

Abstract
Inflammatory Bowel Disease (IBD) is a kind of chronic inflammation, which has increasing incidence and prevalence in recent years. IBD mainly divides into Crohn's disease (CD) and ulcerative colitis (UC). It is hard to cure IBD completely, and novel therapies are urgently needed. Amino acids (AAs) and their metabolites are regarded as important nutrients for humans and animals and also play an important role in IBD amelioration. In the present study, the potential protective effects of AAs and their metabolites on IBD had been summarized with the objective to provide insights into IBD moderating using dietary AAs and their metabolites as a potential adjuvant therapy.
1. Inflammatory Bowel Disease Prevalence and Increasing Incidence
Inflammatory bowel disease (IBD) is a kind of chronically multifactorial inflammatory disorder affecting the gastrointestinal tract [1]. It is triggered as a consequence of excessive proinflammatory cytokines production, persistent macrophage activation, and cell death induced by subsequent bacterial or/and viral infections. IBD is the major intestinal health concerns causing severe diarrhea, abdominal pain, weight loss, metabolic disorder, and malabsorption, and it mainly comprises two major forms: Crohn's disease (CD) and ulcerative colitis (UC) [2, 3]. CD and UC have different inflammatory location in the gastrointestinal tract. CD affects all layers of the gastrointestinal tract and is associated with excess expression of IL-12/IL-23 and IFN-γ/IL-17, while UC mainly occurs in colon affecting the mucosa with primarily excess production of IL-13 [3, 4]. Currently, the incidence and prevalence of IBD are increasing with time around the world, especially the rate within elderly patients [5]. The developed countries (Europe and North America) present the highest occurrence [6]. In the West, the prevalence of IBD is 37 to 249 cases per 100,000 people for UC and from 26 to 319 cases per 100,000 people for CD [6, 7]. By contrast, Eastern Europe, Asia, and other developing countries have a lower IBD incidence [7–9]. The occurrence of UC in Asian countries is higher than that of CD [10].
The pathogenesis of IBD is complex and still unclear. Mounting evidences suggest that IBD is the consequence of abnormal immune regulation induced by genetic and/or environmental factors (e.g., diet, infection) [1, 11]. Various inflammation mediators including reactive oxygen species and proinflammatory cytokines (NF-κB, cytochrome c, and tumor necrosis factor- (TNF-)-α) act as predisposing factors for chronic inflammatory diseases [12]. The typical clinical inflammation of the gastrointestinal tract leads to IBD with excessive production of proinflammatory mediators and loss of the intestinal mucosal barrier integrity. Since there is no complete cure for IBD, the therapies of IBD primarily focus on inducing or maintaining remission and promoting the expression of anti-inflammatory genes [3].
At the present, the conventional treatments of IBD such as biological therapies targeting specific cytokines or pathways and clinical medication have been improved in recent years. Mesalamine, antibiotics, and budesonide are used in patients with mild disease status, while patients with moderately severe IBD take systemic corticosteroids, immune-modulators (thiopurine analogues, azathioprine (AZA), 6-mercaptopurine (6-MP), and methotrexate), and anti-TNFα agents (infliximab, adalimumab, and certolizumab pegol) [13]. However, proper administration selection becomes a common clinical dilemma with those different kinds of medicines. The conventional treatments in severe IBD have a short-term favorable prognosis, while there is still a challenge in the development of alternative therapies character with low-risk and long-term outcomes [14]. Besides, treatments mentioned above are proved to have limited efficacy, adverse effects, drug interactions, and potential toxicity [15, 16]. The use of AZA and 6-MP in IBD patients leads to serious adverse drug reactions such as hepatotoxicity, pancreatitis, and gastrointestinal disturbances [17]. Immunosuppressive and anti-TNF therapy in IBD cause dermatological adverse effects, including skin infections, drug hypersensitivity, psoriasis, eczema, and nonmelanomatous skin cancer [18, 19].
In most cases, patients suffering IBD usually face long duration which becomes a considerable economic burden. With the development of medical and life science, a large number of new strategies are provided to treat IBD. Functional nutrients have aroused growing interest, which help prevent or remit malnutrition, moderate the mucosal immune response, and benefit intestinal homeostasis [20–22]. Essential nutrient has the potential to ameliorate restore redox balance and inflammation in the gastrointestinal tract showing the possibility of nutrients in IBD treatment [3]. Part of the nutrients that benefit the management of IBD had been listed in Table 1.
Table 1.
Studies on dietary nutrients supplementation for the management of IBD.
| Nutrients | Primary components | Chemical structure | Functions | Ref. |
|---|---|---|---|---|
| Corabion | A mixture of vitamin C, vitamin E, ω3-PUFAs (EPA and DHA), and Arg | Reduction of DAI, neutrophil recruitment, oxidative stress, proinflammatory cytokines, and E-cadherin internalization; attenuation of colon shortening and tissue damage | [23] | |
|
| ||||
| Pomegranate extract | Ellagic acid |
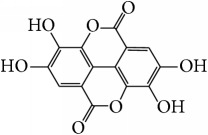
|
Reduction of MPO activity, TNF-α levels, (COX-2), iNOS overexpression, and MAPKs phosphorylation; preventing NF-κB translocation | [24, 25] |
|
| ||||
| Krill oil | ω3-PUFAs and phospholipids | Reduction of DAI, HCS, colon length, and protein oxidation markers; improvement of Pparg1α and (PG)E3 expression | [26] | |
|
| ||||
| Tetradecylthioacetic acid | An artificial 16-carbon fatty acid with a sulphur-substitution in the β-position |
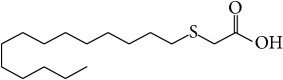
|
Reduction of colonic oxidative damage and colon wall thickness; improvement of expression of Pparg1α; inhibition of the production of inflammatory cytokines (TNF-α, IL-1β, and IL-6) | [27] |
|
| ||||
| Fibre | Nonstarch polysaccharides, resistant oligosaccharides, analogous carbohydrates, and lignin | Maintain remission and reduce lesions of the intestinal mucosa | [28, 29] | |
|
| ||||
| Anthocyanins | Natural agents derived from strawberry, blueberry, barberry, and other plants |
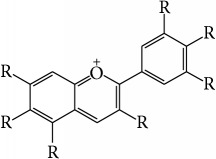
|
Cytoprotection; remission of oxidative stress and inflammatory cytokines; suppression of cellular signaling pathways of inflammatory processes | [30] |
|
| ||||
| α-Linolenic acid | ω3-PUFA: plant-derived oil |
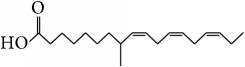
|
Inhibition of the production of IL-6 and TNF-α; reduction of cell apoptosis, intestinal permeability, and bacterial translocation; improving histological repair | [31, 32] |
|
| ||||
| FAAH inhibitors | PF-3845 |
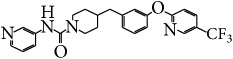
|
Possess anti-inflammatory effect in TNBS-induced colitis in mice; alter the levels of endocannabinoids | [33] |
| FAAH-II |
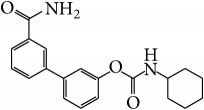
|
Inhibition of inflammatory miRNAs and cytokines; reduction of the number of activated T cells, the frequency of macrophages, and neutrophils in the colon | [34] | |
EPA, eicosapentaenoic acid; DHA, docosahexaenoic acid; (COX-2), cyclooxygenase-2; iNOS, inducible nitric oxide synthase; MAPKs, mitogen-activated protein kinases; MPO, myeloperoxidase; NF-κB, nuclear transcription factor kappa B; PUFAs, polyunsaturated fatty acids; DAI, disease activity index; HCS, histological combined score; Pparg1a, PPAR-g coactivator 1α; (PG)E3, prostaglandin-E3; IL-1β, interleukin-1β; PUFA, polyunsaturated fatty acids; FAAH, fatty acid amide hydrolase.
2. Amino Acids: Application in Inflammatory Bowel Disease Therapies
Among IBD benefit nutrients, amino acids (AAs) act as the key regulatory factor in metabolic pathways controlling and have important effects on keeping the intestinal health. AAs are considered as the building blocks for protein synthesis and also play major roles in other functions, such as cell signaling, gene expression, intracellular protein turnover, maintenance, reproduction, oxidative stress, and immunity [46, 47]. Systemic inflammation can cause malnutrition symptoms and general glutamine (Gln) deprivation which is associated with depression, muscle loss, and emotional fatigue [48]. Both UC and CD disturb AAs metabolism in serum and plasma by increasing levels of isoleucine (Ile) (and its first degradation product 3-methyl-2-oxovalerate), methionine (Met), lysine, glycine (Gly), arginine (Arg), and proline (Pro), while decreasing levels of valine, tyrosine, and serine [49]. Some of the increased AAs were also reported to be increased in fecal extracts [50], while Ile and leucine (Leu) have apparently low concentrations in colonic mucosa of active IBD [51]. Met is an essential amino acid and a precursor of homocysteine, a metabolite shown highly elevated in both plasma and colonic mucosa from IBD patients [52].
AAs also have trophic and cytoprotective effects on health in humans and animals [53, 54]. T cells are regarded as central effectors of the adaptive immune system. T helper (Th) cells (Th1, Th2, and Th 17 cells) differentiate from native CD4+ T cells and are involved in the pathogenesis of several inflammatory immune-mediated disorders such as producing different cytokines in immune responses [55]. CD is a Th1-type T cell-mediated inflammation while UC is a Th2-type T cell-mediated inflammation [4]. AAs directly serve as a fuel source for T cells and are considered to have influence on shaping T cell-mediated immune responses [56]. Additionally, the signaling central integrator of environmental stimuli for the regulation of T cell activation and differentiation is mammalian target of rapamycin (mTOR) [57, 58]. mTOR have two distinct protein complexes, mTOR complex 1 (mTORC1) and mTOR complex 2 (mTORC2). In T cells, AAs work as signaling molecules with mTORC1 acting as a key mediator. AAs regulate the intracellular localization and activation of mTORC1 by the lysosome-based signaling system composed of Ras-related GTPases (Rags) and Regulator v-ATPase, GAP activity towards Rags, and folliculin complexes [59, 60]. AAs could protect whole body and muscle from protein loss via mTOR activation and downstream signaling to protein synthesis through mTORC1 in the acute phase of inflammation [61, 62]. Leu-enriched diet accelerated recovery from muscle damage by alleviating excessive expression of proinflammatory cytokine and preventing invasion of inflammatory cells into muscle [63]. Otherwise, general controlled nonrepressed (GCN2) kinase is a key orchestrator of the integrated stress response which senses AAs depletion. Acute AAs starvation in mice protects the symptoms of colitis, limits Th17 cells, and suppressed IBD via a GCN2-dependent mechanism, providing another mechanism of AAs in regulating IBD [64].
In conclusion, growing evidence shows that the anti-inflammatory activities of tryptophan (Trp), Gln, Met, cysteine (Cys), and Arg have been well established, suggesting a therapeutic role of AAs in IBD, which were listed in Table 2. It is necessary to illustrate the biological activity of specific immunomodulatory AAs in IBD.
Table 2.
Application of partial AAs in inflammatory bowel disease therapies.
| AAs | Dose | Administration method | Models | Major functions and effects | Ref. |
|---|---|---|---|---|---|
| Tryptophan | 88 mg/kg·BW·day | Intragastric infusion | Piglets | Amelioration of clinical symptoms; reduction of gut permeability and cell apoptosis; inhibition of the production of inflammatory cytokines (TNF-α, IL-6, IFN-γ, IL-12p40, IL-1β, and ICAM-1) | [35] |
|
| |||||
| Melatonin | 20 mg/kg·BW·day | Intragastric infusion | Rats | Antioxidant; inhibition of the production of inflammatory cytokines (TNF-α, COX-2, SOD, and NF-κB); accelerating healing of gastric ulcer | [36] |
|
| |||||
| Glutamine | 25% of the total nitrogen | Dietary | Mice | Anti-inflammation; reduction of expression of PSGL-1, LFA-1, and CCR9 by Th cells | [37] |
|
| |||||
| Glutamate | 12 μg Glu/0.3 μL saline | Microinjection | Rats | Neurotransmitter; inhibition of T-cell response and inflammation | [38] |
|
| |||||
| Methionine | 0.12% L-Met | Dietary | Piglets | Protection of the small-intestinal mucosa | [39] |
|
| |||||
| Cysteine | 144 mg/kg·BW·day | Intragastric infusion | Piglets | Reduction of intestinal permeability and cell apoptosis; inhibition of the local expression of inflammatory mediators (IL-6, TNF-α, IL-12p40, and IL-1β) | [40] |
|
| |||||
| Histidine | Not mentioned | Dietary | Mice | Reduction of histologic damage, colon weight, IL-6, and TNF-α production; inhibition of NF-κB | [41] |
|
| |||||
| Arginine | 1% (wt/vol) solution | Drinking water | Mice | Improvement of the clinical parameters and body weight loss; reduction of the colonic permeability; reduction of the proinflammatory cytokine and chemokine expression | [42] |
|
| |||||
| Glycine | 5% Gly | Dietary | Rats | Diarrhea amelioration; prevention of the increases of IL-1β and TNF-α | [43] |
|
| |||||
| Taurine | 30 mM | Rectal administration | Rats | Anti-inflammation; inhibiting NF-κB activity | [44] |
| 2% | Drinking water | Mice | Inhibitory effects on the secretion of MIP-2; cytoprotective functions on the epithelial barrier | [45] | |
TNF-α, tumor necrosis factor-α; IL, interleukin; IFN-γ, interferon-γ; ICAM-1, intracellular adhesion molecule-1; PSGL-1, P-selectin glycoprotein ligand-1; LFA-1, leukocyte function-associated antigen-1; CCR9, C-C chemokine receptor type 9; MIP-2, macrophage inflammatory protein 2.
2.1. Tryptophan
Trp is an essential AA for human and animals and plays an important role in inflammatory regulation beyond building block for proteins synthesis [65]. The serious Trp concentration shows a highly inverse correlation with disease activity in IBD patients [66]. Dietary supplementation of Trp can reduce IBD through its immune-regulatory metabolites [35, 36]. In a porcine model of dextran-sodium sulfate- (DSS-) induced colitis, Trp administration at 80% of the daily recommended intake could ameliorate colitis clinical symptoms, improve histological parameters and intestinal permeability, reduce the expression of proinflammatory cytokines, and increase the expression of proapoptotic genes, which is important for maintaining gut homeostasis [35]. Trp presents strong anti-inflammation activity by triggering calcium-sensing receptor (CaSR) activation in intestine [67], which is a sensing receptor for dietary nutrients in the gastrointestinal tract to maintain mucosal immune homeostasis. Trp treatment allosterically activate CaSR which significantly reduces TNF-α-induced interleukin- (IL-) 8 secretion indicating a novel therapy in intestinal inflammation [67].
In addition, Trp exerts anti-inflammatory function via the metabolites including serotonin (5-hydroxytryptamine, 5-HT) and melatonin (MT). 5-HT is an important compound derived from Trp which serves as a neurotransmitter and inhibiter of the production of inflammatory cytokines and superoxide [46]. MT is abundant in gastrointestinal tract [68, 69] and has a positive impact on IBD with no or negligible side effects due to regulation of macrophage activity, reduction of cell migration and myeloperoxidase activity, and inhibition of NF-κB and TNF-α expression [36, 68, 70]. MT added to omeprazole treatment obviously accelerates chronic gastroduodenal ulcers over the obtained with omeprazole alone [71]. Additionally, in mice with DSS-induced colitis, MT exerts anti-inflammatory effects by alleviating the severity of mucosal injury and recovering the expression of IL-6, IL-17, and adiponectin [72].
2.2. Glutamine and Glutamate
As an abundant AA in the blood and tissues, Gln is mostly used as nitrogen source and/or alternative energy fuel [73]. In various clinical situations, appropriate exogenous Gln supply is safe and can beneficially contribute to diminishing risks of high-dose chemotherapy and radiation [74]. It has been additionally implicated as an immunomodulatory nutrient [75] and has pharmacological function in the IBD treatment [76]. Gln plays a key role in maintaining the integrity of the intestinal mucosa and has been shown to reduce inflammation and relieve CD [77]. In an UC mouse model, dietary Gln supplementation combined with dietary fiber and oligosaccharide has suppressive effects on mucosal damage [78]. A Gln diet replacing 25% of the total nitrogen decreases the expression of chemokine and endothelial adhesion molecules via suppression of T cell migration in mice [37]. These experimental data suggest Gln as a potential nutrient in protecting intestinal integrity and modulating immunity.
Glutamate (Glu) is produced from Gln with the catalysis of glutaminase and generally plays roles in protein synthesis and energy metabolism. Dietary Glu can also function as a signal to regulate the gastrointestinal tract via the gut-brain axis [9]. As a precursor of glutathione (GSH), Arg and Pro, Glu is of critical importance in intestinal metabolism and physiology [53]. Microinjection of Glu into the hypothalamic paraventricular nucleus in UC rats significantly increases the cell proliferation and antioxidant levels and decreases apoptosis and the expression of proinflammatory factors in the colonic mucosa [38]. Poly-γ-glutamate (P-Glu) significantly reduced histopathological evidence of injury, attenuated DSS-induced blood vessel densities, and attenuated DSS-induced expression of VEGF-A and its receptor in C57BL/6 mouse colitis model. P-Glu has potential application in conditions marked by inflammatory-driven angiogenesis and mucosal inflammation [79]. These findings above indicate that dietary supplementation of Gln and Glu is of functional and nutritional importance in intestinal mucosal growth and gut inflammation.
2.3. Sulfur Amino Acids
Sulfur amino acids (SAAs) mainly contain Met, Cys, and cystine. SAAs metabolism mainly takes place in the gastrointestinal tract. Dietary deficiency of SAAs will suppress intestinal mucosal growth, reduce intestinal epithelial cell proliferation, and increase intestinal oxidant stress in piglets [80].
As essential AA, Met has been considered as the first and second or third limiting AA in poultry and nursery pigs, respectively. Piglets fed the diet supplemented with Met present increased growth performance and exhibited improved integrity and barrier function of the small-intestinal mucosa [39]. Dietary supplementation with Met metabolites also can affect the susceptibility to colitis, reduce inflammation and tissue injury, and decrease the expression of multiple inflammatory genes in mice [81]. It is also interesting that Met (twice NRC recommendation) combined with fish oil (2.5%) can enhance immune response in IBD challenged broiler chickens which may be a novel treatment in IBD therapy for poultry [82].
Cys is a nonessential AA playing roles in protein metabolism and is regarded as the key factor in the synthesis of GSH. Dietary supplementation with N-acetyl-L-cysteine improves the clinical symptoms and decreases the chemokines without any side effects [83]. Cys administration can attenuate DSS-induced weight loss and intestinal permeability, decrease the expression of proinflammatory cytokines, and restore susceptibility of activated immune cells to apoptosis, indicating Cys supplementation as a novel therapy for IBD [40]. In addition, γ-glutamyl cysteine treatment can ameliorate DSS-induced clinical signs and histological damage via activating CaSR [84].
Besides, GSH, taurine (Tau), and hydrogen sulfide (H2S), the products of catabolism of SAAs, play major roles in anti-inflammation and antioxidant system [46]. Tau is regarded as an antioxidant and membrane stabilizer against oxidative stress and inflammation by inhibiting chemokine secretion from intestinal cells [85, 86]. Moreover, the cellular metabolite taurine chloramine of Tau in the rectal shows anti-inflammatory property on IBD via inhibition on NF-κB activity [44, 87]. Tau treatment exerts beneficial effects in rats with 2,4,6-trinitrobenzene sulphonic acid- (TNBS-) induced colitis with decreasing inflammatory reactions and apoptosis [88]. H2S is a signaling molecule and a gaseous mediator that exhibits several anti-inflammatory activities and contributes to mucosal protection [89–91]. One of the possible mechanisms of H2S in the resolution of IBD is proved to be mediated via stabilization of hypoxia-inducible factor-1α [92].
2.4. Arginine
Arg is a semiessential AA that has protective effects against oxidative stress. As the substrate for nitric oxide (NO) synthesis, amino acid Arg is reported to be therapeutic in wound healing and has potent anti-inflammatory properties as a mediator of autoimmune diseases [93–96]. Otherwise, exogenous Arg is associated with antiapoptotic effects on the rat intestine and useful in the treatment of intestinal ischemia/reperfusion injury [97]. Dietary supplementation with Arg can improve the immune status of humans and animals and has the potential to be used to supplement current treatments for IBD [12]. Serum Arg concentration is a useful biomarker of UC disease severity [98]. Further, in a DSS-induced fulminant colitis murine model, treatment with hepatocyte growth factor and Arg can decline associated symptoms such as pain and diarrhea [99]. Arg is absorbed and transferred by cationic AA transporters (CAT) in intestine. DSS-induced inflammation reduced the expression of CAT2 in colonic and Arg uptake with body weight loss, reducing colonic permeability. Supplementation with Arg markedly attenuates the clinical parameters above and reduces the expression of proinflammatory cytokine and chemokine [42]. Arg might reduce the inflammation associated with AA-induced colitis through the NF-κB/nitric oxide pathway [100]. NO participates in nutrient metabolism and exerts protective effects against IBD including inhibition of macrophage activation and proinflammatory cytokine levels [101].
2.5. Other AAs
Besides the functional AAs above, other AAs have been reported to possess anti-inflammation functions to some extent. In addition to acting as an important precursor for the biosynthesis of GSH, Gly is proved to ameliorate diarrhea and body weight loss in TNBS induced colitis in the rats, indicating that Gly may be a useful immunomodulating nutrient for the treatment of IBD [43]. Histidine has proven to be a novel therapeutic agent for CD by inhibition of NF-κB activation, downregulating proinflammatory cytokine production in IL10−/− mice [41]. Ergothioneine (EGT) is a natural water-soluble amino acid which can be derived from mushroom or synthesized by nonyeast fungi [102, 103]. In ultraviolet-B-irradiated mice, the administration of EGT inhibited the UV-B-induced inflammatory responses and DNA halogenation, showing the modulatory effects of EGT in inflammation [103].
3. Future Perspectives and Challenge
Lacking of effective medical therapies for IBD makes it of utmost importance to find alternative therapeutic strategies [3]. AAs can relieve intestinal inflammation through regulation of proinflammatory cytokines suggesting a possible approach to IBD treatment. However, further investigations and clinical studies are needed to fully understand the therapeutic mechanism and potential of AAs in preventing inflammation in both humans and animals.
Acknowledgments
This project was supported by the Chinese Academy of Sciences STS Project (KFJ-SW-STS-173), the National Basic Research Program of China (973 Program) (2013CB127301), and the National “948” Project from the Ministry of Agriculture of China (2015Z74).
Disclosure
The funding sources played no role in the study design, data collection and interpretation, or the decision to submit the work for publication.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
References
- 1.Corridoni D., Arseneau K. O., Cominelli F. Inflammatory bowel disease. Immunology Letters. 2014;161(2):231–235. doi: 10.1016/j.imlet.2014.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Himmel M. E., Hardenberg G., Piccirillo C. A., Steiner T. S., Levings M. K. The role of T-regulatory cells and Toll-like receptors in the pathogenesis of human inflammatory bowel disease. Immunology. 2008;125(2):145–153. doi: 10.1111/j.1365-2567.2008.02939.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhang H., Hu C.-A. A., Kovacs-Nolan J., Mine Y. Bioactive dietary peptides and amino acids in inflammatory bowel disease. Amino Acids. 2015;47(10):2127–2141. doi: 10.1007/s00726-014-1886-9. [DOI] [PubMed] [Google Scholar]
- 4.Strober W., Fuss I., Mannon P. The fundamental basis of inflammatory bowel disease. The Journal of Clinical Investigation. 2007;117(3):514–521. doi: 10.1172/jci30587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Taleban S. Challenges in the diagnosis and management of inflammatory bowel disease in the elderly. Current Treatment Options in Gastroenterology. 2015;13(3):275–286. doi: 10.1007/s11938-015-0059-6. [DOI] [PubMed] [Google Scholar]
- 6.Molodecky N. A., Soon I. S., Rabi D. M., et al. Increasing incidence and prevalence of the inflammatory bowel diseases with time, based on systematic review. Gastroenterology. 2012;142(1):46–54.e42. doi: 10.1053/j.gastro.2011.10.001. [DOI] [PubMed] [Google Scholar]
- 7.Loftus E. V., Jr. Clinical epidemiology of inflammatory bowel disease: incidence, prevalence, and environmental influences. Gastroenterology. 2004;126(6):1504–1517. doi: 10.1053/j.gastro.2004.01.063. [DOI] [PubMed] [Google Scholar]
- 8.Ng S. C., Tang W., Ching J. Y., et al. Incidence and phenotype of inflammatory bowel disease based on results from the Asia-Pacific Crohn's and colitis epidemiology study. Gastroenterology. 2013;145(1):158–165.e2. doi: 10.1053/j.gastro.2013.04.007. [DOI] [PubMed] [Google Scholar]
- 9.Lovasz B. D., Golovics P. A., Vegh Z., Lakatos P. L. New trends in inflammatory bowel disease epidemiology and disease course in Eastern Europe. Digestive and Liver Disease. 2013;45(4):269–276. doi: 10.1016/j.dld.2012.08.020. [DOI] [PubMed] [Google Scholar]
- 10.Yang H., Li Y., Wu W., et al. The incidence of inflammatory bowel disease in Northern China: A Prospective Population-based Study. PLOS ONE. 2014;9(7) doi: 10.1371/journal.pone.0101296.e101296 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fiocchi C. Inflammatory bowel disease: etiology and pathogenesis. Gastroenterology. 1998;115(1):182–205. doi: 10.1016/s0016-5085(98)70381-6. [DOI] [PubMed] [Google Scholar]
- 12.Al-Drees A. M., Khalil M. S. Histological and immunohistochemical effects of L-arginine and silymarin on TNBS-induced inflammatory bowel disease in rats. Histology and Histopathology. 2016;31(11):1259–1270. doi: 10.14670/HH-11-757. [DOI] [PubMed] [Google Scholar]
- 13.Dassopoulos T., Sultan S., Falck-Ytter Y. T., Inadomi J. M., Hanauer S. B. American gastroenterological association institute technical review on the use of thiopurines, methotrexate, and anti-TNF-α biologic drugs for the induction and maintenance of remission in inflammatory Crohn's disease. Gastroenterology. 2013;145(6):1464–1478.e5. doi: 10.1053/j.gastro.2013.10.046. [DOI] [PubMed] [Google Scholar]
- 14.Cöeffier M., Marion-Letellier R., Déchelotte P. Potential for amino acids supplementation during inflammatory bowel diseases. Inflammatory Bowel Diseases. 2010;16(3):518–524. doi: 10.1002/ibd.21017. [DOI] [PubMed] [Google Scholar]
- 15.De Silva S., Devlin S., Panaccione R. Optimizing the safety of biologic therapy for IBD. Nature Reviews Gastroenterology and Hepatology. 2010;7(2):93–101. doi: 10.1038/nrgastro.2009.221. [DOI] [PubMed] [Google Scholar]
- 16.Gearry R. B., Barclay M. L., Burt M. J., Collett J. A., Chapman B. A. Thiopurine drug adverse effects in a population of New Zealand patients with inflammatory bowel disease. Pharmacoepidemiology and Drug Safety. 2004;13(8):563–567. doi: 10.1002/pds.926. [DOI] [PubMed] [Google Scholar]
- 17.Schwab M., Schäffeler E., Marx C., et al. Azathioprine therapy and adverse drug reactions in patients with inflammatory bowel disease: impact of thiopurine S-methyltransferase polymorphism. Pharmacogenetics. 2002;12(6):429–436. doi: 10.1097/00008571-200208000-00003. [DOI] [PubMed] [Google Scholar]
- 18.Moran G. W., Lim A. W. K., Bailey J. L., et al. Review article: dermatological complications of immunosuppressive and anti-TNF therapy in inflammatory bowel disease. Alimentary Pharmacology and Therapeutics. 2013;38(9):1002–1024. doi: 10.1111/apt.12491. [DOI] [PubMed] [Google Scholar]
- 19.Torres J., Buche S., Delaporte E., Colombel J.-F. Skin side effects of inflammatory bowel disease therapy. Inflammatory Bowel Diseases. 2013;19(5):1086–1098. doi: 10.1097/MIB.0b013e3182802c07. [DOI] [PubMed] [Google Scholar]
- 20.Neuman M. G., Nanau R. M. Inflammatory bowel disease: role of diet, microbiota, life style. Translational Research. 2012;160(1):29–44. doi: 10.1016/j.trsl.2011.09.001. [DOI] [PubMed] [Google Scholar]
- 21.Ruemmele F. M. Role of diet in inflammatory bowel disease. Annals of Nutrition and Metabolism. 2016;68, supplement 1:33–41. doi: 10.1159/000445392. [DOI] [PubMed] [Google Scholar]
- 22.Altomare R., Damiano G., Abruzzo A., et al. Enteral nutrition support to treat malnutrition in inflammatory bowel disease. Nutrients. 2015;7(4):2125–2133. doi: 10.3390/nu7042125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Vargas Robles H., Citalán Madrid A. F., García Ponce A., et al. Experimental colitis is attenuated by cardioprotective diet supplementation that reduces oxidative stress, inflammation, and mucosal damage. Oxidative Medicine and Cellular Longevity. 2016;2016:9. doi: 10.1155/2016/8473242.8473242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rosillo M. A., Sánchez-Hidalgo M., Cárdeno A., et al. Dietary supplementation of an ellagic acid-enriched pomegranate extract attenuates chronic colonic inflammation in rats. Pharmacological Research. 2012;66(3):235–242. doi: 10.1016/j.phrs.2012.05.006. [DOI] [PubMed] [Google Scholar]
- 25.Ismail T., Sestili P., Akhtar S. Pomegranate peel and fruit extracts: a review of potential anti-inflammatory and anti-infective effects. Journal of Ethnopharmacology. 2012;143(2):397–405. doi: 10.1016/j.jep.2012.07.004. [DOI] [PubMed] [Google Scholar]
- 26.Grimstad T., Bjørndal B., Cacabelos D., et al. Dietary supplementation of krill oil attenuates inflammation and oxidative stress in experimental ulcerative colitis in rats. Scandinavian Journal of Gastroenterology. 2012;47(1):49–58. doi: 10.3109/00365521.2011.634025. [DOI] [PubMed] [Google Scholar]
- 27.Bjørndal B., Grimstad T., Cacabelos D., et al. Tetradecylthioacetic acid attenuates inflammation and has antioxidative potential during experimental colitis in rats. Digestive Diseases and Sciences. 2013;58(1):97–106. doi: 10.1007/s10620-012-2321-2. [DOI] [PubMed] [Google Scholar]
- 28.Pituch-Zdanowska A., Banaszkiewicz A., Albrecht P. The role of dietary fibre in inflammatory bowel disease. Przeglad Gastroenterologiczny. 2015;10(3):135–141. doi: 10.5114/pg.2015.52753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wong C., Harris P. J., Ferguson L. R. Potential benefits of dietary fibre intervention in inflammatory bowel disease. International Journal of Molecular Sciences. 2016;17(6, article E919) doi: 10.3390/ijms17060919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Sodagari H. R., Farzaei M. H., Bahramsoltani R., Abdolghaffari A. H., Mahmoudi M., Rezaei N. Dietary anthocyanins as a complementary medicinal approach for management of inflammatory bowel disease. Expert Review of Gastroenterology and Hepatology. 2015;9(6):807–820. doi: 10.1586/17474124.2015.1002086. [DOI] [PubMed] [Google Scholar]
- 31.Reifen R., Karlinsky A., Stark A. H., Berkovich Z., Nyska A. α-Linolenic acid (ALA) is an anti-inflammatory agent in inflammatory bowel disease. Journal of Nutritional Biochemistry. 2015;26(12):1632–1640. doi: 10.1016/j.jnutbio.2015.08.006. [DOI] [PubMed] [Google Scholar]
- 32.Generoso S. V., Rodrigues N. M., Trindade L. M., et al. Dietary supplementation with omega-3 fatty acid attenuates 5-fluorouracil induced mucositis in mice. Lipids in Health and Disease. 2015;14(1, article 54) doi: 10.1186/s12944-015-0052-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sałaga M., Mokrowiecka A., Zakrzewski P. K., et al. Experimental colitis in mice is attenuated by changes in the levels of endocannabinoid metabolites induced by selective inhibition of fatty acid amide hydrolase (FAAH) Journal of Crohn's and Colitis. 2014;8(9):998–1009. doi: 10.1016/j.crohns.2014.01.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Shamran H., Singh N. P., Zumbrun E. E., et al. Fatty acid amide hydrolase (FAAH) blockade ameliorates experimental colitis by altering microRNA expression and suppressing inflammation. Brain, Behavior, and Immunity. 2016;59:10–20. doi: 10.1016/j.bbi.2016.06.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Kim C. J., Kovacs-Nolan J. A., Yang C., Archbold T., Fan M. Z., Mine Y. L-Tryptophan exhibits therapeutic function in a porcine model of dextran sodium sulfate (DSS)-induced colitis. Journal of Nutritional Biochemistry. 2010;21(6):468–475. doi: 10.1016/j.jnutbio.2009.01.019. [DOI] [PubMed] [Google Scholar]
- 36.Konturek P. C., Burnat G., Brzozowski T., Zopf Y., Konturek S. J. Tryptophan free diet delays healing of chronic gastric ulcers in rat. Journal of Physiology and Pharmacology. 2008;59(2):53–65. [PubMed] [Google Scholar]
- 37.Hou Y.-C., Wu J.-M., Wang M.-Y., et al. Glutamine supplementation attenuates expressions of adhesion molecules and chemokine receptors on T cells in a murine model of acute colitis. Mediators of Inflammation. 2014;2014:14. doi: 10.1155/2014/837107.837107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Li T.-T., Zhang J.-F., Fei S.-J., et al. Glutamate microinjection into the hypothalamic paraventricular nucleus attenuates ulcerative colitis in rats. Acta Pharmacologica Sinica. 2014;35(2):185–194. doi: 10.1038/aps.2013.140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chen Y., Li D., Dai Z., et al. L-Methionine supplementation maintains the integrity and barrier function of the small-intestinal mucosa in post-weaning piglets. Amino Acids. 2014;46(4):1131–1142. doi: 10.1007/s00726-014-1675-5. [DOI] [PubMed] [Google Scholar]
- 40.Kim C. J., Kovacs-Nolan J., Yang C., Archbold T., Fan M. Z., Mine Y. L-cysteine supplementation attenuates local inflammation and restores gut homeostasis in a porcine model of colitis. Biochimica et Biophysica Acta - General Subjects. 2009;1790(10):1161–1169. doi: 10.1016/j.bbagen.2009.05.018. [DOI] [PubMed] [Google Scholar]
- 41.Andou A., Hisamatsu T., Okamoto S., et al. Dietary histidine ameliorates murine colitis by inhibition of proinflammatory cytokine production from macrophages. Gastroenterology. 2009;136(2):564–574.e2. doi: 10.1053/j.gastro.2008.09.062. [DOI] [PubMed] [Google Scholar]
- 42.Coburn L. A., Gong X., Singh K., et al. L-arginine supplementation improves responses to injury and inflammation in dextran sulfate sodium colitis. PLoS ONE. 2012;7(3) doi: 10.1371/journal.pone.0033546.e33546 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tsune I., Ikejima K., Hirose M., et al. Dietary glycine prevents chemical-induced experimental colitis in the rat. Gastroenterology. 2003;125(3):775–785. doi: 10.1016/S0016-5085(03)01067-9. [DOI] [PubMed] [Google Scholar]
- 44.Kim H., Jeon H., Kong H., et al. A molecular mechanism for the anti-inflammatory effect of taurine-conjugated 5-aminosalicylic acid in inflamed colon. Molecular Pharmacology. 2006;69(4):1405–1412. doi: 10.1124/mol.105.020578. [DOI] [PubMed] [Google Scholar]
- 45.Zhao Z., Satsu H., Fujisawa M., et al. Attenuation by dietary taurine of dextran sulfate sodium-induced colitis in mice and of THP-1-induced damage to intestinal Caco-2 cell monolayers. Amino Acids. 2008;35(1):217–224. doi: 10.1007/s00726-007-0562-8. [DOI] [PubMed] [Google Scholar]
- 46.Wu G. Amino acids: metabolism, functions, and nutrition. Amino Acids. 2009;37(1):1–17. doi: 10.1007/s00726-009-0269-0. [DOI] [PubMed] [Google Scholar]
- 47.Terrill J. R., Duong M. N., Turner R., et al. Levels of inflammation and oxidative stress, and a role for taurine in dystropathology of the golden retriever muscular dystrophy dog model for duchenne muscular dystrophy. Redox Biology. 2016;9:276–286. doi: 10.1016/j.redox.2016.08.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Schlemmer M., Suchner U., Schäpers B., et al. Is glutamine deficiency the link between inflammation, malnutrition, and fatigue in cancer patients? Clinical Nutrition. 2015;34(6):1258–1265. doi: 10.1016/j.clnu.2014.12.021. [DOI] [PubMed] [Google Scholar]
- 49.Schicho R., Shaykhutdinov R., Ngo J., et al. Quantitative metabolomic profiling of serum, plasma, and urine by 1H NMR spectroscopy discriminates between patients with inflammatory bowel disease and healthy individuals. Journal of Proteome Research. 2012;11(6):3344–3357. doi: 10.1021/pr300139q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Marchesi J. R., Holmes E., Khan F., et al. Rapid and noninvasive metabonomic characterization of inflammatory bowel disease. Journal of Proteome Research. 2007;6(2):546–551. doi: 10.1021/pr060470d. [DOI] [PubMed] [Google Scholar]
- 51.Balasubramanian K., Kumar S., Singh R. R., et al. Metabolism of the colonic mucosa in patients with inflammatory bowel diseases: an in vitro proton magnetic resonance spectroscopy study. Magnetic Resonance Imaging. 2009;27(1):79–86. doi: 10.1016/j.mri.2008.05.014. [DOI] [PubMed] [Google Scholar]
- 52.Morgenstern I., Raijmakers M. T. M., Peters W. H. M., Hoensch H., Kirch W. Homocysteine, cysteine, and glutathione in human colonic mucosa: elevated levels of homocysteine in patients with inflammatory bowel disease. Digestive Diseases and Sciences. 2003;48(10):2083–2090. doi: 10.1023/a:1026338812708. [DOI] [PubMed] [Google Scholar]
- 53.Wang W. W., Qiao S. Y., Li D. F. Amino acids and gut function. Amino Acids. 2009;37(1):105–110. doi: 10.1007/s00726-008-0152-4. [DOI] [PubMed] [Google Scholar]
- 54.Liu X., Beaumont M., Walker F., et al. Beneficial effects of an amino acid mixture on colonic mucosal healing in rats. Inflammatory Bowel Diseases. 2013;19(13):2895–2905. doi: 10.1097/01.MIB.0000435849.17263.c5. [DOI] [PubMed] [Google Scholar]
- 55.Cosmi L., Maggi L., Santarlasci V., Liotta F., Annunziato F. T helper cells plasticity in inflammation. Cytometry Part A. 2014;85(1):36–42. doi: 10.1002/cyto.a.22348. [DOI] [PubMed] [Google Scholar]
- 56.Poffenberger M. C., Jones R. G. Amino acids fuel T cell-mediated inflammation. Immunity. 2014;40(5):635–637. doi: 10.1016/j.immuni.2014.04.017. [DOI] [PubMed] [Google Scholar]
- 57.Pollizzi K. N., Powell J. D. Regulation of T cells by mTOR: the known knowns and the known unknowns. Trends in Immunology. 2015;36(1):13–20. doi: 10.1016/j.it.2014.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Chi H. Regulation and function of mTOR signalling in T cell fate decisions. Nature Reviews Immunology. 2012;12(5):325–338. doi: 10.1038/nri3198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Bar-Peled L., Sabatini D. M. Regulation of mTORC1 by amino acids. Trends in Cell Biology. 2014;24(7):400–406. doi: 10.1016/j.tcb.2014.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Pezze P. D., Ruf S., Sonntag A. G., et al. A systems study reveals concurrent activation of AMPK and mTOR by amino acids. Nature Communications. 2016;7, article 13254 doi: 10.1038/ncomms13254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Rittig N., Bach E., Thomsen H. H., et al. Amino acid supplementation is anabolic during the acute phase of endotoxin-induced inflammation: a human randomized crossover trial. Clinical Nutrition. 2016;35(2):322–330. doi: 10.1016/j.clnu.2015.03.021. [DOI] [PubMed] [Google Scholar]
- 62.Crozier S. J., Kimball S. R., Emmert S. W., Anthony J. C., Jefferson L. S. Oral leucine administration stimulates protein synthesis in rat skeletal muscle. Journal of Nutrition. 2005;135(3):376–382. doi: 10.1093/jn/135.3.376. [DOI] [PubMed] [Google Scholar]
- 63.Kato H., Miura K., Nakano S., Suzuki K., Bannai M., Inoue Y. Leucine-enriched essential amino acids attenuate inflammation in rat muscle and enhance muscle repair after eccentric contraction. Amino Acids. 2016;48(9):2145–2155. doi: 10.1007/s00726-016-2240-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ravindran R., Loebbermann J., Nakaya H. I., et al. The amino acid sensor GCN2 controls gut inflammation by inhibiting inflammasome activation. Nature. 2016;531(7595):523–527. doi: 10.1038/nature17186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Yao K., Fang J., Yin Y.-L., Feng Z.-M., Tang Z.-R., Wu G. Tryptophan metabolism in animals: important roles in nutrition and health. Frontiers in Bioscience (Scholar Edition) 2011;3(1):286–297. doi: 10.2741/s152. [DOI] [PubMed] [Google Scholar]
- 66.Hisamatsu T., Okamoto S., Hashimoto M., et al. Novel, objective, multivariate biomarkers composed of plasma amino acid profiles for the diagnosis and assessment of inflammatory bowel disease. PLoS ONE. 2012;7(1) doi: 10.1371/journal.pone.0031131.e31131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Mine Y., Zhang H. Calcium-sensing receptor (CaSR)-mediated anti-inflammatory effects of L-amino acids in intestinal epithelial cells. Journal of Agricultural and Food Chemistry. 2015;63(45):9987–9995. doi: 10.1021/acs.jafc.5b03749. [DOI] [PubMed] [Google Scholar]
- 68.Mozaffari S., Abdollahi M. Melatonin, a promising supplement in inflammatory bowel disease: a comprehensive review of evidences. Current Pharmaceutical Design. 2011;17(38):4372–4378. doi: 10.2174/138161211798999357. [DOI] [PubMed] [Google Scholar]
- 69.Söderquist F., Hellström P. M., Cunningham J. L. Human gastroenteropancreatic expression of melatonin and its receptors MT1 and MT2. PLoS ONE. 2015;10(3) doi: 10.1371/journal.pone.0120195.e0120195 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Sayyed H. G., Jaumdally R. J., Idriss N. K., El Sers D. A., Blann A. The effect of melatonin on plasma markers of inflammation and on expression of nuclear factor-kappa beta in acetic acid-induced colitis in the rat. Digestive Diseases and Sciences. 2013;58(11):3156–3164. doi: 10.1007/s10620-013-2811-x. [DOI] [PubMed] [Google Scholar]
- 71.Celinski K., Konturek P. C., Konturek S. J., et al. Effects of melatonin and tryptophan on healing of gastric and duodenal ulcers with Helicobacter pylori infection in humans. Journal of Physiology and Pharmacology. 2011;62(5):521–526. [PubMed] [Google Scholar]
- 72.Kim T. K., Park Y. S., Baik H., et al. Melatonin modulates adiponectin expression on murine colitis with sleep deprivation. World Journal of Gastroenterology. 2016;22(33):7559–7568. doi: 10.3748/wjg.v22.i33.7559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Souba W. W. Glutamine and cancer. Annals of Surgery. 1993;218(6):715–728. doi: 10.1097/00000658-199312000-00004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Kuhn K. S., Muscaritoli M., Wischmeyer P., Stehle P. Glutamine as indispensable nutrient in oncology: experimental and clinical evidence. European Journal of Nutrition. 2010;49(4):197–210. doi: 10.1007/s00394-009-0082-2. [DOI] [PubMed] [Google Scholar]
- 75.Nakaya M., Xiao Y., Zhou X., et al. Inflammatory T cell responses rely on amino acid transporter ASCT2 facilitation of glutamine uptake and mTORC1 kinase activation. Immunity. 2014;40(5):692–705. doi: 10.1016/j.immuni.2014.04.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Xue H., Sufit A. J. D., Wischmeyer P. E. Glutamine therapy improves outcome of in vitro and in vivo experimental colitis models. Journal of Parenteral and Enteral Nutrition. 2011;35(2):188–197. doi: 10.1177/0148607110381407. [DOI] [PubMed] [Google Scholar]
- 77.Akobeng A. K., Elawad M., Gordon M. Glutamine for induction of remission in Crohn's disease. The Cochrane Database of Systematic Reviews. 2016;2 doi: 10.1002/14651858.cd007348.pub2.CD007348 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Joo E., Yamane S., Hamasaki A., et al. Enteral supplement enriched with glutamine, fiber, and oligosaccharide attenuates experimental colitis in mice. Nutrition. 2013;29(3):549–555. doi: 10.1016/j.nut.2012.09.007. [DOI] [PubMed] [Google Scholar]
- 79.Davaatseren M., Hwang J.-T., Park J. H., Kim M.-S., Wang S., Sung M. J. Poly-γ-glutamic acid attenuates angiogenesis and inflammation in experimental colitis. Mediators of Inflammation. 2013;2013:8. doi: 10.1155/2013/982383.982383 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Bauchart-Thevret C., Stoll B., Burrin D. G. Intestinal metabolism of sulfur amino acids. Nutrition Research Reviews. 2009;22(2):175–187. doi: 10.1017/S0954422409990138. [DOI] [PubMed] [Google Scholar]
- 81.Burrin D. G. New insights into sulfur amino acid function in gut health and disease. Proceedings of the 2014 ADSA-ASAS-CSAS Joint Annual Meeting; 2014; Kansas City, Mo, USA. pp. 597–598. [Google Scholar]
- 82.Maroufyan E., Kasim A., Yong Meng G., et al. Effect of dietary combination of methionine and fish oil on cellular immunity and plasma fatty acids in infectious bursal disease challenged chickens. The Scientific World Journal. 2013;2013:9. doi: 10.1155/2013/531397.531397 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Guijarro L. G., Mate J., Gisbert J. P., et al. N-acetyl-L-cysteine combined with mesalamine in the treatment of ulcerative colitis: randomized, placebo-controlled pilot study. World Journal of Gastroenterology. 2008;14(18):2851–2857. doi: 10.3748/wjg.14.2851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhang H., Kovacs-Nolan J., Kodera T., Eto Y., Mine Y. γ-Glutamyl cysteine and γ-glutamyl valine inhibit TNF-α signaling in intestinal epithelial cells and reduce inflammation in a mouse model of colitis via allosteric activation of the calcium-sensing receptor. Biochimica et Biophysica Acta—Molecular Basis of Disease. 2015;1852(5):792–804. doi: 10.1016/j.bbadis.2014.12.023. [DOI] [PubMed] [Google Scholar]
- 85.Santangelo F. The regulation of sulphurated amino acid junctions: fact or fiction in the field of inflammation? Amino Acids. 2002;23(4):359–365. doi: 10.1007/s00726-002-0206-y. [DOI] [PubMed] [Google Scholar]
- 86.Shimizu M., Zhao Z., Ishimoto Y., Satsu H. Dietary taurine attenuates dextran sulfate sodium (DSS)-induced experimental colitis in mice. Advances in Experimental Medicine and Biology. 2009;643:265–271. doi: 10.1007/978-0-387-75681-3_27. [DOI] [PubMed] [Google Scholar]
- 87.Joo K., Lee Y., Choi D., et al. An anti-inflammatory mechanism of taurine conjugated 5-aminosalicylic acid against experimental colitis: Taurine chloramine potentiates inhibitory effect of 5-aminosalicylic acid on IL-1β-mediated NFκB activation. European Journal of Pharmacology. 2009;618(1–3):91–97. doi: 10.1016/j.ejphar.2009.07.009. [DOI] [PubMed] [Google Scholar]
- 88.Giriş M., Depboylu B., Dogru-Abbasoglu S., et al. Effect of taurine on oxidative stress and apoptosis-related protein expression in trinitrobenzene sulphonic acid-induced colitis. Clinical and Experimental Immunology. 2008;152(1):102–110. doi: 10.1111/j.1365-2249.2008.03599.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Motta J.-P., Flannigan K. L., Agbor T. A., et al. Hydrogen sulfide protects from colitis and restores intestinal microbiota biofilm and mucus production. Inflammatory Bowel Diseases. 2015;21(5):1006–1017. doi: 10.1097/MIB.0000000000000345. [DOI] [PubMed] [Google Scholar]
- 90.Medani M., Collins D., Docherty N. G., Baird A. W., O'Connell P. R., Winter D. C. Emerging role of hydrogen sulfide in colonic physiology and pathophysiology. Inflammatory Bowel Diseases. 2011;17(7):1620–1625. doi: 10.1002/ibd.21528. [DOI] [PubMed] [Google Scholar]
- 91.Jin Z., Chan H., Ning J., Lu K., Ma D. The role of hydrogen sulfide in pathologies of the vital organs and its clinical application. Journal of Physiology and Pharmacology. 2015;66(2):169–179. [PubMed] [Google Scholar]
- 92.Flannigan K. L., Agbor T. A., Motta J.-P., et al. Proresolution effects of hydrogen sulfide during colitis are mediated through hypoxia-inducible factor-1α. FASEB Journal. 2015;29(4):1591–1602. doi: 10.1096/fj.14-266015. [DOI] [PubMed] [Google Scholar]
- 93.Zandifar A., Seifabadi S., Zandifar E., et al. Comparison of the effect of topical versus systemic L-arginine on wound healing in acute incisional diabetic rat model. Journal of Research in Medical Sciences. 2015;20(3):233–238. [PMC free article] [PubMed] [Google Scholar]
- 94.de Lima Silva J. J., Pompeu D. G., Ximenes N. C., et al. Effects of kaurenoic acid and arginine on random skin flap oxidative stress, inflammation, and cytokines in rats. Aesthetic Plastic Surgery. 2015;39(6):971–977. doi: 10.1007/s00266-015-0559-8. [DOI] [PubMed] [Google Scholar]
- 95.Li P., Yin Y.-L., Li D., Kim W. S., Wu G. Amino acids and immune function. The British Journal of Nutrition. 2007;98(2):237–252. doi: 10.1017/s000711450769936x. [DOI] [PubMed] [Google Scholar]
- 96.Rees D. D., Palmer R. M. J., Hodson H. F., Moncada S. A specific inhibitor of nitric oxide formation from L-arginine attenuates endothelium-dependent relaxation. British Journal of Pharmacology. 1989;96(2):418–424. doi: 10.1111/j.1476-5381.1989.tb11833.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Taha M. O., De Oliveira J. V., Dias Borges M., et al. L-arginine modulates intestinal inflammation in rats submitted to mesenteric ischemia-reperfusion injury. Transplantation Proceedings. 2016;48(2):512–515. doi: 10.1016/j.transproceed.2015.12.063. [DOI] [PubMed] [Google Scholar]
- 98.Hong S.-K. S., Maltz B. E., Coburn L. A., et al. Increased serum levels of L-arginine in ulcerative colitis and correlation with disease severity. Inflammatory Bowel Diseases. 2010;16(1):105–111. doi: 10.1002/ibd.21035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Zwintscher N. P., Shah P. M., Salgar S. K., et al. Hepatocyte growth factor, hepatocyte growth factor activator and arginine in a rat fulminant colitis model. Annals of Medicine and Surgery. 2016;7:97–103. doi: 10.1016/j.amsu.2016.03.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Farghaly H. S., Thabit R. H. l-arginine and aminoguanidine reduce colonic damage of acetic acid-induced colitis in rats: potential modulation of nuclear factor-κB/p65. Clinical and Experimental Pharmacology and Physiology. 2014;41(10):769–779. doi: 10.1111/1440-1681.12287. [DOI] [PubMed] [Google Scholar]
- 101.Soufli I., Toumi R., Rafa H., Touil-Boukoffa C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World Journal of Gastrointestinal Pharmacology and Therapeutics. 2016;7(3):353–360. doi: 10.4292/wjgpt.v7.i3.353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Cheah I. K., Halliwell B. Ergothioneine; antioxidant potential, physiological function and role in disease. Biochimica et Biophysica Acta - Molecular Basis of Disease. 2012;1822(5):784–793. doi: 10.1016/j.bbadis.2011.09.017. [DOI] [PubMed] [Google Scholar]
- 103.Asahi T., Wu X., Shimoda H., et al. A mushroom-derived amino acid, ergothioneine, is a potential inhibitor of inflammation-related DNA halogenation. Bioscience, Biotechnology and Biochemistry. 2016;80(2):313–317. doi: 10.1080/09168451.2015.1083396. [DOI] [PubMed] [Google Scholar]