

Abstract
Early experience and social context interact to alter the phenotype of complex social behaviors. These early experiences can also result in alterations to cortical organization and connections. Given the ability of the neuropeptide oxytocin (OT) to modulate social and reproductive behavior, OT is likely involved in these cortical processes. However, little is known about the distribution of OT and OT receptors (OTR) within the neocortex. Using autoradiographic and neuroanatomical techniques, we characterized the cortical distribution of oxytocin receptors (OTR) in prairie voles, a socially monogamous rodent species. We found that OTR density was low in the primary sensory areas (including primary somatosensory and auditory regions) but was quite high in association regions (including temporal and parietal association areas, and prelimbic regions). In the primary motor area as well as the temporal and parietal association areas, we observed differences in OTR density across cortical layers. Specifically, cortical layers 2/3 and 5 exhibited greater OTR density than layer 4. Our results point to a role for OT in integrating sensory and motor in the prairie vole brain, providing a complimentary mechanism for the modulation of social interactions. Given the ability of early social experience and developmental manipulations of OT to affect the brain and behavior, these results suggest a novel mechanism for how OT may influence cortical organization.
Keywords: oxytocin, prairie vole, neocortex, monogamy, social behavior
Introduction
Oxytocin (OT; see Table 1 for abbreviations) is a nine amino acid peptide that has long been known for its peripheral effects on female mammalian reproductive physiology, specifically uterine contractions during labor and milk ejection during nursing. But in the past few decades, it was discovered that OT and its analogs, including vasopressin and vasotocin, also have central effects and contribute to a wide variety of behaviors in many vertebrate species (Kelly and Goodson 2014), including food intake (Arletti, et al. 1990, Chaves, et al. 2013), stress response (Jezova, et al. 1995, Gibbs 1984, Williams, et al. 1985), temperature regulation (Chaves, et al. 2013, Young 2013, Kasahara, et al. 2015), grooming (Carter and Wilkinson 2015, Burkett, et al. 2016), and social and reproductive behaviors, including sexual behavior, parental behavior, infant attachment and social bonding (Insel and Shapiro 1992, Insel, et al. 1997, Bales, et al. 2013, Ross, et al. 2009, Bales, et al. 2007). Indeed, the OT system is an integral component to the development and expression of species-typical social behaviors.
Table 1.
Abbreviations
| AC | Auditory cortex |
| Amy | Amygdala |
| BNST | Bed Nucleus of the Stria Terminalis |
| CP | Caudate putamen |
| dpm | disintegrations per minute |
| HC | Hippocampus |
| Ins | Agranular insular area |
| LS | Lateral septum |
| M1 | Primary motor cortex |
| NAcc | Nucleus accumbens |
| OT | Oxytocin |
| OTR | Oxytocin receptor |
| Paa | Parietal association area |
| PLC | Prelimbic cortex |
| PVN | Paraventricular nucleus |
| S1 | Primary somatosensory cortex |
| S2 | Second somatosensory area |
| Taa | Temporal association area |
| V1 | Primary visual cortex |
Many areas containing oxytocin receptors (OTR) receive direct OT projections from the paraventricular nucleus (PVN) of the hypothalamus. But, there is also a considerable incongruency between the density of those projections and the density of OT binding sites, in that the distribution of cells that are immunoreactive for OT does not necessarily match the distribution of its receptors (Veening, et al. 2010). Furthermore, the expression and sensitivity of OTR are extensively controlled in a localized and specific way by many factors, including steroid hormones, thus resulting in a heterogeneous pattern of receptor binding in across the brain (Jimenez, et al. 2015, Mooney, et al. 2015, Ross, et al. 2009).
The neural distribution of OTR may underlie variations in social behavior. This can be seen by comparing OTR localization in the brain of two species of voles: the prairie vole (Microtus ochrogaster), which is monogamous and biparental, and the montane vole (Microtus montanus), which is polygamous (Insel and Shapiro 1992). The prairie vole has high OTR density in the prelimbic cortex (PLC), the bed nucleus of the stria terminalis (BNST), the nucleus accumbens (NAcc), the midline nuclei of the thalamus, and lateral aspects of the amygdala. In contrast, the montane vole has higher OTR density in the lateral septum, the ventromedial nucleus of the hypothalamus, and cortical nucleus of the amygdala. Moreover, in the female montane vole, the OTR distribution in the brain changes within 24h after parturition, more closely resembling the pattern observed in the prairie vole (Insel and Shapiro 1992). Thus, the distribution of the OTR in the brain of the voles can be directly associated with the social organization of the species (Insel and Shapiro 1992, Ross, et al. 2009).
In this study we used the prairie vole (Microtus ochrogaster) as the model species due to its very rare qualities: a rodent as easy to breed in the laboratory as the mouse, but monogamous and biparental (McGraw and Young 2010, Thomas and Birney 1979, Getz, et al. 1981). Moreover, these animals are only a few generations removed from the wild so there is still a great deal of intra-species diversity. As described before, the distribution of OTR in the subcortical regions of adult prairie voles has been well characterized (Insel and Shapiro 1992). But while a great deal is known about the subcortical distribution of OTR, no published studies have quantified its distribution across the prairie vole neocortex.
The mammalian neocortex is a six-layered structure which is involved in processing sensory inputs and generating motor output. Cortical areas are segregated by sensory modality (i.e., somatosensory, visual, and auditory areas), as well as a motor cortical area (Campi, et al. 2007). These functional representations correspond with distinct neuroanatomical characteristics, which can be identified using histological staining techniques. Interspersed between the primary sensory and motor areas are association regions. One common feature of association cortex is that these regions receive and process signals from one or more primary sensory and/or motor areas (Buckner and Krienen 2013). Different regions of association cortex are differentially recruited to integrate sensory and motor functions. The prefrontal cortex (also called the prelimbic cortex in rodents) is arguably the most studied and is involved in memory and decision making (Euston, et al. 2012, Miller and Cohen 2001). Other areas of association cortex are involved in the integration of sensory and motor information. The parietal association area (Paa), for example, integrates visual and sensorimotor processes (Wilber, et al. 2015, Whitlock, et al. 2012, Seelke, et al. 2011) while the temporal association area (Taa) integrates visual and auditory processes (Berger and Ehrsson 2014, Perrodin, et al. 2014).
This project focuses on the distribution of OTR in the cortex of adult prairie voles. In order to best appreciate the anatomical distribution of OTR we used a combination of histological techniques, including traditional coronal sectioning as well as a flattened cortex preparation. This novel approach allowed us to examine where OTR was found throughout cortical layers as well as across the entire cortical sheet. We found several regions of high OTR density within the prairie vole cortex, especially in association areas. Primary sensory and motor regions exhibited very low levels of OTR density. These results may provide insight into how the OT system influences cortical organization, particularly through the modification of association regions.
Experimental Procedures
Subjects
For this study, 12 prairie voles (Microtus ochrogaster) were subjects (6 males and 6 females). Animals were born and housed in the Psychology Department Vivarium at the University of California, Davis. These animals were descendants of a wild stock originally caught near Champaign, Illinois. The animals were weaned at 20 days of age and pair housed with an animal of the same sex (sibling if available, similarly aged non-sibling if not) in small laboratory cages (27 × 16 × 13 cm) in which food and water were available ad libitum. All animals were maintained at approximately 70°F (21°C) on a 14:10 light/dark cycle with the lights on at 6 a.m. All experiments were performed under National Institutes of Health guidelines for the care of animals in research and were approved by the Institutional Animal Care and Use Committee of the University of California, Davis.
Brain removal and slicing
Subjects were euthanized at day 60 via cervical dislocation under anesthesia, and their brains were removed, flash frozen on dry ice and stored at −80°C. Brains from 10 of the animals were sliced on a cryostat into 20μm coronal sections and mounted onto Super-frost slides. Brains from the remaining 2 animals had their cortical hemispheres dissected from subcortical regions, flattened, flash frozen on dry ice, sliced tangential to the pial surface into 20μm sections, and mounted onto Super-frost slides. All slides were stored at −80°C in sealed slide boxes with desiccants until the time of assay. Alternate adjacent sections were stained for Nissl substance (in the case of coronal sections) or myelin (in the case of flattened cortex sections).
Receptor Autoradiography
Receptor autoradiography for OTR was performed as described previously (Perkeybile, et al. 2015). Slides were removed from −80°C and allowed to thaw at room temperature. Next, they were lightly fixed in 0.1% paraformaldehyde (pH 7.4), washed twice in 50 mM Tris base (pH 7.4), and incubated in tracer buffer (pH 7.4) containing 0.05 nM 125I-d(CH2)5[Tyr(Me)2-Tyr-NH2]9-OVT (125I-OVTA; PerkinElmer) for 1 hr. Finally, slides were rinsed in 50 mM Tris base with 10 mM MgCl (pH 7.4) and dipped in deionized water before being allowed to air dry. The slides were exposed to BioMax MR film (Kodak, Rochester, NY) for 3 days and then developed and analyzed.
Nissl staining
The coronal sections that were stained for Nissl substance were left for 30 min in a chloroform-ethanol mix to defat the tissue. Then they underwent a series of alcohol rehydration steps (from 100% ethanol to 30% ethanol) before being immersed in a cresyl violet solution. Cresyl violet binds to DNA and RNA, staining them a blue/purple color. Thus the cell bodies, containing the DNA and the very large majority of the RNA, are apparent and the regions of the brain and layers of the cortex can be distinguished though their differing cellular densities. After the cresyl violet stain, the slides were dehydrated using a series of gradated alcohols (from 70% ethanol to 100% ethanol), cleared in histoclear and coverslipped with DPX mounting medium. Those sections were compared to the adjacent sections used for the autoradiography, so that we could precisely identify the anatomical regions of interest.
Quantification
The anatomical boundaries of regions of interest (ROI) were determined by comparing the sections that underwent autoradiography for OTR to adjacent sections that had been stained for Nissl substance and to a mouse brain atlas (Franklin and Paxinos 2013). Then the 125I-OVTA binding was quantified using ImageJ (NIH; Bethesda, MD, USA) to measure optical density. These techniques have been described previously (Freeman, et al. 2016). Films were scanned and digitized using an Epson Perfection V500 Photo scanner. The digitized autoradiogram images were imported into ImageJ 64, and optical binding density (OBD) values from a set of 125I autoradiography standards (American Radiolabeled Chemicals, Inc., St. Louis, MO) were loaded into the software and used to generate a standard curve, from which OBD values for each ROI could be extrapolated. In each brain, OBD values were calculated for each ROI as well as one background region where no binding was detected. Three measurements were taken for each ROI and background region. Background was quantified for each section of tissue from a subcortical area lacking OT receptors (corpus callosum or part of the thalamus, depending on the caudo-rostral position). The background OBD values were subtracted from each ROI value in order to account for individual differences in nonspecific binding and to yield normalize OBDs across brains.
We measured the optical binding density of the following brain regions: NAcc, PLC, insular cortex (Ins), Paa, Taa, primary motor areas (M1), primary auditory areas (AC) and primary somatosensory areas (S1). Additionally, a qualitative examination of the tissue revealed a heterogenous subcortical distribution of OTR, with the NAcc displaying some of the greatest density. Thus, we included the NAcc as a point of reference to compare cortical OTR density against an area in prairie voles that is characterized by high OTR density. For 4 animals (2 M and 2 F) the Paa and AC were not visible on the slices, so for these two areas, the values were obtained on only 6 animals instead of 10.
Additionally, an examination of several cortical regions, including M1, Taa, and Paa, revealed heterogenous OTR distribution across cortical layers. As before, ROIs were identified by comparing nissl-stained brain sections against adjacent sections that underwent OTR autoradiography. In each ROI, OBDs were determined for the area as a whole, as well as individual layers, including layers 2/3, 4, and 5.
Flattened cortex
While coronal sections allow us to clearly visualize cortical layers, it is difficult to appreciate the spatial relationship between cortical regions. Using a flattened cortical preparation allows for the simultaneous visualization of the entire cortical sheet and the arrangement of the primary sensory and motor areas. However, it is difficult to clearly identify layers using this preparation. Thus, the flattened cortex sections were used to assist in the visualization of the OTR distribution, but were not included in the quantification of data.
Alternating sections of the flattened cortical tissue were stained for myelin. Primary sensory regions within the neocortex contain a greater concentration of myelin than association areas. Thus, a myelin stain can be used to clearly identify architectonic boundaries of cortical regions. This method (Gallyas 1979) has been used previously by this laboratory (Campi, et al. 2007, Seelke, et al. 2012, Seelke, et al. in press). As with the Nissl stain, the myelin stain allowed us to clearly visualize and distinguish cortical areas.
Statistical Analyses
All statistical analyses were performed in R version 3.2.4 (The R Foundation; www.r-project.org) using the lmerTest and lsmeans packages. The lmerTest package utilizes the mixed modeling code found in lme4 but provides additional functionality, including the calculation of p-values using Satterwaite’s approximation for degrees of freedom. Since our experimental design only necessitated within-subject comparisons, we performed repeated measures analyses of variance (ANOVAs) with animal ID as a repeated effect. Post-hoc analyses were accomplished using the lsmeans package. Adjustments for multiple comparisons were computed using the Tukey method. For all analyses, α = 0.05.
Prior to analyzing differences in cortical regions, we first tested to see if OTR differed by hemisphere or sex using one-way ANOVAs for each factor. We found no significant difference between the left and right hemispheres for any ROI so we averaged the values together for further analyses. Similarly, having found no significant difference between the sexes in any of the tested regions we pooled male and female data. In summary, the density of OTR (in disintegrations per minute per milligram of tissue [dpm/mg]) for each ROI was determined by first averaging within subjects (left and right hemispheres) and then between subjects (male and female together).
Results
To quantify the localization and density of OTR in the cortex of adult prairie voles, we first examined coronal sections of vole brains. We compared images of OTR binding to adjacent sections stained with Nissl (Figure 1). Previous work has explored OTR distribution in subcortical regions, and in this study we saw dense OTR staining in areas that have already been described in the literature, including the NAcc, BNST, amygdala, and lateral septal nucleus (Bales, et al. 2007, Wang, et al. 1997, Witt and Insel 1991). Similarly, a close examination of the cortex revealed a non-homogenous distribution of OTR, with OTR density in some areas approaching that of the densest subcortical binding (e.g. NAcc). When we compared OTR binding across cortical regions we found a significant main effect of area (F7,62=98.379, p<2e-16; Figure 2), which confirms the heterogeneity of OTR distribution in the cortex. The amount of OTR did not significantly differ between Ins, Taa and PLC. Importantly, those areas also did not differ from NAcc, demonstrating that some cortical regions contain very high densities of OTR. We also found that there is no significant difference in OTR density between Paa and M1, although they both had significantly lower density than the NAcc, Ins, Taa, and PLC (t ≥ 3.287, p ≤ <0.0309). OTR density in AC was significantly lower than in Paa (p=0.0042), and S1 contained almost no OTR (difference between AC and S1 p<0.0001). Mean values in dpm/mg are shown in the Table 2. The p-value of the difference between every area is displayed in Table 3.
Figure 1.

Anatomical identification of cortical regions of interest. Adjacent sections were stained for Nissl substance (left) or processed for OTR autoradiography (right). In the right column, dark regions correspond to areas that contain high OTR density. The top row represents the most rostral regions of interest, while the bottom row represents the most caudal regions. Specific cortical regions and subcortical nuclei are labeled (see Table 1 for abbreviations). The Nissl stain allowed us to clearly identify specific regions of interest based on neuronal density, the presence of cortical layers, and the intensity of staining. All images were converted to grayscale, and brightness and contrast were adjusted equally across all images. In all images, dorsal is up and lateral is to the right.
Figure 2.
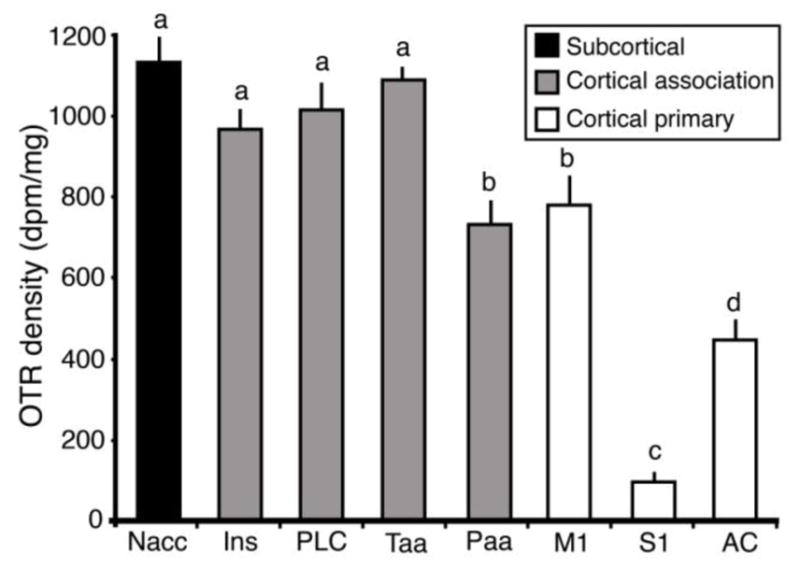
Oxytocin receptor density in different cortical regions. Groups that differ significantly from each other in the overall ANOVA are indicated by different letters. Subcortical regions (NAcc) are indicated in black, association cortical regions are indicated in gray, and primary sensory and motor regions are indicated in white. In the cortex, the association areas contain a higher density of OTR than the primary areas.
Table 2.
Average OTR density in different brain regions, in dpm. Mean ± standard error.
| Area | Mean ± s.e. |
|---|---|
| NAcc | 1137.9 ± 59.0 |
| INS | 975.6 ± 39.1 |
| PLC | 1022.2 ± 57.0 |
| Taa | 1094.5 ± 31.7 |
| Paa | 736.1 ± 52.4 |
| M1 | 789.3 ± 64.7 |
| S1 | 99.4 ± 23.4 |
| AC | 454.1 ± 45.8 |
Table 3.
Table shows p-values for comparisons between areas. Areas are listed on the X and Y axes. Significant differences (p < 0.05) are in bold.
| Area | NAcc | Ins | Lim | Taa | Paa | M1 | A1 | S1 |
|---|---|---|---|---|---|---|---|---|
| NAcc | 0.0816 | 0.435 | 0.9936 | <.0001 | <.0001 | <.0001 | <.0001 | |
| Ins | 0.9902 | 0.3988 | 0.0309 | 0.0256 | <.0001 | <.0001 | ||
| Lim | 0.896 | 0.0033 | 0.0017 | <.0001 | <.0001 | |||
| Taa | 0.0001 | <.0001 | <.0001 | <.0001 | ||||
| Paa | 0.9999 | 0.0042 | <.0001 | |||||
| M1 | 0.0002 | <.0001 | ||||||
| A1 | <.0001 | |||||||
| S1 |
Interestingly, when we partitioned the cortical regions of interest based on function, we found that the non-sensory (Ins, PLC) and association areas (Taa, Paa), had a much higher OTR density than the primary motor (M1; t ≥ 3.354, p ≤ 0.0256) and sensory areas (S1, AC; t ≥ 3.932, p ≤ 0.0042; Figure 2; Table 2). The one exception is the comparison between M1 and Paa, which did not significantly differ (t = 0.435, p = 0.999).
Heterogeneous OTR distribution in cortical layers
In addition to regional differences in OTR density, we also observed selected areas of heterogeneous laminar OTR distribution in coronal sections of vole cortex (Figures 3 and 4; Table 4). In the Paa for example, there was greater OTR density in layers 2/3 and layer 5 than in layer 4 (p<0.0001). It was the same with Paa, with greater OTR density in layers 2/3 and 5 than in layer 4 (p<0.0001). Within M1 we also found more OTR in layer 2/3 than in layer 4 (p<0.0001).
Figure 3.

Anatomical identification of cortical layers. Adjacent sections were stained for Nissl substance (left) or processed for OTR autoradiography (right). Arrows indicate the boundaries of the temporal association area, and cortical layers (1–6) and the amygdala (Amy) are labeled. All images were converted to grayscale and brightness and contrast were adjusted.
Figure 4.
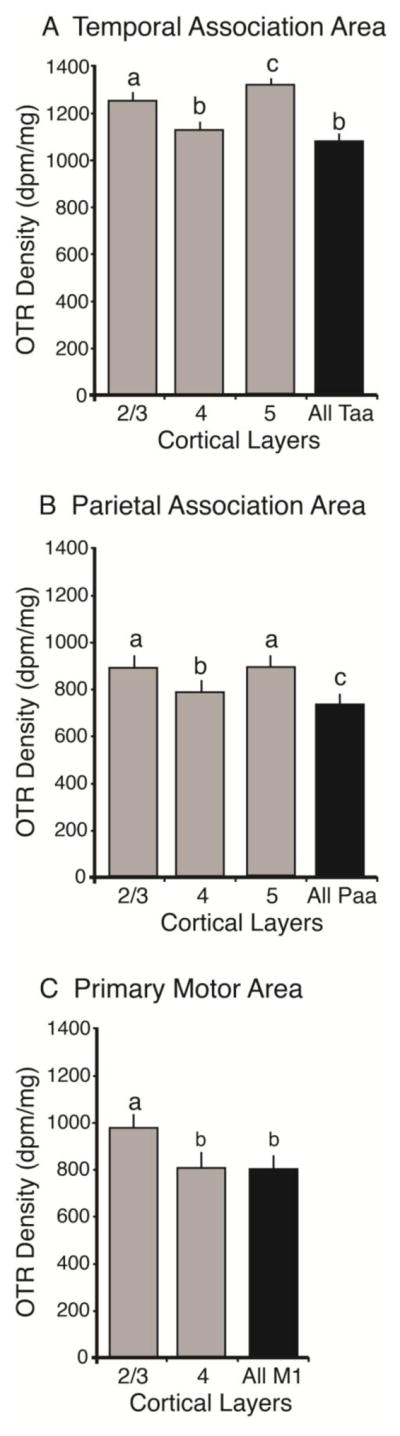
Quantification of oxytocin receptor density in different cortical layers. Groups that differ significantly from each other in the overall ANOVA are indicated by different letters. Individual layers (gray) were compared against each other and the cortical area as a whole (black). In the temporal association area (A) layers 2/3 and 5 had a higher OTR density than layer 4 as well as the area as a whole. In the parietal association area (B), layers 2/3 and 5 had a greater OTR density than layer 4, which in turn had a higher OTR density than the area as a whole. In the primary motor area (C), cortical layer 2/3 had a greater OTR density than layer 4 as well as the area as a whole.
Table 4.
Average OTR density throughout cortical layers, in dpm. Mean ± standard error.
| Taa | Paa | M1 | |
|---|---|---|---|
| Whole area | 1094.5 ± 31.7 | 736.1 ± 52.4 | 789.3 ± 64.7 |
| Layer 2/3 | 1254.1 ± 43.2 | 892.0 ± 56.9 | 691.7 ± 71.2 |
| Layer 4 | 1130.9 ± 35.2 | 787.5 ± 54.7 | 790.4 ± 72.6 |
| Layer 5 | 1327.3 ± 28.2 | 889.7 ± 57.2 |
Distribution of OTR in the flattened cortex
To investigate the distribution of OTR from another point of view, we used a flattened cortex preparation. This method allowed us to look at OTR distribution across the entire cortical sheet in sections parallel to the cortical layers. The flattened cortex preparation confirmed a distinct inverse relationship between OTR density and the location of the primary sensory regions (Figure 5). The architectonic organization of the prairie vole neocortex has been described previously (Campi, et al. 2007, Seelke, et al. 2016). Briefly, the primary visual cortex (V1) is located on the caudal pole of the neocortex and it stains darkly for myelin. The AC is a round structure that is found lateral to V1 and stains darkly for myelin. S1 is found rostral to V1 and AC, and stains darkly, but non-uniformly, for myelin. M1 is found immediately rostral to S1 and stains moderately for myelin. Indeed, one clear characteristic of cortical tissue is that the primary sensory and motor areas stain darkly for myelin. This pattern is in sharp contrast to the distribution of OTR density in flattened cortex, where the primary sensory areas have a low density of OTR. In contrast, the association areas, which stain very lightly for myelin, have a high density of OTR (Figure 5).
Figure 5.

Distribution of OTR across the cortical sheet. A) A flattened section of cortical tissue stained for myelin. The areas that stain darkly for myelin correspond with primary sensory and motor regions. B) The architectonic boundaries of cortical sensory and motor regions reconstructed from myelin-stained tissue. C) A flattened section of cortical tissue upon which OTR autoradiography was performed. Dark regions correspond to areas that contain high OTR density. In contrast to the myelin-stained section, here we see light staining in the sensory and motor areas and dark staining in the association areas, indicating low OTR density in sensory and motor regions and high OTR density in association regions. All images were converted to grayscale and brightness and contrast were adjusted. In all panels, rostral is to the right and medial is up.
Discussion
OT is involved in a wide variety of social behaviors, including pair bonding and parental care (Insel and Shapiro 1992, Bales, et al. 2013, Ross, et al. 2009, Bales, et al. 2007, Insel 1992, Carter, et al. 1995). The distribution of OTR is a critical aspect of the OT system, as the pattern of receptor expression indicates which neural substrates are sensitive to the local release of OT in the brain. Furthermore, differences in the neuroanatomical distribution of OTR have been able to account for some of the differences in social organization across vole species (Insel and Shapiro 1992, Lim, et al. 2004, Young and Wang 2004). The subcortical distribution of OTR has been examined in a wide variety of species, including but not limited to: rats, mice, several vole species, marmosets, coppery titi monkeys, rhesus macaques, and humans (Insel and Shapiro 1992, Freeman, et al. 2016, Vaccari, et al. 1998, Gould and Zingg 2003, Freeman, et al. 2014, Freeman, et al. 2014, Schorscher-Petcu, et al. 2009). However, neocortical OTR distribution has gone largely unquantified, even though in many of these papers there are clearly visible areas of high OTR density within the neocortex. To the best of our knowledge, this study is the first quantitative regional examination of OTR distribution in the neocortex of the prairie vole.
The neocortical distribution of OTR has been examined in other species, including the mouse (Hammock and Levitt 2013) and coppery titi monkey (Freeman, et al. 2014). In adult mice, little to no OTR were found in S1, which was the only region measured. However, developing mice showed significant levels of OTR in several layers within S1 (Hammock and Levitt 2013). The coppery titi monkey did not show appreciable levels of OTR anywhere in the neocortex (Freeman, et al. 2014). This result is in contrast to the data presented here, where some cortical regions, such as Ins, PLC, and Taa exhibit similar levels of OTR density as the Nacc, which contains some of the highest OTR density in the vole brain.
Numerous studies have shown behavioral differences between males and females when OT is involved, however in this study we found no differences in OTR density between male and female brains. Studies in many species have shown sex differences in brain OTR densities (reviewed in (Dumais and Veenema 2016)). Female rats show lower levels of OTR in most forebrain regions than males (Dumais, et al. 2013). Other regions in rats, like the central amygdala, display increased OTR for females relative to males. Conversely, several subcortical structures in the forebrain of prairie voles, including the BNST and amygdala, did not show differences between males and females (Bales, et al. 2007). Furthermore, while it was previously found that the OTR binding in the medial prefrontal cortex, (what we have labeled “prelimbic cortex”) was higher in female than in male prairie voles (Smeltzer, et al. 2006), we did not replicate this finding with our results. This could be explained by a number of statistical considerations including our small sample sizes and subsequent low power to detect sex differences. There is also the possibility that the disparity between the two studies could be explained by differences in methodology. While Smeltzer et al. (2006) simply measured grain density, our values were generated using a standard curve and we controlled for non-specific background binding.
Architectonic analysis of OTR distribution
The examination of cortical organization is complicated by the fact that the cortex is a very large 3-dimensional structure that shows significant variation across both its width and depth. Thus, in order to fully appreciate both its layered structure as well as the relative locations of primary sensory and motor regions, it was helpful to use multiple planes of section to obtain a more complete picture of the spatial distribution and density of OTR in the cortex. As in other mammalian species, the prairie vole neocortex is characterized by six layers that vary based on cell type and density. Sectioning tissue coronally provides a clear visualization of cortical layers and allows for the identification of boundaries between cortical regions (Figure 3). However, in order to observe the relative spatial organization of different cortical regions, it is helpful to view the entire cortical sheet in a single plane of section using a flattened cortical preparation (Figure 5). The flattened cortical preparation is most commonly used in neuroanatomical studies (e.g., (Campi, et al. 2007, Seelke, et al. 2011, Seelke, et al. 2012, Seelke, et al. in press, Seelke, et al. 2016), although is has been used in conjunction with autoradiography when documenting the development of ocular dominance columns in macaques (Horton and Hocking 1996). To our knowledge, this is the first time that the flattened cortex preparation has been used to examine the distribution of neuropeptide receptors.
Using both histological planes of section, we found that association regions, and not primary sensory and motor regions, contained the greatest OTR density. Both the sensory and association cortices likely play an important role in social behaviors: the sensory cortices directly receive information from the environment, and the association cortices integrate the different sensory inputs, which can influence behavioral responses. There have been recent studies that suggest OT plays a role in the processing of sensory information. A study in mice reported that sensory experience during early development regulates OT synthesis and secretion in the brain, which then mediates cross-modal plasticity (Zheng, et al. 2014). OT thus has direct effects on the sensory cortices, but the cross-modal plasticity might be happening via the association cortex, where cortical OTR density is greatest. In the bat, it was found that there are OT immunoreactive neurons in nuclei in the auditory brain stem (Kanwal and Rao 2002), as well as in the frontal and auditory cortices (Prasada Rao and Kanwal 2004). Mother-infant interactions in bats are highly dependent on vocal recognition and auditory feedback, and as adults bats rely on echolocation for spatial navigation as well as food acquisition (Schnitzler, et al. 2003). This result suggests that OT may influence sensory processing to help recognize relevant sensory inputs. A recent study in mice showed that OT disinhibits auditory cortex within seconds, and also leads to long-term modifications that are essential for balancing inhibition with excitation (Marlin, et al. 2015). Even in humans, it was found that variations in OTR gene expression was associated with auditory processing (Tops, et al. 2011). The presence of the presumably more efficient GG phenotype of an OTR gene polymorphism was positively correlated with decreased difficulty in hearing and understanding people within a lot of background noises.
The consequences of early experience
Cortical organization is very sensitive to environmental stimulation during critical periods, which renders it highly dynamic and adaptable to context. In prairie voles, it was shown that different parental rearing styles, measured by examining the amount of parental contact in the first few days after birth, are associated with differences in the organization of the cortical connections (Seelke, et al. 2016). The offspring of voles that were categorized as High Contact (HC) parents exhibited a different pattern of cortical connections than the offspring of voles categorized as Low Contact (LC) parents. The HC offspring had a greater density of intrinsic connections within the S1 compared to LC offspring, while the LC offspring had a wider distribution of labeled cells. Similarly, the size of cortical fields was altered both by differences in parental care as well as exposure to a single dose of OT on the day of birth (Seelke, et al. in press). Early experiences, likely mediated through the OT system, can also result in behavioral changes later in life. As we know, some changes in response to early experience are mediated by OT (see (Bales and Perkeybile 2012) for review), like the amount of alloparental behavior, which is higher in males who have been handled in the first few days of life (Bales, et al. 2011). These behavioral changes are correlated with OTR receptor density variation in some subcortical regions, with less OTR binding in the BNST and the NAcc. This result, taken in context with our findings, has lead us to hypothesize that the changes in cortical connections observed in HC vs LC offspring could be mediated by OTR in the cortex.
Other studies have shown that the OT might balance excitation and inhibition in the cortex. For example, in the rat, the excitatory-to-inhibitory switch of GABA actions in the fetal brain shortly before delivery is triggered by the maternal OT (Tyzio, et al. 2006). Moreover, in the mouse, OT has been shown to suppress excitatory glutamatergic neurotransmission in some part of the prefrontal cortex (Ninan 2011). Our data then suggest that the OT could play an important role at the association cortex level, maybe allowing balance in inhibition and excitation that could lead to cortical plasticity and successful social behaviors. This possibility provides an alternative mechanism by which OT may act to regulate the salience of social cues (Shamay-Tsoory and Abu-Akel 2016).
In this project we also found that the OTR are not homogeneously distributed between the cortical layers. In the motor areas, temporal association areas and posterior parietal association areas, layers 2/3 and 5 seem to express more OTR than the other layers. Interestingly, studies in other species also found more OTR in layer 2/3 of the cortex compared to other layers. In rats there were high levels of OTR mRNA in layers 2 and 3 of the neocortex (Yoshimura, et al. 1993). In mice, there is also a developmental peak of OTR density in the neocortex in pre-weaning mice (P14) (Hammock and Levitt 2013) and more precisely the OTR are prominent in layers 2 and 3 and in the upper layer 5. This heterogeneous OTR density is present in mice only during the early development, but it is very interesting to see that, much like we report here, OTR are always denser in layers 2 and 3, which send and receive projections from other cortical regions. The action of OT at OTR in layers 2/3 may be one mechanism by which OT can contribute to cortical plasticity.
Clinical relevance and future considerations
The research focused on the OT system has greatly increased for the past two decades. It is now seen as the “prosocial hormone” (Shamay-Tsoory and Abu-Akel 2016, Macdonald and Macdonald 2010, Kemp and Guastella 2010, Striepens, et al. 2011), warranting its scrutiny as a possible therapeutic for individuals with autism spectrum disorders (ASD; (Okamoto, et al. 2016, Yamasue 2016, Young and Barrett 2015). There are many clinical trials that are currently underway using intranasal administration of OT to treat patients with ASD. However, the literature surrounding OT as a therapeutic for ASD suffers from a variety of complications: (1) we do not know exactly how the OT system works in the brain (especially in humans), (2) the mechanism behind the effect of intranasal OT administration is still unknown (Quintana, et al. 2015), (3) clinical trials are experiencing a replication problem (Lane, et al. 2016), and (4) there may even be a publication bias for positive effects of OT administration (Lane, et al. 2016, Walum, et al. 2016) which confounds the potential null effect of intranasal OT administration.
But the question of whether OT can successfully be utilized as a treatment for ASD is fundamentally different than the question of whether or not the etiology of ASD involves the OT system. In fact, the idea that OT administration can alleviate the symptoms of ASD has been widely debated. Several studies have provided evidence for an effect while others have failed to do so (reviewed in Anagnostou, et al. 2014, Preti, et al. 2014), and a recent meta-analysis has concluded that OT administration has no effect on social cognition deficits or repetitive behaviors in children with ASD (Ooi, et al. 2016). But despite the lack of conclusive evidence for a therapeutic effect of OT on ASD, two related meta-analyses have confirmed associations between ASD and over a dozen variants of the OTR gene (Kranz, et al. 2016, LoParo and Waldman 2015). Other studies link alterations in OTR gene methylation status (e.g. gene expression activity) to children with ASD (Elagoz Yuksel, et al. 2016, Gregory, et al. 2009). These findings support the idea that the etiology of ASD may involve altered OTR distribution throughout the brain. Unfortunately, studies exploring the neuroanatomical distribution of OTR within the human brain have yet to characterize cortical regions (Freeman, et al. 2016). Given our finding of dense OTR binding with the prairie vole neocortex, further studies are warranted to determine the prevalence of OTR within human neocortex, particularly motor and sensory association regions. Localization of OTR in the human neocortex might help explain the fact that many individuals with ASD also experience comorbid alterations in responsivity to sensory input (Patten, et al. 2013, Ausderau, et al. 2014, Ausderau, et al. 2014, Ausderau, et al. 2016, American Psychiatric Association 2013), providing an alternative mechanism by which the OT system may modulate social experience.
Ultimately, there is a need to expand our understanding of the function of OT more broadly. While our project is an essential step to understanding the multi-faceted nature of the OT system, many more studies are still necessary. It would be quite informative to examine the development trajectory of the distribution of OTR in the neocortex of prairie voles, as OT seems to mediate many aspects of cortical development and plasticity, and the patterns of OTR binding in the brain have been found to change during development (Hammock 2013). According to our data, OT plays a more prominent role in the association cortex than in primary motor and sensory regions in the prairie vole. Having a greater understanding of the implications of these findings may inform how social and sensory experiences during infancy can influence behavioral and brain organization in adulthood.
Highlights.
Oxytocin receptors are heterogeneously distributed throughout the neocortex.
Primary sensory areas have low densities of oxytocin receptors.
Association regions have high densities of oxytocin receptors.
Cortical layers 2/3 and 5 exhibited greater oxytocin receptor density than layer 4.
Acknowledgments
This work was supported by grants to Karen L. Bales (NIH HD060117 and NIH HD071998). We thank Cindy Clayton and the animal care staff at the UC Davis Psychology vivarium for their help.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Kelly AM, Goodson JL. Social functions of individual vasopressin-oxytocin cell groups in vertebrates: What do we really know? Front Neuroendocrin. 2014;35:512–529. doi: 10.1016/j.yfrne.2014.04.005. [DOI] [PubMed] [Google Scholar]
- Arletti R, Benelli A, Bertolini A. Oxytocin inhibits food and fluid intake in rats. Physiol Behav. 1990;48:825–830. doi: 10.1016/0031-9384(90)90234-u. [DOI] [PubMed] [Google Scholar]
- Chaves VE, Tilelli CQ, Brito NA, Brito MN. Role of oxytocin in energy metabolism. Peptides. 2013;45:9–14. doi: 10.1016/j.peptides.2013.04.010. [DOI] [PubMed] [Google Scholar]
- Jezova D, Skultetyova I, Tokarev DI, Bakos P, Vigas M. Vasopressin and oxytocin in stress. Ann N Y Acad Sci. 1995;771:192–203. doi: 10.1111/j.1749-6632.1995.tb44681.x. [DOI] [PubMed] [Google Scholar]
- Gibbs DM. Dissociation of oxytocin, vasopressin and corticotropin secretion during different types of stress. Life Sci. 1984;35:487–491. doi: 10.1016/0024-3205(84)90241-8. [DOI] [PubMed] [Google Scholar]
- Williams TD, Carter DA, Lightman SL. Sexual dimorphism in the posterior pituitary response to stress in the rat. Endocrinology. 1985;116:738–740. doi: 10.1210/endo-116-2-738. [DOI] [PubMed] [Google Scholar]
- Young WS. Shedding heat on oxytocin. Endocrinology. 2013;154:3961–3962. doi: 10.1210/en.2013-1885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasahara Y, Tateishi Y, Hiraoka Y, Otsuka A, Mizukami H, Ozawa K, Sato K, Hidema S, Nishimori K. Role of the oxytocin receptor expressed in the rostral medullary raphe in thermoregulation during cold conditions. Front Endocrinol (Lausanne) 2015;6:180. doi: 10.3389/fendo.2015.00180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter GG, Wilkinson GS. Intranasal oxytocin increases social grooming and food sharing in the common vampire bat desmodus rotundus. Hormones and Behavior. 2015;75:150–153. doi: 10.1016/j.yhbeh.2015.10.006. [DOI] [PubMed] [Google Scholar]
- Burkett JP, Andari E, Johnson ZV, Curry DC, de Waal FB, Young LJ. Oxytocin-dependent consolation behavior in rodents. Science. 2016;351:375–378. doi: 10.1126/science.aac4785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Insel TR, Shapiro LE. Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles. Proc Natl Acad Sci U S A. 1992;89:5981–5985. doi: 10.1073/pnas.89.13.5981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Insel TR, Young L, Wang Z. Central oxytocin and reproductive behaviours. Rev Reprod. 1997;2:28–37. doi: 10.1530/ror.0.0020028. [DOI] [PubMed] [Google Scholar]
- Bales KL, Perkeybile AM, Conley OG, Lee MH, Guoynes CD, Downing GM, Yun CR, Solomon M, Jacob S, Mendoza SP. Chronic intranasal oxytocin causes long-term impairments in partner preference formation in male prairie voles. Biol Psychiatry. 2013;74:180–188. doi: 10.1016/j.biopsych.2012.08.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross HE, Freeman SM, Spiegel LL, Ren X, Terwilliger EF, Young LJ. Variation in oxytocin receptor density in the nucleus accumbens has differential effects on affiliative behaviors in monogamous and polygamous voles. J Neurosci. 2009;29:1312–1318. doi: 10.1523/JNEUROSCI.5039-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bales KL, van Westerhuyzen JA, Lewis-Reese AD, Grotte ND, Lanter JA, Carter CS. Oxytocin has dose-dependent developmental effects on pair-bonding and alloparental care in female prairie voles. Hormones and Behavior. 2007;52:274–279. doi: 10.1016/j.yhbeh.2007.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veening JG, de Jong T, Barendregt HP. Oxytocin-messages via the cerebrospinal fluid: Behavioral effects; a review. Physiol Behav. 2010;101:193–210. doi: 10.1016/j.physbeh.2010.05.004. [DOI] [PubMed] [Google Scholar]
- Jimenez A, Young LJ, Triana-Del Rio R, LaPrairie JL, Gonzalez-Mariscal G. Neuroanatomical distribution of oxytocin receptor binding in the female rabbit forebrain: Variations across the reproductive cycle. Brain Res. 2015;1629:329–339. doi: 10.1016/j.brainres.2015.10.043. [DOI] [PubMed] [Google Scholar]
- Mooney SJ, Coen CW, Holmes MM, Beery AK. Region-specific associations between sex, social status, and oxytocin receptor density in the brains of eusocial rodents. Neuroscience. 2015;303:261–269. doi: 10.1016/j.neuroscience.2015.06.043. [DOI] [PubMed] [Google Scholar]
- Ross HE, Cole CD, Smith Y, Neumann ID, Landgraf R, Murphy AZ, Young LJ. Characterization of the oxytocin system regulating affiliative behavior in female prairie voles. Neuroscience. 2009;162:892–903. doi: 10.1016/j.neuroscience.2009.05.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGraw LA, Young LJ. The prairie vole: An emerging model organism for understanding the social brain. Trends Neurosci. 2010;33:103–109. doi: 10.1016/j.tins.2009.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas JA, Birney EC. Parental care and mating system of the prairie vole, microtus-ochrogaster. Behavioral Ecology and Sociobiology. 1979;5:171–186. [Google Scholar]
- Getz LL, Carter CS, Gavish L. The mating system of the prairie vole, microtus-ochrogaster - field and laboratory evidence for pair-bonding. Behavioral Ecology and Sociobiology. 1981;8:189–194. [Google Scholar]
- Campi KL, Karlen SJ, Bales KL, Krubitzer L. Organization of sensory neocortex in prairie voles (microtus ochrogaster) J Comp Neurol. 2007;502:414–426. doi: 10.1002/cne.21314. [DOI] [PubMed] [Google Scholar]
- Buckner RL, Krienen FM. The evolution of distributed association networks in the human brain. Trends Cogn Sci. 2013;17:648–665. doi: 10.1016/j.tics.2013.09.017. [DOI] [PubMed] [Google Scholar]
- Euston DR, Gruber AJ, McNaughton BL. The role of medial prefrontal cortex in memory and decision making. Neuron. 2012;76:1057–1070. doi: 10.1016/j.neuron.2012.12.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miller EK, Cohen JD. An integrative theory of prefrontal cortex function. Annu Rev Neurosci. 2001;24:167–202. doi: 10.1146/annurev.neuro.24.1.167. [DOI] [PubMed] [Google Scholar]
- Wilber AA, Clark BJ, Demecha AJ, Mesina L, Vos JM, McNaughton BL. Cortical connectivity maps reveal anatomically distinct areas in the parietal cortex of the rat. Front Neural Circuits. 2015;8:146. doi: 10.3389/fncir.2014.00146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitlock JR, Pfuhl G, Dagslott N, Moser MB, Moser EI. Functional split between parietal and entorhinal cortices in the rat. Neuron. 2012;73:789–802. doi: 10.1016/j.neuron.2011.12.028. [DOI] [PubMed] [Google Scholar]
- Seelke AMH, Padberg J, Disbrow E, Purnell S, Recanzone G, Krubitzer L. Topographic maps within brodmann’s area 5 of macaque monkeys. Cereb Cortex. 2011 doi: 10.1093/cercor/bhr257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berger CC, Ehrsson HH. The fusion of mental imagery and sensation in the temporal association cortex. J Neurosci. 2014;34:13684–13692. doi: 10.1523/JNEUROSCI.0943-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perrodin C, Kayser C, Logothetis NK, Petkov CI. Auditory and visual modulation of temporal lobe neurons in voice-sensitive and association cortices. J Neurosci. 2014;34:2524–2537. doi: 10.1523/JNEUROSCI.2805-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perkeybile AM, Delaney-Busch N, Hartman S, Grimm KJ, Bales KL. Intergenerational transmission of alloparental behavior and oxytocin and vasopressin receptor distribution in the prairie vole. Front Behav Neurosci. 2015;9:191. doi: 10.3389/fnbeh.2015.00191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Franklin KBJ, Paxinos G. Paxinos and franklin’s the mouse brain in stereotaxic coordinates. 4. Academic Press, an imprint of Elsevier; Amsterdam: 2013. [Google Scholar]
- Freeman SM, Smith AL, Goodman MM, Bales KL. Selective localization of oxytocin receptors and vasopressin 1a receptors in the human brainstem. Soc Neurosci. 2016:1–11. doi: 10.1080/17470919.2016.1156570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallyas F. Silver staining of myelin by means of physical development. Neurol Res. 1979;1:203–209. doi: 10.1080/01616412.1979.11739553. [DOI] [PubMed] [Google Scholar]
- Seelke AMH, Dooley JC, Krubitzer LA. The emergence of somatotopic maps of the body in s1 in rats: The correspondence between functional and anatomical organization. PLoS One. 2012;7:e32322. doi: 10.1371/journal.pone.0032322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seelke AMH, Yuan S-M, Perkeybile AM, Krubitzer LA, Bales KL. Early experiences can alter the size of cortical fields in prairie voles (microtus ochrogaster) Environmental Epigenetics. doi: 10.1093/eep/dvw019. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bales KL, Plotsky PM, Young LJ, Lim MM, Grotte N, Ferrer E, Carter CS. Neonatal oxytocin manipulations have long-lasting, sexually dimorphic effects on vasopressin receptors. Neuroscience. 2007;144:38–45. doi: 10.1016/j.neuroscience.2006.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang Z, Hulihan TJ, Insel TR. Sexual and social experience is associated with different patterns of behavior and neural activation in male prairie voles. Brain Res. 1997;767:321–332. doi: 10.1016/s0006-8993(97)00617-3. [DOI] [PubMed] [Google Scholar]
- Witt DM, Insel TR. A selective oxytocin antagonist attenuates progesterone facilitation of female sexual behavior. Endocrinology. 1991;128:3269–3276. doi: 10.1210/endo-128-6-3269. [DOI] [PubMed] [Google Scholar]
- Seelke AMH, Perkeybile AM, Grunewald R, Bales KL, Krubitzer LA. Individual differences in cortical connections of somatosensory cortex are associate with parental rearing style in prairie voles (microtus ochrogaster) J Comp Neurol. 2016;524:564–577. doi: 10.1002/cne.23837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Insel TR. Oxytocin - a neuropeptide for affiliation: Evidence from behavioral, receptor autoradiographic, and comparative studies. Psychoneuroendocrino. 1992;17:3–35. doi: 10.1016/0306-4530(92)90073-g. [DOI] [PubMed] [Google Scholar]
- Carter CS, Devries AC, Getz LL. Physiological substrates of mammalian monogamy - the prairie vole model. Neurosci Biobehav R. 1995;19:303–314. doi: 10.1016/0149-7634(94)00070-h. [DOI] [PubMed] [Google Scholar]
- Lim MM, Murphy AZ, Young LJ. Ventral striatopallidal oxytocin and vasopressin v1a receptors in the monogamous prairie vole (microtus ochrogaster) J Comp Neurol. 2004;468:555–570. doi: 10.1002/cne.10973. [DOI] [PubMed] [Google Scholar]
- Young LJ, Wang Z. The neurobiology of pair bonding. Nat Neurosci. 2004;7:1048–1054. doi: 10.1038/nn1327. [DOI] [PubMed] [Google Scholar]
- Vaccari C, Lolait SJ, Ostrowski NL. Comparative distribution of vasopressin v1b and oxytocin receptor messenger ribonucleic acids in brain. Endocrinology. 1998;139:5015–5033. doi: 10.1210/endo.139.12.6382. [DOI] [PubMed] [Google Scholar]
- Gould BR, Zingg HH. Mapping oxytocin receptor gene expression in the mouse brain and mammary gland using an oxytocin receptor-lacz reporter mouse. Neuroscience. 2003;122:155–167. doi: 10.1016/s0306-4522(03)00283-5. [DOI] [PubMed] [Google Scholar]
- Freeman SM, Inoue K, Smith AL, Goodman MM, Young LJ. The neuroanatomical distribution of oxytocin receptor binding and mrna in the male rhesus macaque (macaca mulatta) Psychoneuroendocrino. 2014;45:128–141. doi: 10.1016/j.psyneuen.2014.03.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freeman SM, Walum H, Inoue K, Smith AL, Goodman MM, Bales KL, Young LJ. Neuroanatomical distribution of oxytocin and vasopressin 1a receptors in the socially monogamous coppery titi monkey (callicebus cupreus) Neuroscience. 2014;273:12–23. doi: 10.1016/j.neuroscience.2014.04.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schorscher-Petcu A, Dupre A, Tribollet E. Distribution of vasopressin and oxytocin binding sites in the brain and upper spinal cord of the common marmoset. Neurosci Lett. 2009;461:217–222. doi: 10.1016/j.neulet.2009.06.016. [DOI] [PubMed] [Google Scholar]
- Hammock EA, Levitt P. Oxytocin receptor ligand binding in embryonic tissue and postnatal brain development of the c57bl/6j mouse. Front Behav Neurosci. 2013;7:195. doi: 10.3389/fnbeh.2013.00195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumais KM, Veenema AH. Vasopressin and oxytocin receptor systems in the brain: Sex differences and sex-specific regulation of social behavior. Front Neuroendocrin. 2016;40:1–23. doi: 10.1016/j.yfrne.2015.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dumais KM, Bredewold R, Mayer TE, Veenema AH. Sex differences in oxytocin receptor binding in forebrain regions: Correlations with social interest in brain region- and sex- specific ways. Hormones and Behavior. 2013;64:693–701. doi: 10.1016/j.yhbeh.2013.08.012. [DOI] [PubMed] [Google Scholar]
- Smeltzer MD, Curtis JT, Aragona BJ, Wang Z. Dopamine, oxytocin, and vasopressin receptor binding in the medial prefrontal cortex of monogamous and promiscuous voles. Neurosci Lett. 2006;394:146–151. doi: 10.1016/j.neulet.2005.10.019. [DOI] [PubMed] [Google Scholar]
- Horton JC, Hocking DR. An adult-like pattern of ocular dominance columns in striate cortex of newborn monkeys prior to visual experience. J Neurosci. 1996;16:1791–1807. doi: 10.1523/JNEUROSCI.16-05-01791.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng JJ, Li SJ, Zhang XD, Miao WY, Zhang D, Yao H, Yu X. Oxytocin mediates early experience-dependent cross-modal plasticity in the sensory cortices. Nat Neurosci. 2014;17:391–399. doi: 10.1038/nn.3634. [DOI] [PubMed] [Google Scholar]
- Kanwal JS, Rao PD. Oxytocin within auditory nuclei: A neuromodulatory function in sensory processing? Neuroreport. 2002;13:2193–2197. doi: 10.1097/00001756-200212030-00006. [DOI] [PubMed] [Google Scholar]
- Prasada Rao PD, Kanwal JS. Oxytocin and vasopressin immunoreactivity within the forebrain and limbic-related areas in the mustached bat, pteronotus parnellii. Brain Behav Evol. 2004;63:151–168. doi: 10.1159/000076241. [DOI] [PubMed] [Google Scholar]
- Schnitzler H-U, Moss CF, Denzinger A. From spatial orientation to food acquisition in echolocating bats. Trends Ecol Evol. 2003;18:386–394. [Google Scholar]
- Marlin BJ, Mitre M, D’Amour AJ, Chao MV, Froemke RC. Oxytocin enables maternal behaviour by balancing cortical inhibition. Nature. 2015;520:499–504. doi: 10.1038/nature14402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tops M, van Ijzendoorn MH, Riem MM, Boksem MA, Bakermans-Kranenburg MJ. Oxytocin receptor gene associated with the efficiency of social auditory processing. Front Psychiatry. 2011;2:60. doi: 10.3389/fpsyt.2011.00060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bales KL, Perkeybile AM. Developmental experiences and the oxytocin receptor system. Hormones and Behavior. 2012;61:313–319. doi: 10.1016/j.yhbeh.2011.12.013. [DOI] [PubMed] [Google Scholar]
- Bales KL, Boone E, Epperson P, Hoffman G, Carter CS. Are behavioral effects of early experience mediated by oxytocin? Front Psychiatry. 2011;2:24. doi: 10.3389/fpsyt.2011.00024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tyzio R, Cossart R, Khalilov I, Minlebaev M, Hubner CA, Represa A, Ben-Ari Y, Khazipov R. Maternal oxytocin triggers a transient inhibitory switch in gaba signaling in the fetal brain during delivery. Science. 2006;314:1788–1792. doi: 10.1126/science.1133212. [DOI] [PubMed] [Google Scholar]
- Ninan I. Oxytocin suppresses basal glutamatergic transmission but facilitates activity-dependent synaptic potentiation in the medial prefrontal cortex. J Neurochem. 2011;119:324–331. doi: 10.1111/j.1471-4159.2011.07430.x. [DOI] [PubMed] [Google Scholar]
- Shamay-Tsoory SG, Abu-Akel A. The social salience hypothesis of oxytocin. Biol Psychiatry. 2016;79:194–202. doi: 10.1016/j.biopsych.2015.07.020. [DOI] [PubMed] [Google Scholar]
- Yoshimura R, Kiyama H, Kimura T, Araki T, Maeno H, Tanizawa O, Tohyama M. Localization of oxytocin receptor messenger ribonucleic acid in the rat brain. Endocrinology. 1993;133:1239–1246. doi: 10.1210/endo.133.3.8396014. [DOI] [PubMed] [Google Scholar]
- Macdonald K, Macdonald TM. The peptide that binds: A systematic review of oxytocin and its prosocial effects in humans. Harv Rev Psychiatry. 2010;18:1–21. doi: 10.3109/10673220903523615. [DOI] [PubMed] [Google Scholar]
- Kemp AH, Guastella AJ. Oxytocin: Prosocial behavior, social salience, or approach-related behavior? Biol Psychiatry. 2010;67:e33–34. doi: 10.1016/j.biopsych.2009.11.019. author reply e35. [DOI] [PubMed] [Google Scholar]
- Striepens N, Kendrick KM, Maier W, Hurlemann R. Prosocial effects of oxytocin and clinical evidence for its therapeutic potential. Front Neuroendocrinol. 2011;32:426–450. doi: 10.1016/j.yfrne.2011.07.001. [DOI] [PubMed] [Google Scholar]
- Okamoto Y, Ishitobi M, Wada Y, Kosaka H. The potential of nasal oxytocin administration for remediation of autism spectrum disorders. CNS Neurol Disord Drug Targets. 2016;15:564–577. doi: 10.2174/1871527315666160413120845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamasue H. Promising evidence and remaining issues regarding the clinical application of oxytocin in autism spectrum disorders. Psychiatry Clin Neurosci. 2016;70:89–99. doi: 10.1111/pcn.12364. [DOI] [PubMed] [Google Scholar]
- Young LJ, Barrett CE. Can oxytocin treat autism? Science. 2015;347:825–826. doi: 10.1126/science.aaa8120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quintana DS, Alvares GA, Hickie IB, Guastella AJ. Do delivery routes of intranasally administered oxytocin account for observed effects on social cognition and behavior? A two-level model. Neurosci Biobehav Rev. 2015;49:182–192. doi: 10.1016/j.neubiorev.2014.12.011. [DOI] [PubMed] [Google Scholar]
- Lane A, Luminet O, Nave G, Mikolajczak M. Is there a publication bias in behavioural intranasal oxytocin research on humans? Opening the file drawer of one laboratory. J Neuroendocrinol. 2016;28 doi: 10.1111/jne.12384. [DOI] [PubMed] [Google Scholar]
- Walum H, Waldman ID, Young LJ. Statistical and methodological considerations for the interpretation of intranasal oxytocin studies. Biol Psychiatry. 2016;79:251–257. doi: 10.1016/j.biopsych.2015.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anagnostou E, Soorya L, Brian J, Dupuis A, Mankad D, Smile S, Jacob S. Intranasal oxytocin in the treatment of autism spectrum disorders: A review of literature and early safety and efficacy data in youth. Brain Res. 2014;1580:188–198. doi: 10.1016/j.brainres.2014.01.049. [DOI] [PubMed] [Google Scholar]
- Preti A, Melis M, Siddi S, Vellante M, Doneddu G, Fadda R. Oxytocin and autism: A systematic review of randomized controlled trials. J Child Adolesc Psychopharmacol. 2014;24:54–68. doi: 10.1089/cap.2013.0040. [DOI] [PubMed] [Google Scholar]
- Ooi YP, Weng SJ, Kossowsky J, Gerger H, Sung M. Oxytocin and autism spectrum disorders: A systematic review and meta-analysis of randomized controlled trials. Pharmacopsychiatry. 2016 doi: 10.1055/s-0042-109400. [DOI] [PubMed] [Google Scholar]
- Kranz TM, Kopp M, Waltes R, Sachse M, Duketis E, Jarczok TA, Degenhardt F, Gorgen K, Meyer J, Freitag CM, Chiocchetti AG. Meta-analysis and association of two common polymorphisms of the human oxytocin receptor gene in autism spectrum disorder. Autism Res. 2016;9:1036–1045. doi: 10.1002/aur.1597. [DOI] [PubMed] [Google Scholar]
- LoParo D, Waldman ID. The oxytocin receptor gene (oxtr) is associated with autism spectrum disorder: A meta-analysis. Mol Psychiatry. 2015;20:640–646. doi: 10.1038/mp.2014.77. [DOI] [PubMed] [Google Scholar]
- Elagoz Yuksel M, Yuceturk B, Faruk Karatas O, Ozen M, Dogangun B. The altered promoter methylation of oxytocin receptor gene in autism. J Neurogenet. 2016:1–5. doi: 10.1080/01677063.2016.1202951. [DOI] [PubMed] [Google Scholar]
- Gregory SG, Connelly JJ, Towers AJ, Johnson J, Biscocho D, Markunas CA, Lintas C, Abramson RK, Wright HH, Ellis P, Langford CF, Worley G, Delong GR, Murphy SK, Cuccaro ML, Persico A, Pericak-Vance MA. Genomic and epigenetic evidence for oxytocin receptor deficiency in autism. BMC Med. 2009;7:62. doi: 10.1186/1741-7015-7-62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Patten E, Ausderau KK, Watson LR, Baranek GT. Sensory response patterns in nonverbal children with asd. Autism Research and Treatment. 2013;2013:9. doi: 10.1155/2013/436286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ausderau K, Sideris J, Furlong M, Little LM, Bulluck J, Baranek GT. National survey of sensory features in children with asd: Factor structure of the sensory experience questionnaire (3.0) Journal of Autism and Developmental Disorders. 2014;44:915–925. doi: 10.1007/s10803-013-1945-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ausderau KK, Furlong M, Sideris J, Bulluck J, Little LM, Watson LR, Boyd BA, Belger A, Dickie VA, Baranek GT. Sensory subtypes in children with autism spectrum disorder: Latent profile transition analysis using a national survey of sensory features. Journal of Child Psychology and Psychiatry and Allied Disciplines. 2014;55:935–944. doi: 10.1111/jcpp.12219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ausderau KK, Sideris J, Little LM, Furlong M, Bulluck JC, Baranek GT. Sensory subtypes and associated outcomes in children with autism spectrum disorders. Autism Research. 2016 doi: 10.1002/aur.1626. n/a-n/a. [DOI] [PubMed] [Google Scholar]
- American Psychiatric Association. Dsm 5. American Psychiatric Association; 2013. [Google Scholar]