

ABSTRACT
We present the effects of two novel bumped kinase inhibitors, BKI-1517 and BKI-1553, against Neospora caninum tachyzoites in vitro and in experimentally infected pregnant mice. These compounds inhibited tachyzoite proliferation of a transgenic beta-galactosidase reporter strain cultured in human foreskin fibroblasts with 50% inhibitory concentrations (IC50s) of 0.05 ± 0.03 and 0.18 ± 0.03 μM, respectively. As assessed by an alamarBlue assay, fibroblast IC50s were above 20 μM; however, morphological changes occurred in cultures treated with >5 μM BKI-1517 after prolonged exposure (>6 days). Treatment of intracellular tachyzoites with 5 μM BKI-1553 for 6 days inhibited endodyogeny by interfering with the separation of newly formed zoites from a larger multinucleated parasite mass. In contrast, parasites treated with 5 μM BKI-1517 did not form large complexes and showed much more evidence of cell death. However, after a treatment duration of 10 days in vitro, both compounds failed to completely prevent the regrowth of parasites from culture. BALB/c mice experimentally infected with N. caninum Spain7 (Nc-Spain7) and then treated during 6 days with BKI-1517 or BKI-1553 at different dosages showed a significant reduction of the cerebral parasite load. However, fertility was impaired by BKI-1517 when applied at 50 mg/kg of body weight/day. At 20 mg/kg/day, BKI-1517 significantly inhibited the vertical transmission of N. caninum to pups and increased the rate of survival of offspring. BKI-1553 was less detrimental to fertility and also provided significant but clearly less pronounced protection of dams and offspring. These results demonstrate that, when judiciously applied, this compound class protects offspring from vertical transmission and disease.
KEYWORDS: apicomplexan parasites, Neospora caninum, chemotherapy, vertical transmission, bumped kinase inhibitor, calcium-dependent protein kinase, pregnant mouse model, fertility
INTRODUCTION
Neospora caninum, a cyst-forming apicomplexan parasite that is closely related to Toxoplasma gondii, is one of the most important infectious causes of bovine abortion, stillbirth, and birth of weak calves, collectively named neosporosis (1). Furthermore, N. caninum infection may result in the birth of clinically healthy but persistently infected calves transmitting the parasite to the next generation. Neosporosis has also been described in other ruminants of economic importance, such as sheep and goats; was shown to cause neuromuscular disease in dogs; and has been detected in a wide range of other species and wild animals worldwide (1–3). In cattle, N. caninum infection is estimated to be responsible for annual economic losses amounting to up to 1.28 billion to 2.8 billion UD dollars, considering the data from 10 countries where statistics are available (3). The economic impact of neosporosis can be limited by the testing and culling of seropositive animals, discontinued breeding with offspring from seropositive cows, vaccination of susceptible and infected animals, and chemotherapeutic treatment of calves from seropositive cows, as discussed previously (4).
While vaccination has been widely exploited and is still regarded as potentially the most successful approach to the control of neosporosis (1, 4, 5), chemotherapy has not really been considered a promising strategy due to the expected development of resistance and the fact that drug residues could be retained in meat or milk for extended periods of time. However, there is no efficacious vaccine in sight; a wide range of compounds have been demonstrated to limit tachyzoite proliferation in vitro, and several compounds showed parasiticidal activity. A limited number of drugs was shown to be at least partially active in mice, as reviewed previously (4), and a few have been used to treat neosporosis in dogs (6, 7). A successful strategy for the development of chemotherapeutics against neosporosis may reside in the repurposing of compounds developed against other apicomplexan parasites.
More recently, target-based approaches in antiparasitic drug development have been focusing on kinases (8), especially calcium-dependent protein kinase 1 (CDPK1), which is essential for motility, host cell invasion, and egress of tachyzoites (9, 10). CDPK1 has no homologues in mammalian hosts (9, 11, 12). As a consequence of these efforts, a class of ATP-competitive inhibitors named bumped kinase inhibitors (BKIs) has been developed. The efficacy of these BKIs is linked to the presence of a glycine (Gly) residue in the hydrophobic pocket of the ATP-binding site of CDPK1. This particular amino acid is also called the “gatekeeper residue.” In mammalian kinases, this position is occupied by a larger residue such as methionine (Met), which then blocks the access of the BKI to the pocket. Thus, mammalian kinases are not affected by BKIs, and this renders mammalian cells largely resistant (13). One compound of this class, BKI-1294, is effective against cryptosporidiosis in immunosuppressed mice (14) and against toxoplasmosis in vitro and in vivo (15). BKI-1294 is also effective against N. caninum in vitro and in vivo (16) and strongly interferes with the vertical transmission of neosporosis to newborn pups in a pregnant mouse model of neosporosis (17).
Based on the naphthalenyl-pyrazolopyrimidine scaffold of BKI-1294, a novel compound, BKI-1553, has been developed (Fig. 1). BKI-1553 has improved activity against T. gondii in vitro and lower human ether-a-go-go-related gene (hERG) ion channel inhibition, crosses the blood-brain barrier in mice when orally applied, and reduces the T. gondii burden in brains, lungs, and livers of infected mice (18). Moreover, derivatives of these BKIs with different scaffolds have been created based on structure-activity relationship studies with T. gondii CDPK1 (TgCDPK1) as the main target (19). One of these novel compounds, BKI-1517, has a substituted quinolone-3-pyrazole scaffold (Fig. 1). This compound inhibits TgCDPK1 with a 50% inhibitory concentration (IC50) in the nanomolar range and inhibits T. gondii tachyzoite proliferation in vitro with an IC50 in the submicromolar range. In a mouse model, BKI-1517 given orally has suitable pharmacokinetics (PK) and exhibits high efficacy against intraperitoneally applied T. gondii tachyzoites (19). These results suggest that these compounds could also be highly active against N. caninum.
FIG 1.

Structures of the compounds BKI-1294, BKI-1517, BKI-1553, and quinine.
Here, we present in vitro studies with BKI-1517 and BKI-1553 in human foreskin fibroblasts (HFFs) infected with N. caninum tachyzoites. We show that both compounds strongly interfere with tachyzoite invasion and also inhibit the completion of tachyzoite cytokinesis. However, these compounds differ with respect to the ultrastructural changes that they induce. Moreover, we have performed a series of in vivo studies in a pregnant neosporosis mouse model and show that treatments with BKI-1517 and BKI-1553 prevent acute neosporosis in infected dams and effectively inhibit the vertical transmission of N. caninum. However, at high doses that do not affect the health of nonpregnant mice, both compounds are detrimental to fertility in pregnant mice. Our data provide a proof of concept for the treatment of acute N. caninum infection and the protection of offspring by CDPK1 inhibitors.
RESULTS
In vitro efficacies of BKI-1517 and BKI-1553 against N. caninum tachyzoites.
The inhibitory effects of BKI-1517 and BKI-1553 in comparison to BKI-1294 (16, 17) against N. caninum tachyzoites were assessed by adding the drugs to the cultures at the time point of infection and then measuring parasite proliferation after 3 days (Fig. 2). In parallel, the cytotoxicity of these compounds on HFF host cells, and on two proliferative cell lines, namely, Vero cells and rat hepatoma (RH) cells, was determined by an alamarBlue assay. Similarly to BKI-1294, BKI-1517 and BKI-1553 inhibited N. caninum tachyzoite proliferation at a concentration range of 0.05 to 0.18 μM. As shown in Table 1, the IC50 of BKI-1553 calculated from the inhibition curve was similar to that of BKI-1294, whereas the IC50 of BKI-1517 was about three times lower.
FIG 2.
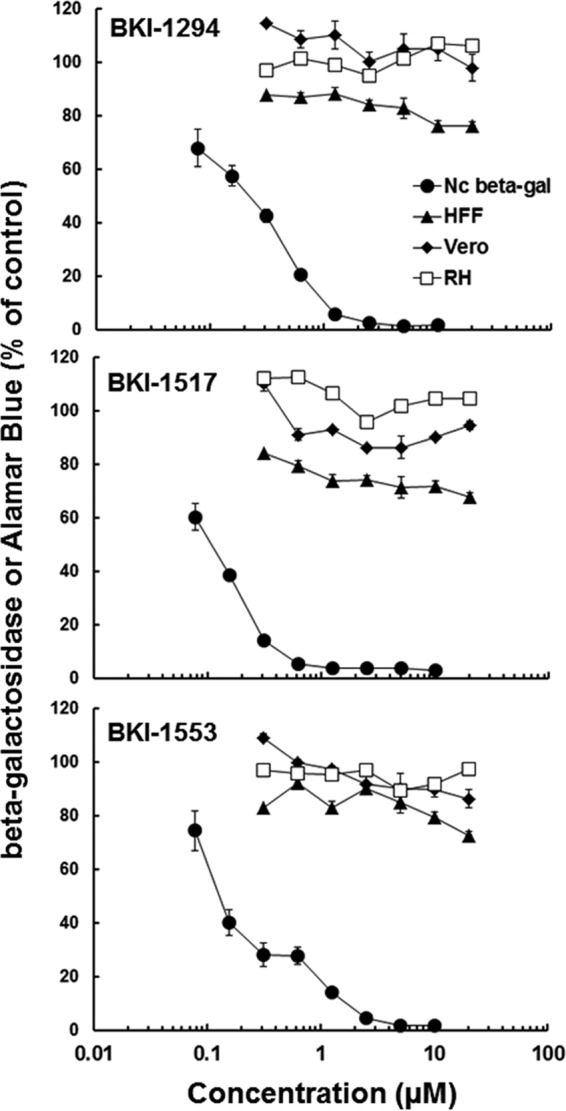
The novel BKIs inhibit N. caninum proliferation and are not cytotoxic in vitro. The inhibition of N. caninum tachyzoite proliferation was determined by using a beta-galactosidase reporter strain as detailed in Materials and Methods. Cytotoxicity was determined for HFF, Vero, and rat RH cells by using an alamarBlue assay as described in Materials and Methods. Mean values ± standard errors for quadruplicates are indicated.
TABLE 1.
In vitro efficacies of the compounds used in this studya
| Cell type | Mean IC50 ± SD |
||
|---|---|---|---|
| BKI-1294 | BKI-1517 | BKI-1553 | |
| N. caninum tachyzoites | 0.17 ± 0.04 | 0.05 ± 0.03 | 0.18 ± 0.03 |
| HFFs | >20 | >20 | >20 |
| Vero cells | >20 | >20 | >20 |
| RH cells | >20 | >20 | >20 |
IC50s in HFFs, Vero cells, and rat hepatoma cells treated for 3 days were above 20 μM, and no cytotoxicity was observed. However, upon prolonged (6 days or more) exposure of HFF monolayers to 5 μM BKI-1517, but not BKI-1553, HFFs started to acquire aberrant morphological features such as forming spheres and partial detachment (data not shown), which is suggestive of some structural interference, despite the absence of discernible alterations by an alamarBlue assay. Thus, longer-term treatment studies of BKI-1517 (up to 10 days) were carried out with a concentration of 2 μM, a concentration at which no effects were observed. Ten days of continuous exposure of N. caninum-infected HFFs to BKI-1553 at 5 μM and BKI-1517 at 2 μM did not lead to parasiticidal activity, and regrowth of parasites occurred at the latest 10 to 12 days after drug-containing medium was replaced by normal medium without the compound.
Morphological and ultrastructural effects exerted by BKI-1553 and BKI-1517.
Immunofluorescence microscopy and transmission electron microscopy (TEM) were employed to visualize the drug-induced alterations in N. caninum-infected HFFs. In control cultures fixed and processed at day 3 postinfection (p.i.), antibody staining with a monoclonal antibody specific for the N. caninum tachyzoite surface antigen SAG1 (NcSAG1) (mAbSAG1) and a tetramethyl rhodamine isocyanate (TRITC)-conjugated secondary antibody revealed the presence of larger parasitophorous vacuoles (PVs) containing numerous newly formed tachyzoites. Staining with an antibody directed against NcMAG1, known to be upregulated in bradyzoite-containing tissue cysts (20), revealed very little labeling and only within the matrix between individual tachyzoites (Fig. 3A to C). When cultures were treated with BKI-1553, larger parasitic masses staining positive for NcSAG1 were already seen after 2 days of treatment, but the effect became more clearly visible after 6 days of treatment (Fig. 3D to I). In addition, MAG1 staining shifted from an internal localization with respect to the PV after 2 days (Fig. 3E) toward more peripheral areas at day 6 (Fig. 3H).
FIG 3.
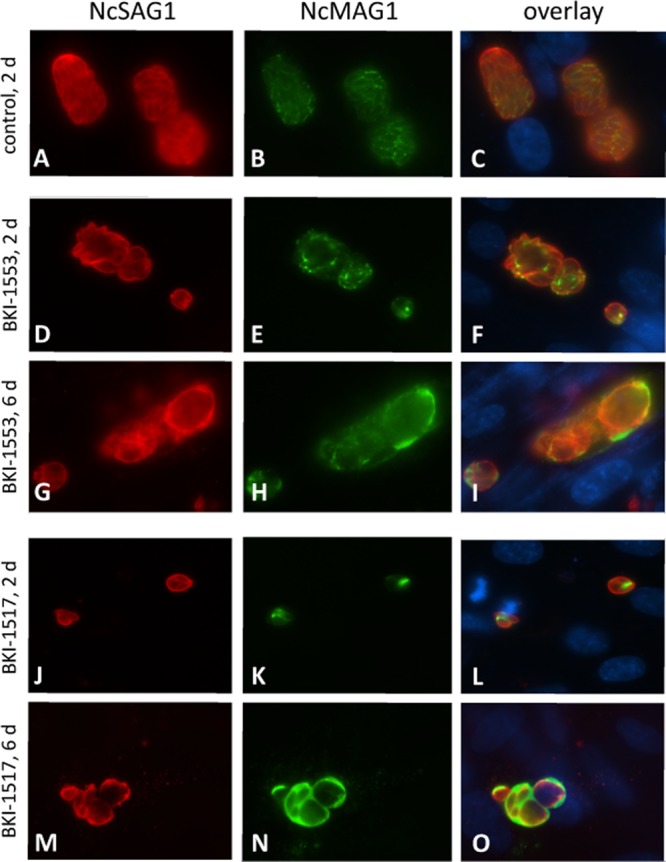
Immunofluorescence labeling of Neospora-infected fibroblasts cultured in the presence and absence of BKIs. Parasites were cultured during 2 days (A to F and J to L) and 6 days (G to I and M to O) either without the drug (A to C) or in the presence of BKI-1553 and BKI-1517 (D to O) and fixed and stained with anti-SAG1 (A, D, G, J, and M) and anti-MAG1 (B, E, H, K, and N). Note the presence of individual tachyzoites in the controls.
In contrast, treatment with BKI-1517 clearly inhibited the intracellular increase of the parasite mass already early on, as indicated in Fig. 3J to L, after 2 days. Also, at day 6 of treatment, PV dimensions were relatively small, containing only a few tachyzoites, and a dramatic increase in the labeling of NcMAG1 was observed, located between individual tachyzoites and at the PV periphery, showing that this antigen was extensively secreted into the PV during treatment (Fig. 3M to O).
These observations were largely corroborated by TEM. Electron microscopy of untreated cultures carried out at day 4 postinfection demonstrated the presence of individual tachyzoites within the PV (Fig. 4A). BKI-1553-treated parasites appeared to still undergo limited proliferation on day 2, as in several PVs, more than one small complex was detected, suggesting that tachyzoites had achieved one or two rounds of full endodyogeny prior to being blocked in their cell cycle (Fig. 4B). On day 6 of BKI-1553 treatment, larger complexes, often with small apical complex precursors sticking out in the periphery, were detected (Fig. 4C). BKI-1517 treatment exhibited more toxic effects. On day 2 of treatment, small PVs containing mostly only one or two parasites were visible (Fig. 5A and B), some of them with large cytoplasmic vacuoles (Fig. 5C). From day 4 onward, progressive signs of cellular distortion were visible, as indicated in Fig. 5D, and the PV matrix became filled with a granular and electron-dense matrix that started to completely enclose the parasites. On day 6, most tachyzoites appeared nonviable and were completely embedded in the granular matrix of the PV (Fig. 5E). Thus, overall, the antiparasitic effect of BKI-1517 seemed to be more pronounced.
FIG 4.

TEM of N. caninum tachyzoites grown in HFFs and treated with BKI-1553. Cultures remained untreated for 4 days (A) or were treated with 5 μM BKI-1553 for 2 days (B) or 6 days (C). The inset in panel C is a low-magnification view of the complex shown in panel C. Note the progressive buildup of multinucleated complexes. Bars, 2.6 μm (A and B), 0.5 μm (C), and 4.8 μm (inset). Note the progressive buildup of multinucleated complexes in BKI-1553-treated cultures.
FIG 5.

TEM of N. caninum tachyzoites treated with BKI-1507. Parasites were cultured in HFFs and treated with 5 μM BKI-1517 for 2 days (A to C), 4 days (D), and 6 days (E). (A to C) After 2 days of treatment, only vacuoles containing one or two parasites, albeit slightly enlarged, were found, and the PV matrix appears rather granular, with some of them exhibiting large vacuolization (C). (D and E) At 4 days of treatment (D), and especially at 6 days (E), parasites show clear signs of structural deterioration and are often embedded in a dense matrix. Bars, 0.7 μm (A and B), 0.9 μm (C and D), 0.5 μm (E). Insets in panels A to C represent the respective low-magnification views (bars, 4.8 μm).
Treatment of pregnant and nonpregnant mice with BKI-1553.
In experiment I, we applied BKI-1553 emulsified in corn oil at 20 mg/kg body weight daily during 5 days. The corresponding placebo group received corn oil during 5 days. None of the nonpregnant mice exhibited clinical signs of neosporosis during 3 weeks p.i. At this point, only 1 of 8 mice from the treated group was positive for N. caninum in brain samples (brain positive), compared to all (8 out of 8) placebo-treated mice (Table 2 and Fig. 6A). These differences were highly significant. In the dams euthanized at 30 days postpartum (p.p.), and thus 6 weeks p.i., the treatment effect was still prominent, with a significantly lower number of brain-positive dams (Table 2) and significantly lower cerebral parasite loads in BKI-1553-treated dams than in the placebo group (Fig. 6B). BKI-1553 had no significant effects on reproductive performance as assessed via the determination of the pregnancy rate and the litter size but unfortunately caused a significantly higher neonatal mortality rate, with 15 out of 40 pups dying within the first 3 days after birth (Table 2). The 25 surviving pups, however, had significantly reduced postnatal mortality rates (Table 2) and a good survival rate (Fig. 6C) compared to those of pups from the placebo group. Only 14 of the 25 surviving pups were brain positive at the end of the experiment on day 30 p.p. compared to pups from the placebo group, all of which were brain positive (Table 2).
TABLE 2.
Effects of BKI-1553 treatment on mortality, fertility, and cerebral N. caninum Nc-Spain7 infection in nonpregnant mice, dams, and pupsc
| Expt | Formulation and duration | No. of mice/total no. of mice |
Litter size (no. of pups/no. of dams) | No. of pups/total no. of pups |
||||
|---|---|---|---|---|---|---|---|---|
| Seropositive | Nonpregnant mice, N. caninum brain positive | Dams, N. caninum brain positive | Neonatal mortalitya | Postnatal mortalityb | N. caninum brain positive | |||
| I | 0 mg/kg for 5 days | 16/16 | 8/8 | 8/8 | 43/8 | 2/43 | 40/41 | 41/41 |
| 20 mg/kg for 5 days | 17/17 | 1/8** | 2/9** | 40/9 | 15/40*** | 9/25*** | 14/25*** | |
| II | 0 mg/kg for 6 days | 16/16 | 6/6 | 9/10 | 56/10 | 1/56 | 55/55 | 55/55 |
| 20 mg/kg for 3 days with 1-day intervals of 0 mg/kg | 16/16 | 2/5* | 8/11 | 60/11 | 0/60 | 48/60*** | 52/60** | |
| Not infected | 0/14 | 0/7 | 0/7 | 47/7 | 0/47 | 0/47 | ||
Proportion of pups born dead or that died within the 2 first days postpartum.
Proportion of pups dead from days 3 to 30 p.p. (considered N. caninum positive).
BALB/c mice were treated with BKI-1553, infected with Nc-Spain7, or neither treated nor infected and euthanized as described in Materials and Methods. Adults were tested for N. caninum seropositivity. Adults and surviving pups were tested for the presence of N. caninum in their brains by real-time PCR. Pups that had died before the end of the experiment were considered N. caninum positive. The respective numbers of animals in the treated and placebo groups were compared by chi-square tests (*, P < 0.05; **, P < 0.01, ***, P < 0.001).
FIG 6.
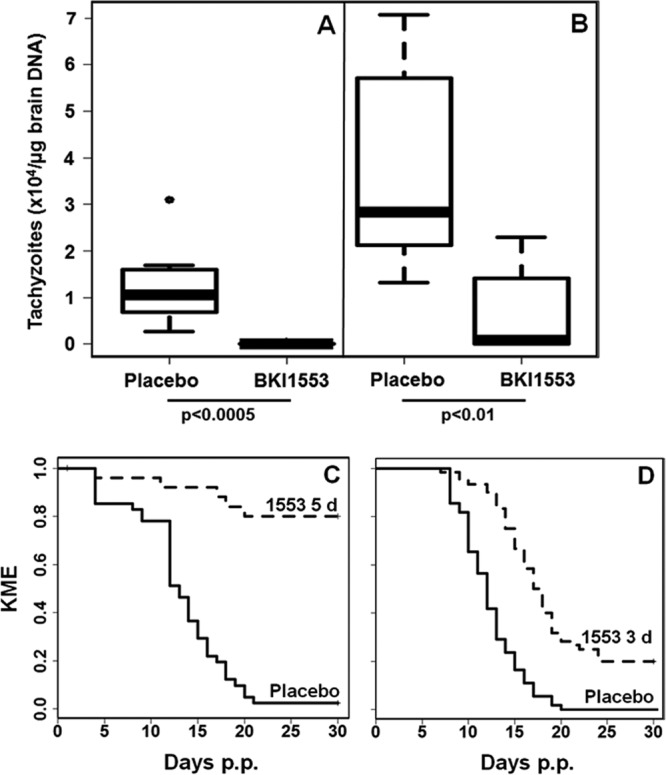
Parasite loads in brains of mice treated with BKI-1553 and the respective survival curves for offspring during 30 days p.p. Female BALB/c mice were mated, infected with Nc-Spain7 tachyzoites, and subsequently treated with BKI-1553 as detailed in Materials and Methods. Nonpregnant mice were euthanized at 3 weeks p.i., and dams were euthanized at 30 days p.p. (and thus at 44 days p.i.). (A and B) After euthanasia, brains were collected, and the tachyzoite load was quantified by real-time PCR and is presented as box plots. (A) Nonpregnant mice; (B) dams. The significance levels (obtained after a Kruskal-Wallis test) are indicated. (C and D) Survival curves of pups during 30 days p.p. with BKI-1553 (1553) (dashed line)- and corn oil (placebo) (solid line)-treated dams. As a parameter for survival, the Kaplan-Meier estimator (KME) is shown. For both treatment schedules, the differences in BKI-1553 and placebo survival curves were highly significant (P < 10−11 [C] and P < 10−10 [D] by a log rank test).
In experiment II, BKI-1553 was administered only 3 times but with 1-day intervals after each drug treatment, during which the mice received corn oil only. The corresponding placebo group was treated during 6 days with corn oil only. In nonpregnant mice, as determined after 3 weeks p.i., treatment showed a clear effect, with significantly reduced numbers of brain-positive females (Table 2) and reduced cerebral parasite loads (data not shown). However, in dams that were analyzed after 6 weeks p.i., this effect was obliterated, with 8 out of 11 animals exhibiting infection in the central nervous system (CNS). In addition, while neonatal mortality could be avoided by using this treatment regimen, the postnatal mortality rate was relatively high (48 out of 60 pups) (Fig. 6D), and 52 out of the 60 pups were brain positive. However, statistically, these parameters were still significantly reduced compared to those of the placebo group (Table 2).
Treatment of pregnant and nonpregnant mice with BKI-1517.
Based on previously reported PK data (19), we applied BKI-1517 at 50 mg/kg of body weight during 6 days, thus in a scheme similar to the one established previously for BKI-1294 (17). The corresponding placebo group received corn oil. At 3 weeks p.i., no parasites were detected in the brains of all six nonpregnant mice of the treated group, compared to 5 out of 6 infected placebo mice (P < 0.001 by a chi-square test) (Table 3). In the dams euthanized at 30 days p.p., and thus at 6 weeks p.i., the treatment effect was still very prominent, with only 2 out of 10 brain-positive dams in the treated group versus 10 out of 10 in the placebo group (P < 0.001 by a chi-square test) (Table 3). However, with this application regimen, BKI-1517 had deleterious effects on reproductive performance, significantly decreasing the litter size and causing neonatal mortality of all born pups (Table 3). Thus, pup survival could not be assessed in this experiment.
TABLE 3.
Effects of BKI-1517 treatment on mortality, fertility, and cerebral Nc-Spain7 infection in nonpregnant mice, dams, and pupsc
| Expt | Formulation and duration | No. of mice/total no. of mice |
Litter size (no. of pups/no. of dams) | No. of pups/total no. of pups |
||||
|---|---|---|---|---|---|---|---|---|
| Seropositive | Nonpregnant mice, N. caninum brain positive | Dams, N. caninum brain positive | Neonatal mortalitya | Postnatal mortalityb | N. caninum brain positive | |||
| I | 0 mg/kg for 6 days | 16/16 | 5/6 | 10/10 | 56/10 | 1/56 | 55/55 | 55/55 |
| 50 mg/kg for 6 days | 16/16 | 0/6*** | 2/10*** | 24/10+ | 24/24*** | |||
| II | 0 mg/kg for 6 days | 16/16 | 6/6 | 10/10 | 38/10 | 0/38 | 38/38 | 38/38 |
| 10 mg/kg for 6 days | 16/16 | 5/6 | 10/10 | 51/10 | 0/51 | 49/51 | 50/51 | |
| 20 mg/kg for 6 days | 16/16 | 2/5* | 8/11 | 60/11 | 0/60 | 13/60*** | 23/60*** | |
| Not infected | 0/8 | 0/8 | 0/8 | 48/8 | 0/48 | 0/48 | ||
Proportion of pups born dead or that died within the 2 first days postpartum.
Proportion of pups dead from days 3 to 30 p.p. (considered N. caninum positive).
BALB/c mice were treated with BKI-1553, infected with Nc-Spain7, or neither treated nor infected and euthanized as described in Materials and Methods. Adults and surviving pups were tested for the presence of N. caninum in their brains by real-time PCR. Pups that had died before the end of the experiment were considered N. caninum positive. The respective numbers of animals in the treated and placebo groups were compared by chi-square tests (+, P < 0.1; *, P < 0.05; **, P < 0.01; ***, P < 0.001).
In experiment II, BKI-1517 was applied at lower doses, namely, 10 and 20 mg/kg of body weight during 6 days. With this setup, only treatment with 20 mg/kg/day reduced the number of infected brains in the nonpregnant group that was analyzed at 6 weeks p.i. (Table 3), but there was no significant effect in terms of cerebral parasite burden in this group compared to the placebo group (Fig. 7A). Analysis of the dams after 6 weeks p.i. also did not reveal any significant differences in the numbers of brain-positive dams. The quantitative parasite load, however, was significantly reduced by both doses, with 20 mg/kg yielding better protection (Fig. 7B). Treatments with BKI-1517 at 10 or 20 mg/kg/day did not affect the litter size and neonatal mortality of the pups. However, postnatal mortality and the number of brain-positive pups were significantly reduced in the 20-mg/kg group (23 out of 60 pups were PCR positive) compared to the placebo group (38 out of 38 were positive) (Table 3), with a significantly improved survival rate. However, the survival rate did not show any improvement for the group treated with 10 mg/kg BKI-1517 (Fig. 7C).
FIG 7.
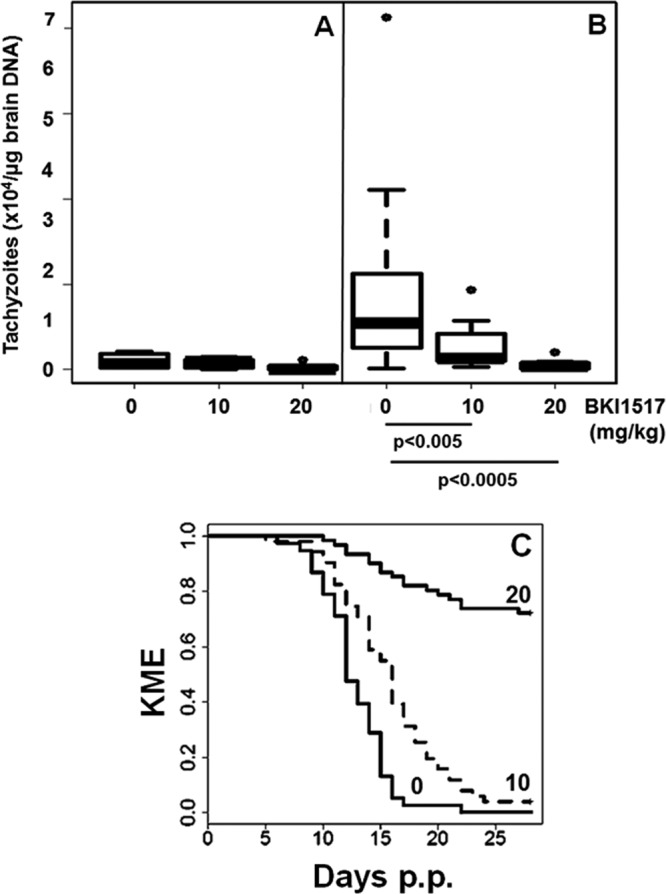
Parasite loads in brains of mice treated with BKI-1517. Female BALB/c mice were mated, infected with Nc-Spain7 tachyzoites, and subsequently treated with BKI-1517 as detailed in Materials and Methods. Nonpregnant mice were euthanized at 3 weeks p.i., and dams were euthanized at 30 days p.p. (and thus at 44 days p.i.). (A and B) After euthanasia, brains were collected, and the amount of tachyzoites was determined by quantitative PCR and are presented as box plots. (A) Nonpregnant mice; (B) dams. The significance levels for comparisons of the treated groups with the control group (obtained after a Kruskal-Wallis test followed by a pairwise Wilcoxon test) are indicated. (C and D) Survival curves of pups during 30 days p.p. of dams treated with BKI-1517 at 20 mg/kg (20) or 10 mg/kg (10) or with corn oil alone (0) for 6 days. As a parameter for survival, the Kaplan-Meier estimator (KME) is shown. For both treatment schedules, the differences in treated and placebo survival curves were highly significant (P < 10−4 for 10 mg/kg and P < 10−14 for 20 mg/kg by a log rank test).
DISCUSSION
We explored the potential of the two novel CDPK1 inhibitors BKI-1517 and BKI-1553 in vitro and in vivo: in vitro by measuring the inhibition of proliferation in short-term-treatment assays and by studying the effects of extended treatment durations by immunofluorescence and electron microscopy and in vivo by investigating the effects on parasite loads in dams and pups in an established pregnant mouse model, which is standardized to the extent that dams and offspring of uninfected, untreated animals survive, while postnatal mortality in offspring occurs only upon dose-dependent infection of the dams. In this model, which uses the N. caninum Spain7 (Nc-Spain7) isolate, which is highly virulent in vitro and in vivo in mice (21, 22) as well as in sheep (23), all pups of infected dams died within 3 weeks; pups of noninfected dams survived. Thus, the model is reproducible, as shown in several recent studies (17, 20, 24).
In vitro, both compounds inhibit N. caninum proliferation at IC50s in the submicromolar range, with BKI-1517 showing a 3-times-lower IC50 than those of BKI-1553 and BKI-1294. The viability of host cells was not affected by both compounds at 20 μM, the highest concentration tested, during a period of 3 days. In accordance to the IC50s, BKI-1517 impaired the viability of N. caninum tachyzoites more efficiently than did BKI-1553, as shown by TEM. A dramatic difference between treated cultures was noted. BKI-1517 much more clearly inhibited parasite proliferation and resulted in a higher frequency of dead parasites that were basically embedded in a granular matrix, which is most likely composed of products secreted by the parasites through dense granule organelles. The correlation between NcMAG1 labeling and TEM observations indicates that this granular matrix contains NcMAG1, a protein stored in dense granules and then secreted by tachyzoites. However, NcMAG1 was shown previously to be much more highly expressed in the PV matrix of bradyzoites than in tachyzoites (25); thus, it is likely that increased physiological stress led to the accumulation of this antigen in the PV. In contrast, BKI-1553 treatment resulted in the buildup of multinuclear complexes similar to those described previously for BKI-1294-treated cultures (16, 17), the extent of NcMAG1 staining was much lower, and no granular matrix was detected within the PV, yet both compounds did not show parasiticidal activity at their respective concentrations after 10 days of treatment, and parasites resumed proliferation once the medium was removed. However, we did not determine the titer of the parasites or try to quantitate the percentages that are viable, so it is likely that a substantial percentage of the parasites are nonviable after 10 days of exposure.
Upon an in vitro treatment duration of more than 6 days, BKI-1517 affected the morphology of fibroblast host cells in our in vitro system, thereby suggesting off-target effects in the host. The existence of such off-target effects in the host was corroborated by the detrimental effects of BKI-1517 on fertility when applied at 50 mg/kg. Whereas the treated dams showed no clinical signs due to either acute neosporosis or the compound and were almost completely devoid of parasites in their brains, the pups died within 2 days p.p. At lower doses (10 and 20 mg/kg), protection of the dams was less complete, but the pups survived and were still statistically significantly protected against neosporosis.
BKI-1553 had a better compatibility with pregnancy albeit impairing fertility to a certain extent when applied during 6 days at 20 mg/kg. The surviving offspring were protected to an extent similar to that with the previously characterized compound BKI-1294 (17). Both BKI-1517 and BKI-1553 also interfered with fertility when applied to uninfected dams (data not shown). Thus, the impairment of fertility is not due to an interaction with neosporosis. The better compatibility with pregnancy of BKI-1294 and BKI-1553 than of BKI-1517 may be due to differences in the backbones of these molecules. BKI-1294 and BKI-1553 have a naphthalenyl-pyrazolopyrimidine scaffold, whereas BKI-1517 is built on a quinolone-3-pyrazole scaffold (Fig. 1), thus sharing structural similarities with quinine. The risks of administering quinine treatment during pregnancy are well documented (26) and occur most likely due to side effects such as hypoglycemia (27), although quinine may be administered to pregnant malaria patients at least during the first trimester if no other alternatives are available (28).
Nevertheless, BKI-1517 is efficacious and well tolerated in adults and may be a compound of choice for the clearance of neosporosis in cattle prior to mating and pregnancy. After the establishment of a suitable treatment scheme, the effects on vertical transmission in cattle can be determined.
Despite the fact that in vitro, both compounds are not parasiticidal, in vivo, they significantly reduced brain positivity in treated dams even after 6 weeks p.i. despite the small treatment window. This suggests that other protective mechanisms, most likely based on innate and/or acquired immunity and similar to those elicited by live vaccines (29), may back up the initial treatment. This could be facilitated by the stimulation of the production of excess parasite antigens, such as SAG1 and MAG1, by BKI-1517 and BKI-1553. Further research will elucidate this point.
MATERIALS AND METHODS
Tissue culture media, biochemicals, and drugs.
If not stated otherwise, all tissue culture media were purchased from Gibco-BRL (Zürich, Switzerland), and biochemical reagents were obtained from Sigma-Aldrich (St. Louis, MO). Kits for molecular biology were purchased from Qiagen (Hilden, Germany). Compounds used in this study were synthesized in the Departments of Chemistry and Biochemistry of the University of Washington and shipped to Switzerland as powders. Prior to use, stock solutions of 20 mM were prepared in dimethyl sulfoxide (DMSO) and stored at −20°C.
Host cell and parasite cultures.
HFFs, Vero cells (30), and RH cells were cultured as described previously (31). N. caninum tachyzoites were maintained in Vero cells and prepared for infection of HFFs or mice as described previously (21).
In vitro determination of IC50s of BKI-1294, BKI-1517, and BKI-1553 in N. caninum and host cells.
IC50s were determined by using an N. caninum beta-galactosidase reporter strain (Nc-beta-gal) (32) and HFFs as host cells. Briefly, 103 HFFs were seeded into 96-well-plates, grown to confluence, and infected with 103 Nc-beta-gal tachyzoites per well in the presence of the compound to be tested or DMSO as a solvent control. After 3 days at 37°C with 5% CO2, medium was removed, and after one wash with phosphate-buffered saline (PBS), cells were overlaid with 0.1 ml PBS containing Triton X-100 (0.05%) and chlorophenyl red–beta-galactoside (Roche, Rotkreuz, Switzerland). Absorption was continuously read at 570 nm by using a 96-well-plate spectrophotometer (VersaMax; Molecular Devices, Sunnyvale, CA) (20). Host cell toxicity was determined not only for HFFs but also for Vero and RH cells grown under the same conditions by using an alamarBlue assay as described previously (24).
Long-term exposure to BKI-1553 and BKI-1517.
Long-term exposure assays were carried out in T25 tissue culture flasks. A total of 3 × 104 HFFs were grown to confluence, infected with 5 × 106 Nc-Spain7 tachyzoites suspended in 5 ml medium, incubated for 4 h at 37°C with 5% CO2, washed twice in HBSS (Hanks' balanced salt solution), and exposed to BKI-1553 (5 μM) or BKI-1517 (2 μM) in medium during 3, 6, 9, and 12 days. After each of these time points, drug-containing medium was replaced by medium alone, and cultures were further maintained at 37°C with 5% CO2, with medium changes being done typically every 3 days. Growth of parasites was monitored closely by light microscopy and recorded on a daily basis.
Immunofluorescence.
Immunofluorescence studies were carried out by using the Nc-Spain7 isolate and HFFs as host cells, essentially as described previously (17). In short, 104 HFFs were seeded onto poly-l-lysine-coated coverslips in 24-well plates and grown to confluence, and each well was infected with 106 Nc-Spain7 tachyzoites. After incubation at 37°C with 5% CO2 for 3 h, BKI-1553 and BKI-1517 were added at a concentration of 5 μM, and cultures treated with corresponding amounts of DMSO served as controls (2 days of incubation only). After 2, 4, and 6 days, samples were washed in PBS and fixed in 3% paraformaldehyde for 10 min, followed by permeabilization in ice-cold methanol and acetone as previously described (33). Unspecific binding sites were blocked in PBS containing 3% bovine serum albumin (BSA) overnight at 4°C. Samples were then stained sequentially with (i) a monoclonal mouse antibody directed against NcSAG1 (34), (ii) TRITC (Sigma-Aldrich)-conjugated anti-mouse antibody (Sigma-Aldrich), (iii) affinity-purified rabbit anti-NcMAG antiserum (25), and (iv) anti-mouse antibody conjugated to fluorescein isothiocyanate (FITC; Sigma-Aldrich). Primary and secondary antibodies were diluted in PBS–0.3% BSA and were applied at room temperature for 45 min each. After three washes in PBS for 5 min each, specimens were embedded in Vectashield mounting medium containing 4′,6-diamidino-2-phenylindole (Vector Laboratories, Burlingame, CA, USA). All specimens were viewed on a Nikon Eclipse E800 digital confocal fluorescence microscope. Processing of images was performed by using Openlab 5.5.2 software (Improvision; PerkinElmer, Waltham, MA, USA).
Transmission electron microscopy.
HFFs were grown to confluence in T25 flasks and infected with 5 × 106 Nc-Spain7 tachyzoites. After 3 h, cultures were supplemented with 5 μM BKI-1553 and BKI-1517, and infected cultures treated with DMSO served as controls. After 2, 4, and 6 days of treatment, the monolayers were washed with 100 mM sodium cacodylate buffer (pH 7.3) and fixed with cacodylate buffer containing 2.5% glutaraldehyde for 10 min prior to collection of the cell material by using a rubber cell scraper and centrifugation for 10 min at 1,000 × g at room temperature. The supernatant was removed, and infected cells were fixed further in glutaraldehyde-cacodylate at 4°C overnight. Postfixation in 2% OsO4, dehydration, embedding in Epon 820 epoxy resin, and cutting of ultrathin sections were done as previously described (25, 33). Specimens were viewed on a CM12 transmission electron microscope operating at 80 kV.
Animal experimentation.
All protocols involving animals were approved by the Animal Welfare Committee of the Canton of Bern under license BE115/14. All animals used in this study were handled in strict accordance with practices made to minimize suffering. Female and male BALB/c mice, 8 weeks of age, were purchased from a commercial breeder (Charles River, Sulzberg, Germany) and were maintained in a common room under controlled temperature and a 14-h/10-h light/dark cycle according to guidelines set up by the animal welfare legislation of the Swiss Veterinary Office.
Assessment of the effects of BKI-1517 and BKI-1553 on nonpregnant and pregnant BALB/c mice infected with the N. caninum Spain7 isolate.
For each experiment, female and male BALB/c mice were housed, and pregnancy was achieved after synchronization of estrus (21). Subsequently, female mice were randomly distributed into groups of 16 mice each and were subcutaneously infected with 105 tachyzoites of the Nc-Spain7 isolate at midgestation (days 5 to 8 postmating) or were left uninfected and received an inoculation of culture medium (21). At day 2 p.i., treatment with BKIs suspended in corn oil at 10 to 50 mg/kg of body weight, or with corn oil alone, was initiated. Prior to gavage, the corn oil-drug mixture was heated to 37°C to enhance the solubility of the drug. A control group remained untouched. Pregnancy was confirmed at between days 15 and 18 of gestation by weighing, and pregnant mice were then allocated into single cages to give birth on days 20 to 22 and to rear their pups for an additional 4 weeks. During this time, those females that had remained nonpregnant were maintained in cages of 3 to 5 mice. Nonpregnant mice were evaluated for clinical signs of disease twice daily and euthanized in a CO2 chamber at day 21 p.i. at the latest. Concerning dams and pups, the litter size, i.e., the number of delivered pups per dam; early pup mortality, defined as the number of full-term dead pups from birth until day 2 p.p.; postnatal mortality, defined as the number of dead pups from days 3 to 30 p.p.; and clinical signs were recorded. Surviving dams and pups were euthanized at 30 days p.p. (and thus 44 days p.i.). Blood was recovered by cardiac puncture, and sera were obtained to assess humoral immune responses. Brains were removed for subsequent determination of parasite load and stored at −20°C until further processing.
Analysis of biological samples from in vivo experiments.
To quantify the parasite load in brains, DNA purification was performed by employing the DNeasy blood and tissue kit (Qiagen, Basel, Switzerland) according to standard protocols suitable for animal tissues. The DNA concentrations in all samples were determined by using the QuantiFluor double-stranded DNA (dsDNA) system (Promega, Madison, WI, USA) according to the manufacturer's instructions and adjusted with sterile DNase-free water to 5 ng/μl. Quantification of parasite loads in brains was performed as described previously (16, 24). Specific antibodies to N. caninum were assessed by an enzyme-linked immunosorbent assay (ELISA) as described previously (35, 36).
Statistics.
Statistical analysis of the parasite burdens in brains was performed by using the Kruskal-Wallis test followed by the Wilcoxon rank sum test. Survival analysis of the pups was performed on the corresponding Kaplan-Meier estimator by using the log rank test. Nominal data were analyzed by using the chi-square test. All analyses were performed by using the R software package (37).
ACKNOWLEDGMENTS
We acknowledge financial support by the Swiss National Science Foundation (grant no. 310030_165782). This work was supported by the Public Health Service, National Institutes of Health, Bethesda, MD (grants R01 AI 111341 and R01 HD 080670), and the U.S. Department of Agriculture (grant 2014-67015-22106).
Wesley C. Van Voorhis is the founder of the company ParaTheraTech Inc., dedicated to bring BKIs to market for animal health indications. None of the other authors have any competing interests in the manuscript.
REFERENCES
- 1.Dubey JP, Schares G, Ortega-Mora LM. 2007. Epidemiology and control of neosporosis and Neospora caninum. Clin Microbiol Rev 20:323–367. doi: 10.1128/CMR.00031-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Monney T, Hemphill A. 2014. Vaccines against neosporosis: what can we learn from the past studies? Exp Parasitol 140:52–70. doi: 10.1016/j.exppara.2014.02.015. [DOI] [PubMed] [Google Scholar]
- 3.Reichel MP, Ayanegui-Alcerreca MA, Gondim LF, Ellis JT. 2013. What is the global economic impact of Neospora caninum in cattle—the billion dollar question. Int J Parasitol 43:133–142. doi: 10.1016/j.ijpara.2012.10.022. [DOI] [PubMed] [Google Scholar]
- 4.Hemphill A, Aguado-Martinez A, Müller J. 2016. Approaches for the vaccination and treatment of Neospora caninum infections in mice and ruminant models. Parasitology 143:245–259. doi: 10.1017/S0031182015001596. [DOI] [PubMed] [Google Scholar]
- 5.Horcajo P, Regidor-Cerrillo J, Aguado-Martinez A, Hemphill A, Ortega-Mora LM. 7 July 2016. Vaccines for bovine neosporosis: current status and key aspects for development. Parasite Immunol doi: 10.1111/pim.12342. [DOI] [PubMed] [Google Scholar]
- 6.Dubey JP, Schares G. 2011. Neosporosis in animals—the last five years. Vet Parasitol 180:90–108. doi: 10.1016/j.vetpar.2011.05.031. [DOI] [PubMed] [Google Scholar]
- 7.Reichel MP, Ellis JT, Dubey JP. 2007. Neosporosis and hammondiosis in dogs. J Small Anim Pract 48:308–312. doi: 10.1111/j.1748-5827.2006.00236.x. [DOI] [PubMed] [Google Scholar]
- 8.Rotella DP. 2012. Recent results in protein kinase inhibition for tropical diseases. Bioorg Med Chem Lett 22:6788–6793. doi: 10.1016/j.bmcl.2012.09.044. [DOI] [PubMed] [Google Scholar]
- 9.Lourido S, Shuman J, Zhang C, Shokat KM, Hui R, Sibley LD. 2010. Calcium-dependent protein kinase 1 is an essential regulator of exocytosis in Toxoplasma. Nature 465:359–362. doi: 10.1038/nature09022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kieschnick H, Wakefield T, Narducci CA, Beckers C. 2001. Toxoplasma gondii attachment to host cells is regulated by a calmodulin-like domain protein kinase. J Biol Chem 276:12369–12377. doi: 10.1074/jbc.M011045200. [DOI] [PubMed] [Google Scholar]
- 11.Ojo KK, Larson ET, Keyloun KR, Castaneda LJ, Derocher AE, Inampudi KK, Kim JE, Arakaki TL, Murphy RC, Zhang L, Napuli AJ, Maly DJ, Verlinde CL, Buckner FS, Parsons M, Hol WG, Merritt EA, Van Voorhis WC. 2010. Toxoplasma gondii calcium-dependent protein kinase 1 is a target for selective kinase inhibitors. Nat Struct Mol Biol 17:602–607. doi: 10.1038/nsmb.1818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Murphy RC, Ojo KK, Larson ET, Castellanos-Gonzalez A, Perera BG, Keyloun KR, Kim JE, Bhandari JG, Muller NR, Verlinde CL, White AC Jr, Merritt EA, Van Voorhis WC, Maly DJ. 2010. Discovery of potent and selective inhibitors of calcium-dependent protein kinase 1 (CDPK1) from C. parvum and T. gondii. ACS Med Chem Lett 1:331–335. doi: 10.1021/ml100096t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Keyloun KR, Reid MC, Choi R, Song Y, Fox AM, Hillesland HK, Zhang Z, Vidadala R, Merritt EA, Lau AO, Maly DJ, Fan E, Barrett LK, Van Voorhis WC, Ojo KK. 2014. The gatekeeper residue and beyond: homologous calcium-dependent protein kinases as drug development targets for veterinarian Apicomplexa parasites. Parasitology 141:1499–1509. doi: 10.1017/S0031182014000857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Castellanos-Gonzalez A, White AC Jr, Ojo KK, Vidadala RS, Zhang Z, Reid MC, Fox AM, Keyloun KR, Rivas K, Irani A, Dann SM, Fan E, Maly DJ, Van Voorhis WC. 2013. A novel calcium-dependent protein kinase inhibitor as a lead compound for treating cryptosporidiosis. J Infect Dis 208:1342–1348. doi: 10.1093/infdis/jit327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Doggett JS, Ojo KK, Fan E, Maly DJ, Van Voorhis WC. 2014. Bumped kinase inhibitor 1294 treats established Toxoplasma gondii infection. Antimicrob Agents Chemother 58:3547–3549. doi: 10.1128/AAC.01823-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ojo KK, Reid MC, Siddaramaiah LK, Müller J, Winzer P, Zhang Z, Keyloun KR, Vidadala RS, Merritt EA, Hol WG, Maly DJ, Fan E, Van Voorhis WC, Hemphill A. 2014. Neospora caninum calcium-dependent protein kinase 1 is an effective drug target for neosporosis therapy. PLoS One 9:e92929. doi: 10.1371/journal.pone.0092929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Winzer P, Müller J, Aguado-Martínez A, Rahman M, Balmer V, Ortega-Mora L, Ojo KK, Fan E, Maly D, Van Voorhis WC, Hemphill A. 2015. In vitro and in vivo effects of the bumped kinase inhibitor 1294 in the related cyst-forming apicomplexans Toxoplasma gondii and Neospora caninum. Antimicrob Agents Chemother 59:6361–6374. doi: 10.1128/AAC.01236-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vidadala RS, Rivas KL, Ojo KK, Hulverson MA, Zambriski JA, Bruzual I, Schultz TL, Huang W, Zhang Z, Scheele S, DeRocher AE, Choi R, Barrett LK, Siddaramaiah LK, Hol WG, Fan E, Merritt EA, Parsons M, Freiberg G, Marsh K, Kempf DJ, Carruthers VB, Isoherranen N, Doggett JS, Van Voorhis WC, Maly DJ. 2016. Development of an orally available and central nervous system (CNS) penetrant Toxoplasma gondii calcium-dependent protein kinase 1 (TgCDPK1) inhibitor with minimal human ether-a-go-go-related gene (hERG) activity for the treatment of toxoplasmosis. J Med Chem 59:6531–6546. doi: 10.1021/acs.jmedchem.6b00760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Huang W, Ojo KK, Zhang Z, Rivas K, Vidadala RS, Scheele S, DeRocher AE, Choi R, Hulverson MA, Barrett LK, Bruzual I, Siddaramaiah LK, Kerchner KM, Kurnick MD, Freiberg GM, Kempf D, Hol WG, Merritt EA, Neckermann G, de Hostos EL, Isoherranen N, Maly DJ, Parsons M, Doggett JS, Van Voorhis WC, Fan E. 2015. SAR studies of 5-aminopyrazole-4-carboxamide analogues as potent and selective inhibitors of Toxoplasma gondii CDPK1. ACS Med Chem Lett 6:1184–1189. doi: 10.1021/acsmedchemlett.5b00319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Müller J, Balmer V, Winzer P, Rahman M, Manser V, Haynes RK, Hemphill A. 2015. In vitro effects of new artemisinin derivatives in Neospora caninum-infected human fibroblasts. Int J Antimicrob Agents 46:88–93. doi: 10.1016/j.ijantimicag.2015.02.020. [DOI] [PubMed] [Google Scholar]
- 21.Arranz-Solis D, Aguado-Martinez A, Müller J, Regidor-Cerrillo J, Ortega-Mora LM, Hemphill A. 2015. Dose-dependent effects of experimental infection with the virulent Neospora caninum Nc-Spain7 isolate in a pregnant mouse model. Vet Parasitol 211:133–140. doi: 10.1016/j.vetpar.2015.05.021. [DOI] [PubMed] [Google Scholar]
- 22.Regidor-Cerrillo J, Arranz-Solis D, Benavides J, Gomez-Bautista M, Castro-Hermida JA, Mezo M, Perez V, Ortega-Mora LM, Gonzalez-Warleta M. 2014. Neospora caninum infection during early pregnancy in cattle: how the isolate influences infection dynamics, clinical outcome and peripheral and local immune responses. Vet Res 45:10. doi: 10.1186/1297-9716-45-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Arranz-Solis D, Benavides J, Regidor-Cerrillo J, Fuertes M, Ferre I, Ferreras MDC, Collantes-Fernandez E, Hemphill A, Perez V, Ortega-Mora LM. 2015. Influence of the gestational stage on the clinical course, lesional development and parasite distribution in experimental ovine neosporosis. Vet Res 46:19. doi: 10.1186/s13567-014-0139-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Müller J, Aguado-Martinez A, Manser V, Balmer V, Winzer P, Ritler D, Hostettler I, Solís D, Ortega-Mora LM, Hemphill A. 2015. Buparvaquone is active against Neospora caninum in vitro and in experimentally infected mice. Int J Parasitol Drugs Drug Resist 5:16–25. doi: 10.1016/j.ijpddr.2015.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Guionaud C, Hemphill A, Mevissen M, Alaeddine F. 2010. Molecular characterization of Neospora caninum MAG1, a dense granule protein secreted into the parasitophorous vacuole, and associated with the cyst wall and the cyst matrix. Parasitology 137:1605–1619. doi: 10.1017/S0031182010000442. [DOI] [PubMed] [Google Scholar]
- 26.Berglund F, Flodh H, Lundborg P, Prame B, Sannerstedt R. 1984. Drug use during pregnancy and breast-feeding. A classification system for drug information. Acta Obstet Gynecol Scand Suppl 126:1–55. [DOI] [PubMed] [Google Scholar]
- 27.Taylor WRJ, White NJ. 2004. Antimalarial drug toxicity: a review. Drug Saf 27:25–61. doi: 10.2165/00002018-200427010-00003. [DOI] [PubMed] [Google Scholar]
- 28.Manyando C, Kayentao K, D'Alessandro U, Okafor HU, Juma E, Hamed K. 2012. A systematic review of the safety and efficacy of artemether-lumefantrine against uncomplicated Plasmodium falciparum malaria during pregnancy. Malar J 11:141. doi: 10.1186/1475-2875-11-141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Mazuz ML, Fish L, Wolkomirsky R, Leibovich B, Reznikov D, Savitsky I, Golenser J, Shkap V. 2015. The effect of a live Neospora caninum tachyzoite vaccine in naturally infected pregnant dairy cows. Prev Vet Med 120:232–235. doi: 10.1016/j.prevetmed.2015.03.020. [DOI] [PubMed] [Google Scholar]
- 30.Schorer M, Debache K, Barna F, Monney T, Müller J, Boykin DW, Stephens CE, Hemphill A. 2012. Di-cationic arylimidamides act against Neospora caninum tachyzoites by interference in membrane structure and nucleolar integrity and are active against challenge infection in mice. Int J Parasitol Drugs Drug Resist 2:109–120. doi: 10.1016/j.ijpddr.2012.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Stadelmann B, Scholl S, Müller J, Hemphill A. 2010. Application of an in vitro drug screening assay based on the release of phosphoglucose isomerase to determine the structure-activity relationship of thiazolides against Echinococcus multilocularis metacestodes. J Antimicrob Chemother 65:512–519. doi: 10.1093/jac/dkp490. [DOI] [PubMed] [Google Scholar]
- 32.Howe DK, Mercier C, Messina M, Sibley LD. 1997. Expression of Toxoplasma gondii genes in the closely-related apicomplexan parasite Neospora caninum. Mol Biochem Parasitol 86:29–36. doi: 10.1016/S0166-6851(97)90003-7. [DOI] [PubMed] [Google Scholar]
- 33.Alaeddine F, Hemphill A, Debache K, Guionaud C. 2013. Molecular cloning and characterization of NcROP2Fam-1, a member of the ROP2 family of rhoptry proteins in Neospora caninum that is targeted by antibodies neutralizing host cell invasion in vitro. Parasitology 140:1033–1050. doi: 10.1017/S0031182013000383. [DOI] [PubMed] [Google Scholar]
- 34.Bjorkman C, Hemphill A. 1998. Characterization of Neospora caninum iscom antigens using monoclonal antibodies. Parasite Immunol 20:73–80. doi: 10.1046/j.1365-3024.1998.00127.x. [DOI] [PubMed] [Google Scholar]
- 35.Debache K, Alaeddine F, Guionaud C, Monney T, Müller J, Strohbusch M, Leib SL, Grandgirard D, Hemphill A. 2009. Vaccination with recombinant NcROP2 combined with recombinant NcMIC1 and NcMIC3 reduces cerebral infection and vertical transmission in mice experimentally infected with Neospora caninum tachyzoites. Int J Parasitol 39:1373–1384. doi: 10.1016/j.ijpara.2009.04.006. [DOI] [PubMed] [Google Scholar]
- 36.Debache K, Guionaud C, Alaeddine F, Mevissen M, Hemphill A. 2008. Vaccination of mice with recombinant NcROP2 antigen reduces mortality and cerebral infection in mice infected with Neospora caninum tachyzoites. Int J Parasitol 38:1455–1463. doi: 10.1016/j.ijpara.2008.04.001. [DOI] [PubMed] [Google Scholar]
- 37.R Core Team. 2012. R: a language and environment for statistical computing. R Core Team, Vienna, Austria. [Google Scholar]