

Abstract
Sepsis causes significant mortality in neonatal foals; however, there is little data describing the cellular and molecular pathways of lung inflammation in septic foals. This study was conducted to characterize lung inflammation in septic foals. Lung tissue sections from control (n = 6) and septic (n = 17) foals were compared using histology and immunohistology. Blinded pathologic scoring of hematoxylin and eosin stained samples revealed increased features of lung inflammation such as thickened alveolar septa and sequestered inflammatory cells in septic foals. Septic foal lungs showed increased expression of von Willebrand factor in blood vessels, demonstrating vascular inflammation. Use of MAC387 antibody to detect calprotectin as a reflection of mononuclear cell infiltration revealed a significant increase in their numbers in alveolar septa of lungs from septic foals compared to those from control foals. The mononuclear cells appeared to be mature macrophages and were located in the septal capillaries, suggesting they were pulmonary intravascular macrophages (PIMs). Finally, lungs from septic foals showed increased expression of Toll-like receptor 4 and 9 in mononuclear cells relative to the control. Taken together, this study is the first to show the expression of inflammatory molecules and an increase in PIMs in lungs from foals that died from sepsis.
Keywords: Toll-like receptor 4 and 9, horses, immunohistochemistry, pulmonary intravascular macrophage, von Willebrand factor
Introduction
Foals, especially those with failure of passive transfer of maternal antibodies, are highly susceptible to bacterial infections and endotoxemia; however, the mechanisms underlying such susceptibilities are not fully understood [19,25]. Non-specific intestinal absorption, hypoglycaemia, omphalophlebitis, unsanitary environmental conditions, and abnormal gestational lengths are common factors that predispose foals to further complications following birth, including septicaemia [16,20,25]. Sepsis is a systemic bacterial invasion that commonly results in systemic inflammatory response syndrome, which can lead to numerous sequelae such as coagulopathies and immune system dysfunction [25]. Indeed, septic shock can result in hypotension, organ hypoperfusion, and multiple organ dysfunction syndrome, which may lead to death of foals, especially neonates [19,20]. Mortality rates of septic neonatal foals generally range from 40 to 55% [25].
The majority of research on human sepsis and its effects on lung inflammation has focused on acute lung injury and/or acute respiratory distress syndrome [4,30]. Indeed there is very little data available regarding lung injury and inflammation in foals suffering from sepsis [16,18,20,25]. Because of the increased awareness of acute lung injury in veterinary species, a consensus statement was released to establish a veterinary specific definition [29]. The defining criteria for acute lung injury in veterinary patients, including foals, are similar to those used in human medicine and include acute onset, known association with systemic inflammation or sepsis, signs of pulmonary capillary leakage in the absence of pulmonary hypertension, and evidence of inefficient gas exchange or hypoxemia. A fifth optional factor is evidence of diffuse pulmonary inflammation seen on radiographs [29]. While septicaemic foals are commonly diagnosed with pulmonary inflammation, most clinical attention has focused on sequelae other than lung inflammation, such as haemostasis, arthritis, or organ dysfunction involving mainly the liver, spleen, and gastrointestinal tract [19,25,26]. Previously, pneumonia was diagnosed either clinically or at post-mortem in 28 to 50% of septic foals [20]. Interestingly, necropsy data have highlighted lung pathologies in septic foals, even when little or no ante-mortem pulmonary signs were observed [18,25]. Regardless of these findings, precise descriptions and information describing the mechanisms of lung inflammation in septic foals are lacking.
Fortunately, studies of other animal species and humans have provided a general mechanistic framework for systemic inflammatory response syndrome and its effects on the mammalian lung. This dysregulated inflammatory response involves activation of innate immune cells such as macrophages and neutrophils through signalling via various pattern recognition receptors, including toll-like receptors (TLRs), which engage and identify conserved molecular patterns of bacteria and other danger signals [15]. For example, Toll-like receptor (TLR) 4 recognizes lipopolysaccharide, while TLR9 recognizes cytosine guanine phosphodiester DNA patterns, which are most common to gram-negative and gram-positive bacteria, respectively [15]. The recognition of pathogenic material by immune cells leads to the release of pro-inflammatory cytokines and chemokines, which induce adhesion phenotype changes in epithelial and endothelial cells to activate the recruitment of neutrophils [13,15]. Neutrophils secrete many cytotoxic molecules, such as free oxygen radicals, which cause cellular damage and increase vascular permeability [5]. In addition, circulating endotoxin promotes activation of the coagulation cascade via tissue factor production [7]. In conjunction, there is increased expression of a glycoprotein known as von Willebrand factor (vWF) on endothelial cells and platelets that promotes cell-endothelial adhesion and platelet activation [14]. The acquisition of a pro-coagulant phenotype by the endothelium may eventually lead to disseminated intravascular coagulation and potentially irreversible vascular damage [19]. However, no molecular characterizations of lung inflammation in septic foals have been conducted to date.
This study was conducted to compare lung tissue from septic and healthy control foals using histological and immunohistological techniques. Through this examination, we tested the hypothesis that lung inflammation is present in septic foals.
Materials and Methods
Tissue samples
The protocols for this study were approved by the Animal Care Assurance Committee of the University of Saskatchewan in accordance with the national guidelines for the use of animals in research. A medical record search was performed to identify foals (n = 17) that died from complications of sepsis and had a complete necropsy preformed, including the collection of pulmonary tissue for histological examination. Information regarding foals was taken from medical records. Clinical diagnosis of sepsis involved analysis of foal history, complete blood count, IgG levels, and evidence of pulmonary embolism. In this study, foals were classified using published septic scoring techniques, with foals considered positive for sepsis if they met any or all of the following criteria: typical clinical characteristics of sepsis, such as multiple sites of infections, positive bacterial cultures from blood smears, and necropsy data highlighting the widespread damage characteristic of septicaemia [2,28]. Tissues were fixed in 10% buffered formalin, embedded in paraffin, and sectioned at 5 µm thickness with a microtome. Lung tissue samples were acquired from normal foals (n = 6) that were used in an previous study by Johnson et al. [12]. These normal foals were delivered normally in a contained facility and found to be normal on clinical examination and complete blood count analyses. These foals received no treatment prior to euthanasia. Briefly, foals were placed in a supine position and the lungs were fixed by intrabronchial instillation of glutaraldehyde/paraformaldehyde fixative. Lung tissue was then randomly sampled using a fractionator design, embedded in paraffin, and sectioned at 5 µm thickness using a microtome [12].
Histopathological analysis
Multiple sections of lung from each foal were stained with hematoxylin and eosin (H&E). One pathologist who was blinded to the identity of the foals examined all H&E stained microscopic lung sections and qualitatively scored them for levels of inflammation. Specifically, lung sections were assessed for the degree of edema, septal inflammation, and alveolar inflammation. Each of these categories was given a score based on severity (0, absent; 1, mild; 2, moderate; 3, severe). An overall score was determined for each lung section based on the sum of the values of the three categories.
Immunohistochemistry
Immunohistochemistry was performed on serial lung sections. Briefly, the sections were incubated with 0.5% hydrogen peroxide in methanol (Sigma-Aldrich, Canada) to quench endogenous peroxidase activity. This was followed by exposure to pepsin (2 mg/mL pepsin in 0.01N HCl, 60 min at room temperature; Sigma-Aldrich) to unmask antigens, and incubation with bovine serum albumin (1% in phosphate buffered saline) for 30 min. Incubation with primary antibodies (Table 1) was performed for 12 h at 4℃ before treatment with horseradish peroxidase-conjugated secondary antibodies at room temperature for 30 min. Color development was conducted using a peroxidase kit (DAB Peroxidase Substrate Kit, Catalog No. SK-4100; Vector Laboratories, Canada), and the sections were counterstained with Methyl green (Vector Laboratories). The negative controls included omission of either the primary or secondary antibody. Each primary antibody was initially titrated to determine the optimal concentration for use in our experiments.
Table 1. Primary antibodies used for immunohistological characterization of the control (n = 6) and septic (n = 17) foal lungs.

vWF, von Willebrand factor.
Analysis of immunohistochemistry
Mononuclear cells stained with MAC387, which recognizes calprotectin, were counted by an observer blinded to treatment in 10 fields at 60× magnification in 3–4 different lung sections per horse. The number of sections was determined by those collected at necropsy. The median value of cells per section was determined for the counts from the septic (n = 17) and control (n = 6) lungs for better comparison. The remaining antibodies (vWF, TLR4, and TLR9) were assessed via light microscopy to determine the presence or absence of staining in various areas of 3–4 different lung sections from each of the control and septic foals. The areas that were assessed included large blood vessel endothelium, alveolar septa, airway epithelium, and mononuclear cells. Positive staining was given a value of 1 and no staining a value of 0. For each area assessed, the number of foal lung sections that stained positive for antibody was calculated as a percentage of the total lung sections extracted from their respective septic or control foal group.
Data analysis was conducted using commercial software (GraphPad Prism 6; GraphPad Software, USA). For all comparisons, significance was assessed at a p value of < 0.05. A Mann-Whitney U test was used to compare histopathological scores, vWF scores, and MAC387 positive cell counts between groups.
Results
Clinical diagnosis of sepsis
The foals were selected for inclusion in this study based on clinical diagnosis of septicaemia along with a complete necropsy. The septic foal time of death after presentation of illness ranged from 6 hours to 8 days. Unfortunately, medical records regarding the origin of sepsis in some of the foals were incomplete. However, 10 of the 17 foals (58%) showed positive results for bacterial cultures (Table 2). It is well known that blood cultures only result in a definitive diagnosis in approximately 50% of cases, with many clinicians using additional septic symptoms as supporting evidence of their diagnosis [10]. Since over 50% of the foals we characterized as septic reflected positive bacterial cultures, we are confident that our classification of sepsis is accurate.
Table 2. Bacterial isolates from foals characterized as septic.

NA, not available.
Histopathology
Lung sections from control foals showed normal histomorphology with thin alveolar septa, few septal cells, and a lack of cells within alveolar spaces (panels A and C in Fig. 1). In contrast, the lungs from all of the septic foals showed septal cellular infiltration, dilated septal vasculature, thickened alveolar septa, and an overall disordered microanatomy (panels B and D in Fig. 1). The majority of cells stained by H&E were mononuclear, while there were very few cells with segmented nuclei, such as neutrophils. The morphology of alveoli, particularly the lack of inflammatory cells contained within them, did not differ between septic foals and control foals.
Fig. 1. Histology of hematoxylin and eosin stained lung sections from control (A and C) and septic (B and D) foals. Lung section from the control foal (A) shows normal lung morphology of bronchiole (†), blood vessels (BV), and alveolar space (*) compared to increased septal thickness and cellularity (arrows) in the lung section from the septic foal (B). Lung section from the control foal (C) shows normal alveolar septal width (double headed arrow) compared to the thickened septa of the septic foal lung section (D). Scale bars = 200 µm (A and B), 20 µm (C), 10 µm (D).
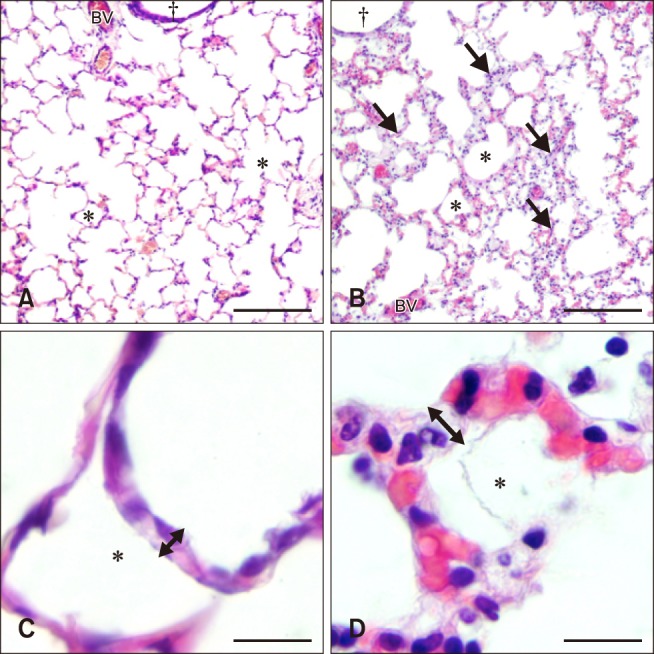
While the lungs of the control foals showed no signs of inflammation across any of the three categories, 47% of the septic foal lungs showed mild edema and 24% showed moderate to severe edema. Furthermore, 59% of the septic foal lungs showed mild levels of septal inflammation. Overall, the lungs from septic foals showed higher scores for edema (median, 1; range, 0–3) and septal inflammation (median, 1; range, 0–1) than those from the control foals (edema: p = 0.007; septal inflammation: p = 0.02); however, there was no difference in alveolar inflammation between groups, with the median value of the septic foals also being zero (p = 0.2). Each control foal received a total inflammation score of 0, while lung sections from septic foals had a median score of 2 (range, 0–6). There was a significant difference in total inflammation score between normal and septic foal lung sections (p = 0.0013).
vWF antibody staining
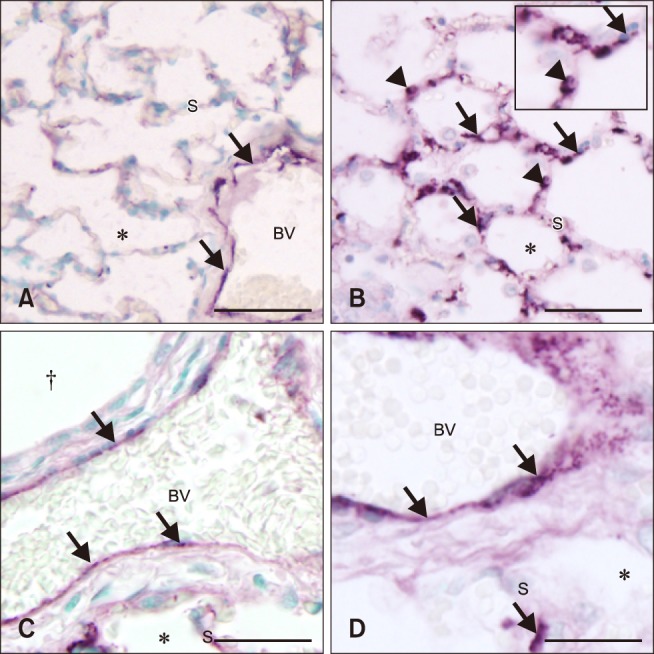
We used the intensity of vWF staining as a marker of activation of endothelial cells. The histologic sections from all control and septic foal lungs showed staining along the endothelium of all large blood vessels (panels A, C and D in Fig. 2). Although the septic foal lung sections appeared to have increased vWF staining in the septal vasculature (panels B and D in Fig. 2) compared to the sections from the control foals (control, 0%; septic, 50%), this difference was not significant (p = 0.058). However, vWF staining was significantly more prevalent on mononuclear cells in lung sections from septic foals than on mononuclear cells found in the control foal lung sections (control, 8%; septic, 59%; p = 0.0183) (panels A and B in Fig. 2).
Fig. 2. Histologic lung sections from control (A and C) and septic (B and D) foals. Von Willebrand factor (vWF) staining (arrows) was present in the endothelium of large blood vessel (BV), but was minimal in alveolar septa (S) in sections from the control foal lung (A and C). The sections from septic foals show increased staining for vWF in alveolar septa and the endothelium in large BVs (B and D). The septic foals (B) also showed an increase in positive vWF staining on mononuclear cells (arrowheads) compared to normal foal lungs. *Alveolar space. †Bronchiole. Scale bars = 100 µm (A and B), 50 µm (C and D).
MAC387 antibody staining
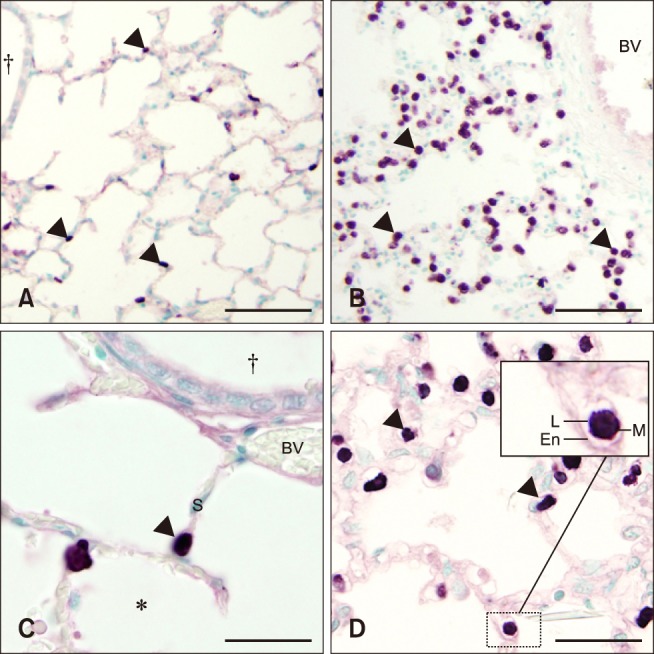
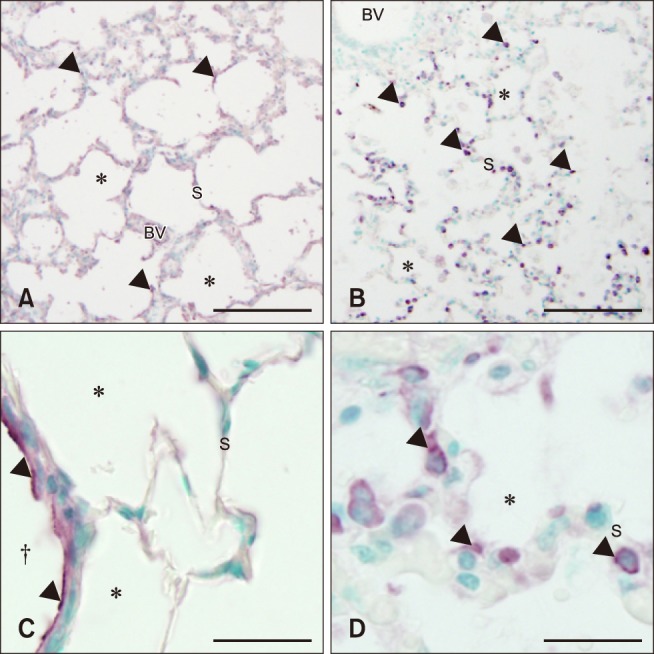
Control foal lung sections had very few septal cells stained with MAC387 antibody (panels A and C in Fig. 3). Lungs from septic foals showed a significant increase in cells positive for MAC387 compared to control foals [control, 12 median cells/10 fields 60× magnification (range, 3–21); septic, 38 median cells/10 fields 60× magnification (range, 21–96); p < 0.001)] (panel B in Fig. 3). Further evaluation of lung sections under higher magnification (100×) showed that MAC387 positive cells were commonly located in the intravascular space, consistent with the location for pulmonary intravascular macrophages (PIMs) (panel D in Fig. 3).
Fig. 3. Histologic lung sections from control (A and C) and septic (B and D) foals with MAC387 staining. Mononuclear cell (arrowheads) infiltration increased in septic (B and D) vs. control (A and C) foals. The inset image in figure d shows macrophage-like cells within the vasculature of the alveolar septa (S), suggesting the presence of a pulmonary intravascular macrophages. BV, blood vessel; En, endothelial cell; L, lumen; M, macrophage. *Alveolar space. †Bronchiole. Scale bars = 200 µm (A and B), 50 µm (C and D).
TLR4 antibody staining
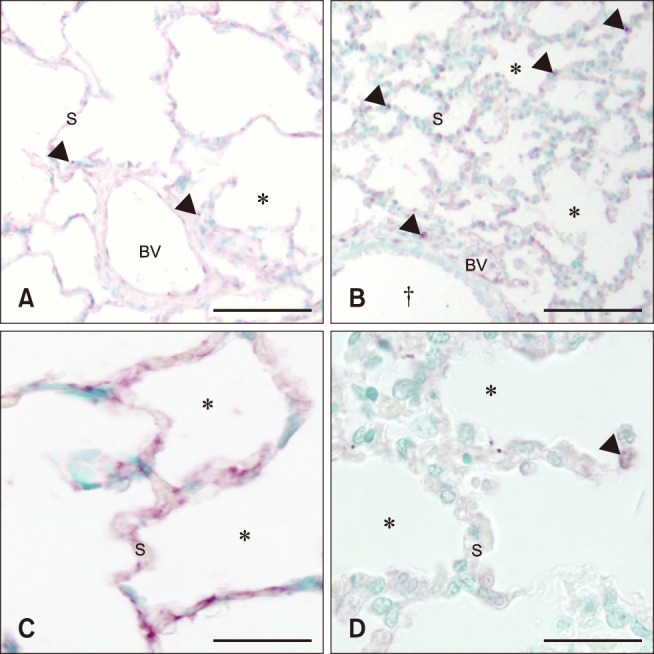
The proportion of TLR4 staining on airway epithelium (control, 67%; septic, 29%), large blood vessel endothelium (control, 89%; septic, 59%), and alveolar septa (control, 50%; septic, 35%) was much lower in the septic foal lung sections than the sections from the control foal lungs (Fig. 4). However, there was increased TLR4 labeling on mononuclear cells in lung sections from septic foals compared to the lung sections of the control foals (control, 8%; septic, 26%) (panels C and D in Fig. 4).
Fig. 4. Histologic lung sections from control (A and C) and septic (B and D) foals. Septic foals (B and D) showed an increase in TLR4 staining (arrowheads) on mononuclear cells, but a slight decrease along bronchiole (†) epithelium, blood vessel (BV) endothelium, and alveolar septa (S). *Alveolar space. Scale bars = 100 µm (A and B), 50 µm (C and D).
TLR9 antibody staining
Similar to the pattern observed for TLR4 staining, there were lower proportions of lung sections from septic foals that showed TLR9 on airway epithelium (control, 100%; septic, 71%), large blood vessel endothelium (control, 67%; septic, 12%), and alveolar septa (control, 67%; septic, 0%) compared to the lungs of the control foals (Fig. 5). However, similar to TLR4 staining, there was increased TLR9 labeling in mononuclear cells in lung sections from septic foals compared to the lung sections from control foals (control, 72%; septic, 82%) (panels D and D in Fig. 5).
Fig. 5. Histologic lung sections from control (A and C) and septic (B and D) foals. Septic foals showed an increase in TLR9 staining (arrowheads) on mononuclear cells, but a slight decrease along bronchiole (†) epithelium, blood vessel (BV) endothelium, and alveolar septa (S). *Alveolar space. Scale bars = 100 µm (A and B), 50 µm (C and D).
Discussion
Although pulmonary pathology is a common sequela to septicaemia in humans and other veterinary species, there is still very little data available regarding the occurrence and characterization of lung inflammation in septic foals. Here, we report the first data to show vascular inflammation, an increase in septal monocytes and macrophages, and increased expression of TLR4 and TLR9 on mononuclear cells in the lungs of septic foals. These data will help improve our understanding of pulmonary complications found in foals experiencing septicaemia.
While the medical records regarding the origin of sepsis in some of the foals were incomplete, the majority of foals had sepsis originating remote from the lung. Therefore, it is likely that foals experience lung inflammation as a sequel to sepsis, similar to humans, as has been previously reported [6]. Based on these findings, foals should be evaluated and treated for pulmonary complications associated with non-pulmonary origin septicaemia.
We observed lung inflammation based on increased migration of cells in the alveolar septa, thickening of the alveolar septa, and through semi-quantitative scoring of the lung sections for MAC387 positive cell infiltration. Lung inflammation is characterized by recruitment of inflammatory cells, such as neutrophils and monocytes, and by increased vascular permeability [23,27]. The recruitment of cells such as platelets, neutrophils, and monocytes is facilitated through activation of the endothelium by cytokines leading to the expression of adhesion molecules. vWF, an adhesive and coagulation protein, is stored in endothelial cytoplasmic Weibel-Palade bodies, which are exocytosed upon endothelial cell activation [14]. The recruitment of platelets, which is facilitated by vWF endothelial expression and other proteins such as selectins, is considered one of the earliest events in acute inflammation [13]. Here, we report the expression of vWF in alveolar septa in lung sections from foals with septicaemia, where it is normally not expressed. It is generally believed that lung microvascular endothelium lacks Weibel-Palade bodies that store preformed vWF. Therefore, the vWF expression in the alveolar septa could be due to accumulation of vWF-containing platelets, or the release and spread of vWF from the endothelium in larger blood vessels, which normally express this protein, into the alveolar capillaries. The increased expression of vWF and accumulation of septal mononuclear cells may suggest a role of vWF in their recruitment. Nevertheless, the increased expression of vWF provides molecular evidence of microvascular inflammation, including a coagulative phenotype of the endothelium in the lungs of septic foals.
Our immunohistochemical data demonstrated mostly monocytes and macrophages in the alveolar septa of septic foals. We previously used calprotectin-binding antibody, MAC387, to identify equine lung macrophages with light and electron microscopy [17,21]. MAC387 can also stain neutrophils; however, we eliminated neutrophils from our counts based on their nuclear morphology. Moreover, we found very few neutrophils in the septa of normal or septic foals. The presence of large numbers of septal monocytes and macrophages, but fewer neutrophils, in the lungs of septic foals is intriguing. In animal species such as the mouse, endotoxin-induced lung inflammation results in initial recruitment of neutrophils followed by infiltration of monocytes. The newly arriving monocytes are known to express anti-inflammatory cytokines such as transforming growth factor beta and interleukin-10 to hasten resolution of inflammation [3]. However, there is evidence that newly recruited intravascular monocytes/macrophages increase the risk of pulmonary inflammation in response to secondary endotoxin challenges [8,9,24]. Preliminary electron microscopic observations of tissues derived from paraffin sections also suggest there are numerous monocytes and macrophages located in the septal capillaries (B. Singh et al., unpublished data). Interestingly, the normal foals in our study had very few intravascular/septal mononuclear cells that stained positive for MAC387, which is similar to the findings from the day old pig [31]. These findings indicate that, similar to pigs, foals may also recruit pulmonary intravascular mononuclear cells such as PIMs during their post-natal life, which may increase host susceptibility for lung inflammation and injury [1,17].
Our study does not address the function of newly recruited septal monocytes/macrophages, but we speculate that the newly recruited macrophages are pro-inflammatory. This speculation is based on our data showing expression of TLR4 and TLR9 on septal mononuclear cells, but not on airway epithelium and endothelium in lungs from septic foals. It is also consistent with previous results showing reduced TLR4 staining in airways and large blood vessels during pulmonary inflammation with increased TLR4 expression in mononuclear cells [27]. However, data describing pulmonary monocytes in rats suggest that after exposure to pro-inflammatory endotoxins, TLR4 expression is initially increased, but then decreases significantly 36 hours after exposure [11]. Previous data has shown increased TLR9 expression on both PIMs and epithelial/endothelial surfaces of adult horses treated with LPS [22]. Since sepsis in foals is most often caused by gram-negative bacteria and their associated endotoxins [26], the increased numbers of septal mononuclear cells expressing TLR4 and TLR9 may lead to more pronounced inflammation in response to circulating endotoxins and other bacterial molecules. Therefore, we cannot concretely attribute the increased staining of TLR4/TLR9 to the recruitment of PIMs in the septic foal. However, it is possible that increased expression of vWF may have contributed to the increased numbers of monocytes and macrophages in lung septa, which may have led to an appearance of increased expression of TLR4 and TLR9. Further data using transmission electron microscopy and immune-gold TLR labeling should be conducted to improve our understanding of lung inflammation in septic foals.
These data, which are the first to characterize inflammation in the lungs of septic foals, show histological inflammation including thickened alveolar septa, increased septal mononuclear cells expressing TLR4 and TLR9, and signs of microvascular inflammation. These cellular and molecular changes in lungs of septic foals suggest association of lung injury with sepsis. Since significant recruitment of septal monocytes and macrophages, most likely PIMs, may increase lung injury, further study is required to better understand their function.
Acknowledgments
This study was funded through a grant from the Ryan-Dube Animal Health Fund. Stacy Anderson was supported by an Interprovincial Graduate Fellowship and Jacqueline Harrison, Alexandra Robinson, and Leah Quanstrom were supported through Interprovincial Undergraduate Student Summer Research Scholarships.
Footnotes
Conflict of Interest: The authors declare no conflicts of interest.
References
- 1.Atwal OS. Pulmonary intravascular macrophages (PIMs) and cationised ferritin-induced phagocytosis of platelets: a morphological study. Comp Haematol Int. 1992;2:179–185. [Google Scholar]
- 2.Brewer BD, Koterba AM. Development of a scoring system for the early diagnosis of equine neonatal sepsis. Equine Vet J. 1988;20:18–22. doi: 10.1111/j.2042-3306.1988.tb01445.x. [DOI] [PubMed] [Google Scholar]
- 3.Charavaryamath C, Janardhan KS, Caldwell S, Singh B. Pulmonary intravascular monocytes/macrophages in a rat model of sepsis. Anat Rec A Discov Mol Cell Evol Biol. 2006;288:1259–1271. doi: 10.1002/ar.a.20401. [DOI] [PubMed] [Google Scholar]
- 4.Cunningham AJ. Acute respiratory distress syndrome—two decades later. Yale J Biol Med. 1991;64:387–402. [PMC free article] [PubMed] [Google Scholar]
- 5.DiStasi MR, Ley K. Opening the flood-gates: how neutrophil-endothelial interactions regulate permeability. Trends Immunol. 2009;30:547–556. doi: 10.1016/j.it.2009.07.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dunkel B, Dolente B, Boston RC. Acute lung injury/acute respiratory distress syndrome in 15 foals. Equine Vet J. 2005;37:435–440. doi: 10.2746/042516405774480094. [DOI] [PubMed] [Google Scholar]
- 7.Esmon CT, Xu J, Lupu F. Innate immunity and coagulation. J Thromb Haemost. 2011;9(Suppl 1):182–188. doi: 10.1111/j.1538-7836.2011.04323.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Frevert CW, Warner AE. Respiratory distress resulting from acute lung injury in the veterinary patient. J Vet Intern Med. 1992;6:154–165. doi: 10.1111/j.1939-1676.1992.tb00331.x. [DOI] [PubMed] [Google Scholar]
- 9.Gill SS, Suri SS, Janardhan KS, Caldwell S, Duke T, Singh B. Role of pulmonary intravascular macrophages in endotoxin-induced lung inflammation and mortality in a rat model. Respir Res. 2008;9:69. doi: 10.1186/1465-9921-9-69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hall KK, Lyman JA. Updated review of blood culture contamination. Clin Microbiol Rev. 2006;19:788–802. doi: 10.1128/CMR.00062-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Janardhan KS, McIsaac M, Fowlie J, Shrivastav A, Caldwell S, Sharma RK, Singh B. Toll like receptor-4 expression in lipopolysaccharide induced lung inflammation. Histol Histopathol. 2006;21:687–696. doi: 10.14670/HH-21.687. [DOI] [PubMed] [Google Scholar]
- 12.Johnson L, Montgomery JB, Schneider JP, Townsend HGG, Ochs M, Singh B. Morphometric examination of the equine adult and foal lung. Anat Rec (Hoboken) 2014;297:1950–1962. doi: 10.1002/ar.22994. [DOI] [PubMed] [Google Scholar]
- 13.Kolaczkowska E, Kubes P. Neutrophil recruitment and function in health and inflammation. Nat Rev Immunol. 2013;13:159–175. doi: 10.1038/nri3399. [DOI] [PubMed] [Google Scholar]
- 14.Lenting PJ, Christophe OD, Denis CV. von Willebrand factor biosynthesis, secretion, and clearance: connecting the far ends. Blood. 2015;125:2019–2028. doi: 10.1182/blood-2014-06-528406. [DOI] [PubMed] [Google Scholar]
- 15.Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev. 2009;22:240–273. doi: 10.1128/CMR.00046-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Morris DD. Endotoxemia in horses: a review of cellular and humoral mediators involved in its pathogenesis. J Vet Intern Med. 1991;5:167–181. doi: 10.1111/j.1939-1676.1991.tb00944.x. [DOI] [PubMed] [Google Scholar]
- 17.Parbhakar OP, Duke T, Townsend HGG, Singh B. Depletion of pulmonary intravascular macrophages partially inhibits lipopolysaccharide-induced lung inflammation in horses. Vet Res. 2005;36:557–569. doi: 10.1051/vetres:2005016. [DOI] [PubMed] [Google Scholar]
- 18.Raisis AL, Hodgson JL, Hodgson DR. Equine neonatal septicaemia: 24 cases. Aust Vet J. 1996;73:137–140. doi: 10.1111/j.1751-0813.1996.tb10006.x. [DOI] [PubMed] [Google Scholar]
- 19.Roy MF. Sepsis in adults and foals. Vet Clin North Am Equine Pract. 2004;20:41–61. doi: 10.1016/j.cveq.2003.12.005. [DOI] [PubMed] [Google Scholar]
- 20.Sanchez LC. Equine neonatal sepsis. Vet Clin North Am Equine Pract. 2005;21:273–293. doi: 10.1016/j.cveq.2005.04.007. [DOI] [PubMed] [Google Scholar]
- 21.Schneberger D, Aharonson-Raz K, Singh B. Pulmonary intravascular macrophages and lung health: what are we missing? Am J Physiol Lung Cell Mol Physiol. 2012;302:L498–L503. doi: 10.1152/ajplung.00322.2011. [DOI] [PubMed] [Google Scholar]
- 22.Schneberger D, Caldwell S, Suri SS, Singh B. Expression of Toll-like receptor 9 in horse lungs. Anat Rec (Hoboken) 2009;292:1068–1077. doi: 10.1002/ar.20927. [DOI] [PubMed] [Google Scholar]
- 23.Shrum B, Anantha RV, Xu SX, Donnelly M, Haeryfar SM, McCormick JK, Mele T. A robust scoring system to evaluate sepsis severity in an animal model. BMC Res Notes. 2014;7:233. doi: 10.1186/1756-0500-7-233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Singh B, Pearce JW, Gamage LN, Janardhan K, Caldwell S. Depletion of pulmonary intravascular macrophages inhibits acute lung inflammation. Am J Physiol Lung Cell Mol Physiol. 2004;286:L363–L372. doi: 10.1152/ajplung.00003.2003. [DOI] [PubMed] [Google Scholar]
- 25.Taylor S. A review of equine sepsis. Equine Vet Educ. 2015;27:99–109. doi: 10.1111/eve.12290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Theelen MJP, Wilson WD, Edman JM, Magdesian KG, Kass PH. Temporal trends in prevalence of bacteria isolated from foals with sepsis: 1979-2010. Equine Vet J. 2014;46:169–173. doi: 10.1111/evj.12131. [DOI] [PubMed] [Google Scholar]
- 27.Wassef A, Janardhan K, Pearce JW, Singh B. Toll-like receptor 4 in normal and inflamed lungs and other organs of pig, dog and cattle. Histol Histopathol. 2004;19:1201–1208. doi: 10.14670/HH-19.1201. [DOI] [PubMed] [Google Scholar]
- 28.Weber EJ, Sanchez LC, Giguère S. Re-evaluation of the sepsis score in equine neonates. Equine Vet J. 2015;47:275–278. doi: 10.1111/evj.12279. [DOI] [PubMed] [Google Scholar]
- 29.Wilkins PA, Otto CM, Baumgardner JE, Dunkel B, Bedenice D, Paradis MR, Staffieri F, Syring RS, Slack J, Grasso S, Pranzo EG. Acute lung injury and acute respiratory distress syndromes in veterinary medicine: consensus definitions: The Dorothy Russell Havemeyer Working Group on ALI and ARDS in Veterinary Medicine. J Vet Emerg Crit Care. 2007;17:333–339. [Google Scholar]
- 30.Wilson RF, Sibbald WJ. Acute respiratory failure. Crit Care Med. 1976;4:79–89. doi: 10.1097/00003246-197603000-00007. [DOI] [PubMed] [Google Scholar]
- 31.Winkler GC. Pulmonary intravascular macrophages in domestic animal species: review of structural and functional properties. Am J Anat. 1988;181:217–234. doi: 10.1002/aja.1001810302. [DOI] [PubMed] [Google Scholar]