

Abstract
Purpose
Dickkopf 1 (DKK1) has been extensively investigated in mouse models of multiple myeloma, which results in osteolytic bone lesions. Elevated DKK1 levels in bone marrow plasma and serum inhibit the differentiation of osteoblast precursors. Present pharmaceutical approaches to target bone lesions are limited to antiresorptive agents. In this study, we developed a cyclized oligopeptide against DKK1-low density lipoprotein receptor-related protein (LRP) 5/6 interaction and tested the effects of the oligopeptide on tumor burden.
Materials and Methods
A cyclized oligopeptide based on DKK1-LRP5/6 interactions was synthesized chemically, and its nuclear magnetic resonance structure was assessed. Luciferase reporter assay and mRNA expressions of osteoblast markers were evaluated after oligopeptide treatment. MOPC315.BM.Luc cells were injected into the tail vein of mice, after which cyclized oligopeptide was delivered subcutaneously 6 days a week for 4 weeks.
Results
The cyclized oligopeptide containing NXI motif bound to the E1 domain of LRP5/6 effectively on surface plasmon resonance analysis. It abrogated the Wnt-β-catenin signaling inhibited by DKK1, but not by sclerostin, dose dependently. RT-PCR and alkaline phosphatase staining showed increased expressions of osteoblast markers according to the treatment concentrations. Bioluminescence images showed that the treatment of cyclized oligopeptide reduced tumor burden more in oligopeptide treated group than in the vehicle group.
Conclusion
The cyclized oligopeptide reported here may be another option for the treatment of tumor burden in multiple myeloma.
Keywords: Multiple myeloma, DKK1, oligopeptide, tumor, burden, Wnt signaling
INTRODUCTION
Multiple myeloma (MM) is a disease of cancerous plasma cells that expand in bone marrow, causing symptoms of anemia, hypercalcemia, and osteolytic bone lesions.1 The disease is characterized by uncontrolled proliferation of a plasma cell clone with an accumulation of monoclonal immunoglobulins. 2 Myeloma and stromal cells produce osteoclast activating factors, such as IL-6, IL-3, receptor activator nuclear factor kappa B ligand (RANKL), TNF-α, and MIP1α, which are responsible for bone resorption.3,4,5,6,7 Furthermore, myeloma cells induce changes in the bone marrow microenvironment, thereby the production of osteoblastic inhibitor factors, such as TGF-β, TNF-α, and IL-3, is increased.8,9 Accordingly, an imbalance in osteoclast and osteoblast activity with the prevalence of bone resorption leads to marrow destruction.
The Wnt signaling pathway plays important role in the process of osteogenic differentiation and in the maintenance of mesenchymal stem cells, as well as in embryonic development. 10 Several attempts have been made to target the Wnt signaling pathway by inhibiting endogenous antagonists and by regulating intracellular mediators for the treatment of osteogenic disorders.11 Antibodies against endogenous Wnt antagonists, such as sclerostin, have shown bone-forming and fracture healing effects.12 Dickkopf 1 (DKK1) is another important endogenous inhibitor of the canonical Wnt signaling pathway, which binds to low density lipoprotein receptor-related protein (LRP) 5/6, together with Kremen receptor, to inhibit the canonical Wnt signaling pathway.13
DKK1 has been extensively investigated in murine models of MM where osteolytic lesions occur. Many studies have confirmed that the interaction of MM cells with the bone marrow microenvironment causes the bone lesions of myeloma.14 MM cancer cells secrete DKK1, which disrupts the balance of osteoblastogenesis and osteoclastogenesis.15 DKK1 disrupts the differentiation of mesenchymal stem cells to osteoblast lineage cells, and this results in a shift of the RANKL/osteoprotegerin (OPG) ratio, leading to excessive bone resorption and marrow destruction.15 Enhanced bone resorption is not followed by neo-matrix deposition due to the inhibition of further differentiation of osteoblast precursors by DKK1.16 Therefore, DKK1 has been a potential therapeutic target for the treatment of MM.
Presently approved medications that target bone diseases are limited to anti-resorptive agents and bone-anabolic agents,17 and bone-protecting anti-resorptive agents have failed to confer significant antitumor activities in clinical studies. 18 A small number of papers have reported that the treatment of antibodies against DKK1 not only reduces bone lesions but also reduces the tumor burden of the disease.19,20 However, exact mechanisms remain under investigation. We hypothesized that DKK1-inhibiting oligopeptide would bind in the place of DKK1 of the Wnt signaling pathway, reducing the binding of the overexpressed DKK1 protein by MM cells. The present study aimed to develop a cyclized oligopeptide containing the NXI motif already known in DKK1 and to test the effects of the oligopeptide on tumor burden.
MATERIALS AND METHODS
Synthesis and purification of peptides
The peptide consisted of 10 amino acids with one disulfide bond; it contains NXI motif, which is found in DKK1. Peptides were synthesized via Fmoc solid phase peptide synthesis using ASP48S (Peptron Inc., Daejeon, Korea) and purified via the reverse phase high-performance liquid chromatography using a Vydac Everest C18 column (250×22 mm, 10 µm). Elution was carried out with a water-acetonitrile linear gradient [3–40% (v/v) of acetonitrile] containing 0.1% (v/v) trifluoroacetic acid. Molecular weights of the purified peptide were confirmed using Liquid Chromatography/Mass Spectrometry (Agilent HP1100 series, Agilent Technologies, Waldbronn, Germany).
Cell culture
MOPC315.BM.Luc cells21 were obtained from Bogen Lab. Briefly, cells were developed from a MOPC315 cell line through s.c. injection twice to obtain MOPC315.4 and i.v. injection to obtain final cell line MOPC315.BM. Then, cells were tagged with luciferase gene. Cells were cultured in RPMI 1640 medium supplemented with fetal bovine serum (FBS) and penicillin-streptomycin (Welgene, Gyeongsan, Korea). ST2 cells were cultured in dulbecco's modified eagle's medicum medium, and luciferase transfected MC3T3-E1 cells were cultured in alpha-minimum essential medium with FBS and penicillin-streptomycin.
Nuclear magnetic resonance (NMR) spectroscopy
Nuclear magnetic resonance (NMR) spectra were acquired at 283 K on a Bruker DRX-500 spectrometer equipped with a triple resonance probe with an x, y, and z-shielded pulsed-field gradient coil. Two-dimensional (2D) NMR spectra were recorded in a phase-sensitive mode using time proportional phase increment for quadrature detection in the t1 domain. Total correlation spectroscopy (TOCSY)22 using a dipsi-2 spinlock pulse sequence with a mixing time of 70 ms and nuclear overhauser enhancement spectroscopy (NOESY)23 with mixing times of 250–600 ms were performed. All NMR spectra were acquired with 2048 complex data points in t2 and 256 increments in the t1 dimension, with 64 scans per each increment. All NMR data were processed using nmrPipe/nmrDraw or XWIN-NMR software (Bruker Instruments, Karlsruhe, Germany) and analyzed using the Sparky 3.95 program.
Structure calculations
Distance restraints were derived from the NOESY spectra in 100% deuterated DMSO solution. The solution structures were calculated using the hybrid distance geometry and dynamical simulated annealing (SA) protocol using the CNS 1.1 program on a Linux workstation. Final structures were analyzed using the PROCHECK20 and MOLMOL programs.24
Surface plasmon resonance (SPR) assay
An surface plasmon resonance (SPR) 36 assay was performed by the Gyeonggi Bio Center (Suwon, Korea), using a ProteOn XPR36 instrument (Bio-Rad Laboratories, Hercules, CA, USA). LRP5 E1 domains were tethered (immobilized) onto a biosensor chip surface. Then, five different concentrations (1, 3, 10, 30, and 100 µM) of the oligopeptide were passed over the LRP5 E1 domain in order for binding interactions to occur. The Langmuir model of the ProteOn Manager™ program (Bio-Rad Laboratory) was used for SPR data analysis.
Transfected MC3T3-E1 cell luciferase reporter assay
In the Wnt signaling study, MC3T3-E1 cells were transfected as previously reported.25 Seeded cells were treated in different concentrations (50 nM, 500 nM, 5 µM) of oligopeptide along with Wnt7a and DKK1 for 24 hours. Cells were collected after lysis in 1X passive lysis buffer (Promega, Madison, WI, USA) and measured for luciferase reporter activity following the Promega protocol.
ST2 cell alkaline phosphatase staining assay (ALP staining)
ST2 cells were seeded and treated with Wnt7a, DKK1, and different concentrations of oligopeptide: 50 nM, 500 nM, 5 µM, and 50 µM. After 48 hours, alkaline phosphatase (ALP) staining assay was performed using an ALP kit (Sigma-Aldrich, St. Louis, MO, USA) according to the manufacturer's protocol. Images were taken using an Olympus IX73 inverted microscope at magnification ×40.
Mice and injections
Five-week-old BALB/c female mice were purchased (Orientbio, Seongnam, Korea) and housed with five mice per cage at the Yonsei Biomedical Research Institute. Mice were maintained for 1 week to allow them to adapt to the new environment. A total of 30 mice were divided into three groups (ten mice each): a negative control, a positive control, and an experimental group. The negative control group received 150 µL of PBS via tail vein injection, while the other 20 mice received 2×105 MOPC315.BM.Luc cells in 150 µL of PBS via tail vein injection. Five days after the cell injection, oligopeptide (75 µg/150 µL/mouse) and vehicle (150 µL of phosphate-buffered saline/mouse) treatments were administered via subcutaneous injection 6 days per week for 4 weeks.
Bioluminescence imaging (BLI)
All mice underwent bioluminescence imaging (BLI) using an IVIS Spectrum (Caliper-Xenogen, Hopkinton, MA, USA) 5 days after the tail vein cell injection and 4 weeks after oligopeptide treatment. D-luciferin (VivoGlo Luciferin, Promega, Madison, WI, USA) was injected (150 mg/kg) with an anesthesia mixture of tiletamine+zolazepam (30 mg/kg) and xylazine (10 mg/kg) 10 minutes prior to taking the image via intraperitoneal injection. Bioluminescence was measured from the dorsal and ventral sides of mice. Images were evaluated with Living Image® 4.4 (Caliper-Xenogen, Hopkinton, MA, USA).
Ethics statement
All experiments were conducted according to the guidelines for the care and use of laboratory animals (National Research Council, Washington, DC, USA), the IACUC, and the 3R principles.
Statistics
Values are expressed as mean±standard deviation and median, and statistical analysis was performed using and Graph-Pad Prism 5 software. Mann-Whitney U test was used to analyze statistical significance, and differences were considered to be statistically significant when p<0.05.
RESULTS
Solution structures of the cyclized oligopeptide
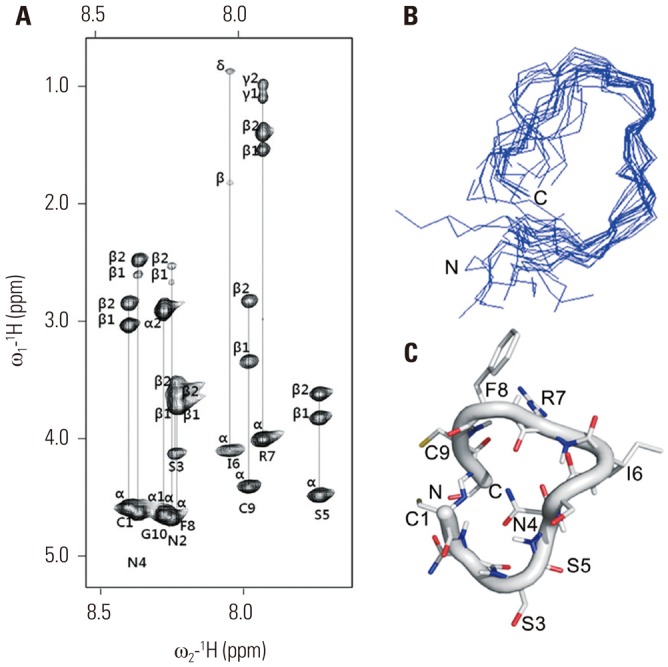
Once the individual spin systems had been classified, backbone sequential resonance assignment was easily completed via dαN (i, i+1) NOE connectivities in the 2D-NOESY spectra. The side chain assignment was performed via TOCSY connectivities (Fig. 1A). All NOEs were observed at mixing times of 600 ms. A total of 100 starting structures were calculated in the initial SA stage. The 20 lowest-energy structures (〈SA〉k) were selected out of the 100 final simulated-annealing structures for structural analysis. The average structure was generated from the geometrical average of 20 structure coordinates and was subjected to restrained energy minimization to correct bond length and angle distortions. The average NMR structure exhibited 0.68-Å root-mean-square deviation for backbone atoms with respect to 20 (〈SA〉k) structures. A best-fit superposition of all final structures and the backbone conformation for the average restrained-energy minimized structure (〈SA〉kr) are displayed in Fig. 1B. The overall fold of the cyclized oligopeptide is presented as a ribbon diagram in Fig. 1C. The cyclized oligopeptide shows a loop conformation with no regular secondary structure.
Fig. 1. NMR structure of cyclized oligopeptide. (A) 2D TOCSY spectrum of DKK1 (1 mM) in 100% DMSO at pH 7.0, 283 K, recorded with a spin-lock mixing time of 70 ms. The spectrum shows the identification of amino acid spin systems based on scalar correlation of the backbone amide protons of (ω2), with the respective side-chain protons (ω1) of each spin system. (B) A backbone superposition of the energy-minimized average structure (〈SA〉kr) over the family of 20 final SA structures (〈SA〉kr). (C) A ribbon diagram of DKK1 with side-chain orientations is displayed as a stick model. NMR, nuclear magnetic resonance; 2D, two-dimensional; TOCSY, total correlation spectroscopy; DKK1, dickkopf 1; SA, simulated annealing.
Cyclized oligopeptide binds to the LRP5 E1 site and targets Wnt signaling
The binding activity increased as the flowing oligopeptide concentration increased from 1 µM to 100 µM (Fig. 2A). MC3T3-E1-S-Top cells were treated with Wnts, Wnt inhibitors, and oligopeptide. When the cells were treated with Wnt3a and DKK1, the activity increased as the concentration of oligopeptide increased; however, when treated with sclerostin, the activity could not overcome the inhibition effect (Fig. 2B). Similar results were observed with Wnt7a treatment (Fig. 2C). These results suggest that the oligopeptide is relatively DKK1 specific and that the oligopeptide targets the Wnt signaling pathway, abrogating the inhibition effects of DKK1.
Fig. 2. Luciferase reporter activity of cells treated with cyclized oligopeptide and ProteOn XPR36 analysis. MC3T3-E1-S-Top cells were treated with Wnt3a, DKK1, sclerostin, and oligopeptide. (A) This graph shows the oligopeptide bound directly to the LRP5 E1 domain and the activity response increased dose dependently. (B) Graph on the left shows the dose increasing with luciferase reporter activity as the concentration of the oligopeptide increased. However, in the graph on the right, when treated with sclerostin, the same trend is not seen. (C) The same cells were treated with Wnt7a, DKK1, sclerostin and oligopeptide. DKK1, dickkopf 1; LRP, lipoprotein receptor-related protein; SOST, sclerositn.

Cyclized oligopeptide reverses the inhibitory effect of DKK1 on osteoblasts in vitro
ST2 cells were treated with Wnt7a, DKK1, and oligopeptide. Wnt7a treatment increased ALP and OPG mRNA expression levels, and the addition of DKK1 decreased mRNA expression levels. However, when cells were treated with oligopeptide, the mRNA expressions of ALP and OPG increased again (Fig. 3A). This result indicated that the oligopeptide activates the canonical Wnt signaling that is inhibited by DKK1. To further confirm this result, ST2 cells were treated with Wnt7a, DKK1, and oligopeptide for ALP staining assay. The results of the ALP staining assay were similar to those seen in the RT-PCR. Cells treated with only Wnt7a expressed a stronger color, and the treatment of DKK1 decreased the staining. Cells treated with Wnt7a, DKK1, and oligopeptide could overcome the inhibition effect of DKK1 dose dependently, shown by the increase in staining intensity (Fig. 3B). These results together suggested that this oligopeptide has bone-anabolic effects in vitro.
Fig. 3. Reverse transcription-PCR and Alkaline phosphatase staining assay. (A) ST2 cells were treated with Wnt7a, DKK1, and oligopeptide then the mRNA expression of both ALP and OPG assessed. (B) When Wnt7a alone was used, the staining was stronger than when Wnt7a and DKK1 were used together. The staining intensity also increased dose dependently with increasing concentration of oligopeptide treatment. Original magnification ×40. (C) The expressions of DKK1 and β-catenin were checked in the MOPC.315.BM.Luc cells prior to the experiment. DKK1, dickkopf 1; ALP, alkaline phosphatase; OPG, osteoprotegerin.

Cyclized oligopeptide inhibits tumor growth in a multiple myeloma mouse model
To determine the effect of oligopeptide on MM cell growth in vivo, DKK1 expression was confirmed. The mRNA expression of both DKK1 and β-catenin were detected (Fig. 3C), indicating that the canonical Wnt signaling pathway may have certain effects on MM progression and bone lesions. After the expression of DKK1 was proven, we injected MOPC315.BM.Luc cells into the tail vein of 20 BALB/c mice (day -5), and five days later (day 0) mice were injected subcutaneously with either the oligopeptide or vehicle alone (Fig. 4A). Tumor burden was measured via BLI (Fig. 4B). On day 0, no significant radiance (p/sec/cm2/sr) was detected in any of the representative mice of three groups. In contrast, on day 28, all mice injected with MM cells invoked a BLI signal. Importantly, larger radiances were detected in the vehicle group than in the oligopeptide group. No significant radiances were detected in the control group, as no cells were injected. The signals of oligopeptide-treated mice were predominantly detected in the spleen, while the signals of the vehicle-treated mice were more diffused all over the body, especially stronger in femur areas. The median radiance fold change of the vehicle group had the highest value, and the oligopeptide group was about three times lower (p=0.046) than the vehicle group (Fig. 4C). This data was divided into three ranges, 10 to less than 50, 50 to 500, and more than 500. In the lower range of fold change, there were more mice from the oligopeptide group (n=7) than the vehicle group (n=3). However, in the fold changes of 50 and higher, the number of the vehicle group mice was much larger (n=7) than the oligopeptide group mice (n=3) (Fig. 4D). All mice from the control group had fold changes of less than two. In accordance with the BLI results, the ELISA assessed M315 protein level of the peptide group showed lowering trend, compared to the vehicle group, with no significance (data not shown). This may be resulted because of the small number of mice in each group and their large variations of the M315 protein level. This observation showed that oligopeptide affects the growth of MM cells in vivo, although the detailed mechanism remains unknown.
Fig. 4. Tumor growth was reduced in the MOPC315.BM.Luc cell induced multiple myeloma mouse model. (A) A brief timeline of the animal study is shown. (B) One representative mouse from each group was chosen based on the closest radiance activity to the median value. Images taken on day 0 and day 28 are shown and compared. On day 0, BLI was taken before the oligopeptide treatment. No radiance was detected in the untreated mouse. (C) Bioluminescence radiance fold changes of each group from day 0 to day 28 are shown. Each group was comprised of 10 mice. (D) Mice numbers according to the fold changes of bioluminescence radiance are shown. Fold changes were calculated simply by dividing the final value by the initial value. *p<0.05 and ***p<0.0001 by Mann-Whitney U test. MM, multiple myeloma.

Cyclized oligopeptide has no direct effect on MOPC315.BM.Luc cell proliferation in vitro
One concern of using this oligopeptide is that it may affect the Wnt signaling pathway and stimulate cancer cell growth. In our in vivo experiment, however, we found that the tumor burdens were reduced with the treatment of the oligopeptide. To better define the effect of the oligopeptide on MM cell growth, we tested the oligopeptide in vitro, examining cell proliferation via WST assay. The MM cell population did not vary significantly in the differently treated groups (data not shown). This result demonstrated that oligopeptide does not affect MM cell growth directly, despite the MM cells exhibiting a Wnt/β-catenin signaling pathway.
These overall findings indicate that the oligopeptide affects MM cells and osteoblastic cells; it remains uncertain as to how and through what kind of mechanism this oligopeptide affects osteoblasts and MM cells. The anti-MM effect may be mediated by the indirect effects on the bone microenvironment by the cyclized oligopeptide. Variable factors in the microenvironment may contribute strongly to the MM tumor burden; however, further studies are needed.
DISCUSSION
In the present study, we showed that a cyclized oligopeptide treatment reduced tumor burden in the MOPC315.BM.Luc MM mouse model. To study the effects of this oligopeptide targeting the DKK1-LRP5/6 binding pocket in MM bone disease, DKK1 NXI motif-containing cyclized oligopeptide was developed, and a series of in vitro and in vivo experiments was performed.
Wnt signaling is well known for the osteoblast lineage specification from mesenchymal cells, further differentiation of osteoblast precursor cells, and skeletal development.24 DKK1, an endogenous Wnt inhibitor, binds to LRP5/6 with Kremen to inhibit the canonical Wnt signaling pathway. Most MM cells express DKK1;14 furthermore, the secreted DKK1 disrupts the RANKL/OPG expression ratio, which enhances osteoclast-mediated osteolysis and simultaneous osteoblast inhibition.15 As DKK1 has been a potential therapeutic target for the treatment of MM, some effective therapies targeting DKK1 have been developed, including DKK1 neutralizing antibodies, proteasome inhibitors, DKK1 vaccines, and tumor-produced endothelin-1.26 A study using the SCID-rab mouse model of myeloma showed that anti-DKK1 antibody treatment increased bone formation and reduced tumor burden in a rabbit bone implanted with MM.19 Human anti-DKK1 monoclonal antibody (BHQ880) was also found to increase osteoblasts and blocked MM cell proliferation when MM cells were cocultured with bone marrow stromal cells (BMSC).20
The cyclized oligopeptide against the DKK1-LRP5/6 binding pocket abrogated the Wnt-β-catenin signaling inhibited by DKK1 (not by sclerostin) dose dependently (IC50≈5×10-8 M). The NMR structure was obtained by complete proton resonance assignment. Isoleucine residue of the cyclized peptide should target the second pocket of LRP5/6. A ribbon diagram (Fig. 1C) showed that DKK1 is a linear peptide chain with a loop conformation.
The MM mice model we studied was developed in the Bogen lab.21 It was stated that the MM mice developed tumor burdens mostly around long bone areas and also developed tumor cells in the bone marrow. How the MM cells affect the bone metabolism or bone marrow, however, was not deeply studied. The administration of the oligopeptide abrogated the suppression of canonical Wnt signaling by DKK1 and inhibited tumor burden significantly. As matrix protein contains many growth factors and cytokines stimulating proliferation of tumor cells, inhibition of bone resorption by decreasing the RANKL/OPG ratio via the oligopeptide could reduce the release of these growth factors and cytokines from matrix proteins and inhibit the overgrowth of MOPC315.BM.Luc cells. Similar to what we have achieved, the DKK1 neutralizing antibody BHQ880 has been shown to not only improve bone formation but also to reduce tumor burden.20 Additionally, studies have reported that DKK1 suppressed β-catenin in myeloid-derived suppressor cells (a heterogeneous population of myeloid lineage immune cells in the stromal compartment) and thus inhibited tumor growth in mice.27 To explore the mechanism of the tumor burden reduction, we studied the effects of oligopeptide on proliferation of tumor cells. However, we could not find any direct effects of oligopeptide on the proliferation of tumor cells, despite the fact that DKK1 is expressed in MOPC315.BM.Luc cells (data not shown). This result was supported by a previous study showing that, while BHQ-880 did not have a direct anti-tumor effect on myeloma cells, it inhibited myeloma growth in the presence of BMSC.20 The MM niches contain bone marrow, fat cells, and immune cells, and they interact with each other. Several other cell types within the bone microenvironment produce significant amounts of DKK1, including megakaryocytes, endothelial cells, and osteoblasts, and may contribute to the regulation of tumor growth. 28,29 TGF-β, VEGF, and FGF, as well as the increase of bone resorption by osteoclasts, seem to support MM cell growth.30,31 In a previous study, DKK1 indirectly contributed to myeloma growth by regulating IL-6 in the bone microenvironment. BHQ-880 significantly reduced the production of IL-6 levels in BMSC supernatants, and the addition of IL-6 reversed the majority of growth inhibitory effects.20 Therefore, the overall results we obtained may have derived from DKK1 acting on these cells differently.
In conclusion, the cyclized oligopeptide based on the DKK1-LRP5/6 interaction abrogates the suppression of canonical Wnt signaling by DKK1. Treatment of the oligopeptide reduced tumor burden significantly in the MOPC315.BM.Luc MM mouse model. Cyclized oligopeptide may be an option for proper management of tumor burden in MM; however, further research is required to determine the details of the mechanisms involved.
ACKNOWLEDGEMENTS
MOPC315.BM.Luc cells and ab2.1-4 antibodies were obtained as generous gift from Bogen lab. This study was supported by Mid-career Research Program (NRF-2013R1A2A2A01068963) through NRF grant funded by the MEST; and Bo Mi Park, Dongdong Zhang, Chu Hyun Bae and Heeyoun Kim are recipients of Brain Korea 21 PLUS grant.
Footnotes
The authors have no financial conflicts of interest.
References
- 1.Raab MS, Podar K, Breitkreutz I, Richardson PG, Anderson KC. Multiple myeloma. Lancet. 2009;374:324–339. doi: 10.1016/S0140-6736(09)60221-X. [DOI] [PubMed] [Google Scholar]
- 2.Balakumaran A, Robey PG, Fedarko N, Landgren O. Bone marrow microenvironment in myelomagenesis: its potential role in early diagnosis. Expert Rev Mol Diagn. 2010;10:465–480. doi: 10.1586/erm.10.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ballester OF, Moscinski LC, Lyman GH, Chaney JV, Saba HI, Spiers AS, et al. High levels of interleukin-6 are associated with low tumor burden and low growth fraction in multiple myeloma. Blood. 1994;83:1903–1908. [PubMed] [Google Scholar]
- 4.Lee JW, Chung HY, Ehrlich LA, Jelinek DF, Callander NS, Roodman GD, et al. IL-3 expression by myeloma cells increases both osteoclast formation and growth of myeloma cells. Blood. 2004;103:2308–2315. doi: 10.1182/blood-2003-06-1992. [DOI] [PubMed] [Google Scholar]
- 5.Heider U, Zavrski I, Jakob C, Bängeroth K, Fleissner C, Langelotz C, et al. Expression of receptor activator of NF-kappaB ligand (RANKL) mRNA in human multiple myeloma cells. J Cancer Res Clin Oncol. 2004;130:469–474. doi: 10.1007/s00432-004-0578-3. [DOI] [PubMed] [Google Scholar]
- 6.Sati HI, Greaves M, Apperley JF, Russell RG, Croucher PI. Expression of interleukin-1beta and tumour necrosis factor-alpha in plasma cells from patients with multiple myeloma. Br J Haematol. 1999;104:350–357. doi: 10.1046/j.1365-2141.1999.01193.x. [DOI] [PubMed] [Google Scholar]
- 7.Choi SJ, Cruz JC, Craig F, Chung H, Devlin RD, Roodman GD, et al. Macrophage inflammatory protein 1-alpha is a potential osteoclast stimulatory factor in multiple myeloma. Blood. 2000;96:671–675. [PubMed] [Google Scholar]
- 8.Mukai T, Otsuka F, Otani H, Yamashita M, Takasugi K, Inagaki K, et al. TNF-alpha inhibits BMP-induced osteoblast differentiation through activating SAPK/JNK signaling. Biochem Biophys Res Commun. 2007;356:1004–1010. doi: 10.1016/j.bbrc.2007.03.099. [DOI] [PubMed] [Google Scholar]
- 9.Ehrlich LA, Chung HY, Ghobrial I, Choi SJ, Morandi F, Colla S, et al. IL-3 is a potential inhibitor of osteoblast differentiation in multiple myeloma. Blood. 2005;106:1407–1414. doi: 10.1182/blood-2005-03-1080. [DOI] [PubMed] [Google Scholar]
- 10.Boland GM, Perkins G, Hall DJ, Tuan RS. Wnt 3a promotes proliferation and suppresses osteogenic differentiation of adult human mesenchymal stem cells. J Cell Biochem. 2004;93:1210–1230. doi: 10.1002/jcb.20284. [DOI] [PubMed] [Google Scholar]
- 11.Kim JH, Liu X, Wang J, Chen X, Zhang H, Kim SH, et al. Wnt signaling in bone formation and its therapeutic potential for bone diseases. Ther Adv Musculoskelet Dis. 2013;5:13–31. doi: 10.1177/1759720X12466608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Li X, Ominsky MS, Warmington KS, Morony S, Gong J, Cao J, et al. Sclerostin antibody treatment increases bone formation, bone mass, and bone strength in a rat model of postmenopausal osteoporosis. J Bone Miner Res. 2009;24:578–588. doi: 10.1359/jbmr.081206. [DOI] [PubMed] [Google Scholar]
- 13.Fedi P, Bafico A, Nieto Soria A, Burgess WH, Miki T, Bottaro DP, et al. Isolation and biochemical characterization of the human Dkk1 homologue, a novel inhibitor of mammalian Wnt signaling. J Biol Chem. 1999;274:19465–19472. doi: 10.1074/jbc.274.27.19465. [DOI] [PubMed] [Google Scholar]
- 14.Mitsiades CS, McMillin DW, Klippel S, Hideshima T, Chauhan D, Richardson PG, et al. The role of the bone marrow microenvironment in the pathophysiology of myeloma and its significance in the development of more effective therapies. Hematol Oncol Clin North Am. 2007;21:1007–1034. doi: 10.1016/j.hoc.2007.08.007. [DOI] [PubMed] [Google Scholar]
- 15.Qiang YW, Chen Y, Stephens O, Brown N, Chen B, Epstein J, et al. Myeloma-derived Dickkopf-1 disrupts Wnt-regulated osteoprotegerin and RANKL production by osteoblasts: a potential mechanism underlying osteolytic bone lesions in multiple myeloma. Blood. 2008;112:196–207. doi: 10.1182/blood-2008-01-132134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tian E, Zhan F, Walker R, Rasmussen E, Ma Y, Barlogie B, et al. The role of the Wnt-signaling antagonist DKK1 in the development of osteolytic lesions in multiple myeloma. N Engl J Med. 2003;349:2483–2494. doi: 10.1056/NEJMoa030847. [DOI] [PubMed] [Google Scholar]
- 17.Rachner TD, Hadji P, Hofbauer LC. Novel therapies in benign and malignant bone diseases. Pharmacol Ther. 2012;134:338–344. doi: 10.1016/j.pharmthera.2012.02.005. [DOI] [PubMed] [Google Scholar]
- 18.Rachner TD, Göbel A, Benad-Mehner P, Hofbauer LC, Rauner M. Dickkopf-1 as a mediator and novel target in malignant bone disease. Cancer Lett. 2014;346:172–177. doi: 10.1016/j.canlet.2014.01.010. [DOI] [PubMed] [Google Scholar]
- 19.Yaccoby S, Ling W, Zhan F, Walker R, Barlogie B, Shaughnessy JD., Jr Antibody-based inhibition of DKK1 suppresses tumor-induced bone resorption and multiple myeloma growth in vivo. Blood. 2007;109:2106–2111. doi: 10.1182/blood-2006-09-047712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Fulciniti M, Tassone P, Hideshima T, Vallet S, Nanjappa P, Ettenberg SA, et al. Anti-DKK1 mAb (BHQ880) as a potential therapeutic agent for multiple myeloma. Blood. 2009;114:371–379. doi: 10.1182/blood-2008-11-191577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hofgaard PO, Jodal HC, Bommert K, Huard B, Caers J, Carlsen H, et al. A novel mouse model for multiple myeloma (MOPC315.BM) that allows noninvasive spatiotemporal detection of osteolytic disease. PLoS One. 2012;7:e51892. doi: 10.1371/journal.pone.0051892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Davis DG, Bax A. Assignment of complex proton NMR spectra via two-dimensional homonuclear Hartnabb-Hahn spectroscopy. J Am Chem Soc. 1985;107:2820–2821. [Google Scholar]
- 23.Jeener J, Meier BH, Bachman P, Ernst RR. Investigation of exchange processes by two-dimensional NMR spectroscopy. J Chem Phys. 1979;71:4546–4553. [Google Scholar]
- 24.Koradi R, Billeter M, Wüthrich K. MOLMOL: a program for display and analysis of macromolecular structures. J Mol Graph. 1996;14:51–55. doi: 10.1016/0263-7855(96)00009-4. [DOI] [PubMed] [Google Scholar]
- 25.Jami A, Gadi J, Lee MJ, Kim EJ, Lee MJ, Jung HS, et al. Pax6 expressed in osteocytes inhibits canonical Wnt signaling. Mol Cells. 2013;35:305–312. doi: 10.1007/s10059-013-2310-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhou F, Meng S, Song H, Claret FX. Dickkopf-1 is a key regulator of myeloma bone disease: opportunities and challenges for therapeutic intervention. Blood Rev. 2013;27:261–267. doi: 10.1016/j.blre.2013.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.D'Amico L, Capietto AH, Zamani A, Faccio R, Bumpass D. Dickkopf-realtaed protein 1 (Dkk1) exerts immune suppressive effects in cancer by regulating expansion and function of myeloid derived suppressor cells. Seattle: Paper presented at Annual Meeting of the American Society for Bone and Mineral Research; 2015. [Google Scholar]
- 28.Wong D, Winter O, Hartig C, Siebels S, Szyska M, Tiburzy B, et al. Eosinophils and megakaryocytes support the early growth of murine MOPC315 myeloma cells in their bone marrow niches. PLoS One. 2014;9:e109018. doi: 10.1371/journal.pone.0109018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Smadja DM, d'Audigier C, Weiswald LB, Badoual C, Dangles-Marie V, Mauge L, et al. The Wnt antagonist Dickkopf-1 increases endothelial progenitor cell angiogenic potential. Arterioscler Thromb Vasc Biol. 2010;30:2544–2552. doi: 10.1161/ATVBAHA.110.213751. [DOI] [PubMed] [Google Scholar]
- 30.Yata K, Yaccoby S. The SCID-rab model: a novel in vivo system for primary human myeloma demonstrating growth of CD138-expressing malignant cells. Leukemia. 2004;18:1891–1897. doi: 10.1038/sj.leu.2403513. [DOI] [PubMed] [Google Scholar]
- 31.Bisping G, Leo R, Wenning D, Dankbar B, Padró T, Kropff M, et al. Paracrine interactions of basic fibroblast growth factor and interleukin-6 in multiple myeloma. Blood. 2003;101:2775–2783. doi: 10.1182/blood-2002-09-2907. [DOI] [PubMed] [Google Scholar]