

Abstract
Bone marrow mesenchymal stem cells (BMSCs) are a type of adult stem cells with a wide range of potential applications. However, BMSCs have a limited life cycle under normal culturing conditions, which has hindered further study and application. Many studies have confirmed that cells modified by telomerase reverse transcriptase (TERT) can maintain the ability to proliferate in vitro over a long period of time. In this study, we constructed a gene expression vector to transfer TERT into sheep BMSCs, and evaluated whether the TERT cell strain was successfully transferred. The abilities of cell proliferation and differentiation were evaluated using the methods including growth curve determination, inheritance stability analysis, multi-directional induction and so on, and the results showed that the cell strain can be cultured to 40 generations, with a normal karyotype rate maintained at 88.24%, and that the cell strain can be transferred and differentiated into neurocytes and lipocytes, proving that it retains the multi-directional transdifferentiation ability.
Keywords: Sheep BMSCs, Stem cells, TERT, Multi-directional differentiation
1. Introduction
The bone marrow mesenchymal stem cell (BMSC) is a kind of multi-potent adult stem cell originating from the bone marrow stromal, and is a type of adult stem cell with a wide range of potential applications in the fields of tissue engineering, and cell and gene therapy (Augello et al., 2010, Austin-Page et al., 2010, Dai et al., 2014, Machado et al., 2009, Nakahara et al., 2009, Tögel et al., 2009, Yang et al., 2011). In the present study, we found that the ability to proliferate decreases along with the number of in vitro passage cultures in BMSCs, which limits the application of BMSCs to a certain extent (Bonab et al., 2006, Estrada et al., 2013). In recent years, different kinds of immortalized cells have been obtained by different methods, but there is no safe way to obtain immortalized cells.
Telomere is an important structure in maintaining chromosome stability and the life span of cells. Telomere length is inversely proportional to the number of chromosome copies. If the telomere length decreases to an extreme value, it will no longer maintain its function of ensuring chromosome stability, which leads to cell death However, telomere contains a reverse transcriptase known as telomerase reverse transcriptase (TERT), which can catalyze reverse transcription of the telomerase into telomere DNA, which is then synthesized into chromosome ends and added to the length of the telomere, thus resulting in continuous cell growth (Kim et al., 2009). Many studies show that exogenous telomerase reverse transcriptase does not produce canceration, and can maintain stem cell self-renewal and multilineage differentiation potential. Therefore, it is of great theoretical significance to study the effect of TERT on the stable passage and differentiation of MSCs.
Therefore, through introduction of the exogenously expressed TERT gene, we further studied the life cycle and biological characteristics of BMSCs as a basis for further application of mesenchymal stems cells in disease treatment and tissue repair technology.
2. Materials and methods
All chemicals and culture media used in this study were of cell culture grade and obtained from Sigma Chemicals Co., (St. Louis, US) unless otherwise indicated. The plastic ware was from Nunc (Roskilde, Denmark).
2.1. Tissue materials and cell culture
Sheep renal tissue was harvested from 12 month old small-tailed Hen sheep which were provided by a slaughterhouse. BMSCs were provided by the Experimental Center of the College of Animal Science and Technology. Cells were inoculated at a density of 2 × 104 cells/ml in DMEM containing 10% FBS, and cultured at 37 °C in a 5% CO2 humidified incubator after thawing at 37 °C. The culture medium was replaced after 24 h, and every 3 days afterward. When cells had grown to a fusion of 80-90%, subculturing of the cells was performed at a ratio of 1:3 with digestion by 0.25% trypsin.
2.2. Construction of eukaryotic expression vector pcDNA 3.1-EGFP-TERT
Total RNA was extracted from the sheep renal tissue, and reversely transcribed into cDNA which was used as a template. A TERT primer was designed containing the Hind III and EcoR I restriction enzyme cutting site, Fwd: CCCAAGCTTGCCACCATGA AGGTGCAGGACTGCG (Hind III), Rev: CGGAATTCTG TCCAAGATGGTCTTGAAGTCT (EcoR I). PCR amplification conditions: 94 °C, 8 min; 94 °C, 40 s; 56 °C, 30 s; 72 °C, 2 min; 35 cycles. The amplified bands were extracted and sequenced after the reaction was terminated. The recombinant plasmid containing the TERT gene and the plasmid pcDNA3.1-EGFP were cut by Hind III and EcoRI restriction enzymes, respectively, and the enzyme fragments were added into T4 DNA ligase to perform the overnight ligation. Double enzyme cutting and sequence identification of the fragments ligated by Hind III and EcoR I were carried out.
2.3. Liposome transfection and Screening of TERT-BMSCs
0.8 μg of normally sequenced plasmids was mixed with 3 μl of liposome in 100 μl of serum-free DMEM culture medium. Then, the mixture was slowly added into a culture containing 70–80% fused cells after 20 min of incubation at room temperature (RT). After the cells were transfected for 24 h, fluorescence was observed under a fluorescence microscope, and the cells were screened by adding G418 with a final concentration of 300 μg/ml. After 7 d, the G418 concentration was reduced by half and cells continued culturing.
2.4. Determination of growth curve
P5 and P40 TERT-BMSCs as well as BMSCs were selected and inoculated at a concentration of 2 × 104 in 24-well plates. The growth curve was determined by calculating the number of cells in 3 wells per day for 9 consecutive days.
2.5. Inheritance stability analysis
Numerous metaphase cells were selected from P10, P20 and P40 TERT-BMSCs and BMSCs. Then, using BEION chromosome karyotype analysis software, the number of chromosomes was analyzed, and the chromosome number and structural stability of the TERT-BMSCs during subculturing was measured.
2.6. RT-PCR analysis
Total RNA was extracted and reversely transcribed into cDNA for use as a template. A primer for the study gene was designed (Table 1). The targeted band was amplified by PCR, and the amplified band was extracted and sequenced after the reaction was terminated.
Table 1.
Details of primers used for gene expression through RT-PCR.
| Sr. no. | Gene | Primer sequence | Product size (bp) | Annealing temp °C |
Source | Accession no. |
|---|---|---|---|---|---|---|
| 1 | GAPDH | CTTCATTGACCTTCACTACATGG(F) | 356 | 57 | Ovis aries | NM_001190390.1 |
| TGCAGGAGGCATTGCTGACAA(R) | ||||||
| 2 | NSE | AGACCTCATCCTGCCTGTGC(F) | 190 | 58 | Ovis aries | XM_004006922.1 |
| GGCGTCCTTGCCATACTTG(R) | ||||||
| 3 | GFAP | CCGCATCACCATTCCTGTTC(F) | 123 | 60 | Ovis aries | XM_004012992.1 |
| CGCATCTCCACGGTCTTCAC(R) | ||||||
| 4 | PPAR | CGTCAGGGTTCCACTATGGAGTT (F) | 175 | 55 | Ovis aries | NM_001100921.1 |
| GACATCCCCACAGCAAGGCACTT (R) | ||||||
| 5 | Leptin | CCAGGATGACACCAAAACCCTCA (F) | 163 | 57 | Ovis aries | KC526876 |
| GATTGCCAATGTCTGGTCCATCT (R) | ||||||
| 6 | TERT | AGCAGGCGGCTGTAGGTG(F) | 191 | 56 | ||
| GCGTTCTTTCTCCAGGTCATCA(R) |
2.7. Multi-directional induction and differentiation
The P30 TERT-BMSCs were selected and inoculated at 2 × 105 cells/ml in 4-well plates. The culture medium supernatant was discarded and replaced with an adipogenic induction culture medium (DMEM-F12 + 10% FBS + 1 μM of dexamethasone + 17 μM of pantothenic acid + 5 mM of indometacin + 1 μM of insulin + 0.5 mM of IBMX) when cells had grown to a fusion of 70–80%, and the culture medium was replaced every 3 d. Cells were cultured for two weeks. For neuroblast induction, the pre-induction medium (DMEM-F12 + 10%FBS + 1 mM BME) was first added, and was then replaced with induction medium (DMEM-F12 + 5 mM BME) after 12 h of induction. The induction continued for 24 h, and changes were observed under a microscope.
2.8. Identification of induction differentiation
Identification of adipogenic induction: The culture medium was discarded after two weeks of cell induction. Cells were then rinsed three times with PBS, and then rinsed three times with distilled water after 20 min of fixation with 10% formaldehyde, then stained with Oil-Red O for 20 min at RT. The results were observed under a microscope. RT-PCR were used to detect the expression of the specifically expressed gene PPAR and Leptin. Identification of neuroblast induction: The culture medium was discarded after cell induction was terminated. Cells were fixed for 20 min by adding 95% ethanol, and rinsed twice. After staining with toluidine blue dye solution for 40 min at 50–60 °C, and rinsing with distilled water for 2 min, cells were observed under an inverted microscope. Expression of the specifically expressed gene NSE and GFAP were detected by RT-PCR.
2.9. Statistical analysis
Data analysis was performed on SPSS 9.2. The effects of different cryopreservation media on pre-freezing and post-thaw viability of cells were tested by a one-way analysis of variance (ANOVA).
3. Results
3.1. Eukaryotic expression vector pcDNA3.1-EGFP-TERT
Three bands (Fig.1A) which were 5 s, 18 s and 28 s could be clearly observed in the extracted sheep renal tissue RNA by electrophoresis detection. A single band (Fig.1B) with higher specificity and a size comparable to that of the anticipated fragment could be seen after PCR amplification, and the sequencing results were the same as the sequence released in NCBI, which proves that the sheep TERT gene was cloned. The enzyme-cut plasmid pcDNA3.1-EGFP was re-ligated and transfected with TERT fragments, from which the expressing plasmid was obtained, and the TERT gene (Fig.1C) was acquired using PCR amplification. The 6132 bp band of pcDNA3.1-EGFP and the 1873 bp band of TERT (Fig.1D) were acquired by double enzyme cutting.
Figure 1.
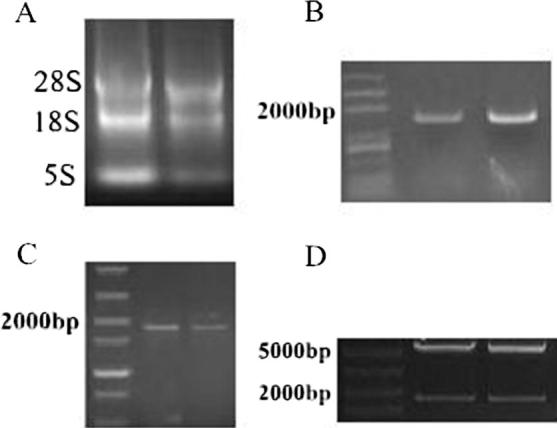
Construction of eukaryotic expression vector pcDNA3.1-EGFP-TERT. (A) The extracted sheep renal tissue RNA under electrophoresis detection; (B) the TERT gene was cloned after PCR amplification; (C) the enzyme-cut plasmid pcDNA3.1-EGFP was re-ligated and transfected with TERT fragments, from which the expressing plasmid was obtained, and the TERT gene (C) was acquired using PCR amplification; (D) The 6132bp band of pcDNA3.1-EGFP and the 1873bp band of TERT (D) were acquired by double enzyme cutting.
3.2. Derivation of TERT-BMSCs
The filtered TERT-BMSCs were subcultured. The cells had a fast growth rate, requiring an average growth period of 3–4 days for each generation. The morphology of the cells was better than that of the BMSCs at higher passages. With the passages increasing, BMSCs gradually grow wider and shorter, eventually taking on a flat polygonal shape, indicating the slow growth caused by cell aging (Fig.2A and B). Meanwhile, TERT-BMSCs maintained their spindle shape, and had no obvious shortening or increase in the number of protuberances, and showed no significant change in growth rate (Fig.2C and D). RT-PCR results show that TERT-BMSCs can express the TERT gene (Fig.2E).
Figure 2.

Derivation of TERT-BMSCs. (A) BMSCs in confluent culture at P3 (×100); (B) BMSCs in confluent culture at P40 (×100); (C) TERT-BMSCs in confluent culture at P3 (×100); (D) BMSCs in confluent culture at P40 (×100). (E) RT-PCR results show that TERT-BMSCs can express the TERT gene, Lane M 2000-bp ladder, lane 1 TERT (191bp), lane 2 negative control.
3.3. Growth curve
The growth curves of TERT-BMSCs and BMSCs from both P5 and P40, take on an “S” shape (Fig. 3), but there is a significant difference between the growth curves of the BMSCs and TERT-BMSCs. The BMSCs ordinarily remain latent for the first 1–2 days, and then enter a logarithmic growth phase on day 3, and a plateau phase on day 7 or 8, with a reduction in the rate of proliferation. While TERT-BMSCs ordinarily begin to grow rapidly from day 2, and enter the plateau phase in advance of day 5–6 due to the growth space constraints. It is to be noted that there is a big difference between the proliferation rates of BMSCs and TERT-BMSCs.
Figure 3.

BMSC and TERT-BMSC growth curves, each value is expressed as mean ± standard error of the mean (SEM).
3.4. The Inheritance stability of TERT-BMSCs
Through karyotype analysis, we found that the normal sheep chromosome karyotype is 2n = 54, which includes 26 pairs of autosomes and 1 chromosome pair. Statistical analysis of the chromosome karyotypes of P5, P20 and P40 TERT-BMSCs and BMSCs showed that the normal karyotype rates of different passages of BMSCs were 96.30%, 72.22% and 31.22%, respectively, and the normal karyotype rates of different passages of TERT-BMSCs were 95.35%, 92.00% and 88.24% (Table 2), respectively. This shows that the TERT-BMSCs maintain excellent inheritance stability over a long period of in vitro passage culturing.
Table 2.
Sheep MSC diploid normal rate of different generations.
| BMSCs (%) | TERT-BMSCs (%) | |
|---|---|---|
| P5 | 96.30% (52/54) | 95.35% (41/43) |
| P20 | 72.22% (26/36) | 92.00% (46/50) |
| P40 | 61.22% (30/49) | 88.24% (45/51) |
3.5. Identification of adipogenic induction
The morphology of TERT-BMSCs begins to change after 24 h of adipogenic induction, gradually changing from the spindle shape to a large ovular shape. Small lipid droplets begin to appear in cytoplasm after 3 days of transfection. Larger lipid droplets appear in some of the cells after 5–6 days of transfection, presenting as a round or ovular shape (Fig.4A). The lipid droplets were stained red using oil-red O dye for observation (Fig.4B) after 9 days of transfection. Meanwhile, no red lipid droplets were observed in the stained control group (Fig.4C). The expression of the specific PPAR and Leptin genes can be detected by RT-PCR.
Figure 4.

Adipogenic (A–C) and Neural (D–F) differentiation potential of sheep TERT-BMSCs. (A) Lipid droplets were seen in the cytoplasm of visible after 9 days of culturing. (B) Oil red O-positive cells. (C) Gene expression profile. Lane 1 250/100bp ladder, lane 2 negative control, lane 3 up: PPAR (175bp); down: Leptin (163bp). (D) Condensed cell bodies and extended dendrites were seen after 24-h culture. (E) Toluidine blue staining, (F) gene expression profile. Lane M 250/100bp ladder, lane 2,3 up: NSE (190bp); down: GFAP (123bp), lane 3,4 negative control.
3.6. Identification of neuroblast induction
No obvious changes were observed in cell morphology after pre-induction of TERT-BMSCs. Enhanced refraction was observed in the BMSC cell bodies, which began to shrink and become rounder 3 h after addition of the induction agent. After 12 h, protuberances began to appear and project out of the cell bodies, causing the cells to form forked ends with large protruding points that can make contact with other cell bodies and points, resembling a synapse structure. The cells became bipolar, multi-polar and tapered, with a morphology like that of neurons after 24 h of induction. At this time, many cells had already intertwined and interconnected with one another with a reticular appearance (Fig.4D). After staining with toluidine blue, nissl bodies appeared as dark blue particles or patches with blue cell nuclei (Fig.4E). RT-PCR was able to detect the expression of the specific NSE and GFAP genes.
4. Discussion
The mesenchymal stem cell (MSC) is a kind of adult stem cell widely applied in tissue repair engineering, and cell and gene therapy. However, MSCs, tend to age and stop proliferating when subcultured in vitro, and there is no way to greatly amplify these cells (Bourgine et al., 2014, Peng et al., 2015, Zimmermann et al., 2003, Røsland et al., 2009, Okada et al., 2016). The present study proves that telomere length may shorten along with cell proliferation. Cells may age and die as the continuously shortened telomere length cannot maintain chromosome stability. Therefore, telomere length is important in guaranteeing cell proliferation stability. Enhancing telomerase activity by introducing the exogenous TERT gene into targeted cells is the primary method used in recent cell immortalization studies (Kaloyianni et al., 2015, Teng et al., 2014, Tsai et al., 2010, Wongkajornsilp et al., 2012). Hamada et al., constructed an hMSC-TERT cell line in 2003, which had biological characteristics that were no different than the original generation of hMSCs, but the detailed molecular mechanism and the function of telomerase remain unclear.
Construction of a eukaryotic expression vector, using the pcDNA3.1-EGFP ring-opening as the expression vector by the restriction enzyme EcoRI and Hind III can ensure the proper insertion direction of exogenous fragments, and can prevent the self-ligation of vectors, which improves recombination efficiency. Furthermore, the expression vector pcDNA3.1-EGFP carries the EGFP gene and the Neo resistance gene, which ensures that it can both express green fluorescence after being introduced into the cell, and can be filtered in eukaryotic cells by G418. In the present study, after constructing a eukaryotic expression vector, we examined the vector from the two dimensions of colony PCR and double enzyme cutting of recombinant plasmids (Zhou et al., 2014). Two types of bands were obtained from the results of double enzyme cutting, with sizes comparable to expectations. One of these was the band of the targeted gene, while the other was the band of the vector, which proves that we successfully constructed the vector.
Cells normally expressing the TERT gene were acquired by G418 filtering after the introduction of the successfully constructed vector into the BMSCs by liposomes (Wongkajornsilp et al., 2012). The acquired cells showed no obvious difference in cell morphology as compared to normal BMSCs when subcultured by amplification to the 40 th passage, while the normal BMSCs showed obvious cell aging and degeneration in cell morphology when cultured to the 20 th passage. These results are analogous to those obtained by (Yao and Hwang, 2012, Yin, 2012). According to the growth curve, the proliferation rate of the BMSCs is obviously decreased when subcultured to the 20th passage, while TERT-BMSCs maintained a normal proliferation rate when subcultured to the 40th passage, the growth curve of this cell maintained the “S” shape, proving that TERT-BMSCs have vigorous proliferation, as reported by (Simonsen et al., 2002). We selected P10, P20 and P40 cells to study the inheritance stability of TERT-BMSCs. Through chromosome karyotype analysis, we found that the cells maintained a karyotype correction rate of 77.78% when subcultured to the 40th passage, which proves that TERT-BMSCs have higher inheritance stability. However, it is necessary to further verify whether TERT-BMSCs are capable of infinite passage culturing.
Adipogenic differentiation assays show that TERT-BMSCs can be differentiated to adipocytes. We find that indomethacin is the most rapid adipogenic supplement, and in 3–4 days of treatment on average, small oil droplets were observed under the inverted microscope. After 7 days of incubation, the cells were stained with Oil-Red O, and red oil vacuoles were obvious in the cytoplasm. The formation of large lipid droplets on the 12th day in adipogenic-induced human MSCs was previously reported. RT-PCR of the differentiated cells shows the adipogenic differentiation-specific genes such as PPAR and Leptin were expressed.
Our neural differentiation assays showed that TERT-BMSCs can differentiate to neurons. BME can support the viability and differentiation of fetal mouse brain neurons (Fortino et al., 2013) and is used as an effective inducer of neural differentiation in MSCs (Latil et al., 2012, Sanchez-Ramos et al., 2000). BME induced dramatic modifications of cellular shape and the expression of neural marker NeuN within 5 h. Nestin expression is a necessary step for neural differentiation of MSCs, and serum in culture medium can inhibit the expression of Netein. We found that TERT-BMSCs also could differentiate into neural cells under serum-free conditions. RT-PCR results confirmed that the specific genes for neural differentiation such as ESE and GFAP were expressed.
5. Conclusions
In the present study, the TERT eukaryotic expression vector was successfully constructed and BMSCs were transfected. Observation of cell morphology and detection of the biological characteristics of BMSCs showed that no early aging occurred, while the stem cell characteristics of the cells were maintained and their life spans prolonged. TERT-BMSCs maintain the potential for multi-directional differentiation after induction. These results can be referenced in the future research of cell immortalization, helping further the discussion of the immortalization mechanism, and laying a foundation for applying the immortalization mechanism in the fields of tissue regeneration and repair, cell transplantation, and gene therapy, etc.
Acknowledgment
This research was supported by Chinese National Natural Science Foundation (grant number: 31402153) and PhD Start-up Fund of College of Animal Science and Technology (13480062).
Footnotes
Peer review under responsibility of King Saud University.

References
- Augello A., Kurth T.B., De Bari C. Mesenchymal stem cells: a perspective from in vitro cultures to in vivo migration and niches. Eur. Cell Mater. 2010;20:121–133. doi: 10.22203/ecm.v020a11. [DOI] [PubMed] [Google Scholar]
- Austin-Page L., Thein S., Lam M., Park A., Li N., Yang R., Gorlick R. Introducing different genetic elements into human mesenchymal stem cells to transform into osteosarcoma. Cancer Res. 2010;70:418. [Google Scholar]
- Bonab M.M., Alimoghaddam K., Talebian F., Ghaffari S.H., Ghavamzadeh A., Nikbin B. Aging of mesenchymal stem cell in vitro. BMC Cell Biol. 2006;71:1. doi: 10.1186/1471-2121-7-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourgine P., LeMagnen C., Pigeot S., Geurts J., Scherberich A., Martin I. Combination of immortalize- ation and inducible death strategies to generate a human mesenchymal stromal cell line with controlled survival. Stem Cell Res. 2014;12:584–598. doi: 10.1016/j.scr.2013.12.006. [DOI] [PubMed] [Google Scholar]
- Dai F., Yang S., Zhang F., Shi D., Zhang Z., Wu J., Xu J. HTERT-and hCTLA4Ig-expressing human bone marrow-derived mesenchymal stem cells: in vitro and in vivo characterization and osteogenic differentiation. J. Tissue Eng. Regener. Med. 2014 doi: 10.1002/term.1924. [DOI] [PubMed] [Google Scholar]
- Estrada J.C., Torres Y., Benguría A., Dopazo A., Roche E., Carrera-Quintanar L., Samper E. Human mesenchymal stem cell-replicative senescence and oxidative stress are closely linked to aneuploidy. Cell Death Dis. 2013;4:e691. doi: 10.1038/cddis.2013.211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fortino V.R., Pelaez D., Cheung H.S. Concise review: stem cell therapies for neuropathic pain. Stem Cells Transl. Med. 2013;2:394–399. doi: 10.5966/sctm.2012-0122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaloyianni M., Pouikli A., Kyrka L., Petrakis S., Koliakos G., Trachana V. Telomerase overexpression in human mesenchymal stem cells offers protection against oxidative DNA damage accumulation. Cytotherapy. 2015;17:S34–S35. [Google Scholar]
- Kim J., Kang J.W., Park J.H., Choi Y., Choi K.S., Park K.D., Kim H.S. Biological characterization of long-term cultured human mesenchymal stem cells. Arch. Pharm. Res. 2009;32:117–126. doi: 10.1007/s12272-009-1125-1. [DOI] [PubMed] [Google Scholar]
- Latil M., Rocheteau P., Châtre L., Sanulli S., Mémet S., Ricchetti M., Chrétien F. Skeletal muscle stem cells adopt a dormant cell state post mortem and retain regenerative capacity. Nat. Commun. 2012;3:903. doi: 10.1038/ncomms1890. [DOI] [PubMed] [Google Scholar]
- Machado C.B., Correa C.R., Medrado G.C., Leite M.F., Goes A.M. Ectopic expression of telomerase enhances osteopontin and osteocalcin expression during osteogenic differentiation of human mesenchymal stem cells from elder donors. J. Stem Cells Regener. Med. 2009;5:49. doi: 10.46582/jsrm.0502010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakahara H., Misawa H., Hayashi T., Kondo E., Yuasa T., Kubota Y., Javed S.M. Bone repair by transplantation of hTERT-immortalized human mesenchymal stem cells in mice. Transplantation. 2009;88:346–353. doi: 10.1097/TP.0b013e3181ae5ba2. [DOI] [PubMed] [Google Scholar]
- Okada M., Kim H.W., Matsu-ura K., Wang Y.G., Xu M., Ashraf M. Abrogation of age-induced MicroRNA-195 rejuvenates the senescent mesenchymal stem cells by reactivating telomerase. Stem Cells. 2016;34:148–159. doi: 10.1002/stem.2211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peng W.X., Wang L.S., Li D.L., Shi L.W., Tan Y.P. Molecular characteristics of pharmacology woody extracts of eucalyptus camaldulensis biomass. Wood Res. 2015;60(6):891–898. [Google Scholar]
- Røsland G.V., Svendsen A., Torsvik A., Sobala E., McCormack E., Immervoll H., Bjerkvig R. Long-term cultures of bone marrow–derived human mesenchymal stem cells frequently undergo spontaneous malignant transformation. Cancer Res. 2009;69:5331–5339. doi: 10.1158/0008-5472.CAN-08-4630. [DOI] [PubMed] [Google Scholar]
- Sanchez-Ramos J., Song S., Cardozo-Pelaez F., Hazzi C., Stedeford T., Willing A., Cooper D.R. Adult bone marrow stromal cells differentiate into neural cells in vitro. Exp. Neurol. 2000;164:247–256. doi: 10.1006/exnr.2000.7389. [DOI] [PubMed] [Google Scholar]
- Simonsen J.L., Rosada C., Serakinci N., Justesen J., Stenderup K., Rattan S.I., Kassem M. Telomerase expression extends the proliferative life-span and maintains the osteogenic potential of human bone marrow stromal cells. Nat. Biotechnol. 2002;20:592–596. doi: 10.1038/nbt0602-592. [DOI] [PubMed] [Google Scholar]
- Teng Y., Hu Y., Li X.S., Wang Z., Wang R. Telomerase reverse transcriptase mediated immortalization of human bone marrow stromal cells. Braz. Arch. Biol. Technol. 2014;57:37–44. [Google Scholar]
- Tögel F., Cohen A., Zhang P., Yang Y., Hu Z., Westenfelder C. Autologous and allogeneic marrow stromal cells are safe and effective for the treatment of acute kidney injury. Stem Cells Dev. 2009;18:475–486. doi: 10.1089/scd.2008.0092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsai C.C., Chen C.L., Liu H.C., Lee Y.T., Wang H.W., Hou L.T., Hung S.C. Overexpression of hTERT increases stem-like properties and decreases spontaneous differentiation in human mesenchymal stem cell lines. J. Biomed. Sci. 2010;17:1. doi: 10.1186/1423-0127-17-64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wongkajornsilp A., Sa-ngiamsuntorn K., Hongeng S. Development of immortalized hepatocyte-like cells from hMSCs. Liver Stem Cells: Methods Protocols. 2012:73–87. doi: 10.1007/978-1-61779-468-1_7. [DOI] [PubMed] [Google Scholar]
- Yang C.Y., Hsiao J.K., Tai M.F., Chen S.T., Cheng H.Y., Wang J.L., Liu H.M. Direct labeling of hMSC with SPIO: the long-term influence on toxicity, chondrogenic differentiation capacity, and intracellular distribution. Mol. Imaging Biol. 2011;13:443–451. doi: 10.1007/s11307-010-0360-7. [DOI] [PubMed] [Google Scholar]
- Yao C.L., Hwang S.M. Immortalization of human mesenchymal stromal cells with telomerase and red fluorescence protein expression. Somatic Stem Cells: Methods Protocols. 2012:471–478. doi: 10.1007/978-1-61779-815-3_28. [DOI] [PubMed] [Google Scholar]
- Yin Z. lmu; 2012. Establishment of a clonal immortalized human mesenchymal stem cell line expressing hTERT using lentiviral gene transfer. (Doctoral dissertation) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou J., Xi P., Zhou Q., Ding D., Cong Y. The putative tumor suppressor C53 interacts with the human telomerase reverse transcriptase hTERT and regulates telomerase activity. Chin. Sci. Bull. 2014;59:2324–2330. [Google Scholar]
- Zimmermann S., Voss M., Kaiser S., Kapp U., Waller C.F., Martens U.M. Lack of telomerase activity in human mesenchymal stem cells. Leukemia. 2003;17:1146–1149. doi: 10.1038/sj.leu.2402962. [DOI] [PubMed] [Google Scholar]