

Abstract
Background
Medicinal plants are becoming more popular in the treatment of various diseases because of the adverse effects of the current therapy, especially antioxidant plant components such as phenols and flavonoids have a protective role against oxidative stress-induced degenerative diseases like diabetes. Thus, the purpose of this study was to investigate β-cell protection and antidiabetic activities of Crassocephalum crepidioides (Asteraceae) Benth. S. Moore.
Method
The in-vitro study was conducted by the pancreatic β-cell culture and α-amylase inhibition technique which includes two methods, namely starch-iodine method and 3,5-dinitrosalicylic acid (DNSA) method. On the other hand, the in-vivo study was performed by oral glucose tolerance test (OGTT) method and alloxan-induced diabetes method by using Wistar albino rat. At the end pancreatic specimens were removed and processed for histopathological study.
Result
The plant extract showed significant (*p < 0.05, **p < 0.01) effect on hyperglycemia as compared to standard (Gliclazide) in OGTT. The plant extract showed efficient protection activity of pancreatic β-cell from cell death in INS-1 cell line by significantly reduced (*p < 0.05, **p < 0.01) the levels alloxan-induced apoptosis and intracellular reactive oxygen species (ROS) accumulation. In addition, the plant extract showed a significant (*p < 0.05, **p < 0.01) effect on hyperglycemia by increases in percent of β-cells present in each islet (45% – 60%) compared to the diabetic group.
Conclusion
The result showed that C. crepidioides had β-cell protection and antidiabetic activities in pancreatic β-cell culture and Wistar albino rat.
Electronic supplementary material
The online version of this article (doi:10.1186/s12906-017-1697-0) contains supplementary material, which is available to authorized users.
Keywords: Crassocephalum crepidioides, β-Cell Protection, Antidiabetes, Alloxan, Apoptosis, Reactive Oxygen Species, INS-1, Wistar Albino Rat
Background
Diabetes mellitus is a metabolic disorder characterized by high blood sugar (hyperglycemia) due to the disturbances of carbohydrate, fat and protein metabolism [1]. It can be classified as type-1 diabetes also known as insulin-dependent diabetes mellitus (IDDM) and type-2 diabetes also called non-insulin dependent diabetes mellitus (NIDDM) [2].
Pancreatic α-amylase inhibitors effectively reduced hyperglycemia through the control of starch breakdown [3]. Flavonoids or phenolic compounds containing plants can inhibit α-amylase enzyme lead to reduce blood glucose level [4]. Alloxan, is a potent diabetogenic agent that selectively damage pancreatic β-cells and it has been widely used as diabetes inducer [5]. It has been reported that alloxan rapidly and selectively accumulates in β-cells in comparison with non β-cells [6]. The alloxan-induced pancreatic β-cell damage through the generation of cytotoxic reactive oxygen species (ROS) [6–9]. The ROS produced by alloxan is responsible for the breakdown of DNA strand of pancreatic β–cells [10]. ROS induces oxidative stress that plays a pathological role in the development and progression of diabetic complications [11]. Thus, the identification of potent antidiabetic agents from natural sources, such as edible plants with minimum side effects, which will aid in the development of beneficial dietary guidelines for diabetic patients, is crucial. Recent studies have demonstrated that medicinal plants can protect β-cell by reducing alloxan-induced oxidative stress by scavenging ROS in-vitro and in-vivo, respectively [12–14].
The Crassocephalum crepidioides (C. crepidioides) Benth. S. Moore (Asteraceae) is an annual, edible herb also known as redflower ragleaf, that is widespread in many tropical and subtropical regions. The fleshy mucilaginous leaves and stems are commonly eaten and different parts of the plant have medical uses [15]. Moreover, it has been reported that C. crepidioides used to treat indigestion, stomach ache, epilepsy, sleeping sickness, swollen lips and it has also antitumor activity associated with nitric oxide production [16]. It is a potent antioxidant and protects against hepatotoxicity [17]. Our previous study found that, methanol extract of C. crepidioides possesses significant antioxidant activity as well as promising antihyperlipidemic activity [18]. Despite long traditional use of C. crepidioides as medicinal herb, no systemic phytochemical and pharmacological works have been carried out on this potential medicinal plant. Therefore, the aim of the present study to find out β-cell protection and antidiabetic effect C. crepidioides.
Methods
Plant material
The aerial parts of C. crepidioides was collected from Fatickchari, Chittagong in October 2013. The herbarium sheet was prepared by the expert following the standard procedure and taxonomic identification was made by Professor Shaikh Bokhtear Uddin, Department of Botany, University of Chittagong (CU), Bangladesh. A voucher specimen (Voucher number: SUS 5396) was deposited in Department of Botany, CU for preservation.
Preparation of extract
The collected sampls were sliced into smaller pieces and dried at room temperature under shade and then powdered finely and subjected to extraction. Five hundred grams of the powder was macerated with about 2500 ml of 80% methanol for 15 days. The extract was then filtered and the marc was remacerated twice using the same volume of solvent to exhaustively extract the plant material [19]. The methanol was then removed from the extract by evaporation under reduced pressure using a rota vapor (BUCHI Rotavapour R-200, Switzerland) at 40 °C. The resulting dry hydroalcoholic extract was weighed and calculated for percentage yield, which was 23% (w/w). The dried plant extract was reconstituted with distilled water for oral administration.
Identification of phenolic and flavonoids compound by chromatographic method
In order to recognize phenolic and flavonoids components of extract, column chromatography (CC) and thin layer chromatography (TLC) were used. The extract was subjected to CC with hexane: ethyl acetate in various solvent ratio. The fractions were collected depending on the visible changes in the colorful bands running out of the column. Then the fractions were subjected for TLC profiling. A variety of indicators, including iodine vapour, p-anisaldehyde-sulphoric acid, chloranil reagent, natural product reagent, aluminum chloride were used in this assay. The indicators were sprayed on TLC plate and observed at 254 nm and 365 nm wavelength under UV light. The Rf values resulted by test samples were 0.88 and 0.93 for extract which were parallel to the Rf values of reference compounds gallic acid and quercetin.
Determination of α-amylase inhibition by starch-iodine color assay
Screening of plant material for α–amylase inhibitors was carried out in a microtitreplate according to based on the starch-iodine test [20]. The total assay mixture composed of 200 μL 0.02 M sodium phosphate buffer (pH 6.9 containing 6 mM sodium chloride), 60 μL of porcine pancreatic α–amylase (PPA) solution and plant extracts at concentration from 0 to 200 mg/mL (w/v) was incubated at 37 °C for 10 min. Then soluble starch 200 μL (1%, w/v) was added to each reaction well and the mixtures were then incubated at 37 °C for 15 min. The enzymatic reaction was stopped by the addition of 0.1 M HCl (120 μL) followed by the addition of 600 μL of iodine reagent (5 mM I2 and 5 mM KI). The color change was noted and the absorbance was read at 620 nm on a microplate reader. The control reaction representing 100% enzyme activity did not contain any plant extract. To eliminate the absorbance produced by plant extract, appropriate extract controls without the enzyme were also included. The known PPA inhibitor, acarbose, was used as positive control at a concentration range of 0–25 mg/mL. A dark-blue color indicated the presence of starch; a yellow color indicated the absence of starch while a brownish color indicated partially degraded starch in the reaction mixture. In the presence of inhibitors of the extracts, the starch added to the enzyme assay mixture was not degraded and gave a dark-blue color complex, whereas no color complex was developed in the absence of the inhibitor, indicating that starch was completely hydrolyzed by α–amylase.
Determination of α-amylase inhibition by 3, 5-dinitrosalicylic acid (DNSA) assay
The inhibition assay was performed using the chromogenic 3, 5-dinitrosalicylic acid (DNSA) method [21]. The total assay mixture composed of 500 μL of 0.02 M sodium phosphate buffer (pH 6.9 containing 6 mM sodium chloride), 20 μL of PPA solution and extracts at concentration from 0 to 200 mg/mL (w/v) were incubated at 37 °C for 10 min. After pre-incubation, 500 μL of 1% (v/v) starch solution in the above buffer were added to each tube and incubated at 37 °C for 15 min. The reaction was terminated with 1.0 mL DNSA reagent, placed in boiling water bath for 5 min, cooled to room temperature, diluted and the absorbance measured at 540 nm. The known PPA inhibitor, acarbose was used a positive control at a concentration range of 0–25 mg/mL.
The IC50 values were determined from plots of percent inhibition versus inhibitor concentration and calculated by linear regression analysis from the mean inhibitory values. The IC50 values were defined as the concentration of the extract containing the α-amylase inhibitor that inhibited 50% of the PPA activity.
The other quantifiers were calculated as follows:
Cell culture
INS-1, a rat insulinoma cell line, was obtained from the American type culture collection (ATCC), USA. The cells were grown in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS), 10 mmol/L HEPES, 2 mmol/L L-glutamine, 50 μmol/L β-mercaptoethanol, 1 mmol/L sodium pyruvate, 100 U/mL penicillin, and 100 μg/mL streptomycin at 37 °C in a humidified atmosphere containing 95% air and 5% CO2.
Determination of cell viability
Cell viability was determined by crystal violet assay. Briefly, INS-1 cells were seeded onto 24-well plate (5 × 104 cells/well), incubated overnight and pretreated with various concentrations of extracts (0–1000 μg/mL) for 24 h. Then, the medium was removed and cells were washed with phosphate buffer solution (PBS). 200 μL of 0.2% crystal violet solution was added to each well and incubated for 10 min at room temperature. Then wash with water and add 100 μL 1% SDS to solubilize the stain solution until color is uniform and no areas of dense coloration in bottom of wells. The samples were read at 590 nm in microplate reader (Spectra MAX, Gemini EM, Molecular Device).
Analysis of the protective effect of the extract on oxidative stress induced cytotoxicity
Alloxan-induced cell survival was determined by crystal violet assay. Briefly, INS-1 cells were seeded (5 × 104 cells/well) and cultured in 24-well culture plates. The cells were then preincubated with or without different concentrations of extract (25, 50, 100, and 200 μg/mL) at 37 °C in a humidified atmosphere of 5% CO2/95% air for 2 h followed by incubation with 200 μM alloxan for 24 h. After that the medium was removed and cells were washed with PBS and 200 μl of 0.2% crystal violet solution was added to each well and incubated for 10 min at room temperature. Then wash with water and add 100 μL 1% SDS to solubilize the stain solution until the color is uniform and no areas of dense coloration in bottom of wells. The samples were read at 590 nm in a microplate reader (Spectra MAX, Gemini EM, Molecular Device).
Measurement of intracellular reactive oxygen species (ROS) level
ROS level was measured by using DCFH-DA method. DCFH-DA is a non-fluorescent compound, and it can be enzymatically converted to a highly fluorescent compound, DCF, in the presence of ROS [22]. In brief, after the treatment, INS-1 cells were washed with PBS and incubated with DCFH-DA at a final concentration of 10 μmol/L for 30 min at 37 °C in darkness. The fluorescence intensity was measured in the microplate reader (Spectra MAX, Gemini EM, Molecular Device) at an excitation wave length of 485 nm and an emission wave length of 538 nm after the cells were washed three times with PBS to remove the extracellular DCFH-DA. The level of intracellular ROS was showed as a percentage of non-treated control.
Apoptosis assay
Hoechst33342 staining distinguishing apoptotic from normal cells based on nuclear chromatin condensation and fragmentation was used for the qualitative and quantitative analyses of the apoptotic cells. INS-1 cells were cultured in 6-well plates for 24 h. After treatment, the cells were incubated with 5 μg/ml Hoechst 33,342 for 15 min, washed twice with PBS, and then visualized by inverted fluorescence microscopy (Axioskop 2 plus microscope, Carl Zeiss, Oberkochen, Germany). The apoptotic nuclei were counted in at least 200 cells from five non overlapping fields in all treatment, and expressed as a percentage of the total number of nuclei counted.
Experimental animal
Wistar albino rats of either sex, aged 8–10 weeks, weighing 120–250 g were used for the experiment. The animals were originally obtained from International Centre for Diarrheal Disease Research, Bangladesh (ICDDR, B). They were kept in clean and dry polypropylene cages with 12 h light, dark cycle at 25 ± 2 °C and 45–55% relative humidity in the animal house. The rats were fed with a standard laboratory diet and water ad libitum. Food was withdrawn 12 h prior to and during the experiment. As these animals are very sensitive to environmental changes, they are kept before the test for at least 3–4 days in the environment where the experiment will take place. The protocol used in this study was carried out with the guidelines of the Institutional Animal Ethics Committee (IAEC) and approval was taken from ethical committee of Faculty of Veterinary Medicine, Chittagong Veterinary and Animal Sciences University.
Acute toxicity studies
Extract at the dose range of 100 mg to 2500 mg/kg were administered orally to different groups of rats, each group comprised of five rats. Mortality was observed after 72 h. Acute toxicity was determined according to the method of Litchfield and Wilcoxon [23].
Oral glucose tolerance test (OGTT)
OGTT is one of the most acceptable methods to evaluate the hypoglycemic activity [24]. Twenty fasted rats were divided into four groups with each group containing five rats. Group 1 was kept as control which received vehicle orally (p.o.), group 2 received gliclazide 10 mg/kg as standard, and group 3 and 4 received extracts 300 mg/kg and 500 mg/kg, respectively. After 30 min of extracts and gliclazide administration all groups were orally treated with 3 g/kg of glucose. The blood glucose level was measured by glucometer using glucose estimation kit at 0, 30, 90 and 120 mins of glucose loading. Blood samples were collected from the tail vein.
Induction of diabetes in experimental animals
The rats were fasted for 12 h prior to the induction of diabetes. Alloxan was freshly prepared in 0.2 ml 0.9% normal saline was administered intraperitoneally (i.p.) at a single dose of 150 mg/kg. Development of diabetes was confirmed by measuring the blood glucose concentration at 48 h after the administration of alloxan. Rats with blood glucose levels of <7 mmol/dL were excluded from the experiment. Treatment with plant extracts was started 48 h after alloxan injection [25].
Experimental design
Animals were divided into five groups with each group receiving solutions as follows:
Normal control (Saline 10 mL/kg, p.o.)
Alloxan treated control (150 mg/kg, i.p.)
Alloxan (150 mg/kg, i.p.) + plant extract (150 mg/kg, p.o.)
Alloxan (150 mg/kg, i.p.) + plant extract (300 mg/kg, p.o.)
Saline and plant extract (150 and 300 mg/kg) were administered to diabetic rats for 14 days and the blood glucose level was measured at 0, 3, 5, 7, 9 and 14th days. After 14 days of plant extract treatment, the animals were sacrificed and blood was collected, sera separated by centrifugation at 3 000 g for 10 min. Details of the estimation of insulin, urea, creatinine and glycosylated haemoglobin are given in Additional file 1.
Haemetoxylin and eosin staining
After plant extract treatment, pancreas from the normal control, alloxan control, and plant extracts (150 and 300 mg/kg)-treated rats were quickly removed for histological studies. Removed pancreatic tissues were washed in saline, fixed in 10% formalin and fixed10% neutral buffered formalin (NBF) for 48 h and processed with paraffin embedding. The sections stained in 0.1% Mayer’s haemetoxylin, counterstained in 0.5% eosin, mounted and were observed under microscope.
Statistical data analysis
All the data were expressed as mean ± SD and one way ANOVA (Analysis of variance) followed by Dunnett’s test was used for the statistical analysis using SPSS software (version 16). *p < 0.05 and **p < 0.01 were considered significant.
Results
Phenols and flavonoids
Phytochemical assay by chromatographic method showed that the main components, including phenolic and flavonoid compounds were present in the extract (Table 1).
Table 1.
Phytochemical result of extract
| Compounds | Reagents | Standards | Results |
|---|---|---|---|
| Phenols | P-Anisaldehyde-Sulphoric acid | Gallic acid | + |
| Chloranil reagent | Gallic acid | + | |
| Flavonoids | Natural product reagent | Quercetin | + |
| Aluminum chloride | Quercetin | + |
α-Amylase inhibition activity
In the present study of the in-vitro α-amylase inhibition activity test, the extract showed significant % inhibition of α-amylase relative to the standard treatment, acarbose in starch-iodine test and DNSA assay, respectively (Figs. 1 and 2). In starch-iodine test, acarbose showed highest α-amylase inhibition of 96.24% at a concentration of 25 mg/mL (Fig. 1a) with an IC50 value 3.11 ± 0.10 mg/mL (Fig. 1c), while the extract showed a highest α-amylase inhibition of 66.37% at a concentration of 200 mg/mL (Fig. 1b) with an 126.85 ± 2.10 mg/mL (Fig. 1c). In DNSA assay, acarbose showed highest inhibition of 85.63% at a concentration of 25 mg/mL (Fig. 2a) with an IC50 value 7.52 ± 0.30 mg/mL (Fig. 2c), while the extract produced a highest inhibition of 57.76% at a concentration of 200 mg/mL (Fig. 2b) with an IC50 value 144.29 ± 5.10 mg/mL (Fig. 2c).
Fig. 1.
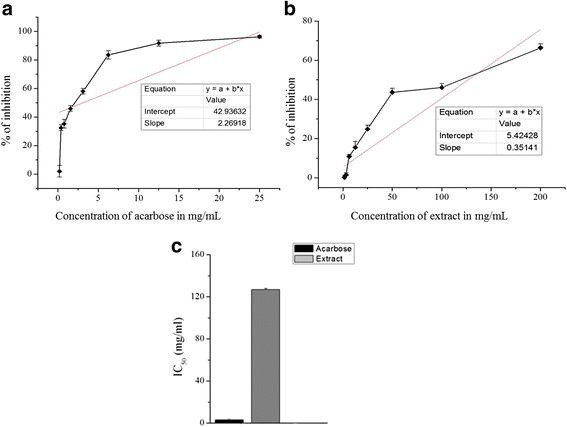
α-amylase inhibition activity of acarbose and C. crepidioides in starch-iodine assay. a Percentage inhibition of acarbose (b) 80% methanol extract of C. crepidioides and (c) IC50 of treatment groups on α -amylase enzyme in vitro. Values were represented as mean ± SD (n = 3)
Fig. 2.
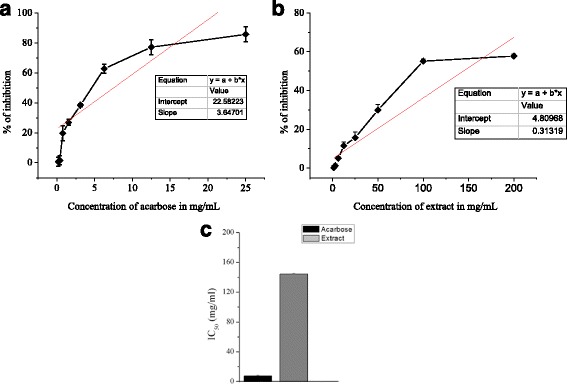
α-amylase inhibition activity of acarbose and C. crepidioides in DNSA assay. a Percentage inhibition of acarbose (b) 80% methanol extract of C. crepidioides and (c) IC50 of treatment groups on α -amylase enzyme in vitro. Values were represented as mean ± SD (n = 3)
Effect of extract on INS-1 cell lines
The cytotoxic effect of the extract on rat pancreatic cell line INS-1 was evaluated by incubating it with various concentrations of extract (1–1000 μg/mL). The toxicity results revealed a decrease in percentage of viability at higher concentrations of the extract and the IC50 value was found to be 464.38 ± 3.1 μg/mL (Fig. 3).
Fig. 3.
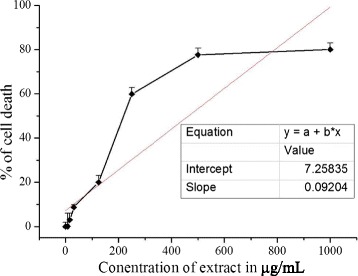
Concentration dependent effect of C. crepidioides extract on INS-1 cell line. Results are expressed as % of cell reduction (= % of cell death). Values were represented as mean ± SD (n = 4). Toxicity results for INS-1 cell line incubated with different concentrations of extract
Protective effect of extract on alloxan induced cytotoxicity
The effect of extract on the viability of INS-1 cells cultured under alloxan-induced toxicity conditions was measured by crystal violet assay. Pretreatment of INS-1 cells for 2 h with extract at concentrations of 25–200 μg/mL significantly protected INS-1 cells from alloxan toxicity. An increase in cell viability was observed in treated cells compared to alloxan control in a dose dependent manner (Fig. 4). The result displayed that extracts (50 μg/mL, 100 μg/mL and 200 μg/mL) possessed the best protective effects (p < 0.01).
Fig. 4.
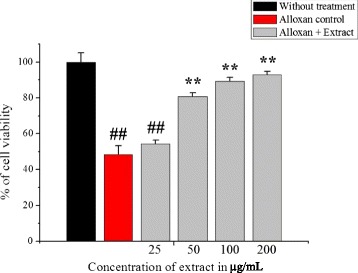
Protective effect of C. crepidioides extract on alloxan induced cell cytotoxicity in INS-1 cells. Values were represented as mean ± SD (n = 4). ## p < 0.01 as compared with the control group; **p < 0.01 as compared with the alloxan control group. Toxicity results of INS-1 cell line induced with 200 μM of alloxan and preincubated with different concentrations of extract
Effect of extract on intracellular ROS activity
As shown in Fig. 5, after exposed to 200 μmol/L alloxan for 24 h, the intracellular ROS level of the INS-1 cells markedly increased to 223.36% relative to the control value (100%, p < 0.01), which suggests that alloxan induce oxidative stress [26, 27]. When the cells were incubated with different concentrations of extract (25, 50, 100 and 200 μg/mL) in the presence of 200 μmol/L alloxan for 24 h, the intracellular ROS levels significantly decreased to 218.22%, 189.73%, 184.07% and 130.8% of the control value (100%, p < 0.01), respectively.
Fig. 5.
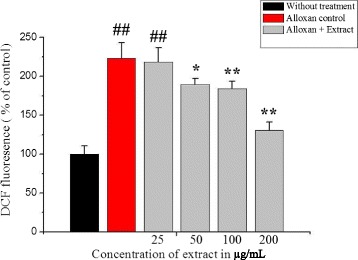
Protective Effect of C. crepidioides extract on alloxan-induced ROS in INS-1 cells. DCF fluorescence reflects ROS level. Values were represented as mean ± SD from three independent experiments. ## p < 0.01 as compared with the control group; *p < 0.05 and **p < 0.01 as compared with the alloxan control group. Intracellular ROS activity results for INS-1 cell line induced with 200 μM of alloxan and pre incubated with different concentrations of extract
Effect of the extract on alloxan-induced apoptosis in INS-1 cells
INS-1 cells treated with 200 μmol/L for 24 h showed typical characteristics of apoptosis, including the condensation of chromatin, the shrinkage of nuclei, alteration of size, shape and number of nuclei using Hoechst 33,342 staining as shown in (Fig. 6a). The amount of apoptotic nuclei was markedly increased, and the apoptosis rate increased to 22.10% relative to the control group (100%, p < 0.01). However, pretreatment with extract (200 μg/ml) in the presence of alloxan (200 μmol/L), the number of apoptotic cells was obviously decreased, and the apoptosis rate reduced to 14.81% relative to the control group (100%, p < 0.01) (Fig. 6b).
Fig. 6.
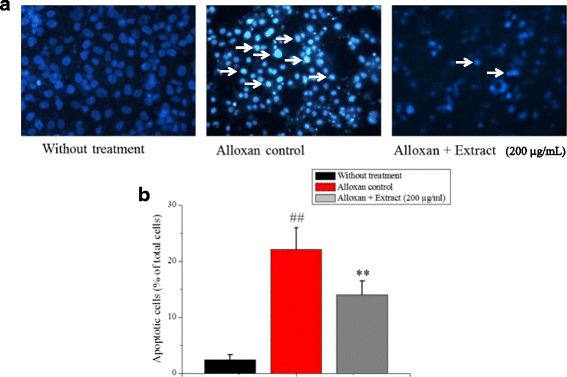
Protective effects of C. crepidioides extract on the cell survival in alloxan-treated INS-1 cells by Hoechst 33,342 staining method. a Representative images by Hoechst 33,342 staining, arrowheads in the pictures indicate the nuclei of the apoptotic cells (Hoechst-positive cells). b The apoptosis rate was determined by calculating the percentage of Hoechst positive cells over the total number of cells. Values were represented as mean ± SD from three independent experiments. ## p < 0.01 as compared with the control group; **p < 0.01 as compared with the alloxan control group. Apoptosis activity results for INS-1 cell line induced with 200 μM of alloxan and pre incubated with different concentrations of extract
Acute toxicity
The plant extract did not show any sign of acute toxicity up to a dose of 2500 mg/kg of body weight. The behavior of the animals was clearly observed for the first 8 h, then at an interval of every 8 h during next 72 h. The extract did not produce a significant change in animal behavior or mortality.
Effect on OGTT
It was observed that the plant extracts showed a significant effect on hyperglycemia as compared to standard treatment, gliclazide (Fig. 7). In the glucose control rats, the blood glucose level was increased after 30 min and remained high over the next 120 min. Plant extracts (300 mg/kg and 500 mg/kg) significantly reduced (p < 0.05) the blood glucose level at 30 min and remain low over the next 120 min when compared with the glucose control.
Fig. 7.
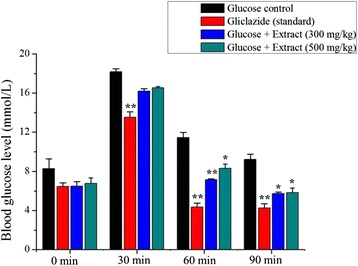
Effect of C. crepidioides extract on OGTT. Values were represented as mean ± SD (n = 6). *p < 0.05 and **p < 0.01 as compared with the glucose control group
Effect on alloxan-induced diabetes
It was observed that both doses (150 mg/kg and 300 mg/kg) of the extract showed successful hypoglycemic activity (Fig. 8). In the diabetic control rats, the blood glucose level was increased and remained high over the 14th days. Plant extracts significantly reduced (p < 0.05) the blood glucose level when compared with the diabetic control.
Fig. 8.
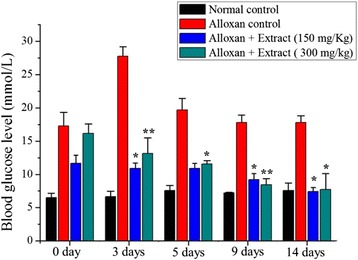
Effect of C. crepidioides extract on alloxan-induced diabetic rats. Values were represented as mean ± SD (n = 6). *p < 0.05 and **p < 0.01 as compared with the alloxan control group
Morphological studies
In histopathological examination, rat pancreatic tissue of the normal group showed regular exocrine tissue with interspersed islets of langerhans. No cellular abnormality or inflammatory infiltration was seen (Fig. 9a). The rat pancreatic tissue of the diabetic control group revealed regular exocrine pancreas with a reduced number of islets of langerhans. As a whole, each islet contained a reduced number of cells. Furthermore, few pyknotic cells along with mild infiltration of lymphocytes were observed within the islets (Fig. 9b). Rat pancreatic tissues of the plant extract 150 mg/kg and 300 mg/kg groups showed regular exocrine pancreas with moderately increased number and size of islets of langerhans as well as increased number of cells per islets with a distinct margin between exocrine and endocrine tissue (Fig. 9c, d). In the plant extract-treated group, morphometric results showed a significant increase in the number of the islets/mm2 (Table 2) and the number of β-cells in islet compared to the diabetic group (Table 3). There was no significant difference in the number of islets/mm2compared to the normal group. There was a significant decrease in the percent of β-cells (25% – 30%) in the diabetic group, while there was a significant increase in the plant extract-treated group (45% – 60%) compared to the diabetic groups (Fig. 9e).
Fig. 9.
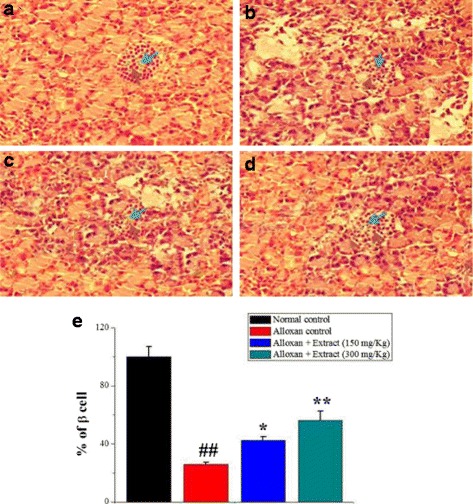
Histopathological examination under microscopy: (a) Normal cellular population in the islets of langerhans in pancreas of vehicle-treated rats. b Extensive damage to the islets of langerhans and reduced dimensions of islets in diabetic control rats. (c, d) The partial restoration of normal cellular population and enlarged size of β-cells were shown by the extracts (150 mg/kg and 300 mg/kg), respectively. e The β-cell regeneration rate was determined by calculating the percentage of H & E positive cells over the total number of cells. Values were represented as mean ± SD. ## p < 0.01 as compared with the control group; *p < 0.05 and **p < 0.01 as compared with the alloxan control group
Table 2.
The number of islets/mm2 in the control and treatment groups
| Groups | Number of islets/mm2 |
|---|---|
| Normal control | 5.2 ± 0.62 |
| Alloxan control | 1.4## ± 0.93 |
| Alloxan + Extract (150 mg/kg) | 2.9* ± 0.44 |
| Alloxan + Extract (300 mg/kg) | 3.9** ± 0.31 |
Values were represented as mean ± SD. ## p < 0.01 as compared with the control group; *p < 0.05 and **p < 0.01 as compared with the alloxan control group
Table 3.
The number of β cells per islet in the control and treatment groups
| Groups | Number of β cells/ islet |
|---|---|
| Normal control | 94.60 ± 6.02 |
| Alloxan control | 28.92## ± 2.13 |
| Alloxan + Extract (150 mg/kg) | 42.2* ± 3.31 |
| Alloxan + Extract (300 mg/kg) | 56.4** ± 4.01 |
Values were represented as mean ± SD. ## p < 0.01 as compared with the control group;
*p < 0.05 and **p < 0.01 as compared with the alloxan control group
Discussion
The blood glucose level has been increased by the α-amylase and α-glucosidase enzyme which break carbohydrate to simple absorbable monosaccharides [28]. The α-amylase and α–glucosidase inhibitors have been useful as oral hypoglycemic drugs for the control of hyperglycemia especially in patients with type II diabetes mellitus [29]. Inhibition of α-amylase delays carbohydrate digestion and prolong overall digestion time lead to reduce rate of glucose absorption and ultimately reducing the blood glucose level [30]. In our study, the plant extract showed significant % inhibition of α-amylase as compared to the standard anti-diabetic drug, acarbose, in both starch-iodine test and DNSA assay that could be attributed to the presence of phenolic and flavonoids components (Table 1) because polyphenols are capable of inhibiting carbohydrate hydrolyzing enzymes due to their ability to bind with proteins [31, 32].
Our results in this study showed pretreatment of INS-1 cells cultured under alloxan-induced cytotoxicity, with extract significantly increased survival and decreased rate of cell death. Treatment of INS-1 cells with extract also reduced ROS intracellular formation and decreased cell apoptosis that indicate the pancreatic β-cell protective effect of extract on the INS-1 cell line [33–35].
Furthermore, the study revealed that the plant extract showed a significant effect on hyperglycemia as compared to standard treatment, gliclazide, in OGTT. OGTT was used to identify the altered carbohydrate metabolism during post glucose administration. The ability of extract to lower the blood glucose level in oral glucose tolerance test suggests that rats treated with plant extract had better glucose utilization capacity [36]. It is also well established that sulfonylureas (e.g. gliclazide) cause hypoglycemia by stimulating insulin release from pancreatic β-cells [37]. Our result showed that the extract groups of diabetic rats significantly increased (p < 0.05 or p < 0.01) the serum insulin (Additional file 1: Table S1). The comparable effect of the extract with gliclazide suggests the possibility of a similar mode of action.
Additionally, our results demonstrated that the plant extracts showed significant effect on hyperglycemia as compared to alloxan control rats. The blood glucose lowering effect of plant extract could be attributed to the presence of flavonoids and phenolic compounds (Table 1) that have been associated with hypoglycemic activity [38, 39].
Histological findings of the pancreatic tissue using haematoxylin of the control group showed bluish-stained β-cells filling the interior of the islet, while pink-stained α-cells were arranged in mainly clusters located at the periphery of the islets and also between β-cells. Alloxan administration exhibited degenerative changes at the center of islets of langerhans and also decreased number of the bluish-stained β-cells, while the pink-stained α-cells remained present at the periphery of the islet. The alloxan caused significant morphological changes in diabetic rats with severe pancreatic β-cells destruction, such as decreasing the islets cell numbers, cell damage, and cell death as compared to the control group [40, 41]. The cytotoxic action of alloxan is mediated by ROS, with a simultaneous massive increase in cytosolic calcium concentration leading to rapid destruction of β-cells [42, 43]. The plant extract showed a significant increase in the number of the islets/mm2 and the number of β-cells in islet compared to diabetic group. The plant extract may act on islet functions (Table 2) lead to increasing the number of pancreatic islets (Table 3) pancreatic β-cells from oxidative damage through regulating ROS production (Fig. 5) and apoptotic rate (Fig. 6) [44, 45]. In summary, the results of this study showed the pancreatic β-cell protective properties of the extract through reducing the ROS production and decreasing the cell apoptosis (Fig. 10) that might be due to presence of flavonoids, polyphenolic compounds (Table 1) [46]. Therefore, this medicinal plant as a Complementary Medicine strategy may be used in metabolic diabetic diseases; however, further study is needed.
Fig. 10.
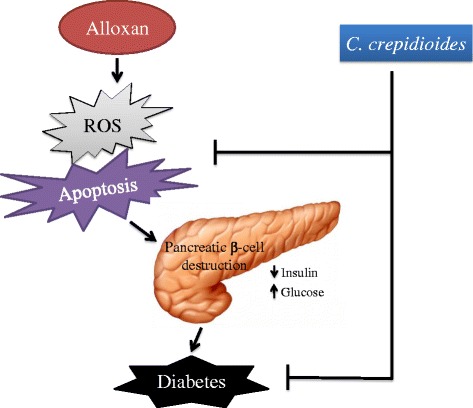
Proposed mechanism of β-cell protection and antidiabetic activities of C. crepidioides against alloxan-induced oxidative stress. The schematic diagram showing the proposed mechanism underlying the β-cell protection and antidiabetic effect of C. crepidioides against the alloxan-induced ROS production, apoptotic degeneration and pancreatic destruction
Conclusions
This study demonstrates that the extract possesses a significant anti-diabetic activity at different dose levels in both in-vitro and animal models. Hence, the study of in-vitro and in-vivo pharmacological and phytochemical evaluation of this plant forms a primary platform for future studies. The anti-apoptotic signaling mechanism of active compounds from this promissory extract should be further investigated as a natural remedy for diabetes treatment.
Acknowledgments
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (2013R1A2A2A01068683).
Funding
National Research Foundation of Korea (NRF) grant funded by the Korea government (2013R1A2A2A01068683).
Availability of data and materials
The data sets supporting the conclusions of this article are presented in this main paper. Plant materials used in this study have been identified at the University of Chittagong (CU) where voucher specimens are deposited.
Authors’ contribution
This study was designed, directed and coordinated by EB; HJ C; HY; MMMH; JA; OR and KM EB; HJC and HY act as the principal investigators, provided conceptual and technical guidance for all aspects of the project. E B planned and performed in-vivo experiments and analyzed data with LGH; LHY; HOR; CMY; KRB and KMA EB performed and analyzed in-vitro experiments with; MMMH; JA; OR and KM The manuscript was written by EB and HY and commented on by all authors. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Ethics approval
Approval was taken from ethical committee of Faculty of Veterinary Medicine, Chittagong Veterinary and Animal Sciences University for using animals by describing the protocols of the study and was performed according to the Guidelines for Animal experimentation established by the Institutional Animal Ethics Committee (IAEC).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- DNSA
5-dinitrosalicylic acid
- h
Hour
- IC50
50% inhibitory concentration
- kg
Kilogram
- mg
Milligram
- min
Minute
- mL
Milliliter
- OGTT
oRal glucose tolerance test
- PBS
Phosphate buffer solution
- PPA
Porcine pancreatic α-amylase
- ROS
Reactive oxygen species
- SD
Standard deviation
- SDS
Sodium dodecyl sulfate
- TLC
Layer chromatography
- UV
Ultraviolet
- μg
Microgram
Additional file
Effect of C. crepidioides on the serum insulin, urea, creatinine and HbA1c. (DOC 41 kb)
Contributor Information
Entaz Bahar, Email: entaz_bahar@yahoo.com.
Kazi-Marjahan Akter, Email: marjahan7silva@yahoo.com.
Geum-Hwa Lee, Email: heloin@jbnu.ac.kr.
Hwa-Young Lee, Email: youngat84@gmail.com.
Harun-Or Rashid, Email: rashid@jbnu.ac.kr.
Min-Kyung Choi, Email: mkelf78@nate.com.
Kashi Raj Bhattarai, Email: meekasik@jbnu.ac.kr.
Mir Mohammad Monir Hossain, Email: monir_pharm@yahoo.com.
Joushan Ara, Email: joushan.sb@gmail.com.
Kishor Mazumder, Email: k.mazumder@pharmacy.ustc.ac.bd.
Obayed Raihan, Email: mo.raihan@just.edu.bd.
Han-Jung Chae, Email: hjchae@jbnu.ac.kr.
Hyonok Yoon, Phone: +82-55-772-2422, Email: hoyoon@gnu.ac.kr.
References
- 1.Oh WK, Lee CH, Lee MS, Bae EY, Sohn CB, Oh H, Kim BY, Ahn JS. Antidiabetic effects of extracts from Psidium guajava. J Ethnopharmacol. 2005;96(3):411–415. doi: 10.1016/j.jep.2004.09.041. [DOI] [PubMed] [Google Scholar]
- 2.Spahr L, Butterworth RF, Fontaine S, Bui L, Therrien G, Milette PC, Lebrun LH, Zayed J, Leblanc A, Pomier-Layrargues G. Increased blood manganese in cirrhotic patients: relationship to pallidal magnetic resonance signal hyperintensity and neurological symptoms. Hepatology. 1996;24(5):1116–1120. doi: 10.1002/hep.510240523. [DOI] [PubMed] [Google Scholar]
- 3.Banting FG, Best CH, Collip JB, Campbell WR, Fletcher AA. Pancreatic Extracts in the Treatment of Diabetes Mellitus. Can Med Assoc J. 1922;12(3):141–146. [PMC free article] [PubMed] [Google Scholar]
- 4.P S, Zinjarde SS, Bhargava SY, Kumar AR. Potent alpha-amylase inhibitory activity of Indian Ayurvedic medicinal plants. BMC Complement Altern Med. 2011;11:5. doi: 10.1186/1472-6882-11-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Rho HW, Lee JN, Kim HR, Park BH, Park JW. Protective mechanism of glucose against alloxan-induced beta-cell damage: pivotal role of ATP. Exp Mol Med. 2000;32(1):12–17. doi: 10.1038/emm.2000.3. [DOI] [PubMed] [Google Scholar]
- 6.Gorus FK, Malaisse WJ, Pipeleers DG. Selective uptake of alloxan by pancreatic B-cells. Biochem J. 1982;208(2):513–515. doi: 10.1042/bj2080513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yamamoto H, Uchigata Y, Okamoto H. Streptozotocin and alloxan induce DNA strand breaks and poly(ADP-ribose) synthetase in pancreatic islets. Nature. 1981;294(5838):284–286. doi: 10.1038/294284a0. [DOI] [PubMed] [Google Scholar]
- 8.Takasu N, Asawa T, Komiya I, Nagasawa Y, Yamada T. Alloxan-induced DNA strand breaks in pancreatic islets. Evidence for H2O2 as an intermediate. J Biol Chem. 1991;266(4):2112–2114. [PubMed] [Google Scholar]
- 9.Malaisse WJ. Alloxan toxicity to the pancreatic B-cell. A new hypothesis. Biochem Pharmacol. 1982;31(22):3527–3534. doi: 10.1016/0006-2952(82)90571-8. [DOI] [PubMed] [Google Scholar]
- 10.Lee JH, Lee JS, Kim YR, Jung WC, Lee KE, Lee SY, Hong EK. Hispidin isolated from Phellinus linteus protects against hydrogen peroxide-induced oxidative stress in pancreatic MIN6N beta-cells. J Med Food. 2011;14(11):1431–1438. doi: 10.1089/jmf.2010.1493. [DOI] [PubMed] [Google Scholar]
- 11.Kaneto H, Nakatani Y, Kawamori D, Miyatsuka T, Matsuoka TA, Matsuhisa M, Yamasaki Y. Role of oxidative stress, endoplasmic reticulum stress, and c-Jun N-terminal kinase in pancreatic beta-cell dysfunction and insulin resistance. Int J Biochem Cell Biol. 2005;37(8):1595–1608. doi: 10.1016/j.biocel.2005.04.003. [DOI] [PubMed] [Google Scholar]
- 12.Diplock AT, Charleux JL, Crozier-Willi G, Kok FJ, Rice-Evans C, Roberfroid M, Stahl W, Vina-Ribes J. Functional food science and defence against reactive oxidative species. Br J Nutr. 1998;80(Suppl 1):S77–112. doi: 10.1079/BJN19980106. [DOI] [PubMed] [Google Scholar]
- 13.Reliene R, Schiestl RH. Antioxidant N-acetyl cysteine reduces incidence and multiplicity of lymphoma in Atm deficient mice. DNA Repair (Amst) 2006;5(7):852–859. doi: 10.1016/j.dnarep.2006.05.003. [DOI] [PubMed] [Google Scholar]
- 14.Sagrista ML, Garcia AE, Africa De Madariaga M, Mora M. Antioxidant and pro-oxidant effect of the thiolic compounds N-acetyl-L-cysteine and glutathione against free radical-induced lipid peroxidation. Free Radic Res. 2002;36(3):329–340. doi: 10.1080/10715760290019354. [DOI] [PubMed] [Google Scholar]
- 15.Grubben GJH. Plant Resources of Tropical Africa: Vegetables. PROTA Foundation Leiden: Backhuys Publishers; 2004. pp. 67–72. [Google Scholar]
- 16.Tomimori K, Nakama S, Kimura R, Tamaki K, Ishikawa C, Mori N. Antitumor activity and macrophage nitric oxide producing action of medicinal herb, Crassocephalum crepidioides. BMC Complement Altern Med. 2012;12:78. doi: 10.1186/1472-6882-12-78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Aniya Y, Koyama T, Miyagi C, Miyahira M, Inomata C, Kinoshita S, Ichiba T. Free radical scavenging and hepatoprotective actions of the medicinal herb, Crassocephalum crepidioides from the Okinawa Islands. Biol Pharm Bull. 2005;28(1):19–23. doi: 10.1248/bpb.28.19. [DOI] [PubMed] [Google Scholar]
- 18.Bahar E, Siddika MS, Nath B, Yoon H. Evaluation of In vitro Antioxidant and In vivo Antihyperlipidemic Activities of Methanol Extract of Aerial Part of Crassocephalum crepidioides (Asteraceae) Benth S Moore. Trop J Pharm Res. 2016;15(3):481–488. doi: 10.4314/tjpr.v15i3.8. [DOI] [Google Scholar]
- 19.Hailu W, Engidawork E. Evaluation of the diuretic activity of the aqueous and 80% methanol extracts of Ajuga remota Benth (Lamiaceae) leaves in mice. BMC Complement Altern Med. 2014;14:135. doi: 10.1186/1472-6882-14-135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Xiao Z, Storms R, Tsang A. A quantitative starch-iodine method for measuring alpha-amylase and glucoamylase activities. Anal Biochem. 2006;351(1):146–148. doi: 10.1016/j.ab.2006.01.036. [DOI] [PubMed] [Google Scholar]
- 21.Lorenz MG. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal Chem. 1959;31(3):426–428. doi: 10.1021/ac60147a030. [DOI] [Google Scholar]
- 22.Yokozawa T, Kim YA, Kim HY, Okamoto T, Sei Y. Protective effect of the Chinese prescription Kangen-karyu against high glucose-induced oxidative stress in LLC-PK1 cells. J Ethnopharmacol. 2007;109(1):113–120. doi: 10.1016/j.jep.2006.07.020. [DOI] [PubMed] [Google Scholar]
- 23.Litchfield JT, Jr, Wilcoxon F. A simplified method of evaluating dose-effect experiments. J Pharmacol Exp Ther. 1949;96(2):99–113. [PubMed] [Google Scholar]
- 24.Maha Abdulrahman Alamin AIY, Yagi SM. Evaluation of Antidiabetic Activity of Plants Used in Western Sudan. Asian Pacific J Trop. Biomed. 2015;5(5):395–402. doi: 10.1016/S2221-1691(15)30375-0. [DOI] [Google Scholar]
- 25.Ahmed MF, Kazim SM, Ghori SS, Mehjabeen SS, Ahmed SR, Ali SM, Ibrahim M. Antidiabetic Activity of Vinca rosea Extracts in Alloxan-Induced Diabetic Rats. Int J Endocrinol. 2010;2010:841090. doi: 10.1155/2010/841090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lenzen S. The mechanisms of alloxan- and streptozotocin-induced diabetes. Diabetologia. 2008;51(2):216–226. doi: 10.1007/s00125-007-0886-7. [DOI] [PubMed] [Google Scholar]
- 27.Katoh M, Sakurai K, Fujimoto Y. Alloxan radical-induced generation of reactive oxygen species in the reaction system of alloxan with ascorbate. Yakugaku Zasshi. 2002;122(10):831–839. doi: 10.1248/yakushi.122.831. [DOI] [PubMed] [Google Scholar]
- 28.Kwon YI, Vattem DA, Shetty K. Evaluation of clonal herbs of Lamiaceae species for management of diabetes and hypertension. Asia Pac J Clin Nutr. 2006;15(1):107–118. [PubMed] [Google Scholar]
- 29.Ceriello A. Postprandial hyperglycemia and diabetes complications: is it time to treat? Diabetes. 2005;54(1):1–7. doi: 10.2337/diabetes.54.1.1. [DOI] [PubMed] [Google Scholar]
- 30.Chipiti T, Ibrahim MA, Singh M, Islam MS. In vitro alpha-amylase and alpha-glucosidase inhibitory effects and cytotoxic activity of Albizia antunesiana extracts. Pharmacogn Mag. 2015;11(Suppl 2):S231–S236. doi: 10.4103/0973-1296.166018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hanamura T, Hagiwara T, Kawagishi H. Structural and functional characterization of polyphenols isolated from acerola (Malpighia emarginata DC.) fruit. Biosci Biotechnol Biochem. 2005;69(2):280–286. doi: 10.1271/bbb.69.280. [DOI] [PubMed] [Google Scholar]
- 32.Kunga MR, Balsamy T, Thayumanavan P, Palanisamy R. Inhibitory effect of Gymnema Montanum leaves on α-glucosidase activity and α-amylase activity and their relationship with polyphenolic content. Med Chem Res. 2010;19(8):948–961. doi: 10.1007/s00044-009-9241-5. [DOI] [Google Scholar]
- 33.Martin MA, Ramos S, Cordero-Herrero I, Bravo L, Goya L. Cocoa phenolic extract protects pancreatic beta cells against oxidative stress. Nutrients. 2013;5(8):2955–2968. doi: 10.3390/nu5082955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lee JS, Kim YR, Park JM, Ha SJ, Kim YE, Baek NI, Hong EK. Mulberry fruit extract protects pancreatic beta-cells against hydrogen peroxide-induced apoptosis via antioxidative activity. Molecules. 2014;19(7):8904–8915. doi: 10.3390/molecules19078904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Islam F, Raihan O, Chowdhury D, Khatun M, Zuberi N, Khatun L, Brishti A, Bahar E. Apoptotic and antioxidant activities of methanol extract of Mussaenda roxburghii leaves. Pak J Pharm Sci. 2015;28(6):2027–2034. [PubMed] [Google Scholar]
- 36.Federiuk IF, Casey HM, Quinn MJ, Wood MD, Ward WK. Induction of type-1 diabetes mellitus in laboratory rats by use of alloxan: route of administration, pitfalls, and insulin treatment. Comp Med. 2004;54(3):252–257. [PubMed] [Google Scholar]
- 37.Underwood EJ. Trace metals in human and animal health. Journal of human nutrition. 1981;35(1):37–48. doi: 10.3109/09637488109143484. [DOI] [PubMed] [Google Scholar]
- 38.Middleton E, Jr, Kandaswami C, Theoharides TC. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer. Pharmacol Rev. 2000;52(4):673–751. [PubMed] [Google Scholar]
- 39.Pareek H, Sharma S, Khajja BS, Jain K, Jain GC. Evaluation of hypoglycemic and anti-hyperglycemic potential of Tridax procumbens (Linn.) BMC Complement Altern Med. 2009;9:48. doi: 10.1186/1472-6882-9-48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Dahech I, Belghith KS, Hamden K, Feki A, Belghith H, Mejdoub H. Oral administration of levan polysaccharide reduces the alloxan-induced oxidative stress in rats. Int J Biol Macromol. 2011;49(5):942–947. doi: 10.1016/j.ijbiomac.2011.08.011. [DOI] [PubMed] [Google Scholar]
- 41.Al-Khalifa A, Mathew TC, Al-Zaid NS, Mathew E, Dashti HM. Therapeutic role of low-carbohydrate ketogenic diet in diabetes. Nutrition. 2009;25(11–12):1177–1185. doi: 10.1016/j.nut.2009.04.004. [DOI] [PubMed] [Google Scholar]
- 42.Bathina S, Srinivas N, Das UN. BDNF protects pancreatic beta cells (RIN5F) against cytotoxic action of alloxan, streptozotocin, doxorubicin and benzo(a)pyrene in vitro. Metabolism. 2016;65(5):667–684. doi: 10.1016/j.metabol.2016.01.016. [DOI] [PubMed] [Google Scholar]
- 43.Bahar E, Kim H, Yoon H. ER Stress-Mediated Signaling: Action Potential and Ca(2+) as Key Players. Int J Mol Sci. 2016;17(9). [DOI] [PMC free article] [PubMed]
- 44.Vessal M, Hemmati M, Vasei M. Antidiabetic effects of quercetin in streptozocin-induced diabetic rats. Comp Biochem Physiol C Toxicol Pharmacol. 2003;135C(3):357–364. doi: 10.1016/S1532-0456(03)00140-6. [DOI] [PubMed] [Google Scholar]
- 45.Polasa K, Naidu AN, Ravindranath I, Krishnaswamy K. Inhibition of B(a)P induced strand breaks in presence of curcumin. Mutat Res. 2004;557(2):203–213. doi: 10.1016/j.mrgentox.2003.10.016. [DOI] [PubMed] [Google Scholar]
- 46.Bahar E, Lee GH, Bhattarai KR, Lee HY, Choi MK, Rashid HO, Kim JY, Chae HJ, Yoon H: Polyphenolic Extract of Euphorbia supina Attenuates Manganese-Induced Neurotoxicity by Enhancing Antioxidant Activity through Regulation of ER Stress and ER Stress-Mediated Apoptosis. Int. J. Mol. Sci. 2017, 18(2). [DOI] [PMC free article] [PubMed]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data sets supporting the conclusions of this article are presented in this main paper. Plant materials used in this study have been identified at the University of Chittagong (CU) where voucher specimens are deposited.