

Abstract
The chimeric antigen receptor T (CAR-T) cell therapy is a newly developed adoptive antitumor treatment. Theoretically, CAR-T cells can specifically localize and eliminate tumor cells by interacting with the tumor-associated antigens (TAAs) expressing on tumor cell surface. Current studies demonstrated that various TAAs could act as target antigens for CAR-T cells, for instance, the type III variant epidermal growth factor receptor (EGFRvIII) was considered as an ideal target for its aberrant expression on the cell surface of several tumor types. CAR-T cell therapy has achieved gratifying breakthrough in hematological malignancies and promising outcome in solid tumor as showed in various clinical trials. The third generation of CAR-T demonstrates increased antitumor cytotoxicity and persistence through modification of CAR structure. In this review, we summarized the preclinical and clinical progress of CAR-T cells targeting EGFR, human epidermal growth factor receptor 2 (HER2), and mesothelin (MSLN), as well as the challenges for CAR-T cell therapy.
Keywords: CAR-T cell, EGFR, HER2, Mesothelin, Solid tumors
Background
Over a century, immunology has been employed to treat malignant tumors, such as monoclonal antibody (mAb), bispecific antibody, tumor vaccine, immune checkpoint blockade, cytokine-induced killer (CIKs), tumor-infiltrating lymphocytes (TILs), and most recently chimeric antigen receptor T (CAR-T) [1]. Application of monoclonal antibodies (Herceptin, cetuximab) in malignant tumor patients showed a satisfying response rate. Immune checkpoint blockades are emerging immunotherapies against tumors. Pembrolizumab, nivolumab (anti-PD-1mAb), and ipilimumab (anti-CTLA-4mAb), which are representative immune checkpoint blocking agent, have been approved by the Food and Drug Administration (FDA) for melanoma patients, as either initial therapy or after relapse [2]. The CAR-T-based immunotherapy has achieved significant progress in malignant hematological diseases. CARs are synthetic receptors consisting of extracellular single-chain variable fragment (scFv), transmembrane domain, and intracellular part of immunoreceptor tyrosine-based activation motifs (ITAMs) and co-stimulatory signal (Fig. 1) [3]. The scFv is responsible for recognizing and binding to tumor-associated antigens (TAAs) expressed on the tumor cell surface. The endodomain plays a pivotal role in T cell activation, proliferation, persistence, and cytotoxicity. The structure of CAR is similar to T cell receptor (TCR), but the scFv of CAR recognizes TAAs independent of major histocompatibility complex (MHC) and targets a variety of antigens expressed on the surface of the tumor cell, including proteins, carbohydrates, and gangliosides (Fig. 1) [4, 5]. The first generation of CARs merely includes activation signal CD3 zeta chain (CD3ζ) or Fc receptor γ (FcRγ) in intracellular motif, thus inducing transient T cell activation [6]. The second and third generation of CARs including one activation domain and one or more costimulatory domains (CD28, 4-1BB, or OX40) were developed and contributed to the expansion, prolonged antitumor activity, and cytokine secretion (such as IL-2, TNFα, and IFN-γ) of T cell (Fig. 1) [7, 8]. Currently, anti-CD19 CAR-T cells were demonstrated to be effective in the treatment of B cell non-Hodgkin lymphoma (NHL), acute lymphoblastic leukemia (ALL), and chronic lymphocytic leukemia (CLL) [9–13]. Anti-CD116 has been developed for treating myelomonocytic leukemia [14].
Fig. 1.
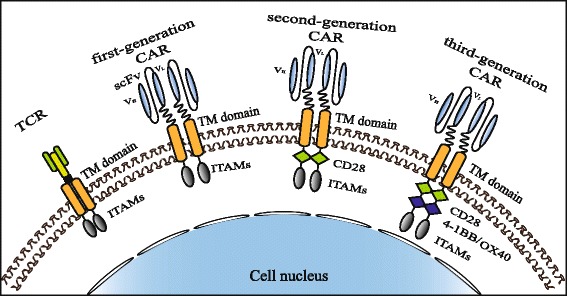
The structure of TCR and the three generations of CAR. T cell receptor (TCR) includes antigen-binding domain, transmembrane domain (TM domain), and immune receptor tyrosine-based activation motifs (ITAMs). The binding domain of CAR consists of a scFv, comprising the light (VL) and heavy (VH) variable fragments of a TAA-specific monoclonal antibody joined by a flexible linker. The intracellular parts are different between the three generations of CAR. The first-generation CAR only has the signal transduction domain of the CD3-zeta chain (CD3ζ) or Fc receptor γ (FcRγ) which mediated transient persistence, inefficient cytotoxicity, and low-level cytokine secretion. The second and third generation CAR add one or more co-stimulatory domains (CD28, 4-1BB, or OX40) to the first generation, which lead to the enhanced cytotoxicity and cytokine secretion along with prolonged T cell persistence
Adoptive cellular therapy (ACT) using CAR-T cells is also a novel way for the treatment of other malignant tumors [15]. In solid tumors, epidermal growth factor receptor (EGFR), human epidermal growth factor receptor2 (HER2), carcinoembryonic antigen (CEA), disialoganglioside 2 (GD2), mesothelin, prostate-specific membrane antigen (PSMA), and interleukin-13Ra2 (IL13Ra2) are known as the targets of CAR-T cells. We summarized the current CAR-T cell-targeted antigens in Table 1. In this review, we mainly introduced the correlated studies of EGFR, HER2, and mesothelin-specific CAR-T cells. Those TAAs are commonly expressed on solid tumors and have been developed by multi-research institutes. More importantly, some studies have achieved promising outcome.
Table 1.
Tumor-associated antigens of CAR-T cell target
| Antigen | Full name | Disease |
|---|---|---|
| EGFR | Epidermal growth factor receptor | NSCLC, epithelial carcinoma, glioma |
| EGFRvIII | Variant III of the epidermal growth factor receptor | Glioblastoma |
| HER2 | Human epidermal growth factor receptor 2 | Ovarian cancer, breast cancer, glioblastoma, colon cancer, osteosarcoma, medulloblastoma |
| MSLN | Mesothelin | Mesothelioma, ovarian cancer, pancreatic adenocarcinoma |
| PSMA | Prostate-specific membrane antigen | Prostate cancer |
| CEA | Carcinoembryonic antigen | Pancreatic adenocarcinoma, breast cancer, colorectal carcinoma |
| GD2 | Disialoganglioside 2 | Neuroblastoma, melanoma |
| IL13Rα2 | Interleukin-13Ra2 | Glioma |
| GPC3 | Glypican-3 | Hepatocellular carcinoma |
| CAIX | Carbonic anhydrase IX | Renal cell carcinoma (RCC) |
| L1-CAM | L1 cell adhesion molecule | Neuroblastoma, melanoma, ovarian adenocarcinoma |
| CA125 | Cancer antigen 125 (also known as MUC16) | Epithelial ovarian cancers |
| CD133 | Cluster of differentiation 133 (also known as prominin-1) | Glioblastoma, cholangiocarcinoma (CCA) |
| FAP | Fibroblast activation protein | Malignant pleural mesothelioma (MPM) |
| CTAG1B | Cancer/testis antigen 1B (also known as NY-ESO-1) | Melanoma and ovarian cancer |
| MUC1 | Mucin 1 | Seminal vesicle cancer |
| FR-α | Folate receptor-α | Ovarian cancer |
Antitumor mechanism of CAR-T cells
CAR-T cells recognize specific tumor antigens in a MHC-independent manner, which lead to the activation and execution of its antitumor function [16]. Once CAR specifically binds with TAAs, T cells are activated through the phosphorylation of immune receptor tyrosine-based activation motifs (ITAMs) and subsequently induce cytokine secretion, T cell proliferation, and cytotoxicity [17]. The original T cells, including CD8+ and CD4+ T cells, are isolated from peripheral blood or tumor tissues of patients. It is generally agreed that CD8+ T cells play a critical part in immune responses against tumors, and CD4+ T cells can help to enhance the efficiency of CD8+ T cell-mediated cytotoxicity [18]. Chimeric immunoreceptor-activated T lymphocytes perform cytotoxicity through two predominant pathways: (1) secretion of perforin and granzyme granules and (2) activation of death receptor signaling via Fas/Fas-ligand (Fas-L) or TNF/TNF-R. CD8+ T cells kill tumor cells through those two pathways. CD4+ T cells destroy target cells primarily via perforin/granzyme, while death receptor-mediated apoptosis is believed to function as a compensatory pathway [19, 20]. Many strategies have been employed to potentiate the functions of CAR-T cells. It has been demonstrated that CAR-T cells with multiple signaling receptors could improve amplification, cytokine production, and cytotoxicity of T cells, as well as reduce antigen-induced cell death (AICD) in vitro and in vivo [21]. CD40L-modified T cells enhanced the proliferation and secretion of proinflammatory Th1 cytokines, including IL-2, IFN-γ, IL-12, and TNF [22]. CD28 costimulation was critical for antigen-specific cytokine secretion and T cell proliferation without obvious effect on the receptor-mediated target cell lysis [23]. IL-12 enhanced the activation of cytotoxic T cell [24], recruited and reinforced the functions of innate immune cells such as NK cell and macrophage [25], enhanced the Th1-type helper T cell response, and exhibited antiangiogenic activities [26]. On this basis, T cells redirected for universal cytokine killing (TRUCK) was developed. TRUCK is a way to redirect CAR-T cells by producing and releasing a transgenic product, such as IL-12, to activate innate immune response against tumor cells which are invisible to CAR-T cells [4]. Besides targeting antigen-specific tumor cell, IFN-γ secreted by CAR-T cells contributed to the antigen-independent destruction of tumor cell through IFNγR expressed in tumor stroma [27]. Neeson et al. developed a novel transgenic mouse model CAR OT-I. CAR OT-I cells not only recognized target tumor cells and secreted cytotoxic granule proteins (perforin, granzyme B) but also induced serial killing which were observed in real time via time-lapse microscopy [28]. In addition, the outcome of clinical application of CAR T cells could be improved by strengthening the function of CAR-T cells through co-activation of macrophage and NK cell (Fig. 2).
Fig. 2.

Antitumor mechanism of CAR-T. a TCR recognizes TAAs depending on the MHC presentation. The advantage is that TCR could recognize intracellular and extracellular antigens. While tumor cells often downregulate MHC expression to escape the killer T cells, b CAR-T cells can specifically recognize the tumor antigens in a MHC-independent manner. And then, the T cells were activated through the phosphorylation of ITAMs followed by enhanced cytokine (include IL-2, IL-4, IFN-γ, IL-12, and TNF) secretion, T cell proliferation, and cytotoxicity. IL-12 could recruit and reinforce the functions of innate immune cells such as NK cell and macrophage. Activated T and CAR-T cells perform cytotoxicity mainly through secretion of perforin and granzyme granules, also through the death receptor pathway such as Fas/Fas-L. Due to added co-stimulatory signal to endodomain, antitumor activity mediated by CARs is stronger than TCRs
Target antigen expressing on solid tumor cell surface
In this part, we summarized the preclinical and clinical studies of CAR-T antigens in solid tumors, focusing on the common targets of EGFR, HER2, and mesothelin. Emphases were put on the scientific basic and progress in preclinical experiments of CAR-T cells.
EGFR
EGFR is a 170-KDa transmembrane receptor tyrosine kinase belonging to the ErbB (also known as HER) oncogene family [29–31]. EGFR is expressed in the skin, gastrointestinal system, kidney, and other normal tissues at the physiological level; however, it is aberrantly activated in many epithelial tumors, such as lung cancer, pancreatic cancer, colorectal cancer, breast cancer, and head and neck squamous cell carcinoma (HNSCC) [32, 33]. EGFR plays central roles in regulation of cellular multiplication, differentiation, and metastasis, and the overexpression of EGFR is related to a more aggressive clinical progression and poor prognosis [34, 35]. In fact, EGFR has been a therapy target for many years. Currently, targeted-EGFR antitumor agent is mainly divided into two categories: anti-EGFR monoclonal antibodies (mAbs) and small-molecule tyrosine kinase inhibitors (TKIs) [36]. Anti-EGFR mAbs prevent the EGF binding and receptor activation by occupying the ligand-binding site of the EGFR. TKIs inhibit autophosphorylation and downstream intracellular signaling of EGFR [37]. Two mAbs (cetuximab and panitumumab) and two TKIs (gefitinib and erlotinib), as first-generation EGFR inhibitors, have been used for the treatment of NSCLC, pancreatic cancer, HNSCC, renal cancer, and colorectal cancer (CRC) [38]. Nevertheless, the therapeutic efficacy of the EGFR inhibitors was attenuated in some patients, resulting from EGFR mutations and the acquired drug resistance. Hence, to create novel therapeutic strategies to overcome the defects is imperative [39, 40]. Anti-EGFR CAR-T therapy is an alternative strategy for EGFR overexpression malignant cancers, although the application of CAR-T therapy toward solid tumors remains challenging [41]. The most common oncogenic EGFR mutant is the type III EGFR (EGFRvIII), which results in an in-frame deletion of exons 2 to 7 [42, 43]. EGFRvIII appears to meet most of the criteria of ideal antigen for CAR-T therapy, for it is the most commonly altered form of EGFR in cancers with no expression in normal tissues [44]. Expression of EGFRvIII promotes tumor cell growth, invasion, migration, and therapeutic resistance and is associated with poor long-term survival [45, 46].
Preclinical studies on EGFR-specific CAR-T cells
Glioblastoma (GBM) remains one of the deadliest primary brain tumors in adults, and standard treatments for GBM do not significantly increase the survival time. EGFRvIII is expressed in GBM cell surface; therefore, CAR-T cell targeting EGFRvIII is a novel strategy worth studying [47]. Morgan et al. conducted a series of experiments to construct competent CARs and evaluated the ability of CAR-engineered T cells recognizing EGFRvIII. Considering that the established cell lines may not keep the molecular characteristics of primary human cancers, Morgan group selected the glioblastoma stem cells (GSCs) expressing EGFRvIII as target cell lines. CAR scFv derived from human mAb 139 recognized GSCs expressing mutant EGFRvIII, but not human normal tissues. T cell signaling transduction domain CD28-41BB-CD3ζ (named 139-28BBZ) made CAR-T keep better survival comparing with the original CAR vector that used CD28-CD3ζ (named 139-28Z) [48–50], but the biological activity and cytotoxicity were at the equal level. The engineered T cells expressing CAR can specifically recognize EGFRvIII+ cell lines, while no reactivity to co-cultured normal tissue cells. At present, a phase I clinical trial (NCT01454596) using anti-EGFRvIII CAR-T cells is recruiting patients with recurrent glioblastoma [51]. Study by Marcela et al. also evaluated the characteristics of anti-EGFRvIII CAR-T cells and verified its antitumor activity against glioblastoma cells in vitro and vivo [52]. The humanized anti-EGFRvIII CAR-T cells produced IFN-γ, IL-2, TNF-α, and only lysed EGFRvIII-expressing target cells. In order to confirm the antitumor activity in vivo, U87-EGFRvIII tumors were implanted subcutaneously and intracranially into NSG mice, respectively. The results indicated that CAR-T-EGFRvIII cells controlled tumor growth and increased median survival time. This group also used mice grafted with normal human skin to test the potential toxicities of anti-EGFRvIII CAR-T cells, and the results of skin graft assay demonstrated that no significant lymphocytic infiltrate by immunohistochemistry. On this basis, Marcela group started a phase 1 clinical trial (NCT02209376) of EGFRvIII-specific CAR-T cells in patients with either residual or recurrent glioblastoma [53]. D-270MG is a tumor cell line that naturally expresses EGFRvIII [54]. Sampson et al. established the D-270MGFLuc/GFP subline that co-expressed firefly luciferase (FLuc) and GFP as the target of EGFR-specific CAR-T cells. The study results demonstrated that anti-EGFRvIII CAR-T cells effectively transpassed the blood-brain barrier (BBB) to arrive at invasive GBM tumors and mediated tumor regression and prolonged survival in NSG mice [55]. Zuo et al. used the EGFR-positive (EGFR+) cells including A549, NCI-H1299, NCI-H460, SGC7901, HT29, and EGFR-knockdown (EGFR−) cells including A549-EGFR−, SGC7901-EGFR−, and HT-29-EGFR− to investigate the antitumor activity of EGFR-specific CAR-CIK cells. The study reported that EGFR-specific CAR observably potentiated cytotoxicity and induced secretion of IFN-γ and IL-2 in EGFR-positive cell lines and xenograft tumor models, but not in EGFR negative ones [56]. In summary, the preclinical studies of EGFR-specific CAR-T cells exhibited potent antitumor effect in vitro and in vivo.
Clinical trials on EGFR-specific CAR-T cells
Multicenter clinical trials using CAR-T cells targeting EGFR or EGFRvIII are underway. We summarized these clinical trials in Table 2. A phase I trial by Han et al. studied the EGFR-targeted CAR-T cells in 11 patients with EGFR-expressing advanced relapsed/refractory NSCLC (NCT01869166). In this study, the six female and five male patients were divided into three cohorts: in cohort 1, EGFR-CAR T cells infused into four patients directly without any conditioning regimens; in cohort 2, two patients were conditioned with cyclophosphamide, followed by CAR-T EGFR therapy; and in cohort 3, two patients were conditioned by cyclophosphamide, pemetrexed, and cisplatin and three were conditioned by cyclophosphamide, docetaxel, and cisplatin, respectively. All patients received EGFR-targeted CAR-T cell infusions at dose ranged from 0.45 to 1.09 × 107 cells/kg. Of 11 patients, there were two persons acquired PR and five kept stable disease (SD). The anti-EGFR CAR-T cells secreted cytokines including IL-2, IL-4, IL-6, IFN-γ, TNF-α, GM-CSF, and granzyme B in co-culture with EGFR-positive tumor cells. However, after the infusion of EGFR-specific CAR-T cells, the serum levels of cytokines observed at different time point were less obvious compared with the experiment in vitro. Investigators monitored the copy numbers of CAR-EGFR transgene in peripheral blood (seven patients) and tumor tissues (four patients) by quantitative real-time PCR. In peripheral blood, the copy numbers of CAR-EGFR transgene hold high level for more than 4 weeks. CAR-EGFR transgene specifically accumulated in tumor tissues. The only tolerable and controllable toxicities reported in the study were skin toxicity, nausea, vomiting, dyspnea, and hypotension, and there was no cytokine storm observed. Therefore, the CAR-T-EGFR cells were found to be feasible and safe in patients of relapsed/refractory NSCLC [57].
Table 2.
Clinical trials of CAR-T cells
| Target | Identifier | Institution | Phase | Status | Disease | Comments |
|---|---|---|---|---|---|---|
| EGFR | NCT02331693 | Shanghai Cancer Institute | I | Recruiting | Glioma | Autologous T cells transduced with a lentiviral vector |
| EGFRvIII | NCT02844062 | Beijing Sanbo Brain Hospital | I | Recruiting | Glioma | Lymphodepletion chemotherapy, followed by CAR-T |
| EGFRvIII | NCT01454596 | National Cancer Institute (NCI) | I/II | Recruiting | Glioma | Autologous T cells, a retroviral vector |
| EGFRvIII | NCT02209376 | University of Pennsylvania/University of California | I | Recruiting | Glioma | Autologous T cells, a lentiviral vector |
| EGFRvIII | NCT02664363 | Duke University Medical Center | I | Not yet recruiting | Glioma | Dose escalation cohorts for 4 dose levels |
| EGFR | NCT01869166 | Chinese PLA General Hospital | I/II | Completed | NSCLC | Safe and feasible |
| HER2 | NCT02713984 | Southwest Hospital, China | I/II | Recruiting | HER2-positive cancer | |
| HER2 | NCT01935843 | Chinese PLA General Hospital | I/II | Recruiting | HER2-positive cancer | |
| HER2 | NCT02547961 | Fuda Cancer Hospital Guangzhou | I/II | Recruiting | Breast cancer | A retrovirus vector, preconditioning treatment |
| HER2 | NCT02442297 | Baylor College of Medicine | I | Recruiting | Glioma | Intracranial injection |
| HER2 | NCT01109095 | Baylor College of Medicine | I | Active, not recruiting | Glioma | CMV-specific cytotoxic T cells (CMV-T cells) |
| HER2 | NCT00889954 | Baylor College of Medicine | I | Active, not recruiting | HER2-positive cancer | TGFBeta resistant HER2/EBV-CTLs |
| HER2 | NCT00924287 | National Cancer Institute (NCI) | I/II | Completed | HER2-positive sarcoma | Results is not encouraging |
| HER2 | NCT00902044 | Baylor College of Medicine | I/II | Completed | HER2-positive sarcoma | Safe and feasible |
| MSLN | NCT02930993 | China Meitan General Hospital | I | Recruiting | Mesothelin-positive tumors | Followed lymphodepletion |
| MSLN | NCT02706782 | Shanghai Renji Hospital | I | Recruiting | Pancreatic cancer | Transcatheter arterial infusion |
| MSLN | NCT02159716 | University of Pennsylvania | I | Active not recruiting | Mesothelin-positive tumors | Lentiviral transduced |
| MSLN | NCT02792114 | Memorial Sloan Kettering Cancer Center | I | Recruiting | Mesothelin-expressing breast cancer | Premedicated with acetaminophen and diphenhydramine, and administered cyclophosphamide |
| MSLN | NCT02465983 | University of Pennsylvania | I | Active not recruiting | Pancreatic cancer | Combination therapy with CART-meso cells and CART19 cells |
| MSLN | NCT02959151 | Shanghai Tumor Hospital | I/II | Recruiting | Pancreatic cancer | Combined with interventional therapy |
| MSLN | NCT02590747 | Chinese PLA General Hospital | I | Recruiting | Mesothelin-positive tumors | Retroviral vector-transduced |
| MSLN | NCT01355965 | University of Pennsylvania | I | Completed | Pleural mesothelioma | Safe and feasible |
| MSLN | NCT02414269 | Memorial Sloan Kettering Cancer Center | I | Recruiting | Malignant pleural disease | With/without chemotherapy |
| MSLN | NCT01583686 | National Cancer Institute (NCI) | I/II | Recruiting | Mesothelin-positive tumors | Followed lymphodepletion |
| MSLN | NCT01897415 | University of Pennsylvania | I | Active not recruiting | Pancreatic ductal adenocarcinoma (PDA) | Transfected with chimeric anti-mesothelin immunoreceptor SS1 |
| IL13Rα2 | NCT00730613 | City of Hope Medical Center | I | Completed | Glioblastoma | Safe and feasible |
| IL13Rα2 | NCT02208362 | City of Hope Medical Center | I | Completed | Glioblastoma | Safe and feasible |
| CEA | NCT01373047 | Roger Williams Medical Center | I | Completed | Liver metastases | Safe and feasible |
| FAP | NCT01722149 | University of Zurich | I | Recruiting | Malignant pleural mesothelioma | Followed lymphodepletion |
| GD2 | NCT00085930 | Baylor College of Medicine | I | Completed | Neuroblastoma | Safe and feasible |
| GD2 | NCT02107963 | National Cancer Institute (NCI) | I | Completed | Sarcoma Osteosarcoma Neuroblastoma Melanoma |
Safe and feasible |
| CD133 | NCT02541370 | Chinese PLA General Hospital | I | Recruiting | CD133-positive malignancies | Relapsed and/or chemotherapy refractory advanced malignancies |
The details of Table 2 derived from http://clinicaltrials.gov/
HER2
HER2 is a 185-KDa transmembrane glycoprotein which also belongs to the family of the EGFR [58, 59]. HER2 gene amplification or HER2 overexpression plays a crucial role in the biologic behavior and pathogenesis of some type of human cancers [60]. HER2 is overexpressed in 25–30% of breast and ovarian cancers [61], up to 60% of human osteosarcomas (OS) [62], approximately 80% of GBM [63], and 40% of medulloblastomas but is not detected in normal cerebellum and other brain tissues [64]. Overexpression of HER2 is associated with cellular transformation and carcinogenesis and also correlated with poor clinical outcome [65, 66]. On this basis, HER2 monoclonal antibody trastuzumab (Herceptin) was first approved for use in patients with HER2-overpressed breast cancer. Trastuzumab alone or in combination with chemotherapy prolongs survival in both primary and metastatic breast cancer [67]. At present, the clinical trials about HER2 tyrosine kinase inhibitors such as lapatinib and neratinib are still ongoing [68]. However, many tumors such as osteosarcoma, glioblastoma, and medulloblastoma expressing HER2 at low levels are ineffectively recognized by trastuzumab [66]. In addition, approximately half of those patients either do not respond to these therapies or develop secondary resistance which results to treatment failure [69, 70]. Therefore, it is necessary to create novel therapeutic approach to treat these patients.
Preclinical studies on HER2-specific CAR-T cells
In GBMs, CD133-positive stem cells keep higher expression of HER2 than CD133-negative counterparts. A study result indicated that HER2-specific CAR-T cells targeted and killed autologous HER2-positive GBMs in vitro and facilitated regression of GBMs in an orthotopic xenograft model [71]. Sun et al. constructed a humanized HER2 CAR-T cell containing chA21scFv and examined its antitumor activity. The results indicated that chA21-28z HER2-specific CAR-T cells recognized and killed HER2+ breast and ovarian cancer cells in vitro. Simultaneously, abundant IFN-γ and IL-2 secretion were also detected. In xenograft model, the HER2-specific CAR-T cells also significantly restricted tumor growth [72]. Another study demonstrated that oligoclonal camelid single-domain antibodies (VHHs) could target a range of different epitopes on HER2 antigen. Based on the potent targeting ability of oligoclonal VHHs, the oligoclonal VHHHER2-CAR-engineered Jurkat T cells exhibited higher expansion, cytokine secretion, and cytotoxicity when exposed to HER2-expressing cells [73]. To reduce antigen escape, Hegdeet et al. created a bispecific CAR molecule co-targeting the two glioma-associated antigens, HER2 and IL-13Rα2, and expanded the CAR-T cells expressing tandem CARs (TanCAR). Encouragingly, the TanCAR effectively redirected T cells to the two antigens and enhanced the function of CAR-T cells and the secretion of cytokines in vitro and in vivo. Therefore, the TanCAR-T cell agents were considered as a potential therapeutic method to control tumor growth as this study reported [74, 75]. Recently, a group combined bispecific antibody αHER2/CD3 and CAR-T therapy. Their data indicated that αHER2/CD3 RNA-engineered T cells exhibited antitumor activity in HER2+ N87 tumor cells and in N87 tumor-bearing mice. Moreover, bystander T cells also showed the similar effects. This new strategy may be a potential therapeutic approach for HER2+ malignancies [76]. To promote the transduction efficiency, EBV-CTLs were modified to express HER2-CAR via the nonviral piggyBac (PB) transposon which had high gene-transfer efficiency and large coding capacity. PB-modified HER2-CTLs could specifically target and kill HER2-positive tumor cells in vivo and suppress tumor growth in xenogeneic murine models [77]. Although 60% human osteosarcoma expressed HER2 [62, 78], a low level of HER2 renders monoclonal antibodies to HER2 ineffective. Hence, a group used genetic-modified T cell targeting HER2 to determine the antitumor activity in osteosarcoma. The HER2-specific CAR-T cells proliferated, produced cytokines, and killed tumor cells after exposure to HER2-positive osteosarcoma cell lines in vitro. Moreover, they created two mouse models: one is locoregional disease in a severe combined immune deficiency (SCID) mouse model and the other is lung metastases model. Adoptive transfer of HER2-specific CAR-T cells caused osteosarcoma regression at the different sites [79]. Similarly, HER2-specific CAR-T cells had the capacity of recognizing and killing HER2-positive medulloblastoma cells in vitro and induced regression of tumors in an orthotopic xenogeneic SCID model [64]. These preclinical studies have achieved encouraging results, promoting HER2-specific CAR-T clinical trials to test the feasibility and safety.
Clinical trials on HER2-specific CAR-T cells
At present, Southwest Hospital in China, Chinese PLA General Hospital, Fuda Cancer Hospital Guangzhou, and Baylor College of Medicine are carrying out clinical trials of HER2-specific CAR-T cells. We summarized these clinical trials in Table 2. Phase I/II clinical study (NCT00924287) sponsored by National Cancer Institute (NCI) has completed. This trial was designed to evaluate the safety and efficacy of HER2-specific CAR-T cells in patients with relapsed/refractory HER2-positive sarcoma. Nineteen patients received escalating doses (range 1 × 104/m2 to 1 × 108/m2) of HER2-specific CAR-T cells including eight dose levels. The study reported that among the detected serum cytokines, only the concentration of IL-8 had significantly increased within 1 week after infusion and persisted for up to 4 weeks. Although HER2-specific CAR-T cells had no expansion after infusion in the peripheral blood, these cells could traffic to tumor sites and maintain at low levels for more than 6 weeks. T cell persistence and copy number were correlated with the infused T cell dose. The clinical benefit of HER2-specific CAR-T cell was not encouraging, only four of nineteen patients acquired stable disease (SD). In the process of HER2-specific CAR-T cell infusion, dose-limiting toxicity was not observed apart from a patient with the highest dose levels within 12 h post-infusion [80].
Mesothelin
Mesothelin (MSLN) is a 40-KDa cell surface tumor differentiation antigen, which derived from the 69-KDa precursor protein encoded by Mesothelin gene [81, 82]. The normal biological function of mesothelin almost remains unknown. Some studies suggest that mesothelin is the receptor of CA125/MUC16, and the interaction between mesothelin-CA125 mediates cell adhesion and may be a critical point in the metastatic of ovarian cancer [83, 84]. Mesothelin overexpression promotes tumor cell proliferation and regional invasion and is associated with poor prognosis, such as worse recurrence-free survival (RFS) and overall survival (OS) [85–87]. As a tumor marker, soluble mesothelin in serum plays an important role in diagnosing and monitoring therapeutic effect for patients with malignant pleural mesothelioma (MPM) and ovarian cancer [88–91]. Mesothelin is expressed at low levels in normal tissues, including pleura, pericardium, peritoneum, tunica vaginalis [92–94], but it is overexpressed in various malignancies including MPM, ovarian cancers, pancreatic cancers, and non-small cell lung cancers [95–98]. Due to the weak expression in normal tissues and strong expression in several cancers, mesothelin is considered as an attractive target for immune-based therapies [81]. With respect to mesothelin-targeted therapies such as anti-mesothelin recombinant immunotoxin SS1P, chimeric anti-mesothelin monoclonal antibody MORAb-009, and mesothelin cancer vaccines CRS-207, investigators performed a lot of preclinical researches and opened a series of clinical trials [99–102]. Simultaneously, a number of studies about CAR-T cells targeting mesothelin are in progress.
Preclinical studies on MSLN-specific CAR-T cells
June et al. demonstrated that the mesothelin-specific T cells exhibit antitumor effects on large pre-established mesothelioma xenografts in NOD/scid/IL2rγ−/− mice. Their data suggested that the combination of CD137 and CD28 improved multifunctional cytokine secretion and enhanced the function of CAR T cells in tumor-bearing mice [103]. In the tumor microenvironment, some inhibitors hampered the function of CAR T cells. For example, diacylglycerol kinase (dgk), as a negative regulator of TCR signaling, is expressed in T cells. Its isoform includes dgkα and dgkζ. Previous studies found that deletion of either dgk isoform induced the activation of DAG-mediated Ras/ERK pathway and proliferation of T cells [104–106]. Based on this, Koretzky et al. demonstrated that deletion of dgks greatly enhanced activity against tumor and improved persistence of CAR-engineered T cells targeting mesothelin in vitro and in implanted tumors. Beyond that, pharmacologic inhibition of dgks also facilitated function of mesothelin-specific CAR-T cells. Moreover, dgk-deficient T cells showed decreased sensitivity to TGFβ and increased FasL and TRAIL expression. Such a combined therapeutic approach might be translated clinically as the study reported [107]. Moon et al. found that a single intravenous injection of human mesoCAR-T cells into immunodeficient mice significantly restrained the tumor growth but did not cure tumor. They considered that upregulation of inhibitory receptors was the main cause of mesoCAR-T cell hypofunction [108]. As an inhibitor within the tumor microenvironment, upregulation of PD-1 limited T cell function [109]. Cherkassky et al. found that PD-1 antibody could reverse PD-1-mediated CAR-T cell exhaustion and mesoCAR-T cells also showed delayed exhaustion upon repeated antigen stimulation. Hence, combination of costimulation and cell-intrinsic PD-1 checkpoint blockade could overcome inhibitory effect on CAR-T cells in MSLN-expressing tumor microenvironment [110]. CAR-T therapy achieved good results in preclinical studies. But the effect was not satisfied in the clinical trials mainly due to its adverse effects. For example, scFv was generally derived from murine monoclonal antibodies; the induction of human anti-mouse antibody (HAMA) might shorten T cell survival time [111]. A study demonstrated that fully human mesothelin-specific CAR-T cells showed potent cytolytic activity toward mesothelin-positive tumor cells and controlled large, well-established ovarian cancer growth in a xenogeneic mouse model. Besides, mesothelin-specific CAR-T cells induced bystander killing of mesothelin-negative tumor cells [112]. On-target/off-tumor toxicity could cause life-threatening adverse effects in the application of CAR-T cells, because the target antigen also expressed on normal cell surface at low levels. Both a-folate receptor (FRa) (90%) and mesothelin (70%) were overexpressed in ovarian cancers [113, 114], and their expression pattern on normal tissues is mainly non-overlapping. Based on the foundation of above studies, Daniel et al. generated trans-signaling CAR T cells engineered to co-express anti-mesoscFv-CD3 and anti- FRascFv-CD28CARs, aiming to diminish the potential toxicity of CAR-T cells to normal tissue cells expressing low levels of TAAs. The result indicated that trans-signaling CAR-T cells exhibited higher antitumor potential in vitro and in vivo. Moreover, trans-signaling CAR-T cells were resistant to antigen-induced cell death (AICD) [115]. The successes achieved by CAR-T cells in hematological malignancies were unable to be accomplished in solid tumor, partly owing to the low efficacy of CAR-T cells homing to tumor sites. Stimulating more chemokine receptors expressed on CAR-T cells or direct regional injection may be valid. Chemokine CCL2 is highly expressed by MPM tumors, but the expression level of CCL2 receptor CCR2 on resting and activated T cells is low. Therefore, Moon et al. transduced the chemokine receptor CCR2b into mesoCAR-T cells to potentiate trafficking of CAR-T cells into tumors. Their study demonstrated that the functional CCR2b in the mesoCAR-T cells significantly increased the number of intratumoral T cells and improved antitumor efficacy in vitro and in vivo [116]. Adusumilli et al. found that compared with intravenous injection, intrapleural administration of anti-mesothelin CAR-T cells exhibited greater antitumor potency and strongly promoted the expansion, differentiation, and persistence of T cells [117].
Clinical trials on MSLN-specific CAR-T cells
Many clinical trials about mesothelin-specific CAR-T cells are ongoing. We summarized these clinical trials in Table 2. Marcela et al. started a clinical study in four patients infused with autologous T cells transducted with mRNA to express CAR derived from a murine antibody to human mesothelin. These results demonstrated that when patients received intermittent infusion of meso-RNA CAR-T cells, the serum IgE levels detected via ELISA assay were elevated which caused anaphylaxis. Therefore, they suggested that a single infusion of stably transducted, long-lived CAR-T cells or constructing CAR based on the humanized antibodies may be safer and more effect [52]. The phase I clinical trial (NCT01355965) conducted by Beatty et al. was designed to improve the feasibility and safety of mRNA-transduced CAR-T cells targeting mesothelin (mesoCAR-T cells) in patients with advanced MPM. They presented two case reports indicating that mRNA CAR-T cells showed potent antitumor activity without evident on-target/off-tumor toxicity against normal tissues, infiltrated solid tumor tissues, and induced humoral epitope spreading after infusion [118].
Other target antigens
In addition, there are lots of tumor-associated antigens studied by investigators in solid tumors. CA125 also called MUC16 is a well-known ovarian tumor antigen routinely used for monitoring disease. To enhance the antitumor efficacy, Brentjens et al. developed T cells co-expressing MUC16 CAR and IL-12, and the results were as expected both in vitro and in vivo [119]. Based on the rationale, they opened a phase I clinical trial in patients with recurrent ovarian cancer [120]. Carbonic anhydrase IX (CAIX) is an attractive target antigen because it is overexpressed in renal cell carcinoma (RCC) but is not found on normal kidney tissue. The CAIX-specific CAR-T cells inhibited tumor growth in xenograft model [121]. Several malignant tumors including pancreatic adenocarcinoma, breast cancer, and colorectal carcinoma overexpressed carcinoembryonic antigen (CEA). Guest et al. generated CAR-T cells for the phase I/II clinical trial of CEA-specific CAR-T therapy in 14 patients with advanced CEA+ malignancy [122]. At present, clinical trials of anti-CEA CAR-T cells in Advanced Liver Malignancy (NCT02959151) and CAR-T Cells Targeting CEA Positive Cancer (NCT02349724) are ongoing. Neuroblastoma is a high-risk extracranial malignant tumor of childhood. Disialoganglioside (GD2) is overexpressed in almost all neuroblastoma. Therefore, GD2 is an ideal candidate of CAR-T cells. The preclinical and clinical studies of GD2-specific CAR-T cells have achieved some progress [123, 124]. Moreover, the clinical trial of GD2-specific CAR-T therapy in 19 patients with advanced neuroblastoma has completed by Louis et al. It was showed that eight achieved remission and 11 with active disease [125]. A study reported that the GD2-specific CAR-T cells showed anti-melanoma activity in vitro and in vivo [126]. Similar to GD2, L1 cell adhesion molecule (L1-CAM) is also overexpressed in neuroblastoma. In addition, ovarian adenocarcinoma, medulloblastoma, and melanoma all highly expressed L1-CAM [127]. Investigators tested the antitumor efficacy and safety in preclinical and clinical studies [128–130]. Glypican 3 (GPC3) is highly expressed in hepatocellular carcinoma (HCC) and hepatoblastoma. Study results demonstrated that all GPC3-CAR-T cells showed potent cytotoxicity to GPC3-positive cells [131]. Aiming at GPC3 and asialoglycoprotein receptor1 (ASGR1) another TAA in HCC, a group developed the dual-targeted CAR-T cells. They found that dual-targeted CAR-T cells caused higher proliferation, antitumor activity, and cytokine secretion than signal-targeted CAR-T cells in vitro [132]. Prostate-specific membrane antigen (PSMA) was expressed in prostate cancer cells. PSMA-targeted CAR-T cells exhibited superior antitumor efficacy in vitro. In established models, PSMA-targeted CAR-T cells also effectively eliminated prostate cancer [133–135]. CD133, as a specific molecular biomarker for CSCs, is an attractive therapeutic target for CAR-T therapy [136, 137]. CD133-specific CAR-T cells in a patient with advanced cholangiocarcinoma have shown antitumor activity [138]. At present, a phase I clinical trial of anti-CD133 CAR-T cells in patients with relapsed and/or chemotherapy refractory advanced malignancies is ongoing (NCT02541370). In addition to above antigens, fibroblast activation protein (FAP) [139, 140], NY-ESO-1 [141], MUC1 [142], foliate receptor [143, 144], and IL13Rα2 [145, 146] are also potential target antigens for immunotherapy.
Conclusions
In this review, we summarized the current preclinical and clinical studies on CAR-T therapy against solid tumors, especially targeting EGFR, HER2, and MSLN. The ideal target for CAR-T cells would be the tumor-specific antigens which are homogenously expressed on the surface of malignant cell and play a critical role in tumorigenesis. Although the curative effect in CAR-T treatments of hematological malignancies are reported, the results of pilot clinical trials on solid cancers are below expectation. Several obstacles have remained to be overcome for a successful application of CAR-T cells in solid tumor, including the lack of ideal TAAs, inefficient trafficking of CAR-T cells to tumor sites, hostile solid tumor microenvironment, and the risk of developing on-target/off-tumor toxicities [15, 17]. To solve the problems, investigators have developed some strategies to potentiate the trafficking of CAR-T cells [116], reduce the inhabitation effect of tumor microenvironment [110], decrease the adverse effects, and so on [115]. In general, the preclinical studies of CAR-T cells in vitro and in vivo showed potent antitumor efficacy; with further exploration to improve the feasibility, safety, and efficiency of CAR-T cells, CAR-T therapy will take the central stage in the treatment of solid tumors.
Acknowledgements
Not applicable.
Funding
This review was supported by National Natural Science Foundation of China (Grant No. 81572608) and the National High Technology Research and Development Program of China (No. 2015AA020301).
Availability of data and materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Authors’ contributions
SY and AL searched the literatures and wrote the manuscript. QL, TL, and XY helped to collect the literatures and participated in the discussion. KW and XH designed the study. All authors read and approved the final manuscript.
Competing interests
The authors declare that they no competing interests.
Consent for publication
All authors have read and approved the final manuscript for publication.
Ethics approval and consent to participate
Not applicable.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- ACT
Adoptive cellular therapy
- AICD
Antigen-induced cell death
- ALL
Acute lymphoblastic leukemia
- ASGR1
Asialoglycoprotein receptor 1
- CAIX
Carbonic anhydrase IX
- CARs
Chimeric antigen receptors
- CD3ζ
CD3 zeta chain
- CEA
Carcinoembryonic antigen
- CIKs
Cytokine-induced killer
- CLL
Chronic lymphocytic leukemia
- CRC
Colorectal cancer
- dgk
diacylglycerol kinase
- EGFR
Epidermal growth factor receptor
- EGFRvIII
Type III variant epidermal growth factor receptor
- Fas-L
Fas-ligand
- FcRγ
Fc receptor γ
- FDA
Food and Drug Administration
- GBM
Glioblastoma
- GD2
Disialoganglioside
- HAMA
Human anti-mouse antibody
- HCC
Hepatocellular carcinoma
- HER2
Human epidermal growth factor receptor2
- HNSCC
Head and neck squamous cell carcinoma
- IL13Ra2
Interleukin-13Ra2
- ITAMs
Immunoreceptor tyrosine-based activation motifs
- mAb
Monoclonal antibody
- MHC
Major histocompatibility complex
- MPM
Malignant pleural mesothelioma
- MSLN
Mesothelin
- NCI
National Cancer Institute
- NHL
Non-Hodgkin lymphoma
- NK
Natural killer
- OS
Osteosarcomas
- PB
PiggyBac
- PSMA
Prostate-specific membrane antigen
- RCC
Renal cell carcinoma
- RFS
Recurrence-free survival
- scFv
Single-chain variable fragment
- SCID
Severe combined immune deficiency
- SD
Stable disease
- TAAs
Tumor-associated antigens
- TanCAR
Tandem CARs
- TCR
T cell receptor
- TILs
Tumor-infiltrating lymphocytes
- TKIs
Tyrosine kinase inhibitors
- TRUCK
T cells redirected for universal cytokine killing
Contributor Information
Shengnan Yu, Email: 1334870996@qq.com.
Anping Li, Email: Li_anping@yahoo.com.
Qian Liu, Email: 521095072@qq.com.
Tengfei Li, Email: 50632953@qq.com.
Xun Yuan, Email: yuanxunjackson@163.com.
Xinwei Han, Email: hanxinwei2006@163.com.
Kongming Wu, Email: kmwu@tjh.tjmu.edu.cn.
References
- 1.Khalil DN, Budhu S, Gasmi B, Zappasodi R, Hirschhorn-Cymerman D, Plitt T, et al. The new era of cancer immunotherapy: manipulating T-cell activity to overcome malignancy. Adv Cancer Res. 2015;128:1–68. doi: 10.1016/bs.acr.2015.04.010. [DOI] [PubMed] [Google Scholar]
- 2.Mellman I, Coukos G, Dranoff G. Cancer immunotherapy comes of age. Nature. 2011;480(7378):480–489. doi: 10.1038/nature10673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Maher J, Wilkie S, Davies DM, Arif S, Picco G, Julien S, Foster J, Burchell J, Taylor-Papadimitriou J. Targeting of Tumor-Associated Glycoforms of MUC1 with CAR T Cells. Immunity. 2016;45(5):945-946. [DOI] [PubMed]
- 4.Chmielewski M, Hombach AA, Abken H. Of CARs and TRUCKs: chimeric antigen receptor (CAR) T cells engineered with an inducible cytokine to modulate the tumor stroma. Immunol Rev. 2014;257(1):83–90. doi: 10.1111/imr.12125. [DOI] [PubMed] [Google Scholar]
- 5.Fesnak AD, June CH, Levine BL. Engineered T cells: the promise and challenges of cancer immunotherapy. Nat Rev Cancer. 2016;16(9):566–581. doi: 10.1038/nrc.2016.97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brocker T, Karjalainen K. Signals through T cell receptor-zeta chain alone are insufficient to prime resting T lymphocytes. J Exp Med. 1995;181(5):1653–1659. doi: 10.1084/jem.181.5.1653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Finney HM, Akbar AN, Lawson ADG. Activation of resting human primary T cells with chimeric receptors: costimulation from CD28, inducible costimulator, CD134, and CD137 in series with signals from the TCR chain. J Immunol. 2003;172(1):104–113. doi: 10.4049/jimmunol.172.1.104. [DOI] [PubMed] [Google Scholar]
- 8.Di S, Li Z. Treatment of solid tumors with chimeric antigen receptor-engineered T cells: current status and future prospects. Sci China Life Sci. 2016;59(4):360–369. doi: 10.1007/s11427-016-5025-6. [DOI] [PubMed] [Google Scholar]
- 9.Liu J, Zhong JF, Zhang X, Zhang C. Allogeneic CD19-CAR-T cell infusion after allogeneic hematopoietic stem cell transplantation in B cell malignancies. J Hematol Oncol. 2017;10(1):35. doi: 10.1186/s13045-017-0405-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang X, Popplewell LL, Wagner JR, Naranjo A, Blanchard MS, Mott MR, et al. Phase 1 studies of central memory-derived CD19 CAR T-cell therapy following autologous HSCT in patients with B-cell NHL. Blood. 2016;127(24):2980–2990. doi: 10.1182/blood-2015-12-686725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schubert ML, Huckelhoven A, Hoffmann JM, Schmitt A, Wuchter P, Sellner L, et al. Chimeric antigen receptor T cell therapy targeting CD19-positive leukemia and lymphoma in the context of stem cell transplantation. Hum Gene Ther. 2016 Jul 31. [Epub ahead of print] [DOI] [PubMed]
- 12.Maude SL, Frey N, Shaw PA, Aplenc R, Barrett DM, Bunin NJ, et al. Chimeric antigen receptor T cells for sustained remissions in leukemia. N Engl J Med. 2014;371(16):1507–1517. doi: 10.1056/NEJMoa1407222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cai B, Guo M, Wang Y, Zhang Y, Yang J, Guo Y, et al. Co-infusion of haplo-identical CD19-chimeric antigen receptor T cells and stem cells achieved full donor engraftment in refractory acute lymphoblastic leukemia. J Hematol Oncol. 2016;9(1):131. doi: 10.1186/s13045-016-0357-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nakazawa Y, Matsuda K, Kurata T, Sueki A, Tanaka M, Sakashita K, et al. Anti-proliferative effects of T cells expressing a ligand-based chimeric antigen receptor against CD116 on CD34+ cells of juvenile myelomonocytic leukemia. J Hematol Oncol. 2016;9(1):27. doi: 10.1186/s13045-016-0256-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Zhang E, Xu H. A new insight in chimeric antigen receptor-engineered T cells for cancer immunotherapy. J Hematol Oncol. 2017;10(1):1. doi: 10.1186/s13045-016-0379-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cartellieri M, Bachmann M, Feldmann A, Bippes C, Stamova S, Wehner R, et al. Chimeric antigen receptor-engineered T cells for immunotherapy of cancer. J Biomed Biotechnol. 2010;2010:956304. doi: 10.1155/2010/956304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kershaw MH, Westwood JA, Slaney CY, Darcy PK. Clinical application of genetically modified T cells in cancer therapy. Clin Transl Immunol. 2014;3(5):e16. doi: 10.1038/cti.2014.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pereira BI, Akbar AN. Convergence of innate and adaptive immunity during human aging. Front Immunol. 2016;7:445. doi: 10.3389/fimmu.2016.00445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Yasukawa M, Ohminami H, Arai J, Kasahara Y, Ishida Y, Fujita S. Granule exocytosis, and not the fas/fas ligand system, is the main pathway of cytotoxicity mediated by alloantigen-specific CD4(+) as well as CD8(+) cytotoxic T lymphocytes in humans. Blood. 2000;95(7):2352–2355. [PubMed] [Google Scholar]
- 20.Hombach A, Kohler H, Rappl G, Abken H. Human CD4+ T cells lyse target cells via granzyme/perforin upon circumvention of MHC class II restriction by an antibody-like immunoreceptor. J Immunol. 2006;177(8):5668–5675. doi: 10.4049/jimmunol.177.8.5668. [DOI] [PubMed] [Google Scholar]
- 21.Long AH, Haso WM, Shern JF, Wanhainen KM, Murgai M, Ingaramo M, et al. 4-1BB costimulation ameliorates T cell exhaustion induced by tonic signaling of chimeric antigen receptors. Nat Med. 2015;21(6):581–590. doi: 10.1038/nm.3838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Curran KJ, Seinstra BA, Nikhamin Y, Yeh R, Usachenko Y, van Leeuwen DG, et al. Enhancing antitumor efficacy of chimeric antigen receptor T cells through constitutive CD40L expression. Mol Ther. 2015;23(4):769–778. doi: 10.1038/mt.2015.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hombach A, Sent D, Schneider C, Heuser C, Koch D, Pohl C, et al. T-cell activation by recombinant receptors: CD28 costimulation is required for interleukin 2 secretion and receptor-mediated T-cell proliferation but does not affect receptor-mediated target cell lysis. Cancer Res. 2001;61(5):1976–1982. [PubMed] [Google Scholar]
- 24.Kerkar SP, Muranski P, Kaiser A, Boni A, Sanchez-Perez L, Yu Z, et al. Tumor-specific CD8+ T cells expressing interleukin-12 eradicate established cancers in lymphodepleted hosts. Cancer Res. 2010;70(17):6725–6734. doi: 10.1158/0008-5472.CAN-10-0735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pegram HJ, Lee JC, Hayman EG, Imperato GH, Tedder TF, Sadelain M, et al. Tumor-targeted T cells modified to secrete IL-12 eradicate systemic tumors without need for prior conditioning. Blood. 2012;119(18):4133–4141. doi: 10.1182/blood-2011-12-400044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Chmielewski M, Kopecky C, Hombach AA, Abken H. IL-12 release by engineered T cells expressing chimeric antigen receptors can effectively muster an antigen-independent macrophage response on tumor cells that have shut down tumor antigen expression. Cancer Res. 2011;71(17):5697–5706. doi: 10.1158/0008-5472.CAN-11-0103. [DOI] [PubMed] [Google Scholar]
- 27.Textor A, Listopad JJ, Wuhrmann LL, Perez C, Kruschinski A, Chmielewski M, et al. Efficacy of CAR T-cell therapy in large tumors relies upon stromal targeting by IFNgamma. Cancer Res. 2014;74(23):6796–6805. doi: 10.1158/0008-5472.CAN-14-0079. [DOI] [PubMed] [Google Scholar]
- 28.Davenport AJ, Jenkins MR, Cross RS, Yong CS, Prince HM, Ritchie DS, et al. CAR-T cells inflict sequential killing of multiple tumor target cells. Cancer Immunol Res. 2015;3(5):483–494. doi: 10.1158/2326-6066.CIR-15-0048. [DOI] [PubMed] [Google Scholar]
- 29.Hynes NE, MacDonald G. ErbB receptors and signaling pathways in cancer. Curr Opin Cell Biol. 2009;21(2):177–184. doi: 10.1016/j.ceb.2008.12.010. [DOI] [PubMed] [Google Scholar]
- 30.Hynes NE, Lane HA. ERBB receptors and cancer: the complexity of targeted inhibitors. Nat Rev Cancer. 2005;5(5):341–354. doi: 10.1038/nrc1609. [DOI] [PubMed] [Google Scholar]
- 31.Olayioye MA, Neve RM, Lane HA, Hynes NE. The ErbB signaling network: receptor heterodimerization in development and cancer. EMBO J. 2000;19(13):3159–3167. doi: 10.1093/emboj/19.13.3159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yano S, Kondo K, Yamaguchi M, Richmond G, Hutchison M, Wakeling A, et al. Distribution and function of EGFR in human tissue and the effect of EGFR tyrosine kinase inhibition. Anticancer Res. 2003;23(5a):3639–3650. [PubMed] [Google Scholar]
- 33.Sasada T, Azuma K, Ohtake J, Fujimoto Y. Immune responses to epidermal growth factor receptor (EGFR) and their application for cancer treatment. Front Pharmacol. 2016;7:405. doi: 10.3389/fphar.2016.00405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Arteaga CL. Epidermal growth factor receptor dependence in human tumors: more than just expression? Oncologist. 2002;7(Suppl 4):31–39. doi: 10.1634/theoncologist.7-suppl_4-31. [DOI] [PubMed] [Google Scholar]
- 35.Shinojima N, Tada K, Shiraishi S, Kamiryo T, Kochi M, Nakamura H, et al. Prognostic value of epidermal growth factor receptor in patients with glioblastoma multiforme. Cancer Res. 2003;63(20):6962–6970. [PubMed] [Google Scholar]
- 36.Yewale C, Baradia D, Vhora I, Patil S, Misra A. Epidermal growth factor receptor targeting in cancer: a review of trends and strategies. Biomaterials. 2013;34(34):8690–8707. doi: 10.1016/j.biomaterials.2013.07.100. [DOI] [PubMed] [Google Scholar]
- 37.Baselga J. Critical update and emerging trends in epidermal growth factor receptor targeting in cancer. J Clin Oncol. 2005;23(11):2445–2459. doi: 10.1200/JCO.2005.11.890. [DOI] [PubMed] [Google Scholar]
- 38.Ciardiello F, Tortora G. A novel approach in the treatment of cancer: targeting the epidermal growth factor receptor. Clin Cancer Res. 2001;7(10):2958–2970. [PubMed] [Google Scholar]
- 39.Ohashi K, Maruvka YE, Michor F, Pao W. Epidermal growth factor receptor tyrosine kinase inhibitor-resistant disease. J Clin Oncol. 2013;31(8):1070–1080. doi: 10.1200/JCO.2012.43.3912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Chong CR, Janne PA. The quest to overcome resistance to EGFR-targeted therapies in cancer. Nat Med. 2013;19(11):1389–1400. doi: 10.1038/nm.3388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhang H, Ye ZL, Yuan ZG, Luo ZQ, Jin HJ, Qian QJ. New strategies for the treatment of solid tumors with CAR-T cells. Int J Biol Sci. 2016;12(6):718–729. doi: 10.7150/ijbs.14405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Pedersen MW, Meltorn M, Damstrup L, Poulsen HS. The type III epidermal growth factor receptor mutation. Biological significance and potential target for anti-cancer therapy. Ann Oncol. 2001;12(6):745–760. doi: 10.1023/A:1011177318162. [DOI] [PubMed] [Google Scholar]
- 43.Heimberger AB, Suki D, Yang D, Shi W, Aldape K. The natural history of EGFR and EGFRvIII in glioblastoma patients. J Transl Med. 2005;3:38. doi: 10.1186/1479-5876-3-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Maus MV, Designing CAR. T cells for glioblastoma. Oncoimmunology. 2015;4(12):e1048956. doi: 10.1080/2162402X.2015.1048956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Luo X, Xie H, Long X, Zhou M, Xu Z, Shi B, et al. EGFRvIII mediates hepatocellular carcinoma cell invasion by promoting S100 calcium binding protein A11 expression. PLoS One. 2013;8(12):e83332. doi: 10.1371/journal.pone.0083332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Del Vecchio CA, Jensen KC, Nitta RT, Shain AH, Giacomini CP, Wong AJ. Epidermal growth factor receptor variant III contributes to cancer stem cell phenotypes in invasive breast carcinoma. Cancer Res. 2012;72(10):2657–2671. doi: 10.1158/0008-5472.CAN-11-2656. [DOI] [PubMed] [Google Scholar]
- 47.Eskilsson E, Rosland GV, Talasila KM, Knappskog S, Keunen O, Sottoriva A, et al. EGFRvIII mutations can emerge as late and heterogenous events in glioblastoma development and promote angiogenesis through Src activation. Neuro Oncol. 2016;18(12):1644–1655. doi: 10.1093/neuonc/now113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Zhong XS, Matsushita M, Plotkin J, Riviere I, Sadelain M. Chimeric antigen receptors combining 4-1BB and CD28 signaling domains augment PI3kinase/AKT/Bcl-XL activation and CD8+ T cell-mediated tumor eradication. Mol Ther. 2010;18(2):413–420. doi: 10.1038/mt.2009.210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhao Y, Wang QJ, Yang S, Kochenderfer JN, Zheng Z, Zhong X, Sadelain M, et al. A Herceptin-based chimeric antigen receptor with modified signaling domains leads to enhanced survival of transduced T lymphocytes and antitumor activity. J Immunol. 2009;183(9):5563–5574. doi: 10.4049/jimmunol.0900447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Song DG, Ye Q, Carpenito C, Poussin M, Wang LP, Ji C, et al. In vivo persistence, tumor localization, and antitumor activity of CAR-engineered T cells is enhanced by costimulatory signaling through CD137 (4-1BB) Cancer Res. 2011;71(13):4617–4627. doi: 10.1158/0008-5472.CAN-11-0422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Morgan RA, Johnson LA, Davis JL, Zheng Z, Woolard KD, Reap EA, et al. Recognition of glioma stem cells by genetically modified T cells targeting EGFRvIII and development of adoptive cell therapy for glioma. Hum Gene Ther. 2012;23(10):1043–1053. doi: 10.1089/hum.2012.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Maus MV, Haas AR, Beatty GL, Albelda SM, Levine BL, Liu X, et al. T cells expressing chimeric antigen receptors can cause anaphylaxis in humans. Cancer Immunol Res. 2013;1(1):26–31. doi: 10.1158/2326-6066.CIR-13-0006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Johnson LA, Scholler J, Ohkuri T, Kosaka A, Patel PR, McGettigan SE, et al. Rational development and characterization of humanized anti-EGFR variant III chimeric antigen receptor T cells for glioblastoma. Sci Transl Med. 2015;7(275):275ra222. doi: 10.1126/scitranslmed.aaa4963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Bigner SH, Humphrey PA, Wong AJ, Vogelstein B, Mark J, Friedman HS, et al. Characterization of the epidermal growth factor receptor in human glioma cell lines and xenografts. Cancer Res. 1990;50(24):8017–8022. [PubMed] [Google Scholar]
- 55.Miao H, Choi BD, Suryadevara CM, Sanchez-Perez L, Yang S, De Leon G, et al. EGFRvIII-specific chimeric antigen receptor T cells migrate to and kill tumor deposits infiltrating the brain parenchyma in an invasive xenograft model of glioblastoma. PLoS One. 2014;9(4):e94281. doi: 10.1371/journal.pone.0094281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ren X, Ma W, Lu H, Yuan L, An L, Wang X, et al. Modification of cytokine-induced killer cells with chimeric antigen receptors (CARs) enhances antitumor immunity to epidermal growth factor receptor (EGFR)-positive malignancies. Cancer Immunol Immunother. 2015;64(12):1517–1529. doi: 10.1007/s00262-015-1757-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Feng K, Guo Y, Dai H, Wang Y, Li X, Jia H, et al. Chimeric antigen receptor-modified T cells for the immunotherapy of patients with EGFR-expressing advanced relapsed/refractory non-small cell lung cancer. Sci China Life Sci. 2016;59(5):468–479. doi: 10.1007/s11427-016-5023-8. [DOI] [PubMed] [Google Scholar]
- 58.Yarden Y, Sliwkowski MX. Untangling the ErbB signalling network. Nat Rev Mol Cell Biol. 2001;2(2):127–137. doi: 10.1038/35052073. [DOI] [PubMed] [Google Scholar]
- 59.Cho HS, Mason K, Ramyar KX, Stanley AM, Gabelli SB, Denney DW, Jr, et al. Structure of the extracellular region of HER2 alone and in complex with the Herceptin Fab. Nature. 2003;421(6924):756–760. doi: 10.1038/nature01392. [DOI] [PubMed] [Google Scholar]
- 60.Hudziak RM, Schlessinger J, Ullrich A. Increased expression of the putative growth factor receptor p185HER2 causes transformation and tumorigenesis of NIH 3T3 cells. Proc Natl Acad Sci U S A. 1987;84(20):7159–7163. doi: 10.1073/pnas.84.20.7159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Slamon DJ, Godolphin W, Jones LA, Holt JA, Wong SG, Keith DE, et al. Studies of the HER-2/neu proto-oncogene in human breast and ovarian cancer. Science. 1989;244(4905):707–712. doi: 10.1126/science.2470152. [DOI] [PubMed] [Google Scholar]
- 62.Gorlick R, Huvos AG, Heller G, Aledo A, Beardsley GP, Healey JH, et al. Expression of HER2/erbB-2 correlates with survival in osteosarcoma. J Clin Oncol. 1999;17(9):2781–2788. doi: 10.1200/JCO.1999.17.9.2781. [DOI] [PubMed] [Google Scholar]
- 63.Zhang JG, Kruse CA, Driggers L, Hoa N, Wisoff J, Allen JC, et al. Tumor antigen precursor protein profiles of adult and pediatric brain tumors identify potential targets for immunotherapy. J Neurooncol. 2008;88(1):65–76. doi: 10.1007/s11060-008-9534-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ahmed N, Ratnayake M, Savoldo B, Perlaky L, Dotti G, Wels WS, et al. Regression of experimental medulloblastoma following transfer of HER2-specific T cells. Cancer Res. 2007;67(12):5957–5964. doi: 10.1158/0008-5472.CAN-06-4309. [DOI] [PubMed] [Google Scholar]
- 65.Thompson SK, Sullivan TR, Davies R, Ruszkiewicz AR. Her-2/neu gene amplification in esophageal adenocarcinoma and its influence on survival. Ann Surg Oncol. 2011;18(7):2010–2017. doi: 10.1245/s10434-011-1554-1. [DOI] [PubMed] [Google Scholar]
- 66.Serrano-Olvera A, Duenas-Gonzalez A, Gallardo-Rincon D, Candelaria M, De la Garza-Salazar J. Prognostic, predictive and therapeutic implications of HER2 in invasive epithelial ovarian cancer. Cancer Treat Rev. 2006;32(3):180–190. doi: 10.1016/j.ctrv.2006.01.001. [DOI] [PubMed] [Google Scholar]
- 67.Valabrega G, Montemurro F, Aglietta M. Trastuzumab: mechanism of action, resistance and future perspectives in HER2-overexpressing breast cancer. Ann Oncol. 2007;18(6):977–984. doi: 10.1093/annonc/mdl475. [DOI] [PubMed] [Google Scholar]
- 68.Nonagase Y, Yonesaka K, Kawakami H, Watanabe S, Haratani K, Takahama T, et al. Heregulin-expressing HER2-positive breast and gastric cancer exhibited heterogeneous susceptibility to the anti-HER2 agents lapatinib, trastuzumab and T-DM1. Oncotarget. 2016;7(51):84860–84871. doi: 10.18632/oncotarget.12743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Cheng YC, Valero V, Davis ML, Green MC, Gonzalez-Angulo AM, Theriault RL, et al. Addition of GM-CSF to trastuzumab stabilises disease in trastuzumab-resistant HER2+ metastatic breast cancer patients. Br J Cancer. 2010;103(9):1331–1334. doi: 10.1038/sj.bjc.6605918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Alexander PB, Chen R, Gong C, Yuan L, Jasper JS, Ding Y, et al. Distinct receptor tyrosine kinase subsets mediate anti-HER2 drug resistance in breast cancer. J Biol Chem. 2017;292(2):748–759. doi: 10.1074/jbc.M116.754960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Ahmed N, Salsman VS, Kew Y, Shaffer D, Powell S, Zhang YJ, et al. HER2-specific T cells target primary glioblastoma stem cells and induce regression of autologous experimental tumors. Clin Cancer Res. 2010;16(2):474–485. doi: 10.1158/1078-0432.CCR-09-1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Sun M, Shi H, Liu C, Liu J, Liu X, Sun Y. Construction and evaluation of a novel humanized HER2-specific chimeric receptor. Breast Cancer Res. 2014;16(3):R61. doi: 10.1186/bcr3674. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Jamnani FR, Rahbarizadeh F, Shokrgozar MA, Mahboudi F, Ahmadvand D, Sharifzadeh Z, et al. T cells expressing VHH-directed oligoclonal chimeric HER2 antigen receptors: towards tumor-directed oligoclonal T cell therapy. Biochim Biophys Acta. 2014;1840(1):378–386. doi: 10.1016/j.bbagen.2013.09.029. [DOI] [PubMed] [Google Scholar]
- 74.Hegde M, Corder A, Chow KK, Mukherjee M, Ashoori A, Kew Y, et al. Combinational targeting offsets antigen escape and enhances effector functions of adoptively transferred T cells in glioblastoma. Mol Ther. 2013;21(11):2087–2101. doi: 10.1038/mt.2013.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Hegde M, Mukherjee M, Grada Z, Pignata A, Landi D, Navai SA, et al. Tandem CAR T cells targeting HER2 and IL13Ralpha2 mitigate tumor antigen escape. J Clin Invest. 2016;126(8):3036–3052. doi: 10.1172/JCI83416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Luo F, Qian J, Yang J, Deng Y, Zheng X, Liu J, et al. Bifunctional alphaHER2/CD3 RNA-engineered CART-like human T cells specifically eliminate HER2(+) gastric cancer. Cell Res. 2016;26(7):850–853. doi: 10.1038/cr.2016.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Nakazawa Y, Huye LE, Salsman VS, Leen AM, Ahmed N, Rollins L, et al. PiggyBac-mediated cancer immunotherapy using EBV-specific cytotoxic T-cells expressing HER2-specific chimeric antigen receptor. Mol Ther. 2011;19(12):2133–2143. doi: 10.1038/mt.2011.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hughes DP, Thomas DG, Giordano TJ, Baker LH, McDonagh KT. Cell surface expression of epidermal growth factor receptor and Her-2 with nuclear expression of Her-4 in primary osteosarcoma. Cancer Res. 2004;64(6):2047–2053. doi: 10.1158/0008-5472.CAN-03-3096. [DOI] [PubMed] [Google Scholar]
- 79.Ahmed N, Salsman VS, Yvon E, Louis CU, Perlaky L, Wels WS, et al. Immunotherapy for osteosarcoma: genetic modification of T cells overcomes low levels of tumor antigen expression. Mol Ther. 2009;17(10):1779–1787. doi: 10.1038/mt.2009.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Ahmed N, Brawley VS, Hegde M, Robertson C, Ghazi A, Gerken C, et al. Human epidermal growth factor receptor 2 (HER2)-specific chimeric antigen receptor-modified T cells for the immunotherapy of HER2-positive sarcoma. J Clin Oncol. 2015;33(15):1688–1696. doi: 10.1200/JCO.2014.58.0225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Hassan R, Bera T, Pastan I. Mesothelin: a new target for immunotherapy. Clin Cancer Res. 2004;10(12 Pt 1):3937–3942. doi: 10.1158/1078-0432.CCR-03-0801. [DOI] [PubMed] [Google Scholar]
- 82.Chang K, Pastan I. Molecular cloning of mesothelin, a differentiation antigen present on mesothelium, mesotheliomas, and ovarian cancers. Proc Natl Acad Sci U S A. 1996;93(1):136–140. doi: 10.1073/pnas.93.1.136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Rump A, Morikawa Y, Tanaka M, Minami S, Umesaki N, Takeuchi M, et al. Binding of ovarian cancer antigen CA125/MUC16 to mesothelin mediates cell adhesion. J Biol Chem. 2004;279(10):9190–9198. doi: 10.1074/jbc.M312372200. [DOI] [PubMed] [Google Scholar]
- 84.Gubbels JA, Belisle J, Onda M, Rancourt C, Migneault M, Ho M, et al. Mesothelin-MUC16 binding is a high affinity, N-glycan dependent interaction that facilitates peritoneal metastasis of ovarian tumors. Mol Cancer. 2006;5(1):50. doi: 10.1186/1476-4598-5-50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Li M, Bharadwaj U, Zhang R, Zhang S, Mu H, Fisher WE, et al. Mesothelin is a malignant factor and therapeutic vaccine target for pancreatic cancer. Mol Cancer Ther. 2008;7(2):286–296. doi: 10.1158/1535-7163.MCT-07-0483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Cheng WF, Huang CY, Chang MC, Hu YH, Chiang YC, Chen YL, et al. High mesothelin correlates with chemoresistance and poor survival in epithelial ovarian carcinoma. Br J Cancer. 2009;100(7):1144–1153. doi: 10.1038/sj.bjc.6604964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kachala SS, Bograd AJ, Villena-Vargas J, Suzuki K, Servais EL, Kadota K, et al. Mesothelin overexpression is a marker of tumor aggressiveness and is associated with reduced recurrence-free and overall survival in early-stage lung adenocarcinoma. Clin Cancer Res. 2014;20(4):1020–1028. doi: 10.1158/1078-0432.CCR-13-1862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Robinson BW, Creaney J, Lake R, Nowak A, Musk AW, de Klerk N, et al. Soluble mesothelin-related protein—a blood test for mesothelioma. Lung Cancer. 2005;49(Suppl 1):S109–111. doi: 10.1016/j.lungcan.2005.03.020. [DOI] [PubMed] [Google Scholar]
- 89.Hassan R, Remaley AT, Sampson ML, Zhang J, Cox DD, Pingpank J, et al. Detection and quantitation of serum mesothelin, a tumor marker for patients with mesothelioma and ovarian cancer. Clin Cancer Res. 2006;12(2):447–453. doi: 10.1158/1078-0432.CCR-05-1477. [DOI] [PubMed] [Google Scholar]
- 90.Cristaudo A, Foddis R, Vivaldi A, Guglielmi G, Dipalma N, Filiberti R, et al. Clinical significance of serum mesothelin in patients with mesothelioma and lung cancer. Clin Cancer Res. 2007;13(17):5076–5081. doi: 10.1158/1078-0432.CCR-07-0629. [DOI] [PubMed] [Google Scholar]
- 91.Scholler N, Fu N, Yang Y, Ye Z, Goodman GE, Hellstrom KE, et al. Soluble member(s) of the mesothelin/megakaryocyte potentiating factor family are detectable in sera from patients with ovarian carcinoma. Proc Natl Acad Sci U S A. 1999;96(20):11531–11536. doi: 10.1073/pnas.96.20.11531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Ordonez NG. Value of mesothelin immunostaining in the diagnosis of mesothelioma. Mod Pathol. 2003;16(3):192–197. doi: 10.1097/01.MP.0000056981.16578.C3. [DOI] [PubMed] [Google Scholar]
- 93.Hassan R, Ho M. Mesothelin targeted cancer immunotherapy. Eur J Cancer. 2008;44(1):46–53. doi: 10.1016/j.ejca.2007.08.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Morello A, Sadelain M, Adusumilli PS. Mesothelin-targeted CARs: driving T cells to solid tumors. Cancer Discov. 2016;6(2):133–146. doi: 10.1158/2159-8290.CD-15-0583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Argani P, Iacobuzio-Donahue C, Ryu B, Rosty C, Goggins M, Wilentz RE, et al. Mesothelin is overexpressed in the vast majority of ductal adenocarcinomas of the pancreas: identification of a new pancreatic cancer marker by serial analysis of gene expression (SAGE) Clin Cancer Res. 2001;7(12):3862–3868. [PubMed] [Google Scholar]
- 96.Ho M, Bera TK, Willingham MC, Onda M, Hassan R, FitzGerald D, et al. Mesothelin expression in human lung cancer. Clin Cancer Res. 2007;13(5):1571–1575. doi: 10.1158/1078-0432.CCR-06-2161. [DOI] [PubMed] [Google Scholar]
- 97.Chang K, Pai LH, Batra JK, Pastan I, Willingham MC. Characterization of the antigen (CAK1) recognized by monoclonal antibody K1 present on ovarian cancers and normal mesothelium. Cancer Res. 1992;52(1):181–186. [PubMed] [Google Scholar]
- 98.Hassan R, Thomas A, Alewine C, Le DT, Jaffee EM, Pastan I. Mesothelin immunotherapy for cancer: ready for prime time? J Clin Oncol. 2016;34(34):4171–4179. doi: 10.1200/JCO.2016.68.3672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Kelly RJ, Sharon E, Pastan I, Hassan R. Mesothelin-targeted agents in clinical trials and in preclinical development. Mol Cancer Ther. 2012;11(3):517–525. doi: 10.1158/1535-7163.MCT-11-0454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Hassan R, Broaddus VC, Wilson S, Liewehr DJ, Zhang J. Anti-mesothelin immunotoxin SS1P in combination with gemcitabine results in increased activity against mesothelin-expressing tumor xenografts. Clin Cancer Res. 2007;13(23):7166–7171. doi: 10.1158/1078-0432.CCR-07-1592. [DOI] [PubMed] [Google Scholar]
- 101.Hassan R, Cohen SJ, Phillips M, Pastan I, Sharon E, Kelly RJ, et al. Phase I clinical trial of the chimeric anti-mesothelin monoclonal antibody MORAb-009 in patients with mesothelin-expressing cancers. Clin Cancer Res. 2010;16(24):6132–6138. doi: 10.1158/1078-0432.CCR-10-2275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Hassan R, Miller AC, Sharon E, Thomas A, Reynolds JC, Ling A, et al. Major cancer regressions in mesothelioma after treatment with an anti-mesothelin immunotoxin and immune suppression. Sci Transl Med. 2013;5(208):208ra147. doi: 10.1126/scitranslmed.3006941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Carpenito C, Milone MC, Hassan R, Simonet JC, Lakhal M, Suhoski MM, et al. Control of large, established tumor xenografts with genetically retargeted human T cells containing CD28 and CD137 domains. Proc Natl Acad Sci U S A. 2009;106(9):3360–3365. doi: 10.1073/pnas.0813101106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Zhong XP, Hainey EA, Olenchock BA, Jordan MS, Maltzman JS, Nichols KE, et al. Enhanced T cell responses due to diacylglycerol kinase zeta deficiency. Nat Immunol. 2003;4(9):882–890. doi: 10.1038/ni958. [DOI] [PubMed] [Google Scholar]
- 105.Olenchock BA, Guo R, Carpenter JH, Jordan M, Topham MK, Koretzky GA, et al. Disruption of diacylglycerol metabolism impairs the induction of T cell energy. Nat Immunol. 2006;7(11):1174–1181. doi: 10.1038/ni1400. [DOI] [PubMed] [Google Scholar]
- 106.Zha Y, Marks R, Ho AW, Peterson AC, Janardhan S, Brown I, et al. T cell energy is reversed by active Ras and is regulated by diacylglycerol kinase-alpha. Nat Immunol. 2006;7(11):1166–1173. doi: 10.1038/ni1394. [DOI] [PubMed] [Google Scholar]
- 107.Riese MJ, Wang LC, Moon EK, Joshi RP, Ranganathan A, June CH, et al. Enhanced effector responses in activated CD8+ T cells deficient in diacylglycerol kinases. Cancer Res. 2013;73(12):3566–3577. doi: 10.1158/0008-5472.CAN-12-3874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Moon EK, Wang LC, Dolfi DV, Wilson CB, Ranganathan R, Sun J, et al. Multifactorial T-cell hypofunction that is reversible can limit the efficacy of chimeric antigen receptor-transduced human T cells in solid tumors. Clin Cancer Res. 2014;20(16):4262–4273. doi: 10.1158/1078-0432.CCR-13-2627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Rizvi NA, Hellmann MD, Snyder A, Kvistborg P, Makarov V, Havel JJ, et al. Cancer immunology. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science. 2015;348(6230):124–128. doi: 10.1126/science.aaa1348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Cherkassky L, Morello A, Villena-Vargas J, Feng Y, Dimitrov DS, Jones DR, et al. Human CAR T cells with cell-intrinsic PD-1 checkpoint blockade resist tumor-mediated inhibition. J Clin Invest. 2016;126(8):3130–3144. doi: 10.1172/JCI83092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Bonifant CL, Jackson HJ, Brentjens RJ, Curran KJ. Toxicity and management in CAR T-cell therapy. Mol Ther Oncolytics. 2016;3:16011. [DOI] [PMC free article] [PubMed]
- 112.Lanitis E, Poussin M, Hagemann IS, Coukos G, Sandaltzopoulos R, Scholler N, et al. Redirected antitumor activity of primary human lymphocytes transduced with a fully human anti-mesothelin chimeric receptor. Mol Ther. 2012;20(3):633–643. doi: 10.1038/mt.2011.256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Hassan R, Kreitman RJ, Pastan I, Willingham MC. Localization of mesothelin in epithelial ovarian cancer. Appl Immunohistochem Mol Morphol. 2005;13(3):243–247. doi: 10.1097/01.pai.00000141545.36485.d6. [DOI] [PubMed] [Google Scholar]
- 114.Kalli KR, Oberg AL, Keeney GL, Christianson TJ, Low PS, Knutson KL, et al. Folate receptor alpha as a tumor target in epithelial ovarian cancer. Gynecol Oncol. 2008;108(3):619–626. doi: 10.1016/j.ygyno.2007.11.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Lanitis E, Poussin M, Klattenhoff AW, Song D, Sandaltzopoulos R, June CH, et al. Chimeric antigen receptor T cells with dissociated signaling domains exhibit focused antitumor activity with reduced potential for toxicity in vivo. Cancer Immunol Res. 2013;1(1):43–53. doi: 10.1158/2326-6066.CIR-13-0008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Moon EK, Carpenito C, Sun J, Wang LC, Kapoor V, Predina J, et al. Expression of a functional CCR2 receptor enhances tumor localization and tumor eradication by retargeted human T cells expressing a mesothelin-specific chimeric antibody receptor. Clin Cancer Res. 2011;17(14):4719–4730. doi: 10.1158/1078-0432.CCR-11-0351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Adusumilli PS, Cherkassky L, Villena-Vargas J, Colovos C, Servais E, Plotkin J, et al. Regional delivery of mesothelin-targeted CAR T cell therapy generates potent and long-lasting CD4-dependent tumor immunity. Sci Transl Med. 2014;6(261):261ra151. doi: 10.1126/scitranslmed.3010162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Beatty GL, Haas AR, Maus MV, Torigian DA, Soulen MC, Plesa G, et al. Mesothelin-specific chimeric antigen receptor mRNA-engineered T cells induce anti-tumor activity in solid malignancies. Cancer Immunol Res. 2014;2(2):112–120. doi: 10.1158/2326-6066.CIR-13-0170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Koneru M, Purdon TJ, Spriggs D, Koneru S, Brentjens RJ. IL-12 secreting tumor-targeted chimeric antigen receptor T cells eradicate ovarian tumors in vivo. Oncoimmunology. 2015;4(3):e994446. doi: 10.4161/2162402X.2014.994446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Koneru M, O'Cearbhaill R, Pendharkar S, Spriggs DR, Brentjens RJ. A phase I clinical trial of adoptive T cell therapy using IL-12 secreting MUC-16(ecto) directed chimeric antigen receptors for recurrent ovarian cancer. J Transl Med. 2015;13:102. doi: 10.1186/s12967-015-0460-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Lo AS, Xu C, Murakami A, Marasco WA. Regression of established renal cell carcinoma in nude mice using lentivirus-transduced human T cells expressing a human anti-CAIX chimeric antigen receptor. Mol Ther Oncolytics. 2014;1:14003. doi: 10.1038/mto.2014.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Guest RD, Kirillova N, Mowbray S, Gornall H, Rothwell DG, Cheadle EJ, et al. Definition and application of good manufacturing process-compliant production of CEA-specific chimeric antigen receptor expressing T-cells for phase I/II clinical trial. Cancer Immunol Immunother. 2014;63(2):133–145. doi: 10.1007/s00262-013-1492-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Seeger RC. Immunology and immunotherapy of neuroblastoma. Semin Cancer Biol. 2011;21(4):229–237. doi: 10.1016/j.semcancer.2011.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Heczey A, Louis CU. Advances in chimeric antigen receptor immunotherapy for neuroblastoma. Discov Med. 2013;16(90):287–294. [PMC free article] [PubMed] [Google Scholar]
- 125.Louis CU, Savoldo B, Dotti G, Pule M, Yvon E, Myers GD, et al. Antitumor activity and long-term fate of chimeric antigen receptor-positive T cells in patients with neuroblastoma. Blood. 2011;118(23):6050–6056. doi: 10.1182/blood-2011-05-354449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Yvon E, Del Vecchio M, Savoldo B, Hoyos V, Dutour A, Anichini A, et al. Immunotherapy of metastatic melanoma using genetically engineered GD2-specific T cells. Clin Cancer Res. 2009;15(18):5852–5860. doi: 10.1158/1078-0432.CCR-08-3163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Rawnaq T, Quaas A, Zander H, Gros SJ, Reichelt U, Blessmann M, et al. L1 is highly expressed in tumors of the nervous system: a study of over 8000 human tissues. J Surg Res. 2012;173(2):314–319. doi: 10.1016/j.jss.2010.10.029. [DOI] [PubMed] [Google Scholar]
- 128.Kunkele A, Taraseviciute A, Finn LS, Johnson AJ, Berger C, Finney O, et al. Preclinical assessment of CD171-directed CAR T-cell adoptive therapy for childhood neuroblastoma: CE7 epitope target safety and product manufacturing feasibility. Clin Cancer Res. 2017;23(2):466–477. doi: 10.1158/1078-0432.CCR-16-0354. [DOI] [PubMed] [Google Scholar]
- 129.Hong H, Stastny M, Brown C, Chang WC, Ostberg JR, Forman SJ, et al. Diverse solid tumors expressing a restricted epitope of L1-CAM can be targeted by chimeric antigen receptor redirected T lymphocytes. J Immunother. 2014;37(2):93–104. doi: 10.1097/CJI.0000000000000018. [DOI] [PubMed] [Google Scholar]
- 130.Park JR, Digiusto DL, Slovak M, Wright C, Naranjo A, Wagner J, et al. Adoptive transfer of chimeric antigen receptor re-directed cytolytic T lymphocyte clones in patients with neuroblastoma. Mol Ther. 2007;15(4):825–833. doi: 10.1038/sj.mt.6300104. [DOI] [PubMed] [Google Scholar]
- 131.Li W, Guo L, Rathi P, Marinova E, Gao X, Wu MF, et al. Redirecting T cells to glypican-3 with 4-1BB zeta chimeric antigen receptors results in Th1 polarization and potent antitumor activity. Hum Gene Ther. 2016 Aug 16. [Epub ahead of print] [DOI] [PMC free article] [PubMed]
- 132.Chen C, Li K, Jiang H, Song F, Gao H, Pan X, et al. Development of T cells carrying two complementary chimeric antigen receptors against glypican-3 and asialoglycoprotein receptor 1 for the treatment of hepatocellular carcinoma. Cancer Immunol Immunother. 2017;66(4):475-489. [DOI] [PMC free article] [PubMed]
- 133.Kloss CC, Condomines M, Cartellieri M, Bachmann M, Sadelain M. Combinatorial antigen recognition with balanced signaling promotes selective tumor eradication by engineered T cells. Nat Biotechnol. 2013;31(1):71–75. doi: 10.1038/nbt.2459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Gade TP, Hassen W, Santos E, Gunset G, Saudemont A, Gong MC, et al. Targeted elimination of prostate cancer by genetically directed human T lymphocytes. Cancer Res. 2005;65(19):9080–9088. doi: 10.1158/0008-5472.CAN-05-0436. [DOI] [PubMed] [Google Scholar]
- 135.Ma Q, Gomes EM, Lo AS, Junghans RP. Advanced generation anti-prostate specific membrane antigen designer T cells for prostate cancer immunotherapy. Prostate. 2014;74(3):286–296. doi: 10.1002/pros.22749. [DOI] [PubMed] [Google Scholar]
- 136.Schmohl JU, Vallera DA. CD133, Selectively targeting the root of cancer. Toxins (Basel). 2016;8(6). doi:10.3390/toxins8060165. [DOI] [PMC free article] [PubMed]
- 137.Wang Z, Wu Z, Liu Y, Han W. New development in CAR-T cell therapy. J Hematol Oncol. 2017;10(1):53. doi: 10.1186/s13045-017-0423-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Feng KC, Guo YL, Liu Y, Dai HR, Wang Y, Lv HY, et al. Cocktail treatment with EGFR-specific and CD133-specific chimeric antigen receptor-modified T cells in a patient with advanced cholangiocarcinoma. J Hematol Oncol. 2017;10(1):4. doi: 10.1186/s13045-016-0378-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Schuberth PC, Hagedorn C, Jensen SM, Gulati P, van den Broek M, Mischo A, et al. Treatment of malignant pleural mesothelioma by fibroblast activation protein-specific re-directed T cells. J Transl Med. 2013;11:187. doi: 10.1186/1479-5876-11-187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Petrausch U, Schuberth PC, Hagedorn C, Soltermann A, Tomaszek S, Stahel R, et al. Re-directed T cells for the treatment of fibroblast activation protein (FAP)-positive malignant pleural mesothelioma (FAPME-1) BMC Cancer. 2012;12:615. doi: 10.1186/1471-2407-12-615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Wang RF, Wang HY. Immune targets and neoantigens for cancer immunotherapy and precision medicine. Cell Res. 2017;27(1):11–37. doi: 10.1038/cr.2016.155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.You F, Jiang L, Zhang B, Lu Q, Zhou Q, Liao X, et al. Phase 1 clinical trial demonstrated that MUC1 positive metastatic seminal vesicle cancer can be effectively eradicated by modified anti-MUC1 chimeric antigen receptor transduced T cells. Sci China Life Sci. 2016;59(4):386–397. doi: 10.1007/s11427-016-5024-7. [DOI] [PubMed] [Google Scholar]
- 143.Kandalaft LE, Powell DJ, Jr, Coukos G. A phase I clinical trial of adoptive transfer of folate receptor-alpha redirected autologous T cells for recurrent ovarian cancer. J Transl Med. 2012;10:157. doi: 10.1186/1479-5876-10-157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Song D-G, Ye Q, Poussin M, Chacon JA, Figini M, Powell DJ. Effective adoptive immunotherapy of triple-negative breast cancer by folate receptor-alpha redirected CAR T cells is influenced by surface antigen expression level. J Hematol Oncol. 2016;9(1):56. doi: 10.1186/s13045-016-0285-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Brown CE, Badie B, Barish ME, Weng L, Ostberg JR, Chang WC, et al. Bioactivity and safety of IL13Ralpha2-redirected chimeric antigen receptor CD8+ T cells in patients with recurrent glioblastoma. Clin Cancer Res. 2015;21(18):4062–4072. doi: 10.1158/1078-0432.CCR-15-0428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Brown CE, Alizadeh D, Starr R, Weng L, Wagner JR, Naranjo A, et al. Regression of glioblastoma after chimeric antigen receptor T-cell therapy. N Engl J Med. 2016;375(26):2561–2569. doi: 10.1056/NEJMoa1610497. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.