

Abstract
Background
Filarial nematodes currently infect up to 54 million people worldwide, with millions more at risk for infection, representing the leading cause of disability in the developing world. Brugia malayi is one of the causative agents of lymphatic filariasis and remains the only human filarial parasite that can be maintained in small laboratory animals. Many filarial nematode species, including B. malayi, carry an obligate endosymbiont, the alpha-proteobacteria Wolbachia, which can be eliminated through antibiotic treatment. Elimination of the endosymbiont interferes with development, reproduction, and survival of the worms within the mamalian host, a clear indicator that the Wolbachia are crucial for survival of the parasite. Little is understood about the mechanism underlying this symbiosis.
Methodology/ Principle findings
To better understand the molecular interplay between these two organisms we profiled the transcriptomes of B. malayi and Wolbachia by dual RNA-seq across the life cycle of the parasite. This helped identify functional pathways involved in this essential symbiotic relationship provided by the co-expression of nematode and bacterial genes. We have identified significant stage-specific and gender-specific differential expression in Wolbachia during the nematode’s development. For example, during female worm development we find that Wolbachia upregulate genes involved in ATP production and purine biosynthesis, as well as genes involved in the oxidative stress response.
Conclusions/ Significance
This global transcriptional analysis has highlighted specific pathways to which both Wolbachia and B. malayi contribute concurrently over the life cycle of the parasite, paving the way for the development of novel intervention strategies.
Author summary
Filarial nematodes currently infect millions of people worldwide and represent a leading cause of disability. Currently available medications are insufficient in reaching elimination of these parasites. Many filarial nematodes, including Brugia malayi, have an Achilles heel of sorts—that is their obligate symbiotic relationship with the bacteria Wolbachia. While it is known that the nematode and the bacteria are co-dependent, the molecular basis of this relationship remains poorly understood. Using deep sequencing, we profiled the transcriptomes of B. malayi and Wolbachia across the life cycle of the parasite to determine the functional pathways necessary for parasite survival provided by the co-expression of nematode and bacterial genes. Defining the mechanisms of endosymbiosis between these two organisms will allow for the exploitation of this relationship for the development of new intervention strategies.
Introduction
Human filarial infections are currently a leading cause of morbidity in the developing world. Despite the large cost to human health, the chronic and debilitating diseases caused by filarial nematodes remain largely neglected. Two of the most prevalent chronic diseases caused by filaria include lymphatic filariasis, caused by Wuchereria bancrofti, Brugia malayi, and Brugia timori, and onchocerciasis, caused by Onchocerca volvulus [1]. Currently 38.5 million people have lymphatic filariasis while 15.5 million people have onchocerciasis, representing in 2015 over 300,000 years lived with disability (YLDs) [2]. While efforts to mitigate the effects of these diseases have been successful in some regions, current medications are insufficient to reach elimination by 2020, particularly in regions of co-endemicity with loasis, caused by the filarial nematode Loa loa [3]. Current mass drug administration relies on a small arsenal of drugs, increasing the likelihood of development of resistance, a phenomenon already observed in their veterinary applications [4]. One such drug, Ivermectin, the primary control strategy for onchocerciasis, is unsafe to use in regions where lymphatic filariasis or onchocerciasis are co-endemic with loasis due to the risk of severe adverse effects in individuals heavily infected with Loa loa.
Most filarial nematodes are hosts for an obligate bacterial endosymbiont, the intracellular bacteria of the genus Wolbachia. As the filariae require these bacteria to develop, reproduce and survive in the human host, they represent an attractive target for intervention. The bacteria reside in the lateral cords of the larval and adult nematodes (male and female) as well as in the ovaries and developing embryos of the adult female worms. While the relationship between the nematode and the bacteria is known to be co-dependent, the molecular basis for this relationship remains poorly understood. Wolbachia are required for the parasite to reproduce and develop in the mammalian host, while the parasite likely provides amino acids required for bacterial growth [5]. Analyses show significant degradation of the Wolbachia genome compared to its free-living relatives, yet it appears to have maintained a number of intact metabolic pathways such as riboflavin, heme, and nucleotide synthesis [5, 6], three pathways that are deficient in the nematode host [7]. As these metabolites are considered essential to all living things, these deficiencies may underlie the symbiotic relationship. Interestingly, in a genome-wide screen for diversifying selection, genes for heme, riboflavin, and nucleotide biosynthesis were found to be under positive selection, again implying they may be integral to the symbiotic relationship [8]. Curiously, however, the recently sequenced L. loa genome, a Wolbachia-free filarial nematode believed to have lost the endosymbiont, also lacks these metabolic pathways and does not appear to have acquired them through horizontal gene transfer [9, 10]. This suggests that filarial worms could also be acquiring these essential metabolites from their mammalian hosts. Thus, the basis of the filaria-Wolbachia co-dependency has still not been clarified with the availability of the genomes.
Clearance of Wolbachia with the use of antibiotics results in significant apoptosis of filarial germline cells, cells of developing embryos in the female worms, as well as somatic cells of the microfilaria. These effects are non cell-autonomous, meaning cell death is not restricted to cells infected with Wolbachia pre-treatment [9]. It is hypothesized that Wolbachia are preventing apoptosis by one or both of two possible mechanisms: i) Wolbachia are interfering with the host apoptotic program to prevent cell death, and ii) Wolbachia secrete some necessary metabolic product(s) that prevent cell death. In this study, we profiled the transcriptomes and inferred co-expression of genes in Wolbachia and B. malayi during the development of male and female worms to identify co-expressed pathways necessary for mediating the endosymbiotic relationship.
Materials and methods
Parasites and study design
Parasites were obtained from FR3 where they were isolated and separated by sex from infected gerbils (Meriones unguiculatus) at 16 (L4), 30, 42 and 120 days post infection (dpi). Worms were flash frozen and shipped to the New York Blood Center for processing.
RNA isolation, library preparation and sequencing
B. malayi worms where homogenized in Trizol (ThermoFisher) using a hand-held pestle in 1.5mL tubes containing the worms. For extraction, 2,000 L4s, 50 male and female juveniles (at 30 dpi and 42 dpi), and 10 male and female adult worms (120 dpi) were used, with two biological replicates for each. Total RNA was extracted by organic extraction using Trizol. A portion of each sample was saved for a DNA extraction while the rest was treated with DNaseI (New England Biolabs). Ribosomal RNA (rRNA) depletion was performed using Terminator (Epicentre), a 5’-phosphate-dependent exonuclease that degrades transcripts with a 5’ monophosphate. Libraries were prepared using the NEBNext Ultra RNA Library Prep Kit for Illumina (New England Biolabs) according to manufacturer instructions. Library quality was assessed using a D1000 ScreenTape Assay (Aligent) prior to sequencing. Library concentrations were assessed using the qPCR library quantification protocol (KAPA biosystems). Libraries were sequenced on the Illumina HiSeq2500 platform with 150bp paired-end reads. To minimize the confounding effects of lane-to-lane variation, libraries were multiplexed and sequenced with technical replicates on multiple lanes. Each developmental stage received an average of 141 million mapped reads.
Sequencing alignment and differential expression analysis
Read quality was assessed using FastQC (Babraham Bioinformatics). Sequence reads from each sample were demultiplexed and analyzed with the Tuxedo suite of tools [11–13]. Reads were mapped to the annotated B. malayi (WormBase.org) and Wolbachia [6] genome assemblies with Tophat2’s (v2.1.1) Bowtie2-very-sensitive algorithm [11]. The resulting BAM files were then used with Cufflinks (v2.2.1) [11–13] to obtain fragments per kilobase of exon per million fragments mapped (FPKMs) for each of the annotated transcripts and with Cuffnorm [11–13] to obtain normalized FPKMs, normalized for library size. The Tophat2 alignment files were also used to determine differentially expressed genes in both organisms by first using HTSeq (v0.6.1p2) [14] to generate read counts for each gene. Raw read counts were used as input to EdgeR (v3.16.5) [15] to obtain differentially expressed genes between life stage. Genes were determined as significantly differentially expressed using a threshold of p <0.05 and a false-discovery rate (FDR) of 5%, standard settings in EdgeR.
Co-expression analysis
To make the co-expression network and identify the co-expressed gene modules in the symbiosis between B. malayi and Wolbachia, we normalized the gene expression profiles of B. malayi and Wolbachia using Cuffnorm [11–13] and then performed weighted gene correlation network analysis (WGCNA) on the combination of normalized gene expression of B. malayi and Wolbachia using the WGCNA package in R [16]. Hierarchical clustering and dynamic branch cutting were used to identify stable modules of densely interconnected genes. GO term information was downloaded from WormBase.org. Metadata including WSP (Wolbachia Surface Protein) and a ratio of wsp to gst (glutathione-S-transferase) were all integrated into the co-expression network.
cDNA synthesis and RT-qPCR
To estimate the relative expression of Wolbachia genes over different stages of worm development, the DNAse-treated RNA stored in aliquots that were prepared for library preparation and sequencing (see above) was used as a template for cDNA synthesis using the SuperScript III First Strand cDNA Synthesis Kit (Invitrogen). The cDNA was prepared from two biological replicates.
Gene expression was estimated using the standard ‘ΔΔCt’ method. For internal control of Wolbachia gene expression, we selected two housekeeping genes (wBm0291 and wBm0528) based on their constitutive expression over the development of the worm according to the RNA-seq data.
DNA isolation and qPCR
DNA was extracted from B. malayi worms (the same samples as RNA) by taking the non-organic fraction of trizol/chloroform solutions (see above). DNA was precipitated by ethanol and diluted in water. Wolbachia numbers per worm were quantified by qPCR using primers for a Wolbachia single-copy gene (wsp) as previously described [17].
Accession numbers
Expression data have been deposited in the Sequence Read Archive (SRA) under Accession number SRP090644.
Results
Transcriptome overview
To obtain a global view of the transcriptional programs of both B. malayi and Wolbachia concurrently, over the course of worm development from L4 to adulthood, we performed dual RNA-seq. In total over 988 million (or 486 paired-end) RNA-seq reads out of 1.5 billion reads (65.9%) obtained were mapped to the B. malayi and Wolbachia reference genomes (Fig 1: Circos plots for Wolbachia). Mapped reads per stage ranged from 77 to 216 million for the B. malayi genome and 2.1 to 3.7 million for the Wolbachia genome (Table 1: Sequencing summary). We found over 96% of B. malayi gene models to be “expressed” in at least one stage (i.e. a minimum of four cumulative reads across the two independent biological replicates) (Fig 2: Clustering of Brugia Expression) (Table 1: Sequencing Summary). Expression in B. malayi was dominated by sex-biased gene expression, with the 120 dpi adult male and females expressing the most genes at the highest expression levels (Fig 2). In Wolbachia, 85% of gene models were classified as expressed (Table 1: Sequencing Summary). Sequence reads from technical replicates—i.e. the same library sequenced on different lanes of the HiSeq—were combined per biological replicate as they contained the same insert size distribution. Using multidimensional scaling analysis, we clustered biological replicates for each B. malayi developmental stage (S1a Fig: B. malayi MDS). All biological replicates clustered closely to each other with the exception of the F42 replicates, where F42b clustered more closely with the F30 replicates than with F42a. However, when we clustered biological replicates for Wolbachia reads, F42a and F42b clustered closely together (S1b Fig: Wolbachia MDS). It is thus unlikely that the observed B. malayi disparate clustering for that stage is due to the mislabeling or contamination of the sample and more an effect of natural population variation. Generally, clustering of the stages indicates good reproducibility of the biological replicates, with the 30 dpi samples of both males and females clustering more closely with the mixed-gender L4s than to each other, followed by the 42 dpi samples. As expected, the 120 dpi male and female samples are found to be the most different from each other than the other samples from earlier life stages.
Fig 1. Circos plots of stage-specific Wolbachia gene expression.

Wolbachia expression profiles using normalized FPKMs over development of male and female worms from L4 to 120 days post infection (dpi); genomic location is shown around the perimeter, and the black rectangles represent the CDS.
Table 1. Dual RNA-seq sequencing summary.
The table shows the total reads sequenced and mapped in each biological replicate at each developmental stage, L4 to 120 days post infection (dpi) males (M) and females (F), lower case a and b refer to separate biological replicates.
| Sample | Total reads (million) | Left mapped reads (million) | Right mapped reads (million) | Total mapped reads (million) | % Mapped reads | Stage total reads mapped to Brugia (million) | % Brugia Genes Expressed | Stage total reads mapped to Wolbachia (million) | % Wolbachia genes expressed |
|---|---|---|---|---|---|---|---|---|---|
| L4a | 104 | 24 | 23 | 47 | 45 | 94 | 69 | 2 | 66 |
| L4b | 91 | 25 | 24 | 49 | 54 | ||||
| F30a | 82 | 20 | 19 | 39 | 48 | 87 | 75 | 3 | 61 |
| F30b | 101 | 25 | 25 | 50 | 50 | ||||
| F42a | 172 | 75 | 73 | 149 | 87 | 185 | 78 | 3 | 60 |
| F42b | 74 | 20 | 19 | 39 | 53 | ||||
| F120a | 195 | 85 | 81 | 166 | 85 | 212 | 94 | 4 | 84 |
| F120b | 105 | 25 | 25 | 50 | 48 | ||||
| M30a | 57 | 16 | 16 | 32 | 56 | 75 | 78 | 2 | 62 |
| M30b | 83 | 23 | 23 | 46 | 55 | ||||
| M42a | 158 | 71 | 68 | 139 | 88 | 206 | 82 | 3 | 62 |
| M42b | 119 | 35 | 35 | 70 | 59 | ||||
| M120a | 90 | 40 | 39 | 78 | 87 | 110 | 91 | 3 | 65 |
| M120b | 56 | 17 | 17 | 34 | 61 | ||||
| Total | 1486 | 502 | 487 | 989 | 969 | 97 | 19 | 85 |
Fig 2. Clustering of stages based on B. malayi gene expression.

Clustering of B. malayi genes and developmental stages, L4 to 120 days post infection (dpi) Male (M) and Female (F), based on gene expression in normalized FPKMs. Expression was scaled using Z score prior to clustering, with red representing high expression and blue representing low expression. Biological replicates were combined prior to analysis.
To validate the use of RNA-seq for the purpose of transcriptional analysis, seven Wolbachia genes, with ten pair-wise comparisons, were selected for qRT-PCR analysis of their relative expression. Four of the genes (wsp, Hsp90, DnaK, and GroEL), with seven significant pair-wise comparisons, were chosen based on the criteria that they were found to be significantly differentially expressed and had over 50 read counts per stage. We also included three genes (RibA, HemA, and AfuA) that were constitutively expressed, based on an FDR of 1 in EdgeR, which indicates that they were the least likely to be differentially expressed. We observed a spearman correlation between the qRT-PCR and RNA-seq results of 0.987 and a p-value < 2.2e-16 (S2 Fig: qPCR validation graph).
Differential expression is dominated by female stages
Differential expression in B. malayi was dominated by sex-biased gene expression, as previously observed [18], with the largest number of sex-biased genes at 120 dpi, with 2,753 genes showing male bias, and 3,109 showing female biased expression (S1 Table: Brugia DE Female, S2 Table: Brugia DE Male, and S3 Table: Brugia DE Male to Female). We find that 82% of the genes previously determined to be significantly up-regulated in adult male worms and 79% of the genes significantly up-regulated in females worms [18], were, in our new data set, also up-regulated in male worms (M120) or female worms (F120), respectively, as compared to worms of the opposite sex. This shows good reproducibility between the two studies, although it should be noted that many additional genes were found to be differentially expressed between F120 and M120 worms in the new data set. This is likely due to the use of biological and technical replicates, as well as a higher depth of coverage.
To uncover the role Wolbachia may play in worm development, we analyzed differentially expressed Wolbachia genes in male and female worms at each developmental stage. Pair-wise differential expression analysis was performed using EdgeR, after removing all genes with zero expression in two or more samples per comparison. The percentage of differentially expressed genes in any pair-wise comparison ranged from 0–4.8% of Wolbachia genes expressed (Table 1).
We identified a total of 62 differentially expressed (DE) Wolbachia genes across a single or multiple pair-wise comparisons (Fig 3: Clustering of Wolbachia DE genes). The largest number of Wolbachia DE genes (34 genes) is in the females from 42 dpi (F42) to 120 dpi (F120), while there are no DE genes in the 30 to 42dpi comparisons in both males and females (Table 2: DE summary). In comparing stages between genders, there were 17 DE genes between females and males at 120 dpi (F120 and M120), and no genes differentially expressed between both sexes at 30 or 42 dpi. Comparisons between female stages consistently resulted in more DE genes than did comparisons between male stages (Table 2: DE summary): 40 Wolbachia genes were determined as DE over the course of female growth, but were absent in any male comparisons. Because these genes appear to be differentially regulated during female worm development only, they are potentially required for female-specific processes known to be dependent on Wolbachia infection, such as maturity of female gonads and germline development, as well as embryogenesis. The ten Wolbachia genes determined as DE in both males and females represent potential expression in the lateral cords, required for the development of both the male and female germlines. Twelve Wolbachia genes were determined as DE in males only (S4 Table: Wolbachia DE genes).
Fig 3. Clustering of stages and Wolbachia DE genes.

Hierarchical clustering of Wolbachia DE genes and developmental stages, L4 through 120 days post infection (dpi) male (M) and female (F), based on gene expression in normalized FPKMs. Expression values were scaled prior to clustering using a Z score calculation, with red representing high expression and blue representing low expression. Biological replicates were combined prior to clustering.
Table 2. Summary of differential expression.
The table shows the results of the EdgeR pair-wise comparisons between developmental stages, L4 to 120 days post infection (dpi) in males (M) and females (F). Genes are said to be up-regulated if they are higher in the second developmental stage in the comparison, and down-regulated if they are lower in the second developmental stage.
| Stage | L4-F30 | F30-F42 | F42-F120 | F30-F120 | L4-M30 | M30-M42 | M42-M120 | M30-M120 | F30-M30 | F42-M42 | F120-M120 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Wolbachia Genes | 682 | 673 | 728 | 728 | 682 | 681 | 689 | 691 | 673 | 678 | 734 |
| Total DE Wolbachia | 7 (1.0%) | 0 | 34 (4.7%) | 27 (3.7%) | 3 (0.4%) | 0 | 7 (1.0%) | 4 (0.6%) | 0 | 0 | 17 (2.3%) |
| Wolbachia Up | 1 | 0 | 17 | 15 | 0 | 0 | 3 | 3 | 0 | 0 | 12 |
| Wolbachia Down | 6 | 0 | 17 | 12 | 3 | 0 | 4 | 1 | 0 | 0 | 5 |
| Brugia Genes | 9,269 | 9,431 | 10,424 | 10,295 | 9,384 | 9,929 | 10,386 | 10,356 | 9,517 | 9,892 | 10,643 |
| Total DE Brugia | 1,129 (12.2%) | 13 (0.1%) | 1,350 (13.0%) | 4,113 (40.0%) | 930 (9.9%) | 2,036 (20.5%) | 3,101 (29.9%) | 3,580 (34.6%) | 1,243 (13.1%) | 456 (4.6%) | 5,281 (49.6%) |
| Brugia Up | 474 | 12 | 805 | 1,995 | 538 | 1,118 | 1,398 | 1,535 | 697 | 265 | 2,817 |
| Brugia Down | 655 | 1 | 545 | 2,118 | 392 | 918 | 1,703 | 2,045 | 546 | 191 | 2,464 |
Up-regulated Wolbachia genes during female development are enriched for chaperone function, energy production and translation
The pattern of Wolbachia differential expression in the female stages was dominated by chaperone protein expression (Table 3: DE expression of Chaperones). Integral membrane proteins, translation, antioxidants, oxidative phosphorylation, DNA replication, and peptidase function were also highly represented (S4 Table). GO-term enrichment analysis of Wolbachia DE genes during female development using Fisher’s exact test revealed an enrichment of GO terms associated with chaperone function including protein-folding and unfolded protein-binding (Table 3). We detected seven genes with chaperone function that were significantly up-regulated in F30 as well as in F42, four of which are also significantly up-regulated in F120 as compared to M120. The four genes found to be significantly up-regulated in all female stages include wBm0350 groEL and co-chaperonin wBm0349 groES, which work in complex as an integral part of several stress responses in bacteria, including the oxidative stress response, where they recover oxidized proteins [19–23]. Also highly up-regulated in all female stages is the molecular chaperone wBm0533 grpE, shown to assist in protein refolding during oxidative stress, as well as HslU, a subunit of an ATP-dependent protease with chaperone function [24, 25]. Among the chaperones significantly up-regulated in the F30 and F42 stages, but not F120, are DnaK and DnaJ, another molecular chaperone system shown to be required for cell division in bacteria as well as for resistance to heat shock [23].
Table 3. DE expression of chaperones.
The table shows the differential expression of genes encoding chaperone proteins over the development of the parasite, L4 to 120 days post infection (dpi) males (M) and females (F). The stage at which the gene is up-regulated is listed, and the stage at which it was compared to is in parentheses.
| Gene | Differential Expression | Description | logFC | logCPM | PValue | FDR |
|---|---|---|---|---|---|---|
| Wbm0138 | Upregulated in F30 (F120) | Heat shock protein 90 | -1.30 | 6.72 | 8.25E-06 | 6.01E-04 |
| Wbm0350 | Upregulated in F30 (F120) | Molecular chaperone GroEL | -1.48 | 9.79 | 4.90E-16 | 1.78E-13 |
| Wbm0785 | Upregulated in F30 (F120) | Molecular chaperone DnaJ | -1.51 | 6.38 | 3.13E-06 | 3.20E-04 |
| Wbm0533 | Upregulated in F30 (F120) | Molecular chaperone GrpE (heat shock protein) | -1.85 | 4.85 | 4.06E-04 | 1.65E-02 |
| Wbm0495 | Upregulated in F30 (F120) | Molecular chaperone DnaK | -1.87 | 9.10 | 1.09E-13 | 2.65E-11 |
| Wbm0349 | Upregulated in F30 (F120) | Co-chaperonin GroES | -1.95 | 7.55 | 2.83E-11 | 5.14E-09 |
| Wbm0723 | Upregulated in F30 (F120) | ATP-dependent protease ATP-binding subunit HslU | -1.99 | 6.41 | 8.21E-08 | 1.19E-05 |
| Wbm0349 | Upregulated in F42 (F120) | Co-chaperonin GroES | -1.72 | 7.60 | 2.07E-05 | 1.51E-03 |
| Wbm0785 | Upregulated in F42 (F120) | Molecular chaperone DnaJ | -1.94 | 6.32 | 1.60E-07 | 1.66E-05 |
| Wbm0495 | Upregulated in F42 (F120) | Molecular chaperone DnaK | -2.13 | 9.06 | 2.42E-13 | 8.81E-11 |
| Wbm0138 | Upregulated in F42 (F120) | Heat shock protein 90 | -2.14 | 6.57 | 1.42E-10 | 3.46E-08 |
| Wbm0533 | Upregulated in F42 (F120) | Molecular chaperone GrpE (heat shock protein) | -2.20 | 4.82 | 7.75E-05 | 4.34E-03 |
| Wbm0350 | Upregulated in F42 (F120) | Molecular chaperone GroEL | -2.34 | 9.63 | 1.74E-19 | 1.27E-16 |
| Wbm0723 | Upregulated in F42 (F120) | ATP-dependent protease ATP-binding subunit HslU | -2.39 | 6.37 | 2.35E-09 | 3.43E-07 |
| Wbm0350 | Upregulated in F120 (M120) | Molecular chaperone GroEL | -1.75 | 9.92 | 1.80E-04 | 1.62E-02 |
| Wbm0723 | Upregulated in F120 (M120) | ATP-dependent protease ATP-binding subunit HslU | -1.88 | 6.61 | 9.17E-04 | 3.99E-02 |
| Wbm0533 | Upregulated in F120 (M120) | Molecular chaperone GrpE (heat shock protein) | -2.09 | 4.97 | 7.59E-04 | 3.98E-02 |
| Wbm0349 | Upregulated in F120 (M120) | Co-chaperonin GroES | -2.50 | 7.64 | 1.63E-07 | 1.19E-04 |
Among the Wolbachia DE genes in adult female worms (F120) are a number of genes involved in combating oxidative stress. wBm0439 coenzyme Q-binding protein, an antioxidant, is significantly up-regulated from F30 to F120 as well as from F42 to F120. We also find wBm0220 SodA, a superoxide dismutase, to be significantly up-regulated from F42 to F120. SodA catalyzes the conversion of superoxide radicals to hydrogen peroxide and oxygen and is known to be essential in combating oxidative stress [19]. Additionally, we detect significant upregulation from F42 to F120 of wBm0674, a malic enzyme responsible for the interconversion of L-malate and pyruvate. This reaction is essential for maintaining cellular pools of NADPH, required for a number of downstream processes including reducing oxidative stress [26, 27].
A number of genes determined as DE during female development are involved in energy production. It is hypothesized that a key mechanism of the Wolbachia-host symbiosis is aerobic energy production by the bacteria for the worm [28–30]. NADH dehydrogenase subunit B (wBm0242), which is involved in oxidative phosphorylation, is significantly up-regulated in F30 as compared to L4s. ATP synthase subunit C, which creates ATP using a proton gradient, is also up-regulated in F120 as compared to F30, and in M120 as compared to F120. Two proteins involved in iron-sulfur cluster formation are also up-regulated in F120 as compared to earlier female stages: wBm0756, an iron-sulfur cluster assembly scaffold protein, and wBm0448, a succinate dehydrogenase flavoprotein. Iron-sulfur clusters are essential co-factors for respiratory chain proteins involved in ATP production [28].
Several glycolytic enzymes were significantly up-regulated in F120 and M120 as compared to earlier stages, including transaldolase (wBm0686), an enzyme linking the pentose phosphate pathway to glycolysis, and wBm0097, a fructose-bisphophase aldolase. Notably, the Wolbachia genome lacks two glycolytic enzymes (6-phosphofructokinase and pyruvate kinase) likely rendering the glycolytic pathway defective. Wolbachia may therefore depend on products from the B. malayi glycolytic cycle such as pyruvate, as well as TCA cycle intermediates derived from amino acids. Accordingly, wBm0207 pyruvate dehydrogenase, which transforms pyruvate into acetyl-CoA that can then be used in the citric acid cycle, was differentially expressed during female development. Correspondingly, in B. malayi we see an upregulation of Bm5241 in F120, involved in the glycogen catabolic process. We also see differential expression of wBm0384, an extracellular metalloprotease potentially involved in the breakdown of filarial peptides for amino acids during female development [6]. Additionally we see the two Zn-dependent peptidases cluster with most of the TCA cycle enzymes based on expression. Together with the up-regulation of pyruvate dehydrogenase, this co-expression suggests an increased dependence on B. malayi products for energy production.
Another functional category represented in the up-regulated genes in F120 as compared to younger female stages is that of DNA replication. DNA polymerase III gamma/tau subunit (wBm0434) and recJ (wBm0124), a single-stranded DNA-specific exonuclease involved in single-strand break repair, and DNA/RNA helicase (wBm0708), required for both DNA replication and transcription, are significantly up-regulated at this stage, as is the RNA polymerase omega subunit (wBm0387), indicating an increase in transcription during this developmental stage. We determined two ribosomal proteins, S4 and S15, to also be significantly up-regulated at F120. The ribosomal protein S4 is essential for protein synthesis through its function in RNA binding, leading to fewer errors, while S15 plays an essential role in the assembly of the central domain of the small ribosomal subunit [31]. Our observations are consistent with findings in the Wolbachia populations in the gonads of O. ochengi showing differential regulation of Wolbachia genes required for DNA replication and translation, including ribosomal proteins in the germline [29]. Genes involved in lipid II/ peptidoglycan biosynthesis (wBm0493, metC, and wBm0492, murE) and wBm0490, a protein shown to interact with ftsH, a gene required for cell division, were also found to be significantly up-regulated in F120.
Among the genes differentially expressed over the course of female development are three peptidases including wBm0384, an extracellular metallopeptidase unique to the Wolbachia of B. malayi, that are potentially involved in the breakdown of filarial peptides for amino acids [6]. The other two genes, wBm0772 and wBm0552, encode ATP-dependent protease subunits.
Genomic analysis revealed that Wolbachia has maintained the biosynthetic pathways for purines and pyrimidines while B. malayi has not, suggesting that Wolbachia are potentially provisioning nucleotides to their filarial hosts, especially during times of increased need. In accordance with this hypothesis, wBm0443 guanosine monophosphate synthase, an essential enzyme in de novo purine biosynthesis, is differentially expressed during female worm development. Additionally, we find wBm0255 amidophosphoribosyltransferase, also involved in de novo purine biosynthesis, to be differentially expressed in both males and females. This supports a potential role of nucleotide production by Wolbachia in the lateral cords in male and female worm development.
It was proposed that Wolbachia might be inhibiting apoptosis in the worm host through the manipulation of the host apoptotic pathway [9, 29]. While very little is understood about how this may be occurring, we find significant up-regulation of three genes putatively involved in the manipulation of the apoptotic pathways in F120. One such gene is wBm0152, a Wolbachia surface protein, shown to inhibit apoptosis of purified human polymorphonuclear cells in vitro [32]. We also find significant up-regulation of wBm0296, an ankyrin repeat-containing protein, hypothesized to be an effector protein of the Type-IV secretion system (T4SS) able to mediate interactions with the host cells as they are for other intracellular bacteria [33–35]. Lastly, wBm0490, a gene with high homology to a bax-inhibitor in the Wolbachia of Drosophila, is significantly up-regulated in F120. Manipulation of the host apoptotic pathway through the expression of bax-inhibitors is believed to be responsible for the suppression of apoptosis of host cells by the obligate intracellular bacteria, Chlamydia trachomatis [36].
Constitutive expression of biosynthetic pathways in Wolbachia that are missing in B. malayi
The expression in Wolbachia of three biosynthetic pathways (heme, riboflavin, and FAD) potentially important for symbiosis as well as Wolbachia transporters, were examined in each developmental stage (Fig 4: Heatmap of gene expression of pathways of interest). Common to all Wolbachia genomes sequenced thus far is the presence of nearly all genes necessary for the synthesis of the iron-containing cofactor, heme, except for hemG, which is missing in many heme-producing bacteria [6, 28, 37]. Heme is an essential cofactor for cytochromes, peroxidases, and catalases, which are involved in a number of critical cellular processes including oxidative phosphorylation and electron transport. Heme is a co-factor for peroxidases essential for molting and might possibly also be a co-factor for steroids involved in molting of filarial parasites [28, 38]. Unlike what was shown for O. ochengi where very low expression of the heme biosynthetic pathway in adult tissues was detected [29], we found all Wolbachia genes involved in heme biosynthesis to be expressed in all sampled stages, with the highest expression at the L4 and F120 stages. Additionally, we find constitutively high expression of iron ABC transporters responsible for importing iron into the bacterial cell as well as heme ABC-transporters responsible for transporting heme from the bacterial cystoplasm into the periplasmic space and potentially involved in the transport of heme into the cytoplasm of the filarial host cell.
Fig 4. Heatmap of gene expression of pathways of interest.
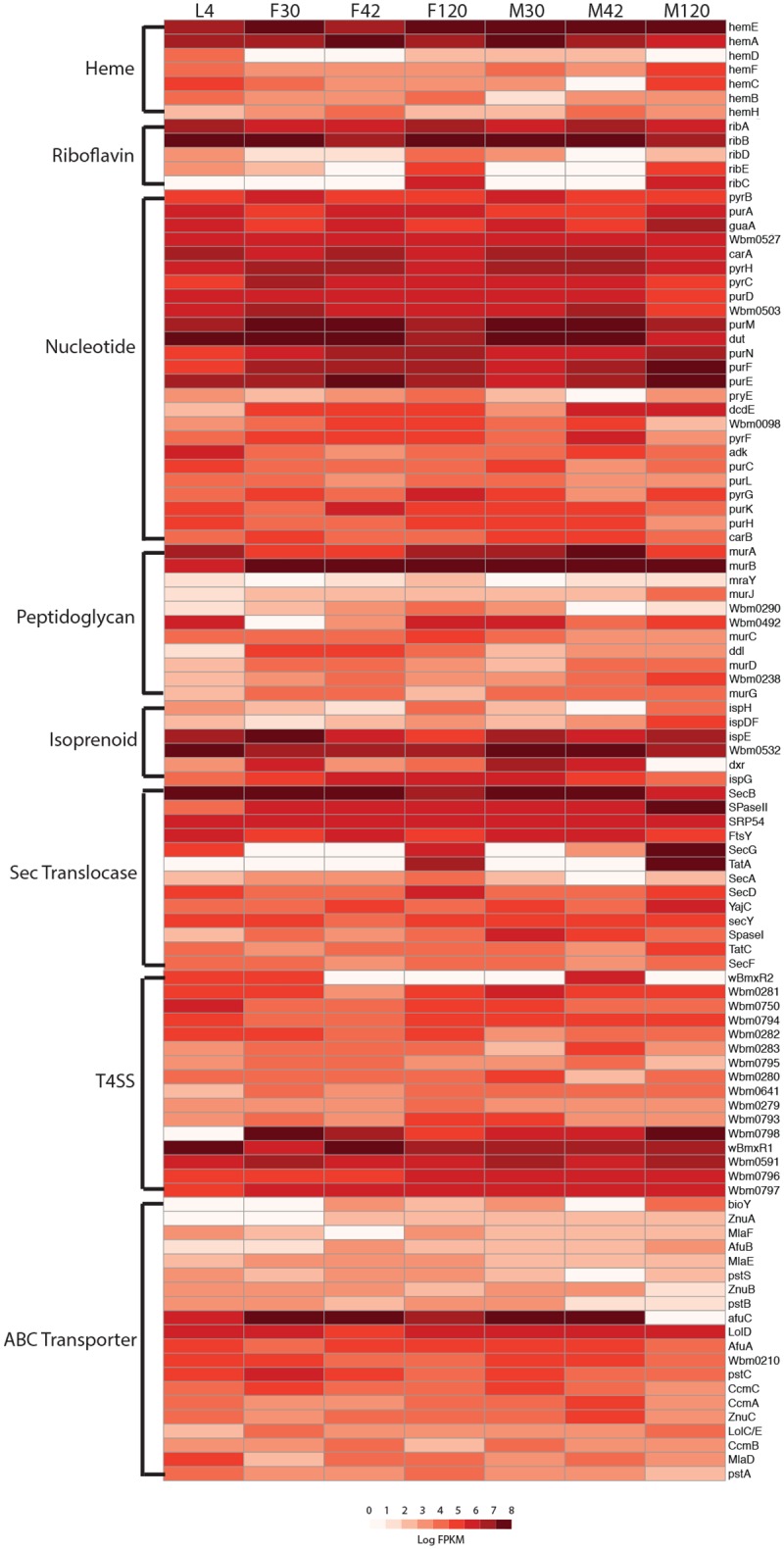
Expression, in log FPKMs, is plotted over the development of the nematode, L4 to 120 days post infection (dpi) male (M) and female (F), by pathway for heme, riboflavin, FAD, peptidoglycan, and isoprenoid biosynthesis as well as expression of the main secretion systems in Wolbachia including Sec translocase, Type IV secretion system, and ABC transporters.
Unlike Rickettsia, Wolbachia has maintained the ability to synthesize both riboflavin and FAD [6]. As riboflavin biosynthesis has been lost in B. malayi, it was hypothesized that Wolbachia were provisioning this cofactor to their filarial hosts. In support of this hypothesis we found that wBm0416, involved in FAD biosynthesis, as well as RibA and RibB are constitutively highly expressed across all stages of worm development. RibA is a bifunctional enzyme that catalyzes the first two essential steps in riboflavin biosynthesis, and is co-regulated with the T4SS [39]. F120 is the only stage in which we find all genes in the FAD/ riboflavin biosynthetic pathways to be classified as expressed, and at particularly high levels.
The Sec translocase system is responsible for the majority of protein trafficking across the bacterial cytoplasmic membrane into the periplasm with the use of ATP [40]. SecY is a transmembrane protein constituting the core of the protein-translocating complex. SecY was constitutively highly expressed across all stages of the life cycle. SecG associates with SecY to form a heterotrimeric complex. While not necessary for general function of the system, SecG has been shown to facilitate transport at low temperatures (20°C), or when the proton-motive-force is reduced [41, 42]. We find expression of secG only in L4, M42, F120, and M120 where we see particularly high expression. While Wolbachia lack the tatB gene, part of the Sec-independent twin arginine translocation (Tat) protein system present in most bacteria, they do maintain TatA and TatC genes. Thus, as in other alpha-proteobacteria, it is likely that this system is still functional [43]. TatA was highly expressed in F120 and M120 exclusively, while TatC was expressed constitutively across all stages. Secretion in Wolbachia requires not only translocation into the periplasmic space by either the Sec or Tat systems, but transport across the outer membrane as well. This is accomplished by the T4SS, a leader-peptide independent mechanism for transporting effector proteins and virulence factors found in many pathogenic and endosymbiotic bacteria [35, 44–46]. We find constitutive expression of nearly all genes in this pathway at most stages except for wBm0798 in L4.
ATP-binding cassette transporters (ABC transporters) are composed of two transmembrane domains and two cytoplasmic ATP-binding domains. They are involved in the uptake of a variety of nutrients and the extrusion of drugs and metabolites [47]. As previously mentioned, we saw constitutive expression of all four heme ABC transporters encoded in the Wolbachia genome, as well as two lipoprotein transporters. Constitutive expression of the lipoprotein transport system LolCDE is required to export lipoproteins to the outer membrane [48]. Lipoproteins have been shown to be agonists of inflammatory pathogenesis in lymphatic filariasis, recognized by the TLR-2 and TLR-6 in the human host [49]. Correspondingly, we see significant up-regulation in adult female worms of the wBm0152 peptidoglycan-associated lipoprotein-like outer membrane protein shown to be localized to numerous sites on the bacterial membrane [50]. We also find constitutively high expression of two phosphate transporters, potentially required for importing phosphate for nucleotide production. Experiments in L. sigmodontis show that when Wolbachia is depleted with tetracycline, expression of a filarial phosphate transporter is significantly increased to compensate for the decrease in Wolbachia-produced nucleotides that are essential for worm embryogenesis and survival [51].
Co-expressed nematode and bacterial genes
To determine which genes were being co-expressed between B. malayi and Wolbachia, we built a co-expression network for the two organisms using WGCNA (Fig 5: The co-expression network for B. malayi and Wolbachia). WGCNA is a well-established method by which expression data and trait data are integrated to identify co-expressed pathways. We used hierarchical clustering and dynamic cutting to determine modules of co-expression. A module is a cluster of interconnected genes with high correlation based on their expression profiles (S5 Table: Summary of module membership). GO term enrichment of each resulting module was performed to determine which modules were biologically significant (S6 Table: GO term Enrichment). To identify modules of gene co-expression with the most interest based on the symbiotic interaction of the two organisms, we evaluated the correlation of each module to a measure of Wolbachia population, wsp/gst. We found three modules that had the highest negative correlations with the wsp/gst ratio (brown, -0.57 p-value = 0.03; yellow and green, -0.65 p-value = 0.01). A fourth module had the highest positive correlation with the wsp/gst ratio (salmon, 0.63 p-value = 0.02). Because the F120 samples have the lowest wsp/gst ratio ratio due to the large size of the female worms, we determined that the three modules with the highest negative correlation were indicative of adult female gene expression.
Fig 5. The co-expression network for B. malayi and Wolbachia.
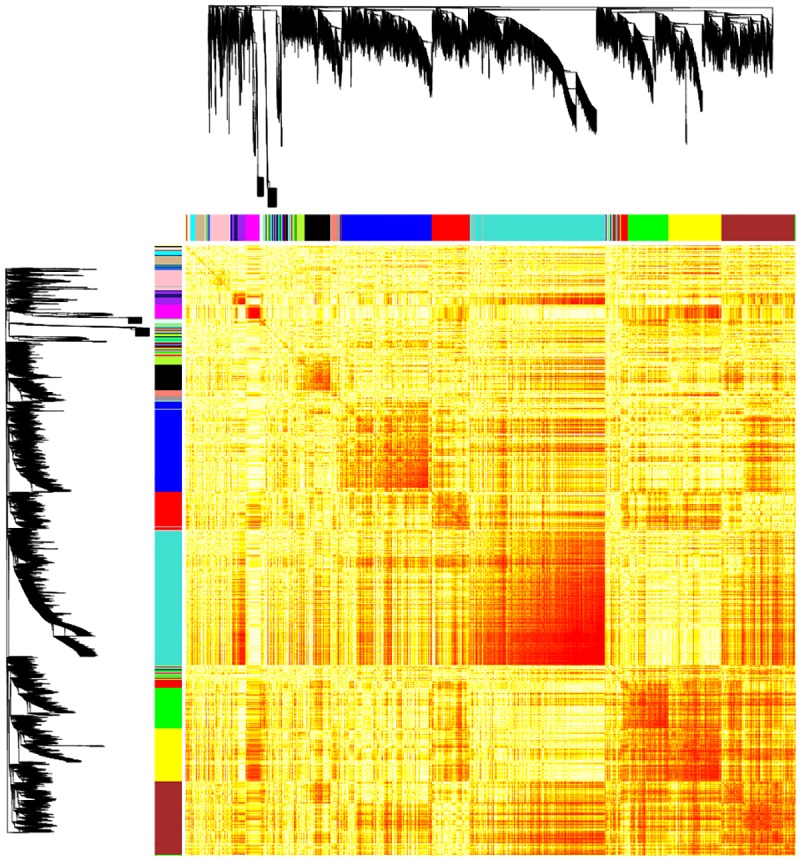
The network heatmap plot of network connectivity for B. malayi and Wolbachia calculated using weighted gene correlation network analysis (WGCNA). The black branches show the hierarchical clustering dendrograms, which were assigned to clusters using dynamic tree cutting to identify modules of co-expressed genes, shown as colored bars. High co-expression interconnectedness is indicated by increasingly saturated orange and red coloring. Modules correspond to groups of highly interconnected genes.
The green module contains 2,312 B. malayi genes and 28 Wolbachia genes (S5 Table); it shows an enrichment of DE genes that are up-regulated in the adult females, with a p-value < 2.2e-16. The most significantly enriched GO terms in the green module include intracellular signal transduction, proteolysis, transmembrane transport, cyclic nucleotide biosynthetic process, cysteine-type peptidase activity, and mitochondrion. Interestingly, this module also contains the cathepsin-like cysteine protease Bma-cpl-6 found to be involved in development and embryogenesis in the worm as well as in Wolbachia expansion (S6 Table) [52]. The yellow module contains 2,782 B. malayi genes and 39 Wolbachia genes and shows an enrichment of GO terms that include the regulation of transcription, oxidoreductase activity, sequence-specific DNA binding, and cell redox homeostasis. Interestingly, the co-expressed Wolbachia genes in this module include six of the seven chaperones found to be differentially expressed during female development as well as a gene involved in redox sensing and two genes involved in cytochrome c biogenesis. Cytochrome c is known to play a role in the electron transport chain and cell apoptosis, as well as being an antioxidative enzyme by removing superoxide and hydrogen peroxide. The brown module contains 4,148 B. malayi genes and 64 Wolbachia genes and shows a significant under-representation of DE genes (p-values < 2.2e-16) in F120 as compared to F42. As one of the modules that are the most highly negatively correlated to the Wolbachia population, the brown module contains a number of genes implicated in the host/endosymbiont relationship including genes involved in riboflavin and nucleotide biosynthesis (S5 Table). Among the most enriched GO terms in the brown module are metabolism and transport. Co-expressed Wolbachia genes include a number of genes also involved in transport, such as three ABC transporters, including an iron importer, a general permease exporter, and a polyamine transporter. Among the many B. malayi genes involved in transport co-expressed in this module is Bm4941, one of three genes in the genome predicted to have nucleoside transmembrane transporter activity based on protein domain information. In the brown module, we find the GO term glycogen catabolic process (GO:0005980) to be significantly enriched, as well as the genes Bm5241 in B. malayi and wBm0207 in Wolbachia, previously mentioned in relation to pyruvate metabolism, to be co-expressed in this module. Among the Wolbachia genes co-expressed in this module are four genes involved in de novo purine and pyrimidine biosynthesis. Also in the brown module are two Wolbachia genes in the riboflavin biosynthesis pathway, including RibA, which are co-expressed with a number of B. malayi riboflavin or flavin-requiring proteins. Another set of enriched GO terms in the brown module includes DNA repair and replication, a function also represented with the co-expressed Wolbachia genes that include five genes involved in DNA replication and repair, including ribonucleotide reductase, an enzyme integral in controlling the rate of DNA synthesis [53]. Correspondingly, two genes involved in cell division in Wolbachia are co-expressed in this module. Interestingly, the salmon module, the module with the highest positive correlation to Wolbachia population, is enriched for GO terms related to regulation of apoptosis, response to oxidative stress, oxidase activity, and heme binding.
Discussion
The dual transcriptional profiling that we performed revealed potential stage-specific requirements from Wolbachia during filarial development and embryogenesis. Differentially expressed Wolbachia genes during the course of female development generally fell into the functional categories of chaperone function, energy production, nucleotide biosynthesis, DNA replication, and anti-oxidative defense. These categories include genes that are likely to be required for specific developmental processes, including germline and embryonic development in the nematode and Wolbachia invasion of the gonad. Similar studies have been performed on the filarial nematode Dirofilaria immitis, or dog heartworm, and its Wolbachia endosymbiont, wDi [30, 54]. While the first study found no wDi genes differentially expressed between adult male and female worms, a second tissue-specific study reported differentially expressed wDi genes in the uterus and female body wall. Our findings including wBm genes differentially expressed over the course of female development correspond to those wDi genes found to be up-regulated in the uterus, including multiple genes encoding ribosomal proteins, DNA replication and repair machinery, a tRNA synthetase, and a component of the purine biosynthetic pathway. This indicates that the role Wolbachia play in certain life stages of the filaria may be well conserved.
It has been proposed that the ability of Wolbachia to perform aerobic respiration and metabolize iron whilst responding to oxidative stress may be an essential mechanism of the endosymbiotic relationship with filarial worms [28, 29]. Studies in Litomosoides sigmodontis, for example, have found that targeting Wolbachia with antibiotics resulted in the up-regulation of components of the mitochondrial respiratory chain [55]. Experiments in O. ochengi showed that worms treated with antibiotics lose motility and that Wolbachia density in infected cells greatly exceeded that of mitochondria [56]. These results point to the potential ATP provisioning by Wolbachia to the filarial host [56]. The up-regulation of genes involved in the ATP transport chain and in iron-sulfur cluster formation—which are essential co-factors for respiratory chain proteins in ATP production—provide support for this hypothesis but it is difficult to prove. Alternatively, the up-regulation of ATP production could be required for the increased propagation of Wolbachia at this stage.
The potential production of ATP by Wolbachia for its filarial host likely contributes to oxidative stress of their cellular environment by generating ROS as by-products of aerobic metabolism. Consistent with this hypothesis, many genes that encode proteins known to be involved in combating oxidative stress were highly differentially expressed during female development, including chaperone proteins and a gene involved in single-strand break repair, a potential consequence of an increasingly oxidative environment. A number of these chaperone proteins have been shown to be part of the oxidative stress response in bacteria and to maintain their stability under oxidative conditions [57, 58]. It was proposed that overexpression of groEL is an important adaptation allowing for the obligate intracellular lifestyle of Wolbachia within a cytoplasmic vesicle [59, 60]. We find that these requirements appear to be of special importance during female development, potentially as a consequence of the up-regulation of oxidative phosphorylation. Chaperones GroEL, HSP60, and DnaK were found to be among the proteins with the most abundant peptide counts in proteomic analysis of B. malayi in the adult stages of the worm [61]. This study did not however look at abundance over the course of worm development, L4 through 30–42 dpi. The chaperone HslU forms a complex with the peptidase HslV, which was also found to be significantly up-regulated in the F30 and F42 stages. HslV, or HSP20, has additionally been shown to be involved in bacteria-host interactions in Helicobacter pylori [62]. Interestingly, the DnaJ/K chaperones are among the Wolbachia genes found to be inserted in the nuclear genomes as well as expressed by the Wolbachia-free filariae A. viteae and O. flexuosa [63].
While many endosymbionts and parasites, including B. malayi, as well as members of the Rickettsia genus have lost the pathways for de novo purine and pyrimidine synthesis, Wolbachia has maintained these biosynthetic pathways. Additionally, Wolbachia was shown to lack the ADP/ATP translocases used by other endosymbionts, including the parasitic Rickettsia and Chlamydia, and the mutualist Buchnera, to scavenge for nucleotides from the host [8]. These observations, combined with the evidence for positive selection on genes in this pathway suggest that Wolbachia produce nucleotides not only for internal consumption but also for the host at times when the requirement for DNA synthesis is particularly high, such as during oogenesis and embryogenesis [6, 8]. During the mitotic proliferation of the B. malayi oogonia, Wolbachia divides rapidly, requiring increased expression of the replication machinery [64]. Correspondingly, we found in F120 significant up-regulation of the Wolbachia DNA replication machinery and genes involved in transcription and translation.
In our quest to capture the basis of the endosymbiotic relationship between B. malayi and Wolbachia, we looked at the expression of genes that are part of biosynthetic pathways in Wolbachia that are missing in B. malayi. The absence of the heme, riboflavin and FAD biosynthetic pathways in filaria led to the hypothesis that Wolbachia could be providing these to the filarial host. Evidence for this, however, remains elusive. Several studies have shown that inhibitors of the heme biosynthetic pathway such as 5-aminolevulinate (ALAD) and N-methyl mesoporphyrin (NMMP) have adverse effects on B. malayi, causing a marked reduction in motility [38]. While this suggests a role for Wolbachia in provisioning heme, adverse effects were also observed on C. elegans, which also lack the heme biosynthetic pathways and are Wolbachia-free, suggesting non-specific effects. We found constitutive expression of all genes in the heme biosynthetic pathway of Wolbachia as well as of heme ABC transporters at almost all sampled worm stages, with the highest expression at the L4 and F120 stages. This suggests the importance of heme synthesis and transport in the symbiotic relationship at these stages [29]. However, it remains unknown if or how B. malayi might receive heme from its bacterial endosymbiont. Examination of the riboflavin and FAD biosynthetic pathways in Wolbachia revealed F120 as the only stage in which all genes are highly expressed, suggesting an increased need in adult females for riboflavin and FAD for embryogenesis. This finding is consistent with the observation that when adult worms are grown in the presence of doxycycline, causing severe adverse effects in embryogenesis, supplementation with riboflavin is able to rescue embryogenesis in adult female worms by approximately 50% [39].
If Wolbachia are indeed provisioning B. malayi with metabolites or nutrients, they would require active secretion and transport systems to do so. We determined that the Sec-dependent and Sec-independent systems appear to be constitutively expressed, especially in adult females. We also find genes in the T4SS, responsible for transport across the outer membrane, to be constitutively expressed across all sampled stages with the exception of wBm0798 in L4. This suggests that the T4SS is not only active in Wolbachia, but also important in all stages of development that were included in this study. These results confirm the potential for the filarial dependence on Wolbachia products at all stages of the life cycle. Constitutive expression of the heme, phosphate, and lipoprotein ABC transporters is also consistent with the expression of these biosynthetic pathways as phosphate is an essential molecule in nucleotides.
The co-expression network analysis of Wolbachia and B. malayi genes was another approach to define interactions. The use of WGCNA to construct a co-expression network for both Wolbachia and B. malayi genes revealed co-expression of important pathways. A number of resulting modules were significantly correlated with Wolbachia density, either positively or negatively. Analysis of GO enrichment of the modules as well as module membership revealed a number of pathways of interest including redox homeostasis and oxidative stress responses as well as the co-expression of DNA repair and replication between the two organisms. Transport mechanisms were also co-expressed, including a nucleoside transporter in B. malayi co-expressed with Wolbachia genes involved in de novo purine and pyrimidine biosynthesis. We plan to expand this co-expression network to include additional stages of parasite development, including molting larvae and microfilaria, in order to better represent the dynamics of endosymbiosis over the entire parasitic lifecycle.
In conclusion, our study provides novel insight into the complexity of the interactions between B. malayi and its endosymbiotic bacteria, Wolbachia. We find that it is unlikely that this obligate symbiotic relationship relies on a single process or pathway, but rather on more complex interactions that likely vary over the life cycle of the parasite. This work paves the way for functional validation of the essential role of these associations through the use of RNAi experiments. Elucidation of essential pathways involved in the endosymbiosis between Wolbachia and B. malayi will allow for the identification of novel drug targets.
Supporting information
a. Clustering of biological replicates using multidimensional scaling analysis of the top 50% most highly expressed B. malayi genes. b. Clustering of biological replicates using multidimensional scaling analysis of the top 50% most highly expressed Wolbachia genes. Samples taken during nematode development from L4 to 120 days post infection (dpi), in males (M) and females (F), with a and b denoting separate biological replicates.
(TIF)
Seven genes (10 pair-wise comparisons) were chosen for confirmation with RT-qPCR. Four of the genes (WSP, Hsp90, life cycle DnaK, and GroEL), with seven pair-wise comparisons, were chosen based on the criteria that they were found to be significantly differentially expressed and had over 50 read counts per stage. Also included are three genes (RibA, HemA, and AfuA) that were found to be constitutively expressed, based on an FDR of 1 in EdgeR. Spearman correlation of 0.987 and a p-value < 2.2e-16.
(TIF)
Table showing results of pair-wise differential expression analyses between female stages.
(XLSX)
Table showing results of pair-wise differential expression analyses between female stages.
(XLSX)
Table showing results of pair-wise differential expression analyses between male and female stages.
(XLSX)
Table showing results from all pair-wise differential expression analyses for Wolbachia.
(XLSX)
Table showing the module assignments resulting from the hierarchical clustering for B. malayi and Wolbachia genes.
(XLSX)
Table showing the results of the GO term enrichment analysis for each module, p-value < 0.01.
(XLSX)
Acknowledgments
The authors would like to thank Mr. Saheed Bachu for his assistance with the RNA isolation and qPCR experiments as well as FR3 for providing the parasite material.
Data Availability
All sequencing files are available from the Sequence Read Archive (SRA), Accession number SRP090644 https://www.ncbi.nlm.nih.gov/sra.
Funding Statement
This work was supported by the National Institute of Allergy and Infectious Diseases, National Institutes of Health www.niaid.nih.gov under grants R56 AI118936 and R56 AI101372 awarded to SL, TRU, and EG. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Lustigman S., et al. , A research agenda for helminth diseases of humans: the problem of helminthiases. PLoS Negl Trop Dis, 2012. 6(4): p. e1582 10.1371/journal.pntd.0001582 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Disease G.B.D., Injury I., and Prevalence C., Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: a systematic analysis for the Global Burden of Disease Study 2015. Lancet, 2016. 388(10053): p. 1545–1602. 10.1016/S0140-6736(16)31678-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hoerauf A., et al. , Filariasis in Africa—treatment challenges and prospects. Clin Microbiol Infect, 2011. 17(7): p. 977–85. 10.1111/j.1469-0691.2011.03586.x [DOI] [PubMed] [Google Scholar]
- 4.Taylor M.J., Hoerauf A., and Bockarie M., Lymphatic filariasis and onchocerciasis. Lancet, 2010. 376(9747): p. 1175–85. 10.1016/S0140-6736(10)60586-7 [DOI] [PubMed] [Google Scholar]
- 5.Slatko B.E., et al. , Wolbachia endosymbionts and human disease control. Mol Biochem Parasitol, 2014. 195(2): p. 88–95. 10.1016/j.molbiopara.2014.07.004 [DOI] [PubMed] [Google Scholar]
- 6.Foster J., et al. , The Wolbachia genome of Brugia malayi: endosymbiont evolution within a human pathogenic nematode. PLoS Biol, 2005. 3(4): p. e121 10.1371/journal.pbio.0030121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ghedin E., et al. , Draft genome of the filarial nematode parasite Brugia malayi. Science, 2007. 317(5845): p. 1756–60. 10.1126/science.1145406 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brownlie J.C., et al. , Diversifying selection and host adaptation in two endosymbiont genomes. BMC Evol Biol, 2007. 7: p. 68 10.1186/1471-2148-7-68 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Landmann F., et al. , Anti-filarial activity of antibiotic therapy is due to extensive apoptosis after Wolbachia depletion from filarial nematodes. PLoS Pathog, 2011. 7(11): p. e1002351 10.1371/journal.ppat.1002351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Desjardins C.A., et al. , Genomics of Loa loa, a Wolbachia-free filarial parasite of humans. Nat Genet, 2013. 45(5): p. 495–500. 10.1038/ng.2585 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kim D., et al. , TopHat2: accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol, 2013. 14(4): p. R36 10.1186/gb-2013-14-4-r36 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Trapnell C., et al. , Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol, 2010. 28(5): p. 511–5. 10.1038/nbt.1621 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Trapnell C., et al. , Differential analysis of gene regulation at transcript resolution with RNA-seq. Nat Biotechnol, 2013. 31(1): p. 46–53. 10.1038/nbt.2450 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Anders S., Pyl P.T., and Huber W., HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics, 2015. 31(2): p. 166–9. 10.1093/bioinformatics/btu638 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Robinson M.D., McCarthy D.J., and Smyth G.K., edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics, 2010. 26(1): p. 139–40. 10.1093/bioinformatics/btp616 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Langfelder P. and Horvath S., WGCNA: an R package for weighted correlation network analysis. BMC Bioinformatics, 2008. 9: p. 559 10.1186/1471-2105-9-559 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.McGarry H.F., Egerton G.L., and Taylor M.J., Population dynamics of Wolbachia bacterial endosymbionts in Brugia malayi. Mol Biochem Parasitol, 2004. 135(1): p. 57–67. [DOI] [PubMed] [Google Scholar]
- 18.Choi Y.J., et al. , A deep sequencing approach to comparatively analyze the transcriptome of lifecycle stages of the filarial worm, Brugia malayi. PLoS Negl Trop Dis, 2011. 5(12): p. e1409 10.1371/journal.pntd.0001409 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cabiscol E., Tamarit J., and Ros J., Oxidative stress in bacteria and protein damage by reactive oxygen species. Int Microbiol, 2000. 3(1): p. 3–8. [PubMed] [Google Scholar]
- 20.Ericsson M., et al. , Increased synthesis of DnaK, GroEL, and GroES homologs by Francisella tularensis LVS in response to heat and hydrogen peroxide. Infect Immun, 1994. 62(1): p. 178–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Farr S.B. and Kogoma T., Oxidative stress responses in Escherichia coli and Salmonella typhimurium. Microbiol Rev, 1991. 55(4): p. 561–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Susin M.F., et al. , GroES/GroEL and DnaK/DnaJ have distinct roles in stress responses and during cell cycle progression in Caulobacter crescentus. J Bacteriol, 2006. 188(23): p. 8044–53. 10.1128/JB.00824-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Walkup L.K. and Kogoma T., Escherichia coli proteins inducible by oxidative stress mediated by the superoxide radical. J Bacteriol, 1989. 171(3): p. 1476–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Seong I.S., et al. , The HslU ATPase acts as a molecular chaperone in prevention of aggregation of SulA, an inhibitor of cell division in Escherichia coli. FEBS Lett, 2000. 477(3): p. 224–9. [DOI] [PubMed] [Google Scholar]
- 25.Azim M.K., et al. , Characterization of the HslU chaperone affinity for HslV protease. Protein Sci, 2005. 14(5): p. 1357–62. 10.1110/ps.04970405 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Tchankouo-Nguetcheu S., et al. , Differential protein modulation in midguts of Aedes aegypti infected with chikungunya and dengue 2 viruses. PLoS One, 2010. 5(10). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rzezniczak T.Z. and Merritt T.J., Interactions of NADP-reducing enzymes across varying environmental conditions: a model of biological complexity. G3 (Bethesda), 2012. 2(12): p. 1613–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gill A.C., Darby A.C., and Makepeace B.L., Iron necessity: the secret of Wolbachia's success? PLoS Negl Trop Dis, 2014. 8(10): p. e3224 10.1371/journal.pntd.0003224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Darby A.C., et al. , Analysis of gene expression from the Wolbachia genome of a filarial nematode supports both metabolic and defensive roles within the symbiosis. Genome Res, 2012. 22(12): p. 2467–77. 10.1101/gr.138420.112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Luck A.N., et al. , Tissue-specific transcriptomics and proteomics of a filarial nematode and its Wolbachia endosymbiont. BMC Genomics, 2015. 16: p. 920 10.1186/s12864-015-2083-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Held W.A., et al. , Assembly mapping of 30 S ribosomal proteins from Escherichia coli. Further studies. J Biol Chem, 1974. 249(10): p. 3103–11. [PubMed] [Google Scholar]
- 32.Bazzocchi C., et al. , Wolbachia surface protein (WSP) inhibits apoptosis in human neutrophils. Parasite Immunol, 2007. 29(2): p. 73–9. 10.1111/j.1365-3024.2006.00915.x [DOI] [PubMed] [Google Scholar]
- 33.Pan X., et al. , Ankyrin repeat proteins comprise a diverse family of bacterial type IV effectors. Science, 2008. 320(5883): p. 1651–4. 10.1126/science.1158160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Rikihisa Y. and Lin M., Anaplasma phagocytophilum and Ehrlichia chaffeensis type IV secretion and Ank proteins. Curr Opin Microbiol, 2010. 13(1): p. 59–66. 10.1016/j.mib.2009.12.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Rances E., et al. , Genetic and functional characterization of the type IV secretion system in Wolbachia. J Bacteriol, 2008. 190(14): p. 5020–30. 10.1128/JB.00377-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fischer S.F., et al. , Protection against CD95-induced apoptosis by chlamydial infection at a mitochondrial step. Infect Immun, 2004. 72(2): p. 1107–15. 10.1128/IAI.72.2.1107-1115.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hamza I. and Dailey H.A., One ring to rule them all: trafficking of heme and heme synthesis intermediates in the metazoans. Biochim Biophys Acta, 2012. 1823(9): p. 1617–32. 10.1016/j.bbamcr.2012.04.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wu B., et al. , The heme biosynthetic pathway of the obligate Wolbachia endosymbiont of Brugia malayi as a potential anti-filarial drug target. PLoS Negl Trop Dis, 2009. 3(7): p. e475 10.1371/journal.pntd.0000475 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li Z. and Carlow C.K., Characterization of transcription factors that regulate the type IV secretion system and riboflavin biosynthesis in Wolbachia of Brugia malayi. PLoS One, 2012. 7(12): p. e51597 10.1371/journal.pone.0051597 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.du Plessis D.J., Nouwen N., and Driessen A.J., The Sec translocase. Biochim Biophys Acta, 2011. 1808(3): p. 851–65. 10.1016/j.bbamem.2010.08.016 [DOI] [PubMed] [Google Scholar]
- 41.Nishiyama K., Hanada M., and Tokuda H., Disruption of the gene encoding p12 (SecG) reveals the direct involvement and important function of SecG in the protein translocation of Escherichia coli at low temperature. EMBO J, 1994. 13(14): p. 3272–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hanada M., Nishiyama K., and Tokuda H., SecG plays a critical role in protein translocation in the absence of the proton motive force as well as at low temperature. FEBS Lett, 1996. 381(1–2): p. 25–8. [DOI] [PubMed] [Google Scholar]
- 43.Nunez P.A., Soria M., and Farber M.D., The twin-arginine translocation pathway in alpha-proteobacteria is functionally preserved irrespective of genomic and regulatory divergence. PLoS One, 2012. 7(3): p. e33605 10.1371/journal.pone.0033605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Masui S., Sasaki T., and Ishikawa H., Genes for the type IV secretion system in an intracellular symbiont, Wolbachia, a causative agent of various sexual alterations in arthropods. J Bacteriol, 2000. 182(22): p. 6529–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Pichon S., et al. , Conservation of the Type IV secretion system throughout Wolbachia evolution. Biochem Biophys Res Commun, 2009. 385(4): p. 557–62. 10.1016/j.bbrc.2009.05.118 [DOI] [PubMed] [Google Scholar]
- 46.Christie P.J., Whitaker N., and Gonzalez-Rivera C., Mechanism and structure of the bacterial type IV secretion systems. Biochim Biophys Acta, 2014. 1843(8): p. 1578–91. 10.1016/j.bbamcr.2013.12.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ren Q. and Paulsen I.T., Comparative analyses of fundamental differences in membrane transport capabilities in prokaryotes and eukaryotes. PLoS Comput Biol, 2005. 1(3): p. e27 10.1371/journal.pcbi.0010027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Yakushi T., et al. , A new ABC transporter mediating the detachment of lipid-modified proteins from membranes. Nat Cell Biol, 2000. 2(4): p. 212–8. 10.1038/35008635 [DOI] [PubMed] [Google Scholar]
- 49.Turner J.D., et al. , Wolbachia lipoprotein stimulates innate and adaptive immunity through Toll-like receptors 2 and 6 to induce disease manifestations of filariasis. J Biol Chem, 2009. 284(33): p. 22364–78. 10.1074/jbc.M901528200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Voronin D., et al. , Wolbachia lipoproteins: abundance, localisation and serology of Wolbachia peptidoglycan associated lipoprotein and the Type IV Secretion System component, VirB6 from Brugia malayi and Aedes albopictus. Parasit Vectors, 2014. 7: p. 462 10.1186/s13071-014-0462-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Heider U., et al. , Differential display of genes expressed in the filarial nematode Litomosoides sigmodontis reveals a putative phosphate permease up-regulated after depletion of Wolbachia endobacteria. Int J Med Microbiol, 2006. 296(4–5): p. 287–99. 10.1016/j.ijmm.2005.12.019 [DOI] [PubMed] [Google Scholar]
- 52.Lustigman S., et al. , Potential involvement of Brugia malayi cysteine proteases in the maintenance of the endosymbiotic relationship with Wolbachia. Int J Parasitol Drugs Drug Resist, 2014. 4(3): p. 267–77. 10.1016/j.ijpddr.2014.08.001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Herrick J. and Sclavi B., Ribonucleotide reductase and the regulation of DNA replication: an old story and an ancient heritage. Mol Microbiol, 2007. 63(1): p. 22–34. 10.1111/j.1365-2958.2006.05493.x [DOI] [PubMed] [Google Scholar]
- 54.Luck A.N., et al. , Concurrent transcriptional profiling of Dirofilaria immitis and its Wolbachia endosymbiont throughout the nematode life cycle reveals coordinated gene expression. BMC Genomics, 2014. 15: p. 1041 10.1186/1471-2164-15-1041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Strubing U., et al. , Mitochondrial genes for heme-dependent respiratory chain complexes are up-regulated after depletion of Wolbachia from filarial nematodes. Int J Parasitol, 2010. 40(10): p. 1193–202. 10.1016/j.ijpara.2010.03.004 [DOI] [PubMed] [Google Scholar]
- 56.Gilbert J., et al. , Antibiotic chemotherapy of onchocerciasis: in a bovine model, killing of adult parasites requires a sustained depletion of endosymbiotic bacteria (Wolbachia species). J Infect Dis, 2005. 192(8): p. 1483–93. 10.1086/462426 [DOI] [PubMed] [Google Scholar]
- 57.Melkani G.C., Zardeneta G., and Mendoza J.A., Oxidized GroEL can function as a chaperonin. Front Biosci, 2004. 9: p. 724–31. [DOI] [PubMed] [Google Scholar]
- 58.Melkani G.C., et al. , Divalent cations stabilize GroEL under conditions of oxidative stress. Biochem Biophys Res Commun, 2008. 368(3): p. 625–30. 10.1016/j.bbrc.2008.01.126 [DOI] [PubMed] [Google Scholar]
- 59.Fares M.A., Moya A., and Barrio E., GroEL and the maintenance of bacterial endosymbiosis. Trends Genet, 2004. 20(9): p. 413–6. 10.1016/j.tig.2004.07.001 [DOI] [PubMed] [Google Scholar]
- 60.Brennan L.J., et al. , The endosymbiont Wolbachia pipientis induces the expression of host antioxidant proteins in an Aedes albopictus cell line. PLoS One, 2008. 3(5): p. e2083 10.1371/journal.pone.0002083 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Bennuru S., et al. , Stage-specific proteomic expression patterns of the human filarial parasite Brugia malayi and its endosymbiont Wolbachia. Proc Natl Acad Sci U S A, 2011. 108(23): p. 9649–54. 10.1073/pnas.1011481108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Du R.J. and Ho B., Surface localized Heat Shock Protein 20 (HslV) of Helicobacter pylori. Helicobacter, 2003. 8(4): p. 257–67. [DOI] [PubMed] [Google Scholar]
- 63.McNulty S.N., et al. , Endosymbiont DNA in endobacteria-free filarial nematodes indicates ancient horizontal genetic transfer. PLoS One, 2010. 5(6): p. e11029 10.1371/journal.pone.0011029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Landmann F., et al. , Asymmetric Wolbachia segregation during early Brugia malayi embryogenesis determines its distribution in adult host tissues. PLoS Negl Trop Dis, 2010. 4(7): p. e758 10.1371/journal.pntd.0000758 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
a. Clustering of biological replicates using multidimensional scaling analysis of the top 50% most highly expressed B. malayi genes. b. Clustering of biological replicates using multidimensional scaling analysis of the top 50% most highly expressed Wolbachia genes. Samples taken during nematode development from L4 to 120 days post infection (dpi), in males (M) and females (F), with a and b denoting separate biological replicates.
(TIF)
Seven genes (10 pair-wise comparisons) were chosen for confirmation with RT-qPCR. Four of the genes (WSP, Hsp90, life cycle DnaK, and GroEL), with seven pair-wise comparisons, were chosen based on the criteria that they were found to be significantly differentially expressed and had over 50 read counts per stage. Also included are three genes (RibA, HemA, and AfuA) that were found to be constitutively expressed, based on an FDR of 1 in EdgeR. Spearman correlation of 0.987 and a p-value < 2.2e-16.
(TIF)
Table showing results of pair-wise differential expression analyses between female stages.
(XLSX)
Table showing results of pair-wise differential expression analyses between female stages.
(XLSX)
Table showing results of pair-wise differential expression analyses between male and female stages.
(XLSX)
Table showing results from all pair-wise differential expression analyses for Wolbachia.
(XLSX)
Table showing the module assignments resulting from the hierarchical clustering for B. malayi and Wolbachia genes.
(XLSX)
Table showing the results of the GO term enrichment analysis for each module, p-value < 0.01.
(XLSX)
Data Availability Statement
All sequencing files are available from the Sequence Read Archive (SRA), Accession number SRP090644 https://www.ncbi.nlm.nih.gov/sra.