruskal-Wallis test; error bars indicate SEM. Vehicle: DMSO.
(C) Western blotting shows that PCSK9 is induced by simvastatin in iHeps.
(D) LDL uptake capacity of healthy and heterozygous FH iHeps was measured with microplate readers and indicated as relative fluorescence units. Cells were treated with simvastatin and/or PCSK9 antibodies for 16 hr on days 17–21 of differentiation. Statistical tests were performed on three independent cell lines for both healthy (+/+, WT iHeps, and iHeps from another unaffected FH family member) and heterozygous FH iHeps (+/−, FH-1, and FH-2 iHeps). p Values are indicated on the figure and were obtained using one-way ANOVA adjusted with Dunnett's multiple comparison; error bars indicate SEM.
In the clinic, the inefficiency of statins in reducing LDL-C in some FH patients is partly due to increased PCSK9 expression, as this promotes LDLR degradation (Maxwell et al., 2005). To confirm whether PCSK9 is also upregulated by statins in iHeps in vitro, we performed western blotting of FH-1 and WT iHeps untreated or treated with simvastatin, observing that the treatment increases PCSK9 expression (Figure 2C). We then developed a 48-well-based assay to compare systematically the effect of different concentrations of statins and PCSK9 antibodies on LDL uptake in iHeps. In isolation, PCSK9 antibodies were more potent than simvastatin in inducing LDL uptake in healthy and heterozygous FH iHeps, and their combination did not increase it further (Figure 2D). Neither simvastatin nor PCSK9 antibodies had any effect on LDL uptake in −/− iHeps (Figure S3F). These in vitro results reproduce clinical observations showing that PCSK9 antibodies are a remarkable substitute for statins in treating heterozygous FH patients but are ineffective for homozygous FH patients (Stein et al., 2013).
Engrafted iHeps Reduce Plasma LDL-C Levels in an FH Chimeric Mouse Model
To further expand our iHep platform and perform drug testing in vivo, we crossed Ldlr−/− mice (Ishibashi et al., 1993) with Rag2−/−/Il2rg−/− mice, and so generated triple-knockout LRG mice (Figure 3A). LRG mice lack an effective adaptive immune system, allowing the transplantation of human iHeps into an LDLR null background without rejection. To demonstrate this, we engrafted iHeps from FH iPSCs into LRG mice via intrasplenic injection. To improve the repopulation efficiency, we exposed mice to 3 Gy of γ-ray radiation 24 hr prior to engraftment to induce moderate liver injury and so enable proliferation of the engrafted iHeps. In addition, we suspended the injected iHeps in 50% Matrigel to provide a microenvironment that facilitates migration into the liver from the spleen (Figure 3B). The evidence of liver repopulation with iHeps in LRG mice was assessed by immunohistochemical staining for LDLR, human albumin (hALB), and human nuclear antigen (hNA) (Figures 3C and 3D). To determine iHep engraftment efficiency, we first stained chimeric mouse liver sections with hALB antibodies and then scanned the whole sections to quantify hALB+ areas (Figures 3E, 3F, and S4A). With this strategy, we were able to assess iHep engraftment efficiency in multiple lobules and lobes rather than in small-area images. All tested iHeps showed significant repopulation in LRG mouse liver 3 weeks after transplantation, based on hALB staining, although with slight variations in efficiency (ranging from ∼5% to ∼10%) (Figure 3G). Because hALB is a secreted protein and dispersion could influence the assessment, we also used hNA to calculate iHep engraftment efficiency, which showed a moderately higher value than with hALB (∼15% on average for different iHeps) (Figure 3H). No evidence of hALB+ or hNA+ cells was observed in LRG mouse liver engrafted with vehicle (Figures 3D and S4A). Likewise, there was no positive signal in the spleen of animals injected with iHeps (Figure S4B), demonstrating that transplanted cells do not engraft there after injection.
Figure 3.

Generation of Chimeric Mice Engrafted with FH iHeps
(A) Schematic view of derivation of LRG mice.
(B) Strategy for generation of FH chimeric mice, and assessment of LDL-C level and endothelial function.
(C and D) Immunohistochemical staining for LDLR, hNA, and hALB in LRG mouse liver repopulated with the indicated iHeps or Matrigel (vehicle). Zoomed images for LDLR and hNA are shown. Scale bars represent 50 μm (C) and 200 μm (D).
(E and F) Whole-slide scan image (E) and snapshot images (F) of hALB staining in mouse liver repopulated with +/+ iHeps. The location of snapshot images in the whole section is shown in the corner. Scale bars represent 2 mm (E and top left image in F) and 400 μm (remaining three images in F).
(G) Scatter diagram shows the percentage of engrafted hALB+ iHeps in LRG mouse liver calculated based on whole-slide scanning images. Each dot represents one mouse with no less than four sections analyzed. Error bars indicate SEM.
(H) Bar graph shows the percentage of repopulated hNA+ iHeps in LRG mouse liver (n = 4, including sections of LRG mice engrafted with +/+ iHeps × 1, +/− iHeps × 2, and −/− iHeps × 1); quantification of hNA+ cells was performed by manual counting (see Supplemental Experimental Procedures for detailed description). Error bars indicate SEM.
(I) Plasma LDL-C levels in LRG mice engrafted with the indicated iHeps under normal diet (left panel) and HFHC diet (right panel) assessed at the indicated time points after iHep transplantation; n indicates number of mice. p Values are indicated on the figure and were obtained using a Kruskal-Wallis test; error bars indicate SEM.
(J) Percentage change of plasma LDL-C from baseline at weeks 1, 4, 8, and 12 post-transplantation in LRG mice fed with HFHC diet and engrafted with +/+ iHeps or pHH; n indicates number of mice. p Values are indicated on the figure and were obtained using a Kruskal-Wallis test; error bars indicate SEM.
(K) Immunohistochemical staining for hALB and hNA in LRG mouse liver repopulated with +/+ iHeps and pHH 12 weeks post-engraftment. Scale bars represent 200 μm.
Fed a normal diet, LRG mice engrafted with +/+ iHeps had lower, but not statistically significant, levels of plasma LDL-C (n = 6, 57.9 ± 3.1 mg/dL) compared with vehicle (n = 4, 112.7 ± 20.6 mg/dL) 3 weeks after transplantation, while +/− iHeps (n = 4, 72.5 ± 12.0 mg/dL) and FH-1 iHeps (n = 5, 70.1 ± 12.7 mg/dL) were less effective (Figure 3I, left panel). As expected, −/− iHeps had no effect on LDL-C clearance (n = 6, 119.6 ± 6.9 mg/dL) compared with vehicle (Figure 3I, left panel). We then fed LRG mice with a high-fat, high-cholesterol (HFHC) diet to see whether this magnifies differences in LDL-C lowering after iHep transplantation. LRG mice showed around a 3-fold increase in LDL-C plasma after 1 week on HFHC diet (Figure 3I, right panel). Under this condition, transplanted +/+ iHeps (n = 5, 275.1 ± 19.0 mg/dL), and to a lesser extent +/− iHeps (n = 6, 439.3 ± 35.6 mg/dL) and FH-1 iHeps (n = 6, 414.7 ± 42.0 mg/dL), significantly downregulated plasma LDL-C levels compared with vehicle (n = 6, 606.7 ± 27.7 mg/dL) or −/− iHeps (n = 6, 617.3 ± 22.8 mg/dL) (p < 0.01, Figure 3I, right panel). To assess the long-term effects of engrafted iHeps compared with primary human hepatocytes (pHH), we tested LDL-C levels 12 weeks after transplantation of +/+ iHeps and pHH into LRG mice fed with HFHC diet, observing notable and rather comparable plasma LDL-C clearance in both cases compared with vehicle (p < 0.05, Figure 3J). The evidence of engrafted cells at this time point was assessed by staining for hALB and hNA in liver sections (Figure 3K). Overall, these experiments demonstrate that transplantation of LDLR-competent but not LDLR null iHeps promotes plasma LDL-C clearance in LRG mice.
Alirocumab Shows Stronger LDL-Lowering Ability than Statins in Chimeric LRG Mice
We then used HFHC diet to assess the effect of PCSK9 antibodies compared with statins on iHeps engrafted into LRG mice (Figure 4A). Importantly, we used a clinical-grade formulation of PCSK9 monoclonal antibodies, alirocumab (Sanofi and Regeneron Pharmaceuticals), which has completed phase III clinical trials (Robinson et al., 2015) and was approved by the Food and Drug Administration in July 2015 as a cholesterol-lowering drug. Alirocumab at 10 mg/kg/week enhanced plasma LDL-C clearance by engrafted +/− iHeps (Figure 4B; n = 3, percentage reduction from baseline 50.06% ± 7.38%, p < 0.05 compared with +/− iHeps alone) and FH-1 iHeps (Figure 4C; n = 3, percentage reduction from baseline 61.29% ± 10.80%, p < 0.05 compared with FH-1 iHeps alone) 21 days after treatment. Simvastatin at 10 mg/kg/day also reduced LDL-C clearance by engrafted +/− iHeps (n = 3, percentage reduction 33.37% ± 10.55%, p > 0.05 compared with iHeps alone) and FH-1 iHeps (n = 5, percentage reduction 28.39% ± 9.55%, p > 0.05 compared with iHeps alone), but less significantly than alirocumab, and their combination was not synergistic (Figures 4B and 4C). Moreover, in chimeric mice engrafted with −/− iHeps, alirocumab alone had no effect, while simvastatin alone or combined with alirocumab had only a modest effect on LDL-C clearance (Figure 4D). This moderate effect of simvastatin (alone or in combination) on engrafted −/− iHeps is likely due to inhibition of HMG-CoA reductase (Endo, 1992). Notably, the observed percentage reduction of plasma LDL-C with alirocumab in LRG mice engrafted with +/− iHeps or FH-1 iHeps is similar to that reported in human studies (Robinson et al., 2015, Roth et al., 2012). These results show the potential utility of LRG mice engrafted with FH iHeps for preclinical testing of novel drugs for FH.
Figure 4.

In Vivo Drug Testing with FH Chimeric Mice
(A) Strategy for in vivo drug testing using LRG mice engrafted with FH iHeps.
(B–D) Percentage change of plasma LDL-C from baseline at the indicated time points after iHep transplantation in LRG mice fed with HFHC diet and treated with simvastatin and/or PCSK9 antibodies; n indicates number of mice. p Values are indicated on the figure panel and were obtained using a Kruskal-Wallis test and error bars indicate SEM.
Improvement of Endothelium-Dependent Vasodilation in Chimeric LRG Mice by Alirocumab and Statins
One consequence of FH is the accelerated development of atherosclerosis on the arterial wall (Vanhoutte et al., 2016), but we did not observe any obvious development of atherosclerotic lesions (assessed by oil red O staining) in our LRG mice fed with HFHC diet (data not shown). This is likely due to their compromised immune system and the relatively short period on the HFHC diet. Impaired endothelial function is an early indicator of atherosclerosis in children and young adults with FH (Wiegman et al., 2004), which manifests as reduced flow-mediated endothelium-dependent vasodilation (EDV) and increased proinflammatory gene expression, while EDV in response to glyceryl trinitrate in these patients is comparable with that in control subjects (Aggoun et al., 2000). We found that EDV induced by cumulative concentrations of acetylcholine (ACh) was significantly improved in aortic rings of LRG mice engrafted with +/+ iHeps compared with LRG mice engrafted with FH-1 iHeps, +/− iHeps, or −/− iHeps (p < 0.01, Figure 5A). However, endothelium-independent vasodilation induced by sodium nitroprusside was similar in endothelium-denuded aortic rings from all LRG groups (Figure 5B). Alirocumab at 10 mg/kg/week improved EDV significantly in LRG mice engrafted with +/− (p < 0.01) or FH-1 iHeps (p < 0.05, Figures 5C and 5D), while there was no effect on LRG mice engrafted with −/− iHeps or vehicle (Figures 5E and 5F). In contrast, 10 mg/kg/day simvastatin improved EDV in aortas of all groups (Figures 5C–5F), supporting the knowledge that the vasoprotective effect of simvastatin is partially independent of LDLR status (Tian et al., 2011). Moderate synergistic effect of simvastatin and alirocumab on EDV in aortic rings was observed in LRG mice engrafted with +/− iHeps or FH-1 iHeps, but not −/− iHeps or vehicle (Figures 5C–5F). Simvastatin or alirocumab did not have any effect on sodium nitroprusside-induced relaxation in aortic rings from LRG mice engrafted with all groups of iHeps or vehicle (Figures S5A–S5D). Furthermore, RT-qPCR of aorta samples from LRG mice engrafted with +/− iHeps showed that treatment with alirocumab, simvastatin, or their combination significantly reduced the expression of cytokines, chemokine receptors, and adhesion molecules related to vascular inflammation (Figure S5E). Therefore, EDV in aortas is a good readout for monitoring the consequences of FH in LRG mice engrafted with FH iHeps, and is responsive to treatment with PCSK9 antibodies or statins.
Figure 5.

Alirocumab and Statins Improve EDV in Chimeric LRG Mice
(A and B) EDV in response to cumulative concentrations of acetylcholine (ACh) (A) and endothelium-independent vasodilation to sodium nitroprusside (SNP) (B) in aortas (precontracted with phentolamine [Phe]) from LRG mice fed with HFHC diet. Mice were engrafted with the indicated iHeps or vehicle; n indicates number of mice. p Values are indicated on the figure and were obtained using two-way ANOVA adjusted with Dunnett's multiple comparison; error bars indicate SEM.
(C–F) EDV in response to ACh in aortae (precontracted with Phe) from LRG mice fed with HFHC diet. Mice were engrafted with the indicated iHeps and treated with simvastatin (daily) and/or alirocumab (weekly); n indicates number of mice. p Values are indicated on the figure and were obtained using two-way ANOVA adjusted with Dunnett's multiple comparison; error bars indicate SEM.
Discussion
Patient-specific iPSCs can be differentiated into disease-relevant cells, thus providing an unlimited source of human cells for in vitro studies (Grskovic et al., 2011). Using this disease-in-a-dish approach, multiple human conditions including liver diseases have been modeled (Li et al., 2012, Rashid et al., 2010, Zhang et al., 2011). In this study, we have employed patient-specific FH iPSCs and genetically engineered isogenic FH iPSCs to create a platform for in vitro and in vivo drug testing of LDL-C-lowering therapies.
Several groups have previously reported FH iPSCs harboring mutations in LDLR and PCSK9 and the ability of the derived iHeps to increase LDL uptake in vitro upon treatment with statins (Cayo et al., 2012, Ramakrishnan et al., 2015, Si-Tayeb et al., 2016). Our work has focused on testing therapies for FH, in particular anti-PCSK9 antibodies, in vitro and in vivo using FH iHeps, although this has been previously studied in vitro using pHH (Zhang et al., 2012). Likewise, FH disease modeling with isogenic iPSCs, which allow healthy versus diseased comparison of iHeps that share the same genetic background, had not been described beforehand. The addition of isogenic iPSCs may overcome the shortcomings of using a small number of patient-specific iPSCs and clones for identifying FH phenotypes in this study. Consistent with previous studies using FH iHeps (Cayo et al., 2012, Ramakrishnan et al., 2015, Si-Tayeb et al., 2016), our in vitro experiments confirm that statins increase LDL uptake in FH iHeps and also demonstrate a concomitant reduction in TC and APOB level. To facilitate the systematic screening of LDL-C-lowering medications with FH iHeps, we designed a 48-well assay based on labeled LDL capture. This assay showed higher efficacy in increasing LDL capture of PCSK9 antibodies compared with statins, and in the future may prove useful for identifying new candidate drugs for treating FH through systematic screening. In this regard, our assay may be adaptable to 96-well plates through further optimization.
Human liver chimeric mice carrying human pHH can facilitate in vivo study of viral hepatitis infections (Bissig et al., 2010, Carpentier et al., 2014) and liver genetic diseases (Bissig-Choisat et al., 2015, Yusa et al., 2011), and the testing of small molecular drug candidates and biological components including antibodies (Legrand et al., 2009). Embryonic stem cell (ESC)- or iPSC-derived iHeps have been used to generate chimeric mice by engrafting them into the liver of non-obese diabetic severe combined immunodeficient (NOD-SCID) mice (Chen et al., 2012), NOD/Lt-SCID/IL-2Rγ−/− (NSG) mice (Liu et al., 2011), or uPA transgenic mice (Basma et al., 2009, Carpentier et al., 2014), where iHeps showed further maturation and regeneration potential and displayed key activities of primary hepatocytes. Our study has produced a mouse model that lacks the gene (Ldlr) involved in a human disease (FH) and is engrafted with patient-specific iHeps to mimic the specific human disease condition and perform in vivo drug testing. This approach has significant advantages over studies using Ldlr knockout mice alone (Ishibashi et al., 1993), as endogenous hepatocytes in the latter mouse model are not responsive to drugs modulating the LDLR pathway. A recent study showed engraftment of FH patient-specific iHeps injected subcutaneously rather than into the liver parenchyma of Rag1−/−/Ldlr−/− mice, but the authors did not perform an assay to demonstrate functional recovery (Ramakrishnan et al., 2015). Another report demonstrated the feasibility of directly engrafting FH (due to compound heterozygosity in LDLR gene) pHH into Fah−/−/Rag2−/−/Il2rg−/− mice, and the phenotype of FH was successfully rescued by LDLR gene therapy using adeno-associated virus (Bissig-Choisat et al., 2015). Because of the lack of Fah, these mice had higher repopulation efficiency (>70%) (Bissig-Choisat et al., 2015) than our mice. However, a problem of this approach is that a population of mouse hepatocytes with intact LDLR remains while the engrafted patients' pHH did not contain functional LDLR, which can be confounding factors when performing in vivo testing of drugs acting on the LDLR pathway. Moreover, primary FH hepatocytes are difficult to obtain and cannot be expanded in vitro. Although liver repopulation in our chimeric mice is significantly lower than for Fah−/−/Rag2−/−/Il2rg−/− mice engrafted with pHH (Azuma et al., 2007, Bissig-Choisat et al., 2015), it is comparable with results of recent reports describing NSG mice or Gunn rats (a model of Crigler-Najjar syndrome 1) engrafted with iPSC-derived iHeps (2%–17% [Liu et al., 2011] and 5.1% [Chen et al., 2015]). In the future, crossing our LRG mice with Fah knockout mice could help achieve higher liver chimerism, and thus further upgrade in vivo drug testing studies using engrafted FH iHeps. It should also be considered that mice and humans bear differences in lipoprotein metabolism, and therefore it may be desirable to genetically engineer other mammalian species (e.g., rabbits) (Shiomi and Ito, 2009) for more accurate studies using transplanted human FH iHeps. Nevertheless, despite the physiological differences between mice and humans and the moderate chimerism of our own model, the lowering of plasma LDL-C achieved with LDLR-competent iHeps was significant, while FH iHeps were less effective but exhibited good response to statins and particularly to PCSK9 antibodies. Besides plasma LDL-C levels, we measured endothelial function in aortas of LRG mice to assess the efficacy of engrafted iHeps and these two medications in vivo, as this parameter is affected early in patients with FH (Wiegman et al., 2004). Our results showed that cell therapy and the two medications could improve EDV in chimeric mice, indicating that endothelial function can be used as a parameter to evaluate disease progression during preclinical testing in chimeric LRG mice or other similar animal models. Importantly, we also noticed that the engrafted iHeps could be maintained in LRG mouse liver, and were functional, for at least 3 months. Overall, these results highlight the potential relevance of transplanting iPSC-derived iHeps for the treatment of hereditary metabolic liver disorders (Cantz et al., 2015), and the use of chimeric animals humanized with disease-specific iHeps for preclinical evaluation of novel therapies. However, the long-term therapeutic efficacy of iPSC-based approaches remains unclear, and future studies are necessary to address this as well as safety concerns.
In summary, our work provides a state-of-the-art in vitro and in vivo platform for testing drugs and experimental cell therapies using FH iHeps. Similar platforms using other gene knockout mice and alternative patient-specific iHeps may be useful to investigate pathological mechanisms and perform preclinical studies for unrelated inherited liver diseases.
Experimental Procedures
DNA Sample Preparation and Sequencing
Genomic DNA was extracted from blood of subjects with FH using the QIAamp DNA blood kit (Qiagen). All exons of the LDLR gene locus were sequenced by Sanger sequencing, and mutations were identified by aligning with the reference sequence GenBank: NM_000527.4 from NCBI.
Cell Culture and Reprogramming
Urinary cells were obtained and expanded according to our published protocol (Zhou et al., 2012) with approval from the Institutional Review Board (HKCTR-725, http://www.hkclinicaltrials.com). Written informed consent from all patients is available upon request. Cells were reprogrammed with episomal vectors (Addgene) that encode human OCT4, SOX2, KLF4, MYC, LIN28, and SV40LT (Yu et al., 2009), using a previously reported protocol (Yu et al., 2011) with the combination of small molecular compounds containing 0.5 μM PD0325901 (Stemgent), 3 μM CHIR99021 (Stemgent), 0.5 μM A-83-01 (Stemgent), 1,000 U/mL human LIF (Millipore), 10 μM Y27632 (Sigma-Aldrich), and 100 ng/mL basic fibroblast growth factor (Peprotech). Human ESC-like colonies were picked at around day 25 and expanded in mTeSR1 (StemCell) on Matrigel (Corning)-coated plates with Accutase (Sigma-Aldrich).
Knockout Cell Line Generation
Sigma-Aldrich donated the LDLR ZFNs. LDLR-ZFN mRNAs were generated through in vitro transcription of LDLR-ZFN plasmids with a MessageMAX T7 ARCA-Capped Message Transcription Kit (Cambio) according to the manufacturer's specifications. The parental iPSC clone was reported previously (UC C0406-iPS C4) (Zhou et al., 2011). Detailed procedures are described in Supplemental Experimental Procedures.
In Vitro Differentiation of iPSCs into iHeps
Human iPSCs were differentiated into iHeps as reported previously (Kajiwara et al., 2012) but with small modifications at the endoderm induction stage: RPMI-1640 containing 100 ng/mL activin A (PeproTech) and 25 ng/mL WNT3a (R&D Systems) was employed for 1 day when the iPSCs had reached around 70% confluence, after which WNT3a was omitted from the medium for the following 2 days. HGF was purchased from PeproTech, Oncostatin M from R&D Systems, RPMI-1640, KnockOut DMEM, KnockOut Serum Replacement, and GlutaMAX Supplement from Life Technologies, DMSO from Sigma-Aldrich, and the HCM Bullet Kit from Lonza.
hALB, TC, and APOB Measurements
hALB, TC, and APOB measurements were performed on supernatants of day-17 to -18 (24-hr interval) iHeps culture by ELISA using a kit (Cell Biolabs for hALB and TC; MABTECH for APOB), in 96-well assay plates according to the manufacturer's specifications.
LDL Uptake Assays
LDL uptake assays employed fluorescently labeled LDL and 3,3′-dioctadecylindocarbocyanine (DiI)-LDL (Life Technologies), which were measured by microscopy or flow cytometry. Detailed procedures are described in Supplemental Experimental Procedures.
PCSK9 Antibodies and Simvastatin Treatments In Vitro
iPSCs were split onto 48-well plates and differentiated into iHeps as described above. At days 17–21 of differentiation, iHeps were treated with simvastatin and/or PCSK9 monoclonal antibodies (Cayman Chemical) for 16 hr and then incubated with 5 μg/mL DiI-LDL for 1 hr. Cells were washed with Ca2+/Mg2+-free PBS (CMF-PBS, Hyclone) and the DiI-LDL signal was captured with a CLARIOstar microplate reader (BMG LABTECH). Cells were then fixed with 4% paraformaldehyde and stained with ASGPR antibodies (Santa Cruz Biotechnology). Signal was read again with the microplate reader. Fluorescence signals were analyzed by MARS Data Analysis Software and normalized by the number of iHeps indicated by the ASGPR signal. Relative fluorescence units refer to the fold change of fluorescence values compared with the baseline (null treatment).
Chimeric Mice Generation
The animal study was approved by the Institutional Review Board (CULATR-2592-11 and 3003-13). Ldlr–/– mice (B6.129S7-Ldlrtm1Her/J, The Jackson Laboratory) were crossed with Rag2−/−/Il2rg–/– mice (available in our lab by crossing of B6(Cg)-Rag2tm1.1Cgn/J mice with B6.129S4-Il2rgtm1.wWjl/J mice purchased from The Jackson Laboratory) to generate LRG mice. Genotype was confirmed by PCR and sequencing using genomic DNA from mouse ear. As recipient for iHeps, we used 8- to 16-week-old LRG mice. Mice were irradiated with 3 Gy of γ-ray using a Gammacell 3000 Elan II (MDS Nordion) machine 24 hr prior to cell engraftment. A total of 106 iHeps or pHH (Lonza) suspended in 50 μL of 5.1 mg/mL Matrigel (Basement Membrane Matrix, Corning 354234) in CMF-PBS solution were engrafted into mouse liver by intrasplenic injection.
In Vivo Drug Testing
For the HFHC diet group, mice were fed with HFHC diet from 7 days prior to engraftment. For the drug treatment group, drugs were administered from 7 days post-engraftment. The clinical-grade PCSK9 monoclonal antibody formulation alirocumab (SAR236553/REGN727, Sanofi and Regeneron Pharmaceuticals) was injected subcutaneously (10 mg/kg/week) with or without simvastatin (Zocor; Merck Sharp & Dohme) treatment (10 mg/kg/day in drinking water) for 21 days. Blood samples were collected from the facial vein at the indicated time points and the day of engraftment of iHeps was set as day 0 or baseline. Plasma LDL-C level was evaluated using an L type LDL-C detection kit (Wako). Functional studies of EDV in response to ACh were performed in aortas precontracted with phentolamine using a myograph (Danish Myo Technology) as previously reported (Tian et al., 2011). Detailed procedures are described in Supplemental Experimental Procedures.
Statistical Analyses
Results are represented as the mean ± SEM unless otherwise indicated. Statistical analysis was performed with one-way or two-way ANOVA implemented in Prism 6 (version 6.03), or with Kruskal-Wallis test implemented in Scipy (version 0.18.1). All statistical tests were evaluated at a significance level of p < 0.05.
Author Contributions
M.A.E. and H.-F.T. conceived the idea for this study and supervised the experiments. M.A.E. and H.-F.T. provided most of the funding. X.Z., B.Q., and X.B. contributed to the funding. J.Y., Y.W., T.Z., L.-Y.W., X.-Y.T., X.H., R.W., Y.L., L.-H.C., G.L., and Z.H. conducted the experiments. W.-H.L., K.-W.A., and A.P.H. provided technical support. W.F., P.Z., X.W., D.P.I., Z.L., Y.L., S.C., D.W, and L. Li provided materials or patient samples. X.Z., L. Lai, B.Q., X.B., Y.H., and C.-W.S. provided relevant advice. M.A.E., J.Y., and H.-F.T. analyzed the data and wrote the manuscript. M.A.E. and H.-F.T. approved the final version.
Acknowledgments
We thank all members of our laboratories for their support. We also thank Jenny C.Y. Ho for her technical contribution to this work, Sigma-Aldrich for donation of LDLR ZFNs, and Sanofi and Regeneron Pharmaceuticals for providing PCSK9 antibodies (alirocumab). This work was supported by the Hong Kong Research Grant Council Theme Based Research Scheme (T12-705/11), The National Key Research and Development Program of China (2016YFA0100102), the Strategic Priority Research Program of the Chinese Academy of Sciences (XDA01020106), the Hong Kong Government Innovation and Technology Support Program (Tier 3) (ITS/303/12), the Cooperation Program of the Research Grants Council (RGC) of the Hong Kong Special Administrative Region and the National Natural Science Foundation of China (N-HKU 730/12 and 81261160506), the National Natural Science Foundation of China (8157050838 and 31370995), the Guangdong Province Science and Technology Program (2013B050800010, 2014A030312001, and 2016B030229007), the Guangzhou Science and Technology Cooperation Program (201508030027), the Shenzhen Science and Technology Council Basic Research program (JCYJ20150331142757383), the Pearl River Science and Technology Nova Program of Guangzhou (201610010107), and the Youth Innovation Promotion Association of the Chinese Academy of Sciences (2015294). H.-F.T. is the National Coordinator and Investigator of the ODYSSEY OUTCOMES study sponsored by Sanofi and Regeneron Pharmaceuticals.
Published: March 2, 2017
Footnotes
Supplemental Information includes Supplemental Experimental Procedures, five figures, and two tables and can be found with this article online at http://dx.doi.org/10.1016/j.stemcr.2017.01.027.
Contributor Information
Miguel A. Esteban, Email: miguel@gibh.ac.cn.
Hung-Fat Tse, Email: hftse@hku.hk.
Supplemental Information
References
- Aggoun Y., Bonnet D., Sidi D., Girardet J.P., Brucker E., Polak M., Safar M.E., Levy B.I. Arterial mechanical changes in children with familial hypercholesterolemia. Arterioscler. Thromb. Vasc. Biol. 2000;20:2070–2075. doi: 10.1161/01.atv.20.9.2070. [DOI] [PubMed] [Google Scholar]
- Azuma H., Paulk N., Ranade A., Dorrell C., Al-Dhalimy M., Ellis E., Strom S., Kay M.A., Finegold M., Grompe M. Robust expansion of human hepatocytes in Fah(-/-)/Rag2(-/-)/Il2rg(-/-) mice. Nat. Biotechnol. 2007;25:903–910. doi: 10.1038/nbt1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basma H., Soto-Gutierrez A., Yannam G.R., Liu L., Ito R., Yamamoto T., Ellis E., Carson S.D., Sato S., Chen Y. Differentiation and transplantation of human embryonic stem cell-derived hepatocytes. Gastroenterology. 2009;136:990–999. doi: 10.1053/j.gastro.2008.10.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benda C., Zhou T., Wang X., Tian W., Grillari J., Tse H.-F., Grillari-Voglauer R., Pei D., Esteban M.A. Urine as a source of stem cells. Adv. Biochem. Eng. Biotechnol. 2013;129:19–32. doi: 10.1007/10_2012_157. [DOI] [PubMed] [Google Scholar]
- Bissig K.D., Wieland S.F., Tran P., Isogawa M., Le T.T., Chisari F.V., Verma I.M. Human liver chimeric mice provide a model for hepatitis B and C virus infection and treatment. J. Clin. Invest. 2010;120:924–930. doi: 10.1172/JCI40094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bissig-Choisat B., Wang L., Legras X., Saha P.K., Chen L., Bell P., Pankowicz F.P., Hill M.C., Barzi M., Leyton C.K. Development and rescue of human familial hypercholesterolaemia in a xenograft mouse model. Nat. Commun. 2015;6:7339. doi: 10.1038/ncomms8339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown M.S., Goldstein J.L. A receptor-mediated pathway for cholesterol homeostasis. Science. 1986;232:34–47. doi: 10.1126/science.3513311. [DOI] [PubMed] [Google Scholar]
- Cannon C.P., Blazing M.A., Giugliano R.P., McCagg A., White J.A., Theroux P., Darius H., Lewis B.S., Ophuis T.O., Jukema J.W. Ezetimibe added to statin therapy after acute coronary syndromes. N. Engl. J. Med. 2015;372:2387–2397. doi: 10.1056/NEJMoa1410489. [DOI] [PubMed] [Google Scholar]
- Cantz T., Sharma A.D., Ott M. Concise review: cell therapies for hereditary metabolic liver diseases-concepts, clinical results, and future developments. Stem Cells. 2015;33:1055–1062. doi: 10.1002/stem.1920. [DOI] [PubMed] [Google Scholar]
- Carpentier A., Tesfaye A., Chu V., Nimgaonkar I., Zhang F., Lee S.B., Thorgeirsson S.S., Feinstone S.M., Liang T.J. Engrafted human stem cell-derived hepatocytes establish an infectious HCV murine model. J. Clin. Invest. 2014;124:4953–4964. doi: 10.1172/JCI75456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cayo M.A., Cai J., DeLaForest A., Noto F.K., Nagaoka M., Clark B.S., Collery R.F., Si-Tayeb K., Duncan S.A. JD induced pluripotent stem cell–derived hepatocytes faithfully recapitulate the pathophysiology of familial hypercholesterolemia. Hepatology. 2012;56:2163–2171. doi: 10.1002/hep.25871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y.F., Tseng C.Y., Wang H.W., Kuo H.C., Yang V.W., Lee O.K. Rapid generation of mature hepatocyte-like cells from human induced pluripotent stem cells by an efficient three-step protocol. Hepatology. 2012;55:1193–1203. doi: 10.1002/hep.24790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen Y., Li Y.F., Wang X., Zhang W., Sauer V., Chang C.J., Han B., Tchaikovskaya T., Avsar Y., Tafaleng E. Amelioration of hyperbilirubinemia in Gunn rats after transplantation of human induced pluripotent stem cell-derived hepatocytes. Stem Cell Rep. 2015;5:22–30. doi: 10.1016/j.stemcr.2015.04.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dormuth C.R., Filion K.B., Paterson J.M., James M.T., Teare G.F., Raymond C.B., Rahme E., Tamim H., Lipscombe L. Higher potency statins and the risk of new diabetes: multicentre, observational study of administrative databases. BMJ. 2014;348:g3244. doi: 10.1136/bmj.g3244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubuc G., Chamberland A., Wassef H., Davignon J., Seidah N.G., Bernier L., Prat A. Statins upregulate PCSK9, the gene encoding the proprotein convertase neural apoptosis-regulated convertase-1 implicated in familial hypercholesterolemia. Arterioscler. Thromb. Vasc. Biol. 2004;24:1454–1459. doi: 10.1161/01.ATV.0000134621.14315.43. [DOI] [PubMed] [Google Scholar]
- Endo A. The discovery and development of HMG-CoA reductase inhibitors. J. Lipid. Res. 1992;33:1569–1582. [PubMed] [Google Scholar]
- Grskovic M., Javaherian A., Strulovici B., Daley G.Q. Induced pluripotent stem cells—opportunities for disease modelling and drug discovery. Nat. Rev. Drug Discov. 2011;10:915–929. doi: 10.1038/nrd3577. [DOI] [PubMed] [Google Scholar]
- Ishibashi S., Brown M.S., Goldstein J.L., Gerard R.D., Hammer R.E., Herz J. Hypercholesterolemia in low density lipoprotein receptor knockout mice and its reversal by adenovirus-mediated gene delivery. J. Clin. Invest. 1993;92:883–893. doi: 10.1172/JCI116663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kajiwara M., Aoi T., Okita K., Takahashi R., Inoue H., Takayama N., Endo H., Eto K., Toguchida J., Uemoto S. Donor-dependent variations in hepatic differentiation from human-induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA. 2012;109:12538–12543. doi: 10.1073/pnas.1209979109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khoo K.L., van Acker P., Defesche J.C., Tan H., van de Kerkhof L., Heijnen-van Eijk S.J., Kastelein J.J., Deslypere J.P. Low-density lipoprotein receptor gene mutations in a Southeast Asian population with familial hypercholesterolemia. Clin. Genet. 2000;58:98–105. doi: 10.1034/j.1399-0004.2000.580202.x. [DOI] [PubMed] [Google Scholar]
- Legrand N., Ploss A., Balling R., Becker P.D., Borsotti C., Brezillon N., Debarry J., de Jong Y., Deng H.K., Di Santo J.P. Humanized mice for modeling human infectious disease: challenges, progress, and outlook. Cell Host Microbe. 2009;6:5–9. doi: 10.1016/j.chom.2009.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li W., Wang X., Fan W., Zhao P., Chan Y.C., Chen S., Zhang S., Guo X., Zhang Y., Li Y. Modeling abnormal early development with induced pluripotent stem cells from aneuploid syndromes. Hum. Mol. Genet. 2012;21:32–45. doi: 10.1093/hmg/ddr435. [DOI] [PubMed] [Google Scholar]
- Liu H., Kim Y., Sharkis S., Marchionni L., Jang Y.Y. In vivo liver regeneration potential of human induced pluripotent stem cells from diverse origins. Sci. Transl. Med. 2011;3:82ra39. doi: 10.1126/scitranslmed.3002376. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maxwell K.N., Fisher E.A., Breslow J.L. Overexpression of PCSK9 accelerates the degradation of the LDLR in a post-endoplasmic reticulum compartment. Proc. Natl. Acad. Sci. USA. 2005;102:2069–2074. doi: 10.1073/pnas.0409736102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moehle E.A., Rock J.M., Lee Y.-L., Jouvenot Y., DeKelver R.C., Gregory P.D., Urnov F.D., Holmes M.C. Targeted gene addition into a specified location in the human genome using designed zinc finger nucleases. Proc. Natl. Acad. Sci. USA. 2007;104:3055–3060. doi: 10.1073/pnas.0611478104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramakrishnan V.M., Yang J.Y., Tien K.T., McKinley T.R., Bocard B.R., Maijub J.G., Burchell P.O., Williams S.K., Morris M.E., Hoying J.B. Restoration of physiologically responsive low-density lipoprotein receptor-mediated endocytosis in genetically deficient induced pluripotent stem cells. Sci. Rep. 2015;5:13231. doi: 10.1038/srep13231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rashid S.T., Corbineau S., Hannan N., Marciniak S.J., Miranda E., Alexander G., Huang-Doran I., Griffin J., Ahrlund-Richter L., Skepper J. Modeling inherited metabolic disorders of the liver using human induced pluripotent stem cells. J. Clin. Invest. 2010;120:3127–3136. doi: 10.1172/JCI43122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reiner Z. Management of patients with familial hypercholesterolaemia. Nat. Rev. Cardiol. 2015;12:565–575. doi: 10.1038/nrcardio.2015.92. [DOI] [PubMed] [Google Scholar]
- Robinson J.G., Farnier M., Krempf M., Bergeron J., Luc G., Averna M., Stroes E.S., Langslet G., Raal F.J., El Shahawy M. Efficacy and safety of alirocumab in reducing lipids and cardiovascular events. N. Engl. J. Med. 2015;372:1489–1499. doi: 10.1056/NEJMoa1501031. [DOI] [PubMed] [Google Scholar]
- Roth E.M., McKenney J.M., Hanotin C., Asset G., Stein E.A. Atorvastatin with or without an antibody to PCSK9 in primary hypercholesterolemia. N. Engl. J. Med. 2012;367:1891–1900. doi: 10.1056/NEJMoa1201832. [DOI] [PubMed] [Google Scholar]
- Shiomi M., Ito T. The Watanabe heritable hyperlipidemic (WHHL) rabbit, its characteristics and history of development: a tribute to the late Dr. Yoshio Watanabe. Atherosclerosis. 2009;207:1–7. doi: 10.1016/j.atherosclerosis.2009.03.024. [DOI] [PubMed] [Google Scholar]
- Si-Tayeb K., Idriss S., Champon B., Caillaud A., Pichelin M., Arnaud L., Lemarchand P., Le May C., Zibara K., Cariou B. Urine-sample-derived human induced pluripotent stem cells as a model to study PCSK9-mediated autosomal dominant hypercholesterolemia. Dis. Model. Mech. 2016;9:81–90. doi: 10.1242/dmm.022277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sniderman A.D., Tsimikas S., Fazio S. The severe hypercholesterolemia phenotype: clinical diagnosis, management, and emerging therapies. J. Am. Coll. Cardiol. 2014;63:1935–1947. doi: 10.1016/j.jacc.2014.01.060. [DOI] [PubMed] [Google Scholar]
- Soldner F., Laganière J., Cheng A.W., Hockemeyer D., Gao Q., Alagappan R., Khurana V., Golbe L.I., Myers R.H., Lindquist S. Generation of isogenic pluripotent stem cells differing exclusively at two early onset Parkinson point mutations. Cell. 2011;146:318–331. doi: 10.1016/j.cell.2011.06.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stein E.A., Honarpour N., Wasserman S.M., Xu F., Scott R., Raal F.J. Effect of the PCSK9 monoclonal antibody, AMG 145, in homozygous familial hypercholesterolemia. Circulation. 2013;128:2113–2120. doi: 10.1161/CIRCULATIONAHA.113.004678. [DOI] [PubMed] [Google Scholar]
- Stroes E.S., Thompson P.D., Corsini A., Vladutiu G.D., Raal F.J., Ray K.K., Roden M., Stein E., Tokgozoglu L., Nordestgaard B.G. Statin-associated muscle symptoms: impact on statin therapy—European Atherosclerosis Society consensus panel statement on assessment, aetiology and management. Eur. Heart J. 2015;36:1012–1022. doi: 10.1093/eurheartj/ehv043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takahashi K., Tanabe K., Ohnuki M., Narita M., Ichisaka T., Tomoda K., Yamanaka S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. doi: 10.1016/j.cell.2007.11.019. [DOI] [PubMed] [Google Scholar]
- Tian X.Y., Wong W.T., Xu A., Chen Z.Y., Lu Y., Liu L.M., Lee V.W., Lau C.W., Yao X., Huang Y. Rosuvastatin improves endothelial function in db/db mice: role of angiotensin II type 1 receptors and oxidative stress. Br. J. Pharmacol. 2011;164:598–606. doi: 10.1111/j.1476-5381.2011.01416.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanhoutte P.M., Zhao Y., Xu A., Leung S.W. Thirty years of saying NO: sources, fate, actions, and misfortunes of the endothelium-derived vasodilator mediator. Circ. Res. 2016;119:375–396. doi: 10.1161/CIRCRESAHA.116.306531. [DOI] [PubMed] [Google Scholar]
- Wiegman A., Hutten B.A., de Groot E., Rodenburg J., Bakker H.D., Buller H.R., Sijbrands E.J., Kastelein J.J. Efficacy and safety of statin therapy in children with familial hypercholesterolemia: a randomized controlled trial. JAMA. 2004;292:331–337. doi: 10.1001/jama.292.3.331. [DOI] [PubMed] [Google Scholar]
- Yu J., Hu K., Smuga-Otto K., Tian S., Stewart R., Slukvin I.I., Thomson J.A. Human induced pluripotent stem cells free of vector and transgene sequences. Science. 2009;324:797–801. doi: 10.1126/science.1172482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu J., Chau K.F., Vodyanik M.A., Jiang J., Jiang Y. Efficient feeder-free episomal reprogramming with small molecules. PLoS One. 2011;6:e17557. doi: 10.1371/journal.pone.0017557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yusa K., Rashid S.T., Strick-Marchand H., Varela I., Liu P.Q., Paschon D.E., Miranda E., Ordonez A., Hannan N.R., Rouhani F.J. Targeted gene correction of alpha1-antitrypsin deficiency in induced pluripotent stem cells. Nature. 2011;478:391–394. doi: 10.1038/nature10424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang S., Chen S., Li W., Guo X., Zhao P., Xu J., Chen Y., Pan Q., Liu X., Zychlinski D. Rescue of ATP7B function in hepatocyte-like cells from Wilson's disease induced pluripotent stem cells using gene therapy or the chaperone drug curcumin. Hum. Mol. Genet. 2011;20:3176–3187. doi: 10.1093/hmg/ddr223. [DOI] [PubMed] [Google Scholar]
- Zhang L.W., McCabe T., Condra J.H., Ni Y.G., Peterson L.B., Wang W.R., Strack A.M., Wang F.B., Pandit S., Hammond H. An anti-PCSK9 antibody reduces LDL-cholesterol on top of a statin and suppresses hepatocyte SREBP-regulated genes. Int. J. Biol. Sci. 2012;8:310–327. doi: 10.7150/ijbs.3524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou T., Benda C., Duzinger S., Huang Y., Li X., Li Y., Guo X., Cao G., Chen S., Hao L. Generation of induced pluripotent stem cells from urine. J. Am. Soc. Nephrol. 2011;22:1221–1228. doi: 10.1681/ASN.2011010106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou T., Benda C., Dunzinger S., Huang Y., Ho J.C., Yang J., Wang Y., Zhang Y., Zhuang Q., Li Y. Generation of human induced pluripotent stem cells from urine samples. Nat. Protoc. 2012;7:2080–2089. doi: 10.1038/nprot.2012.115. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.