

Abstract
The aim of the present study was to investigate the key genes associated with traumatic spinal cord injuries (TSCI). The dataset GSE52763 was downloaded from the Gene Expression Omnibus, for which lumbar spinal cord samples were obtained from rats at 1 and 3 weeks following contusive spinal cord injury and 1 week subsequent to a sham laminectomy, and used to identify differentially expressed genes (DEGs). Functional enrichment analysis, co-expression analysis and transcription factor (TF) identification were performed for DEGs common to the 1 and 3 week injury samples. In total, 234 upregulated and 51 downregulated DEGs were common to the 1 and 3 week injury samples. The upregulated DEGs were significantly enriched in Gene Ontology terms concerning immunity (e.g. Itgal and Ccl2) and certain pathways, including natural killer cell mediated cytotoxicity [e.g. Ras-related C3 botulinum toxin substrate 2 (Rac2) and TYRO protein tyrosine kinase binding protein (Tyrobp)]. The downregulated DEGs were highly enriched in female gonad development [e.g. progesterone receptor (Pgr)], and the steroid biosynthesis pathway. A total of 139 genes had co-expression associations and the majority of them were upregulated genes. The upregulated co-expressed genes were predominantly enriched in biological regulation, including TGFB induced factor homeobox 1 (Tgif1) and Rac2. The downregulated co-expressed genes were enriched in anatomical structure development (e.g. Dnm3). A total of 92 co-expressed genes composed the protein-protein interaction network. Additionally, 9 TFs (e.g. Pgr and Tgif1) were identified from the DEGs. It was hypothesized that the genes including Tgif1, Rac2, Tyrobp, and Pgr may be closely associated with TSCI.
Keywords: co-expression, differentially expressed gene, network, traumatic spinal cord injury, transcription factor
Introduction
Patients with traumatic spinal cord injuries (TSCI) endure low health-associated quality of life and high healthcare costs. They also have a higher mortality rate compared with the general population. In 2012, the estimated incidence of acute spinal cord injury in the United States was 54 cases per 1 million (1). The biological processes of TSCI involve a diverse group of cells and molecules from the nervous, immune and vascular systems. For instance, connexin 43 functions as a mediator of central nervous system inflammation and chronic pain following spinal cord injury (2), altered expression of E2F-associated phosphoprotein regulates reactive astrogliosis and neuronal apoptosis (3) and ginsenoside Rb1 upregulates the expression of Bcl-xL and vascular endothelial growth factor at 7 days after spinal cord injury (4). Investigation of gene changes has contributed to the understanding of the molecular mechanisms of TSCI.
Gene expression profiling by microarray has been used to uncover molecular variations in spinal cord repair and degeneration (5–7). In 2014, using microarray analysis, Shin et al (8) identified that numerous inflammation-associated genes were upregulated in the lumbar spinal cord at 1 and 3 weeks after traumatic injury, and locomotor function was improved in part by treadmill locomotor training (TMT). However, the molecular mechanisms of TSCI remain to be elucidated and regulatory factors associated with TSCI, including transcription factors (TFs), have not been investigated to the best of the authors' knowledge.
The present study used the microarray data obtained by Shin et al (8) and screened differentially expressed genes (DEGs) common to the 1 and 3 week injury samples, and then analyzed the functions and interactions of DEGs. Additionally, TFs in DEGs were identified to reveal the regulatory associations of DEGs. These results may provide novel information to aid the understanding of the molecular mechanisms of TSCI.
Materials and methods
Affymetrix microarray data
The raw gene expression profile data GSE52763 (8) were obtained from the public database Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/), which is based on the platform of GPL1355 (Rat230_2) Affymetrix Rat Genome 230 2.0 Array. The dataset contained eight rat lumbar spinal cord samples obtained from rats 1 (n=4) and 3 (n=4) weeks following contusive spinal cord injury at the T9 level (designated as 1 week injury and 3 weeks injury samples), three lumbar spinal cord samples obtained from rats 1 week following sham laminectomy (designated as sham samples), four lumbar spinal cord samples obtained at 3 weeks following contusive spinal cord injury with treadmill training (designated as 3 weeks injury + TMT samples), and three lumbar spinal cord samples from rats which underwent a sham laminectomy followed by 2 weeks of treadmill training (designated as sham + TMT samples). All of the samples were taken from adult (8 weeks) female Sprague-Dawley rats (250–300 g). Only 1 week injury, 3 weeks injury and sham samples were used for analysis in the present study.
CEL files were downloaded and the gene expression data of all samples were preprocessed through background correction, quantile normalization and probe summarization using the Robust Microarray Analysis algorithm of the affy package of Bioconductor (http://www.bioconductor.org/packages/release/bioc/html/) (9).
DEGs screening
The linear models for the microarray data package of Bioconductor (10) was used to identify DEGs between 1 week and 3 weeks injury samples and sham samples. The P-value for each gene was calculated by t-test and only the genes with P-value <0.05 and fold change ≥1.5 were selected as DEGs. Subsequently, the DEGs common to the 1 and 3 week injury samples were screened for subsequent analyses.
Enrichment analysis
Gene Ontology (GO) functional and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses for DEGs were conducted using the Database for Annotation, Visualization and Integrated Discovery (http://david.abcc.ncifcrf.gov) database, which provides a set of functional annotation tools to aid investigators in comprehending the biological importance underlying numerous genes (11). P<0.05 and gene count ≥2 were set as the cut-off criteria.
Co-expression analysis
The Pearson correlation coefficient was calculated to analyze the co-expression associations between DEGs (11). The co-expression pairs with a Pearson correlation coefficient >0.9 were screened out, and the co-expression network was visualized using Cytoscape [http://cytoscape.org; (12)].
Subsequently, GO functional enrichment analysis in biological process was performed using the plug-in Bingo (13) in Cytoscape. P<0.05 was set as the cut-off criterion. Additionally, the Search Tool for the Retrieval of Interacting Genes/Proteins (http://string-db.org) database was used to analyze the protein-protein interactions (PPIs) of co-expressed genes, and the PPI network was visualized by Cytoscape.
Identification of TFs from DEGs
TFs in the DEGs common to the 1 and 3 week injury samples were identified using the Animal Transcription Factor Database [AnimalTFDB; http://www.bioguo.org/AnimalTFDB/species_index.php; (14)].
Results
Identification of DEGs
Based on the normalization of the microarray data, the boxplot of preprocessed data displayed that medians of each sample data were almost on a line, indicating that the data after preprocessing met the standard for further analyses (Fig. 1).
Figure 1.
The boxplots for microarray data prior and subsequent to normalization. The abscissa displays the samples and the ordinate represents gene expression. ‘1 week sham’ represents lumbar spinal cord samples obtained from rats 1 week following a sham laminectomy, three repeats; ‘1 week injury’ and ‘3 week injury’ represents lumbar spinal cord samples obtained from rats 1 and 3 weeks following contusive spinal cord injury at the T9 level (four repeats for each treatment).
In total, 322 upregulated and 78 downregulated DEGs were screened between 1 week injury and sham samples, in addition to 354 upregulated and 285 downregulated ones between 3 week injury and sham samples. Among them, 234 upregulated and 51 downregulated DEGs were common to the 1 and 3 week injury samples. The hierarchical cluster analysis of the data demonstrated that the DEGs can be used to accurately distinguish 1 and 3 week injury samples from sham samples (Fig. 2).
Figure 2.

The cluster heat maps for the differentially expressed genes between the sham samples and the 1 and 3 week injury samples. Each row represents a single gene and each column represents a spinal cord sample. The gradual color change from red to green represents the changing process from upregulation to downregulation. ‘1 week sham’ represents lumbar spinal cord samples obtained from rats 1 week following a sham laminectomy, three repeats; ‘1 week injury’ and ‘3 week injury’ represents lumbar spinal cord samples obtained from rats 1 and 3 weeks following contusive spinal cord injury at the T9 level (four repeats for each treatment).
GO functional and KEGG pathway enrichment analyses
To identify the functions of DEGs common to the 1 and 3 week injury samples, GO functional and KEGG pathway enrichment analyses were performed. According to GO functional enrichment analysis, the upregulated DEGs were significantly enriched in several GO terms concerning immunity, including immune response [e.g. integrin subunit α L (Itgal), similar to guanylate binding protein family, member 6 and C-C motif chemokine ligand 2 (Ccl2)], defence response (e.g. TNF α induced protein 8 like 2, apolipoprotein B MRNA editing enzyme catalytic subunit 1 and Ccl2) and cell activation (e.g. exonuclease 1, Intercellular Adhesion Molecule 1 and pleckstrin; Table I). The downregulated DEGs were highly enriched in female gonad development [e.g. Pgr, vascular endothelial growth factor A (Vegfa) and BCL2 like 1], neuron projection [e.g. ATPase plasma membrane Ca2+ transporting 2, dynamin 3 (Dnm3) and glutamate metabotropic receptor 7 (Grm7)] and gated channel activity [e.g. γ-aminobutyric acid type A receptor α3 subunit (Gabra3), Grm7 and calcium voltage-gated channel auxiliary subunit β4; Table II].
Table I.
Top five enriched GO terms with the highest P-value in biological process, cellular component and molecular function for the up-regulated differentially expressed genes common to the 1 week and 3 weeks injury samples.
| Category | Term | Count | P-value | Genes |
|---|---|---|---|---|
| BP | GO:0006955-immune response | 37 | 3.71E-22 | Tnfaip8L2, Itgal, Loc685067, Ccl2, Apobec1, C3, Endou, Tlr2, Rsad2, Tlr7, C1Qc, Btk, Gch1, Klhl6, Fcgr1A, Cfh, Fcer1G, Inpp5D, Fcgr3A, Exo1, Ptpn6, Icam1, Ptprc, Tnfsf4, Rt1-Ce12, Myo1F, Vav1, Psmb8, Psmb9, C1Qa, C1Qb, Cd86, Cybb, Fcgr2B, Cd300A, Cxcl16, Gbp2 |
| GO:0006952-defense response | 29 | 8.07E-15 | Tnfaip8L2, Apobec1, Ccl2, C3, Tlr2, Rsad2, Itgb2, Tlr7, C1Qc, Btk, Gch1, Casp4, Hmox1, Cybb, Fcgr1A, Cfh, Pycard, Fcer1G, Ptpn6, Ptprc, Tnfsf4, Pdpn, Hck, Myo1F, C1Qa, C1Qb, Cd86, Fcgr2B, Cxcl16 | |
| GO:0002252-immune effector process | 18 | 3.17E-14 | Exo1, Ptprc, Ptpn6, Icam1, Apobec1, C3, Myo1F, Rsad2, Tlr7, C1Qc, Btk, C1Qa, C1Qb, Fcgr2B, Fcgr1A, Cfh, Fcer1G, Inpp5D | |
| GO:0001775-cell activation | 21 | 4.15E-12 | Exo1, Icam1, Ptprc, Itgal, Tnfsf4, Plek, Aif1, Tlr2, Myo1F, Itgb2, Tlr7, Vav1, Itgam, Timp1, Btk, Cd48, Cd86, Fcgr2B, Fcer1G, Fcgr3A, Blnk | |
| GO:0045321-leukocyte activation | 19 | 3.69E-11 | Exo1, Icam1, Ptprc, Itgal, Tnfsf4, Aif1, Tlr2, Myo1F, Itgb2, Tlr7, Vav1, Itgam, Btk, Cd48, Cd86, Fcgr2B, Fcer1G, Fcgr3A, Blnk | |
| CC | GO:0009897-external side of plasma membrane | 13 | 5.95E-07 | Cd48, Icam1, Itgal, Ptprc, Cd86, Emr1, Fcgr2B, Fcgr1A, Tlr2, Fcer1G, Cd22, Clec7A, Itgam |
| GO:0009986-cell surface | 16 | 1.03E-05 | Icam1, Ptprc, Itgal, Tnfsf4, Tlr2, Itgb2, Cd53, Itgam, Cd48, Cd86, Fcgr2B, Emr1, Fcgr1A, Cd22, Fcer1G, Clec7A | |
| GO:0044459-plasma membrane part | 33 | 7.58E-05 | Itgal, Gna15, Aif1, Tlr2, Rsad2, Itgb2, Abca1, Itgam, Cd48, P2Ry6, Laptm5, Fcgr1A, Hmox1, Cd22, Fcer1G, Gpnmb, Ptpn6, Icam1, Ptprc, Plek, Rt1-Ce12, Pdpn, Anxa1, Axl, Stom, Cd86, Cybb, Gngt2, Emr1, Fcgr2B, Clec7A, Cp, Gbp2 | |
| GO:0005886-plasma membrane | 47 | 2.18E-04 | Itgal, Gna15, Aif1, Ifitm3, Tlr2, Rsad2, Itgb2, Abca1, Itgam, Slc7A7, Cd48, P2Ry6, Laptm5, Mall, Fcgr1A, Hmox1, Cfh, Fcer1G, Cd22, Pik3Ap1, Inpp5D, Fcgr3A, Gpnmb, Csf1R, Blnk, Ptprc, Ptpn6, Icam1, F10, Plek, Rt1-Ce12, Pdpn, Anxa1, Axl, Clic1, Stom, Cd86, Cybb, Gngt2, Gpr34, Fcgr2B, Emr1, Cd300A, Clec7A, Pcyox1, Cp, Gbp2 | |
| GO:0005576-extracellular region | 30 | 3.59E-04 | Fmod, Ccl2, Spock3, C3, Tnc, C1Qc, Timp1, Lgals3Bp, Glipr1, Fcn1, Cfh, Pycard, Ptn, Casp1, Tfpi2, Matn2, Icam1, Ctsz, F10, Tnfsf4, Lgals3, Plek, Anxa1, C1Qa, C1Qb, Grn, Cxcl16, Cp, Pcyox1, Pros1 | |
| MF | GO:0030246-carbohydrate binding | 19 | 8.31E-08 | Ptprc, Ccl2, Lgals3, Spock3, Endou, Hexb, Tlr2, Itgam, Tnfaip6, Pygl, Fcn1, Cfh, Grifin, Clec4A1, Ptn, Clec4A3, Clec7A, Gpnmb, Cd302 |
| GO:0030247-polysaccharide binding | 11 | 1.06E-06 | Tnfaip6, Ptprc, Ccl2, Spock3, Endou, Cfh, Tlr2, Ptn, Clec7A, Gpnmb, Itgam | |
| GO:0001871-pattern binding | 11 | 1.06E-06 | Tnfaip6, Ptprc, Ccl2, Spock3, Endou, Cfh, Tlr2, Ptn, Clec7A, Gpnmb, Itgam | |
| GO:0019865-immunoglobulin binding | 5 | 8.51E-06 | Lgals3, Fcgr2B, Fcgr1A, Fcer1G, Fcgr3A | |
| GO:0005539-glycosaminoglycan binding | 9 | 2.63E-05 | Tnfaip6, Ptprc, Ccl2, Spock3, Cfh, Tlr2, Ptn, Gpnmb, Itgam |
GO, Gene Ontology; BP, biological process; CC, cellular component; MF, molecular function; Tnfaip8L2, TNF α induced protein 8 like 2; Itgal, Integrin subunit αL; Loc685067, similar to guanylate binding protein family, member 6; Ccl2, C-C motif chemokine ligand 2; Psmb8, proteasome subunit β8; C1Qa, complement C1q A chain; C1Qb, complement C1q B chain; Cd86, CD86 molecule; Cybb, cytochrome B-245 b chain Fcgr2B, fc fragment of IgG receptor IIb; Apobec1, apolipoprotein B MRNA editing enzyme catalytic subunit 1; C3, complement C3; Tlr2, toll like receptor 2; Rsad2, radical s-adenosyl methionine domain containing 2; Itgb2, integrin subunit b2; Tlr7, toll like receptor 7; C1Qc, complement C1q C chain; Btk, bruton tyrosine kinase; Exo1, exonuclease 1; Ptprc, protein tyrosine phosphatase, receptor type C; Ptpn6, protein tyrosine phosphatase, non-receptor type 6; Icam1, Intercellular Adhesion Molecule 1; Myo1F, myosin IF; Vav1, vav guanine nucleotide exchange factor 1; Itgam, integrin subunit α M; Timp1, TIMP metallopeptidase inhibitor 1; Cd48, CD48 molecule; Emr1, EGF-Like Module Receptor 1; Fcgr1A, fc fragment of IgG receptor Ia; Tlr2, toll like receptor 2; Fcer1G, Fc fragment of IgE receptor Ig; Clec7A, C-type lectin domain family 7 member A; Gna15, G protein subunit α15; Abca1, ATP binding cassette subfamily A member 1; Gpnmb, glycoprotein Nmb; Plek, pleckstrin; Slc7A7, solute carrier family 7 member 7; P2Ry6, pyrimidinergic receptor P2Y6; Laptm5, lysosomal protein transmembrane 5; Fcn1, ficolin 1; F10, coagulation factor X; Tnfsf4, tumor necrosis factor superfamily member 4; Lgals3, galectin 3; Pcyox1, prenylcysteine oxidase 1; Pygl, phosphorylase, glycogen, liver; Cfh, complement factor H; Grifin, galectin-related inter-fiber protein; Clec4A1, C-type lectin domain family 4 member A1; Clec4A3, C-type lectin domain family 4 member A3; Cd302, CD302 molecule; Tnfaip6, TNF α induced protein 6; Spock3, SPARC/osteonectin, cwcv and kazal like domains proteoglycan 3; Endou, endonuclease, poly(U) specific; Ptn, pleiotrophin; Fcgr3A, Fc fragment of IgG receptor IIIa.
Table II.
Top five enriched GO terms with the highest P-value in biological process, cellular component and molecular function for the downregulated differentially expressed genes common to the 1 week injury and 3 week injury samples.
| Category | Term | Count | P-value | Genes |
|---|---|---|---|---|
| BP | GO:0008585-female gonad development | 5 | 7.32E-05 | Pgr, Vegfa, Bcl2L1, Pcyt1B, Vgf |
| GO:0046545-development of primary female sexual characteristics | 5 | 9.52E-05 | Pgr, Vegfa, Bcl2L1, Pcyt1B, Vgf | |
| GO:0022602-ovulation cycle process | 5 | 1.05E-04 | Pgr, Vegfa, Bcl2L1, Pcyt1B, Vgf | |
| GO:0046660-female sex differentiation | 5 | 1.28E-04 | Pgr, Vegfa, Bcl2L1, Pcyt1B, Vgf | |
| GO:0042698-ovulation cycle | 5 | 1.46E-04 | Pgr, Vegfa, Bcl2L1, Pcyt1B, Vgf | |
| CC | GO:0043005-neuron projection | 8 | 6.07E-04 | Pgr, Atp2B2, Dnm3, Grm7, Dicer1, Slc18A3, Vgf, Pex5L |
| GO:0045202-synapse | 7 | 1.68E-03 | Dnm3, Clstn2, Gabra4, Gabra3, Grm7, Rps6Kb1, Rph3A | |
| GO:0044456-synapse part | 6 | 1.80E-03 | Dnm3, Clstn2, Gabra4, Gabra3, Grm7, Rph3A | |
| GO:0030425-dendrite | 5 | 8.17E-03 | Atp2B2, Dnm3, Grm7, Dicer1, Pex5L | |
| GO:0005886-plasma membrane | 16 | 9.36E-03 | Cadm3, Clstn2, Gabra4, Gabra3, Trhr, Rps6Kb1, Bcl2L1, Cacnb4, Rph3A, Slc9A3R2, P2Rx5, Atp2B2, Mast1, Grm7, Rasgrp1, Slc6A5 | |
| MF | GO:0022836-gated channel activity | 5 | 1.32E-02 | P2Rx5, Gabra4, Gabra3, Grm7, Cacnb4 |
| GO:0005230-extracellular ligand-gated ion channel activity | 3 | 2.14E-02 | P2Rx5, Gabra4, Gabra3 | |
| GO:0005216-ion channel activity | 5 | 2.74E-02 | P2Rx5, Gabra4, Gabra3, Grm7, Cacnb4 | |
| GO:0022838-substrate specific channel activity | 5 | 3.00E-02 | P2Rx5, Gabra4, Gabra3, Grm7, Cacnb4 | |
| GO:0046983-protein dimerization activity | 6 | 3.33E-02 | Zbtb7B, Cadm3, Grm7, Trhr, Vegfa, Bcl2L1 |
GO, Gene Ontology; BP, biological process; CC, cellular component; MF, molecular function; Pgr, progesterone receptor; Vegfa, vascular endothelial growth factor A; Bcl2L1, BCL2 like 1; Pcyt1B, phosphate cytidylyltransferase 1, choline, β; Vgf, VGF nerve growth factor inducible; Atp2B2, ATPase plasma membrane Ca2+ transporting 2; Dnm3, dynamin 3; Grm7, glutamate metabotropic receptor 7; Dicer1, Dicer 1, Ribonuclease III; Slc18A3, solute carrier family 18 member A3; Pex5L, peroxisomal biogenesis factor 5 like; Clstn2, calsyntenin 2; Gabra4, γ-aminobutyric acid type A receptor α4 subunit; Gabra3, γ-aminobutyric acid type A receptor α3 subunit; Rps6Kb1, ribosomal protein S6 kinase B1; Rph3A, rabphilin 3A; Cadm3, cell adhesion molecule 3; Trhr, thyrotropin releasing hormone receptor; Cacnb4, calcium voltage-gated channel auxiliary subunit β 4; Slc9A3R2, SLC9A3 regulator 2; P2Rx5, purinergic receptor P2X 5; Zbtb7b, zinc finger and BTB domain containing 7B.
Meanwhile, a set of upregulated DEGs were markedly enriched in certain pathways, including natural killer cell mediated cytotoxicity [e.g. Ras-related C3 botulinum toxin substrate 2 (Rac2) and TYRO protein tyrosine kinase binding protein (Tyrobp)] and the B cell receptor signaling pathway [e.g. fc fragment of IgG receptor IIb (Fcgr2B) and Rac2). Several downregulated DEGs were significantly enriched in the pathways of steroid biosynthesis (cytochrome P450 family 51, transmembrane 7 superfamily member 2 and lanosterol synthase) and neuroactive ligand-receptor interaction (e.g. Gabra3 and Grm7; Table III).
Table III.
Top five enriched pathways for the upregulated differentially expressed genes and two enriched pathways for the downregulated differentially expressed genes common to the 1 week injury and 3 week injury samples.
| Category | Term | Count | P-value | Genes |
|---|---|---|---|---|
| Upregulated | rno04650:Natural killer cell mediated cytotoxicity | 11 | 9.78E-07 | Cd48, Itgal, Ptpn6, Icam1, Fcgr2B, Rac2, Fcer1G, Itgb2, Fcgr3A, Vav1, Tyrobp |
| rno04662:B cell receptor signaling pathway | 9 | 7.93E-06 | Ptpn6, Fcgr2B, Rac2, Cd22, Pik3Ap1, Inpp5D, Vav1, Btk, Blnk | |
| rno04666:Fc γ R-mediated phagocytosis | 8 | 2.01E-04 | Arpc1B, Ptprc, Fcgr2B, Rac2, Hck, Fcgr1A, Inpp5D, Vav1 | |
| rno05322:Systemic lupus erythematosus | 8 | 2.32E-04 | C1Qa, C1Qb, Cd86, Fcgr2B, C3, Fcgr1A, Fcgr3A, C1Qc | |
| rno04610:Complement and coagulation cascades | 7 | 3.92E-04 | C1Qa, C1Qb, F10, C3, Cfh, Pros1, C1Qc | |
| Downregulated | rno00100:Steroid biosynthesis | 3 | 1.94E-03 | Cyp51, Tm7Sf2, Lss |
| rno04080:Neuroactive ligand-receptor interaction | 5 | 1.77E-02 | P2Rx5, Gabra4, Gabra3, Grm7, Trhr |
Cd48, CD48 molecule; Itgal, Integrin subunit α L; Icam1, Intercellular Adhesion Molecule 1; Fcgr2B, fc fragment of IgG receptor IIb; Rac2, Ras-related C3 botulinum toxin substrate 2; Fcer1G, Fc fragment of IgE receptor Ig; Itgb2, integrin subunit β2; Fcgr3A, Fc fragment of IgG receptor IIIa; Vav1, vav guanine nucleotide exchange factor 1; Tyrobp, TYRO protein tyrosine kinase binding protein; Ptpn6, protein tyrosine phosphatase, non-receptor type 6; Cd22, CD22 molecule; Pik3Ap1, phosphoinositide-3-kinase adaptor protein 1; Inpp5D, inositol polyphosphate-5-phosphatase D; Btk, bruton tyrosine kinase; Blnk, B-cell linker; Arpc1B, actin related protein 2/3 complex subunit 1B; Ptprc, protein tyrosine phosphatase, receptor type C; Hck, HCK proto-oncogene, src family tyrosine kinase; Fcgr1A, fc fragment of IgG receptor Ia; Inpp5D, inositol polyphosphate-5-phosphatase D; C1Qa, complement C1q A chain; C1Qb, complement C1q B chain; Cd86, CD86 molecule; C3, complement C3; C1Qc, complement C1q C chain; F10, coagulation factor X; Cfh, complement factor H; Pros1, Protein S (α); Cyp51, cytochrome P450 family 51; Tm7Sf2, transmembrane 7 superfamily member 2; Lss, lanosterol synthase; P2Rx5, purinergic receptor P2X 5; Gabra4, gamma-aminobutyric acid type A receptor a4 subunit; Gabra3, gamma-aminobutyric acid type A receptor a3 subunit; Grm7, glutamate metabotropic receptor 7; Trhr, thyrotropin releasing hormone receptor.
Analysis of co-expressed genes
Gene co-expression analysis is a powerful method to predict the function of genes and/or to identify genes that are functionally associated with query genes. Based on the cut-off criterion of Pearson correlation coefficient >0.9, 1894 co-expression pairs in 139 DEGs were obtained (Fig. 3). Notably, the majority of co-expressed genes were upregulated DEGs.
Figure 3.

The co-expression network of the differentially expressed genes common to the 1 week and 3 week injury samples. The red nodes represent upregulated genes; and the green nodes represent downregulated genes. A line between two nodes indicates that there is a co-expression association between these two nodes.
According to GO functional enrichment analysis, the upregulated co-expressed genes [e.g. Rac2, fc fragment of IgG receptor Ia and including TGFB induced factor homeobox 1 (Tgif1)] were significantly enriched in a series of GO terms, including biological regulation and response to stimulus. The downregulated co-expressed genes were markedly enriched in certain GO terms, including anatomical structure development (e.g. Dnm3 and Vegfa) and intracellular signal transduction (e.g. mitogen-activated protein kinase kinase kinase kinase 5 and Grm7) (Table IV).
Table IV.
Top five enriched GO terms with the highest P-value in biological process for the co-expressed up- and downregulated genes.
| Category | Term | Count | P-value | Genes |
|---|---|---|---|---|
| Co-expressed upregulated genes | GO:0065007-biological regulation | 44 | 2.39E-02 | S100A4, Tspo, Spock3, C3, Aif1, Rbm3, Unc93B1, Rsad2, Itgb2, Cd53, C1Qc, Tiprl, Timp1, Cd48, P2Ry6, Rac2, Fcgr1A, Hmox1, Pycard, Cfh, Ptn, Fcer1G, Inpp5D, Csf1R, Ptprc, Pigz, Plek, Pdpn, Bcl2A1, Anxa1, Pkib, Il6R, Anxa3, Psmb8, Arhgap25, Psmb9, C1Qa, C1Qb, Arpc1B, Grn, Cxcl16, Tgif1, Cp, Smarca1 |
| GO:0050789-regulation of biological process | 43 | 1.12E-02 | S100A4, Tspo, Spock3, C3, Aif1, Rbm3, Unc93B1, Rsad2, Itgb2, Cd53, C1Qc, Tiprl, Timp1, Cd48, P2Ry6, Rac2, Fcgr1A, Hmox1, Pycard, Cfh, Ptn, Fcer1G, Inpp5D, Csf1R, Ptprc, Pigz, Plek, Pdpn, Bcl2A1, Anxa1, Pkib, Il6R, Anxa3, Psmb8, Arhgap25, Psmb9, C1Qa, C1Qb, Arpc1B, Grn, Cxcl16, Tgif1, Smarca1 | |
| GO:0050896-response to stimulus | 41 | 8.03E-04 | Itgal, Tspo, Rtp4, Ifitm1, C3, Aif1, Rbm3, Ifitm3, Endou, Rsad2, Itgb2, C1Qc, Itgam, Tiprl, Timp1, Cd48, Rac2, Alox5Ap, Fcgr1A, Hmox1, Pycard, Cfh, Ptn, Fcer1G, Inpp5D, Fcgr3A, Ptprc, Plek, Rt1-Ce12, Pdpn, Hck, Anxa1, Il6R, Anxa3, C1Qa, C1Qb, Cybb, Cxcl16, Tgif1, Ctsc, Cp | |
| GO:0048518-positive regulation of biological process | 33 | 3.61E-07 | S100A4, Tspo, C3, Aif1, Rbm3, Unc93B1, Itgb2, C1Qc, P2Ry6, Rac2, Fcgr1A, Hmox1, Pycard, Cfh, Ptn, Fcer1G, Inpp5D, Csf1R, Ptprc, Pigz, Plek, Pdpn, Anxa1, Il6R, Anxa3, Psmb8, Psmb9, C1Qa, C1Qb, Cxcl16, Grn, Tgif1, Smarca1 | |
| GO:0002376-immune system process | 28 | 6.07E-15 | Itgal, Aif1, C3, Endou, Unc93B1, Rsad2, Itgb2, C1Qc, Itgam, Cd48, Fcgr1A, Cfh, Fcer1G, Inpp5D, Fcgr3A, Blnk, Ptprc, Plek, Rt1-Ce12, Bcl2A1, Anxa3, Psmb8, Psmb9, C1Qa, C1Qb, Cybb, Cxcl16, Ctse | |
| Co-expressed downregulated genes | GO:0048856-anatomical structure development | 8 | 4.26E-02 | Pcsk2, Lingo1, Atp2B2, Dnm3, Vegfa, Rps6Kb1, Fgf1, Slc9A3R2 |
| GO:0007399-nervous system | 6 | 4.26E-02 | Pcsk2, Lingo1, Atp2B2, Dnm3, Vegfa, Fgf1 | |
| GO:0035556-intracellular signal transduction | 5 | 4.26E-02 | Lingo1, Map4K5, Grm7, Rps6Kb1, Fgf1 | |
| GO:0023014-signal transmission via phosphorylation event | 4 | 3.54E-02 | Lingo1, Map4K5, Rps6Kb1, Fgf1 | |
| GO:0007243-intracellular protein kinase cascade | 4 | 3.54E-02 | Lingo1, Map4K5, Rps6Kb1, Fgf1 |
GO, Gene Ontology; BP, biological process; Itgb2, integrin subunit β2; Cd53, CD53 Molecule; Rac2, Ras-related C3 botulinum toxin substrate 2; Fcgr1A, fc fragment of IgG receptor Ia; Hmox1, heme oxygenase 1; C1Qb, complement C1q B chain; Arpc1B, actin related protein 2/3 complex subunit 1B; Grn, granulin; Cxcl16, C-X-C motif chemokine ligand 16; Tgif1, TGFB induced factor homeobox 1; P2Ry6, pyrimidinergic receptor P2Y6; Arhgap25, r GTPase activating protein 25; Psmb9, proteasome subunit β9; C1Qa, complement C1q A chain; Smarca1, WI/SNF related, matrix associated, actin dependent regulator of chromatin, subfamily A, member 1; Itgam, integrin subunit α M; Tiprl, TOR signaling pathway regulator; Timp1, TIMP metallopeptidase inhibitor 1; Cd48, CD48 molecule; Pycard, PYD and CARD domain containing; Cfh, complement factor H; Cybb, cytochrome B-245 β chain Plek, pleckstrin; Pdpn, podoplanin; Anxa1, annexin A1; Il6R, interleukin 6 receptor; Anxa3, annexin A3; Tyrobp, TYRO protein tyrosine kinase binding protein; Itgal, Integrin subunit α L; Blnk, B-cell linker; Ptprc, protein tyrosine phosphatase, receptor type C; Bcl2L1, BCL2 like 1; Pcsk2, proprotein convertase subtilisin/kexin type 2; Lingo1, leucine rich repeat and Ig domain containing 1; Atp2B2, ATPase plasma membrane Ca2+ transporting 2; Dnm3, dynamin 3; Vegfa, vascular endothelial growth factor A; Rps6Kb1, ribosomal protein S6 kinase B1; Fgf1, fibroblast growth factor 1; Slc9A3R2, SLC9A3 regulator 2; Map4K5, mitogen-activated protein kinase kinase kinase kinase 5; Grm7, glutamate metabotropic receptor 7.
The PPI network was composed of 92 co-expressed genes (83 upregulated and 9 downregulated) and 351 interactions. The connectivity degree of six genes was more than 20 and they were Tyrobp (degree=35), CD68 molecule (Cd68; degree=34), Rac2 (degree=29), integrin subunit β2; (degree=28), CD53 molecule (degree=25), C-type lectin domain family 4 member A (degree=22). Tyrobp interacted with multiple genes, including Cd68 and Rac2 (Fig. 4).
Figure 4.

The protein-protein interaction network of co-expressed genes. The red nodes represent upregulated genes; and the green nodes represent downregulated genes. A dotted line between two nodes indicates that there is an interaction between these two nodes.
Analysis of TFs
Based on the AnimalTFDB database, a total of 9 TFs were identified from the DEGs common to the 1 and 3 week injury samples, including cold shock domain containing C2, Pgr, zinc finger and BTB domain containing 7B, SRY-box 18, activating TF 3 (Atf3), MAF BZIP TF B (Mafb), Tgif1, Fli-1 proto-oncogene, ETS TF (Fli1) and T-box 4 (Tbx4). Among them, Atf3, Mafb, Tbx4, Tgif1 and Fli1 were all upregulated in 1 and 3 week injury samples, compared with sham samples, while the others were downregulated (Fig. 5).
Figure 5.
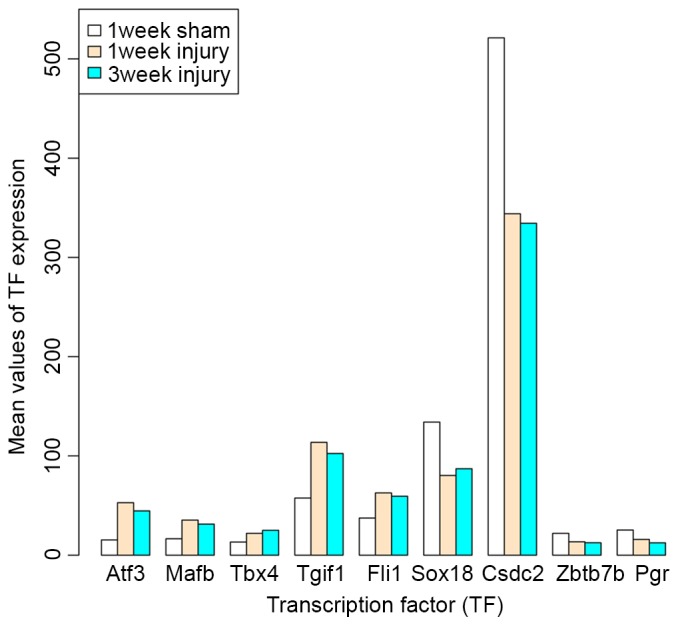
The bar diagram displaying the mean expression of TFs identified from the differentially expressed genes common to the 1 week and 3 week injury samples. The white bars represent lumbar spinal cord samples obtained from rats 1 week following a sham laminectomy; the yellow bars and blue bars represent lumbar spinal cord samples obtained from rats 1 and 3 weeks following contusive spinal cord injury at the T9 level. TF, transcription factor.
Discussion
In the present study, 234 upregulated and 51 downregulated DEGs were common to the 1 week and 3 week injury samples, compared with the sham samples. Among them, 139 genes had co-expression associations and the majority of them were upregulated genes. The upregulated co-expressed genes were predominantly enriched in several GO terms of biological regulation, including Tgif1 and Rac2.
Tgif1 was identified as a TF in the present study. It belongs to the three-amino acid loop extension superclass of atypical homeodomains (15). Studies (16–18) have showed that Tgif1 exerts crucial functions in the nervous system. Additionally, a previous study (19) identified TGIF1 as a novel regulator of macrophage activation in immune response. In the present study, Tgif1 had a co-expression associations with Rac2 and Tyrobp, the two of which had a higher degree in the PPI network. Rac2 encodes a member of the Ras superfamily of small guanosine triphosphate (GTP)-metabolizing proteins, and it modulates diverse processes, including secretion, cell polarization and phagocytosis (20). In the present study, Rac2 was identified to be significantly enriched in several pathways, including natural killer cell-mediated cytotoxicity. Natural killer cells participate in immune processes after spinal cord injury (21) and there is evidence that suppression of Rac activity in the injured spinal cord enhances cell survival (22). It has been demonstrated that the expression of Rac2 is activated in inflammatory responses (23). Furthermore, Ras GTPases exert critical functions in multiple procedures during axonogenesis in injured spinal cords (24). Tyrobp is a transmembrane signaling polypeptide which has an immunoreceptor tyrosine-based activation motif and it serves a role in signal transduction, brain myelination, and inflammation (25,26). In the current study, Tyrobp was enriched in natural killer cell mediated cytotoxicity, interacted with Rac2 in the PPI network and co-expressed with Tgif1. Together, Tgif1, Rac2 and Tyrobp may play pivotal roles in TSCI.
Among the downregulated genes, Pgr, identified as a TF, was highly enriched in female gonad development and the ovulation cycle process. In axonal regeneration, gonadal steroids function as promoting factors (27). Estrogens have direct neuroprotective effects, including modification of humoral immune responses, and gestagens can prevent neuronal death and promote the growth of nervous cells and the formation of new synapses (28). A previous study (29) confirmed that progesterone provides neuroprotection to the injured central and peripheral nervous system in the injured spinal cord. Therefore, Pgr may serve a key role in the regulation of nervous regeneration in spinal cord injuries.
In conclusion, 234 upregulated and 51 downregulated DEGs were differentially expressed in 1 and 3 week injury samples. Among them, the upregulated genes Rac2 and Tyrobp, which are associated with natural killer cell-mediated cytotoxicity, may have crucial functions in TSCI. Tgif1 and Pgr may exert a regulatory function in TSCI. These observations require experimental validation, however they are expected to aid the elucidation of the molecular mechanisms in TSCI.
References
- 1.Jain NB, Ayers GD, Peterson EN, Harris MB, Morse L, O'Connor KC, Garshick E. Traumatic spinal cord injury in the United States, 1993–2012. Jama. 2015;313:2236–2243. doi: 10.1001/jama.2015.6250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Chen MJ, Kress B, Han X, Moll K, Peng W, Ji RR, Nedergaard M. Astrocytic CX43 hemichannels and gap junctions play a crucial role in development of chronic neuropathic pain following spinal cord injury. Glia. 2012;60:1660–1670. doi: 10.1002/glia.22384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Chen M, Ni Y, Liu Y, Xia X, Cao J, Wang C, Mao X, Zhang W, Chen C, Chen X, Wang Y. Spatiotemporal expression of EAPP modulates neuronal apoptosis and reactive astrogliosis after spinal cord injury. J Cell Biochem. 2015;116:1381–1390. doi: 10.1002/jcb.25096. [DOI] [PubMed] [Google Scholar]
- 4.Zhu P, Hata R, Nakata K, Cao F, Samukawa K, Fujita H, Sakanaka M. Intravenous infusion of ginsenoside Rb1 ameliorates compressive spinal cord injury through upregulation of Bcl-xL and VEGF. Int J Neurol Neurother. 2015;2:1. doi: 10.23937/2378-3001/2/1/1017. [DOI] [Google Scholar]
- 5.De Biase A, Knoblach SM, Di Giovanni S, Fan C, Molon A, Hoffman EP, Faden AI. Gene expression profiling of experimental traumatic spinal cord injury as a function of distance from impact site and injury severity. Physiol Genom. 2005;22:368–381. doi: 10.1152/physiolgenomics.00081.2005. [DOI] [PubMed] [Google Scholar]
- 6.Byrnes KR, Garay J, Di Giovanni S, De Biase A, Knoblach SM, Hoffman EP, Movsesyan V, Faden AI. Expression of two temporally distinct microglia-related gene clusters after spinal cord injury. Glia. 2006;53:420–433. doi: 10.1002/glia.20295. [DOI] [PubMed] [Google Scholar]
- 7.Sharp J, Frame J, Siegenthaler M, Nistor G, Keirstead HS. Human embryonic stem cell-derived oligodendrocyte progenitor cell transplants improve recovery after cervical spinal cord injury. Stem Cells. 2010;28:152–163. doi: 10.1002/stem.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Shin HY, Kim H, Kwon MJ, Hwang DH, Lee K, Kim BG. Molecular and cellular changes in the lumbar spinal cord following thoracic injury: Regulation by treadmill locomotor training. PLoS One. 2014;9:e88215. doi: 10.1371/journal.pone.0088215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gautier L, Cope L, Bolstad BM, Irizarry RA. Affy-analysis of Affymetrix GeneChip data at the probe level. Bioinformatics. 2004;20:307–315. doi: 10.1093/bioinformatics/btg405. [DOI] [PubMed] [Google Scholar]
- 10.Smyth GK. Bioinformatics and computational biology solutions using R and Bioconductor. Springer; 2005. Limma: Linear models for microarray data; pp. 397–420. [DOI] [Google Scholar]
- 11.Obayashi T, Kinoshita K. Rank of correlation coefficient as a comparable measure for biological significance of gene coexpression. DNA Res. 2009;16:249–260. doi: 10.1093/dnares/dsp016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kohl M, Wiese S, Warscheid B. Data Mining in Proteomics. Springer; 2011. Cytoscape: Software for visualization and analysis of biological networks; pp. 291–303. [DOI] [PubMed] [Google Scholar]
- 13.Maere S, Heymans K, Kuiper M. BiNGO: A Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics. 2005;21:3448–3449. doi: 10.1093/bioinformatics/bti551. [DOI] [PubMed] [Google Scholar]
- 14.Zhang HM, Liu T, Liu CJ, Song S, Zhang X, Liu W, Jia H, Xue Y, Guo AY. Animal TFDB 2.0: A resource for expression, prediction and functional study of animal transcription factors. Nucleic Acids Res. 2015;43(Database Issue):D76–D81. doi: 10.1093/nar/gku887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yang Y, Hwang CK, D'Souza UM, Lee SH, Junn E, Mouradian MM. Three-amino acid extension loop homeodomain proteins Meis2 and TGIF differentially regulate transcription. J Biol Chem. 2000;275:20734–20741. doi: 10.1074/jbc.M908382199. [DOI] [PubMed] [Google Scholar]
- 16.Huang J, Wah IY, Pooh RK, Choy KW. Molecular genetics in fetal neurology. Semin Fetal Neonatal Med. 2012;17:341–346. doi: 10.1016/j.siny.2012.07.007. [DOI] [PubMed] [Google Scholar]
- 17.Kerr TC, Cuykendall TN, Luettjohann LC, Houston DW. Maternal Tgif1 regulates nodal gene expression in Xenopus. Dev Dyn. 2008;237:2862–2873. doi: 10.1002/dvdy.21707. [DOI] [PubMed] [Google Scholar]
- 18.Sha L, Kitchen R, Porteous D, Blackwood D, Muir W, Pickard B. SOX11 target genes: Implications for neurogenesis and neuropsychiatric illness. Acta Neuropsychiatrica. 2012;24:16–25. doi: 10.1111/j.1601-5215.2011.00583.x. [DOI] [PubMed] [Google Scholar]
- 19.Ramsey SA, Klemm SL, Zak DE, Kennedy KA, Thorsson V, Li B, Gilchrist M, Gold ES, Johnson CD, Litvak V, et al. Uncovering a macrophage transcriptional program by integrating evidence from motif scanning and expression dynamics. PLoS Comput Biol. 2008;21:e1000021. doi: 10.1371/journal.pcbi.1000021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Didsbury J, Weber RF, Bokoch GM, Evans T, Snyderman R. Rac, a novel ras-related family of proteins that are botulinum toxin substrates. J Biol Chem. 1989;264:16378–16382. [PubMed] [Google Scholar]
- 21.Iversen PO, Hjeltnes N, Holm B, Flatebo T, Strom-Gundersen I, Ronning W, Stanghelle J, Benestad HB. Depressed immunity and impaired proliferation of hematopoietic progenitor cells in patients with complete spinal cord injury. Blood. 2000;96:2081–2083. [PubMed] [Google Scholar]
- 22.Numano F, Inoue A, Enomoto M, Shinomiya K, Okawa A, Okabe S. Critical involvement of Rho GTPase activity in the efficient transplantation of neural stem cells into the injured spinal cord. Mol Brain. 2009;2:37. doi: 10.1186/1756-6606-2-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Tian Y, Autieri MV. Cytokine expression and AIF-1-mediated activation of Rac2 in vascular smooth muscle cells: A role for Rac2 in VSMC activation. Am J Physiol Cell Physiol. 2007;292:C841–C849. doi: 10.1152/ajpcell.00334.2006. [DOI] [PubMed] [Google Scholar]
- 24.Hall A, Lalli G. Rho and Ras GTPases in axon growth, guidance, and branching. Cold Spring Harb Perspect Biol. 2010;2:a001818. doi: 10.1101/cshperspect.a001818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Lanier LL, Corliss BC, Wu J, Leong C, Phillips JH. Immunoreceptor DAP12 bearing a tyrosine-based activation motif is involved in activating NK cells. Nature. 1998;391:703–707. doi: 10.1038/35642. [DOI] [PubMed] [Google Scholar]
- 26.Gingras MC, Lapillonne H, Margolin JF. TREM-1, MDL-1 and DAP12 expression is associated with a mature stage of myeloid development. Mol Immunol. 2002;38:817–824. doi: 10.1016/S0161-5890(02)00004-4. [DOI] [PubMed] [Google Scholar]
- 27.Jones KJ. Gonadal steroids as promoting factors in axonal regeneration. Brain Res Bull. 1993;30:491–498. doi: 10.1016/0361-9230(93)90283-H. [DOI] [PubMed] [Google Scholar]
- 28.Pakulski C. Neuroprotective properties of sex hormones. Anestezjol Intens Ter. 2011;43:113–118. (In Polish) [PubMed] [Google Scholar]
- 29.Gonzalez SL, Labombarda F, González Deniselle MC, Guennoun R, Schumacher M, De Nicola AF. Progesterone up-regulates neuronal brain-derived neurotrophic factor expression in the injured spinal cord. Neuroscience. 2004;125:605–614. doi: 10.1016/j.neuroscience.2004.02.024. [DOI] [PubMed] [Google Scholar]