

ABSTRACT
Drosophila adults have been utilized as a genetically tractable model organism to decipher the molecular mechanisms of humoral innate immune responses. In an effort to promote the utility of Drosophila larvae as an additional model system, in this study, we describe a novel aspect of an induction mechanism for innate immunity in these larvae. By using a fine tungsten needle created for manipulating semi-conductor devices, larvae were subjected to septic injury. However, although Toll pathway mutants were susceptible to infection with Gram-positive bacteria as had been shown for Drosophila adults, microbe clearance was not affected in the mutants. In addition, Drosophila larvae were found to be sensitive to mechanical stimuli with respect to the activation of a sterile humoral response. In particular, pinching with forceps to a degree that might cause minor damage to larval tissues could induce the expression of the antifungal peptide gene Drosomycin; notably, this induction was partially independent of the Toll and immune deficiency pathways. We therefore propose that Drosophila larvae might serve as a useful model to analyze the infectious and non-infectious inflammation that underlies various inflammatory diseases such as ischemia, atherosclerosis and cancer.
KEY WORDS: Innate immunity, Drosophila, Larvae
Editors' choice: Drosophila larvae represent a useful model of infectious inflammation and sterilely induced humoral innate response, which are relevant for disorders associated with chronic inflammation.
INTRODUCTION
Drosophila adults have been used as a leading model organism to investigate molecular mechanisms of innate immunity (Lemaitre and Hoffmann, 2007; Buchon et al., 2014) since it was first demonstrated in 1996 that the Toll pathway, which was initially characterized as an essential pathway for dorsoventral patterning in Drosophila embryos (Anderson et al., 1985a,b), was required for the induction of the antifungal peptide gene Drosomycin (Drs) upon fungal infection (Lemaitre et al., 1996). In particular, Drosophila adult models have contributed to identifying genes required for the humoral innate immune responses and for the production of antimicrobial peptides (AMPs) and melanization factors (Lemaitre et al., 1995; Rämet and Hultmark, 2014). In Drosophila adults, AMP induction upon challenge with microbes is controlled by two distinct signaling pathways, the Toll and immune deficiency (IMD) pathways (Lemaitre and Hoffmann, 2007; Valanne et al., 2011; Myllymäki et al., 2014). The Toll pathway is required for the induction of Drs and for survival following systemic infection with Gram-positive bacteria or fungi (Ferrandon et al., 2007). Specifically, the recognition of lysine-type peptidoglycans or β-glucans from microbes by the PGRP-SA/GNBP1 complex or by GNBP3 in the hemolymph activates modular serine protease (ModSP), followed by activation of Spätzle (Spz)-processing enzyme and cleavage of Spz, a protein ligand of the Toll receptor (Gottar et al., 2002, 2003, 2006; Jang et al., 2006; Buchon et al., 2009,b). In addition, so-called ‘danger signals’ also activate the Toll pathway through the protease Persephone (Psh). For example, exogenous danger signals such as PR1 secreted from pathogenic fungi, as well as endogenous danger signals generated in apoptosis-deficient mutants, lead to the activation of Psh and subsequent processing of Spz (Chamy et al., 2008; Ming et al., 2014; Obata et al., 2014). The active form of Spz induces conformational changes in the Toll receptor, activates Toll intracellular signaling (Kanoh et al., 2015b) and ultimately leads to the nuclear translocation of nuclear factor-kappa B (NF-κB) proteins Dif and Dorsal, inducing the expression of antimicrobial peptide genes including Drs (Lindsay and Wasserman, 2014). Conversely, the IMD pathway recognizes diaminopimelic acid-type peptidoglycans derived from Gram-negative bacteria via peptidoglycan recognition protein (PGRP)-LC and PGRP-LE (Kleino and Silverman, 2014). These receptors facilitate downstream signaling via the adaptor protein IMD, activate the NF-κB protein Relish, and induce the expression of antimicrobial peptides such as Diptericin (Dpt) (Paquette et al., 2010). Notably, these pathways are essentially characterized in Drosophila adults.
In contrast, Drosophila larvae have been largely utilized for dissecting cellular immune responses, particularly for nematode and wasp infections (Paddibhatla et al., 2010; Arefin et al., 2015; Kucerova et al., 2015; Hillyer, 2016). Insect hemocytes, representing blood cells, are composed of three cell types: plasmatocytes, crystal cells, and lamellocytes. These play central roles in cellular immunity by phagocytosing bacteria (plasmatocytes), involvement in the melanization process (crystal cells) and forming capsules around wasp eggs, a process referred to as encapsulation (lamellocytes) (Honti et al., 2014; Gold and Brückner, 2015; Parsons and Foley, 2016). For example, recent studies have begun to unravel the complex encapsulation processes by using Drosophila larvae upon infection with parasitoid wasps such as Leptopilina boulardi (Kari et al., 2016). In addition, the fat body, an immune-responsive organ in flies functionally resembling the mammalian liver, expresses edin and utilizes Toll signaling to control the numbers of plasmatocytes (Schmid et al., 2014; Vanha-aho et al., 2015). Finally, JAK-STAT signaling in somatic muscles is important for inducing the encapsulation reaction and controls the number of circulating lamellocytes (Yang et al., 2015).
By contrast, only a handful of studies have been published related to use of the Drosophila larval model of bacterial infection to analyze humoral immune responses (Ferrandon et al., 1998; Manfruelli et al., 1999; Ligoxygakis et al., 2002; Shia et al., 2009; Yamamoto-Hino et al., 2015; Yamamoto-Hino and Goto, 2016). Because these studies implicate intriguing differences in terms of the induction mechanisms of AMPs between larvae and adults, a larval model might thus have the potential to identify novel molecular mechanisms. However, it is possible that the limited numbers of publications on larval bacterial infection might partly be due to technical difficulties in the manufacture of uniform tungsten wires sharpened by electrolysis and their use in introducing infections (Romeo and Lemaitre, 2008) without causing severe damage that leads to the death of the larvae. Consistent with this likelihood, the survival and colony-forming assays upon systemic infection in larvae have been seldom reported. Here, we present a method to perform larval infection using a tungsten needle provided by a manufacturer that produces pins for testing semi-conductor devices. By using this uniform and solid needle, we were able to successfully perform and investigate bacterial infection in Drosophila larvae. In addition, we found that mechanical stimuli generated by pinching larvae with forceps resulted in the sterile induction of a antimicrobial peptide, providing a novel model for non-infectious activation of the humoral innate immune response.
RESULTS
The Toll pathway is required for survival against Gram-positive bacterial infection in larvae but not for bacterial removal
To easily and consistently perform infection using third instar larvae, we employed a fine tungsten needle used for the examination of semiconductor devices. With this needle, over 80% of larvae were able to survive following a clean injury in the wild type and in Toll pathway and IMD pathway mutants (Fig. 1A). By pricking larvae with a needle dipped into a pellet of Gram-positive bacteria Staphylococcus saprophyticus, we found that Toll pathway mutants were susceptible to the infection (Fig. 1B), although the number of bacteria in the infected whole mutant larvae after any time point was similar to that in the wild type (Fig. 1C). These results suggest that the Toll pathway is dispensable for bacterial clearance in larvae, showing a sharp contrast to the results from Drosophila adults in which the Toll pathway is required for the removal of bacteria upon Gram-positive bacterial challenge. Notably, although the induction of the antifungal peptide gene Drs was slightly lower in Toll pathway mutants than in wild-type larvae, substantial induction of Drs still remained in the mutants (Fig. 1D), consistent with the results of Manfruelli et al. (1999).
Fig. 1.

Systemic infection in Drosophila larvae by septic injury with a fine tungsten needle. (A) Survival analysis of larvae upon clean injury. Oregon R wild-type larvae, wild-type control y w, Toll pathway mutant Difnmc, psh and modSPKO double mutant, and the IMD pathway mutant relishE20 were used. (B) Survival analysis of larvae upon septic injury with S. saprophyticus. Larvae of Oregon R, Difnmc, modSPKO and imd1 mutant were used. (C) Colony forming unit (CFU) assay before (0 h) and after septic injury with S. saprophyticus at the indicated time points. Larvae of y w, Difnmc, and psh1 and modSPKO double mutant were used. (D) Real-time qPCR analysis of antimicrobial peptide Drs expression upon septic injury with S. saprophyticus at the indicated time points with larvae of Oregon R, Difnmc, and the psh1 and modSPKO double mutant. (E) Survival analysis of larvae upon septic injury with Ecc15. Larvae of Oregon R, Difnmc, modSPKO and imd1 mutants were used. Each survival curve is representative of at least two independent experiments of 60 larvae each (A,B,E). P-values were calculated using the log-rank test. (F) Real-time qPCR analysis of antimicrobial peptide Dpt expression upon septic injury with Ecc15 at the indicated time points with larvae of Oregon R, Difnmc, and imd1 mutants. Data are representative of more than two independent experiments performed in 20 larvae (C,D,F) (*P<0.05).
We next challenged larvae with Gram-negative bacteria using the needle. Fig. 1E and F show that IMD mutant larvae were not sensitive to infection with Ecc15, although the induction of the antibacterial peptide gene Dpt was almost abrogated in the mutant. From these results, we conclude that survival, AMP expression and bacterial number upon bacterial infection by septic injury with a tungsten needle could be consistently measured in Drosophila larvae, and that the role of the Toll pathway was somewhat different during this process compared with the adult infection model.
Pinching by forceps induces the expression of AMP in larvae
We found that clean injury with the needle induced the expression of Drs and Dpt (Fig. 2A,B). Furthermore, even pinching larvae using forceps, a normal means of handling larvae, caused strong Drs induction (Fig. 2A). Time-course experiments showed that Drs expression was induced from 2 h, maximized at 4 h and continued to 12 h (Fig. 2C). The level of Drs after eclosion was not increased compared with the level in untreated flies (Fig S4A). After pinching, 10% of larvae showed small melanized spots (Fig. 2D), although extremely weakly pinched larvae did not show melanization and the level of Drs induction was marginal (Fig. S4B), implying that pinching might cause minor injury in larval tissues. Next, we examined which tissues exhibited Drs expression. Fig. 2E shows that Drs reporter larvae exhibited GFP signals in the whole fat body and that the position of pinching was not connected with Drs induction. Consistent with this result, quantitative real-time-polymerase chain reaction (real-time qPCR) analysis showed that the induction of Drs was detected in the fat body dissected out from other tissues (Fig. 2F). These results indicate that Drs is induced in the fat body upon pinching with forceps.
Fig. 2.

Characterization of antimicrobial peptide induction following pinching larvae with forceps. (A-C) Real-time qPCR analysis of Drs expression (A,C) or Dpt expression (B) upon clean injury or pinching larvae with forceps at the indicated time points with larvae of Oregon R, w1118 and y w. Pinching was performed with larvae of w1118 in C. (D) Melanization spots after pinching larvae with forceps after 4 h. The indicated magnification of the objective lens was used; arrowheads indicate melanization spots. The right bar graph shows the percentage of larvae that exhibited melanization spots. Data were analyzed by Student's t-test and values represent the means±s.e. of three independent experiments with 30 larvae each. (E) Drs-GFP reporter analysis using a fluorescence stereomicroscope upon pinching larvae with forceps after 4 h. Larvae of Oregon R, Drs-GFP Dpt-lacZ (DD1); c564-GAL4, DD1; Dif1 and DD1; Difnmc were used. Brightfield (top row) and fluorescence images (bottom 2 rows) of single larva or multiple larvae (bottom row); the green signal indicates GFP fluorescence. (F) Real-time qPCR analysis of Drs expression following pinching of Oregon R, w1118, and y w larvae with forceps at the indicated time points. The larval fat body was dissected out from other tissues. (G,H) Real-time qPCR analysis of bacterial genomic DNA coding for 16S rRNA, which was normalized by the Drosophila genomic region for 12S RNA from conventionally reared or germ-free Oregon R larvae before pinching (G), or of Drs expression following pinching larvae with forceps at 4 h with conventionally reared or germ-free Oregon R larvae (H). Data were analyzed by the Student's t-test and values represent the means±s.e. of three independent experiments with 10 larvae each (F,G). *P<0.05.
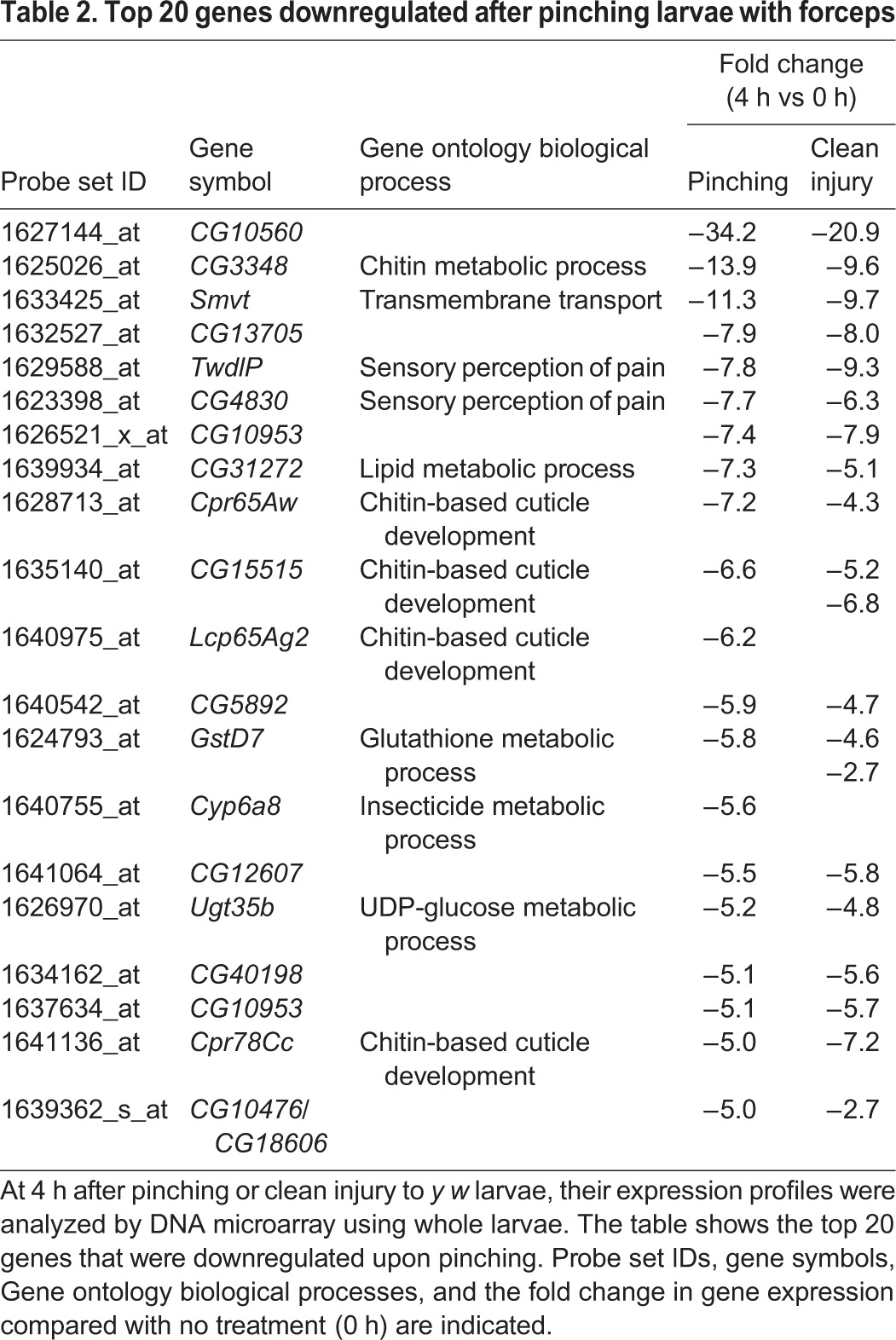
As Drosophila possess commensal bacteria (Kuraishi et al., 2013), Drs induction by pinching might be caused by such infections. To assess this possibility, germ-free larvae (Fig. 2G) were used for pinching experiments. Fig. 2H shows that the level of induction of Drs in germ-free larvae was not reduced compared with that in conventionally reared larvae, indicating that Drs expression is sterilely induced by pinching with forceps. Next, we performed microarray analysis using pinched larvae in order to examine whether Drs was uniquely induced by pinching or whether other defense response genes that respond to infection in adults (De Gregorio et al., 2001, 2002) were also induced. We found that in addition to Drs, several immune-related genes such as IM1, IM3, IM10 and Attacin were induced over 10-fold upon pinching with forceps (Table 1). In addition, stress responsive genes such as TotA, TotB and TotC were induced in the larvae. This result suggests that pinching larvae with forceps induces a humoral innate immune response that is similar to that observed in systemic infection in adults. We also noticed that a number of chitin metabolic genes were also downregulated upon pinching (Table 2).
Table 1.
Top 20 genes upregulated after pinching larvae with forceps
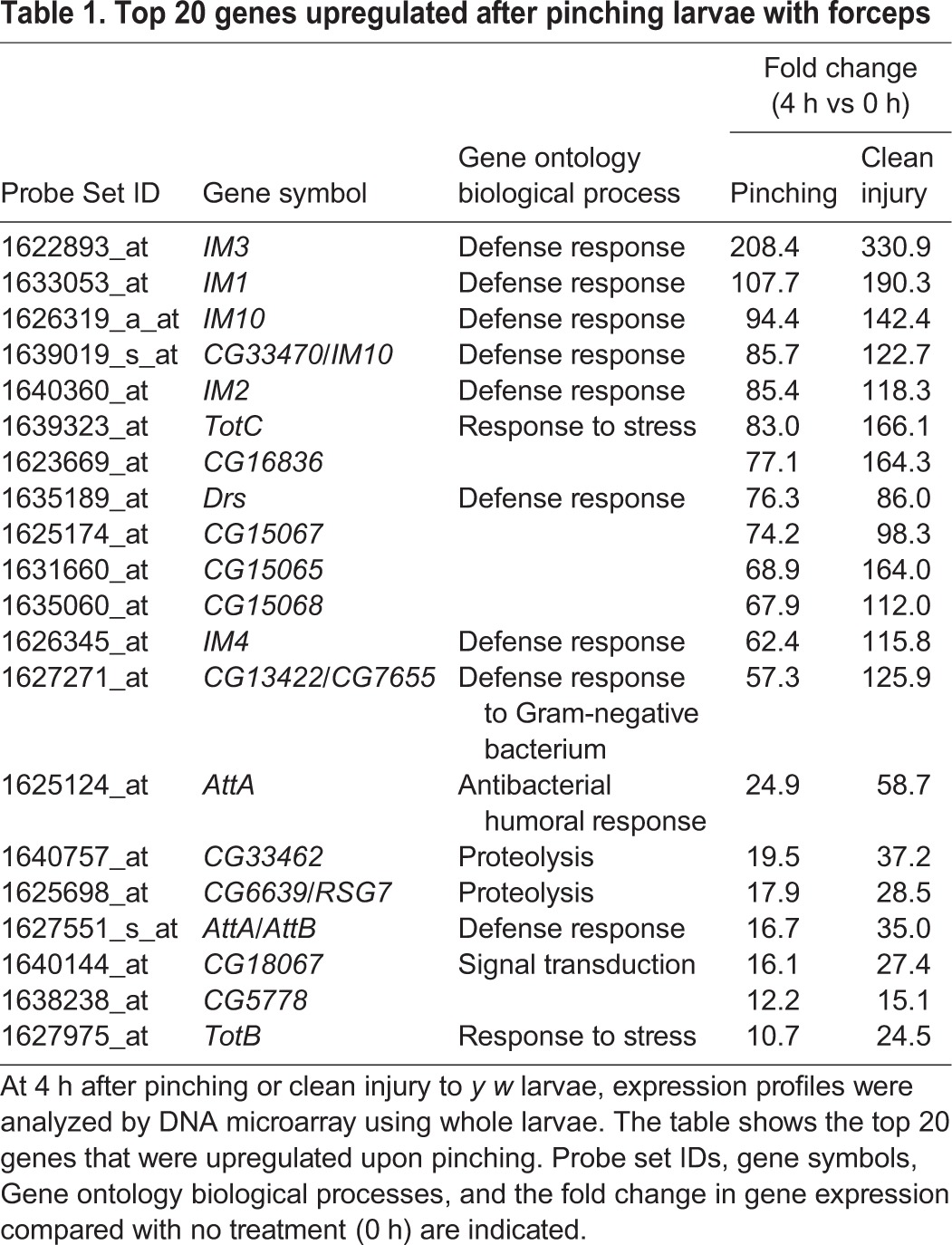
Table 2.
Top 20 genes downregulated after pinching larvae with forceps
Toll pathway genes contribute to the induction of Drs upon pinching with forceps
Next, we asked which signaling pathway is involved in the induction of Drs upon larval pinching. Real-time qPCR analysis showed that the level of induction in the spz mutant or dMyd88 mutant was approximately half that of the wild-type larvae (Fig. 3A). In contrast, the induction of Drs was comparable to that in the wild-type in the Dif mutant, or psh and modSP double mutants (Fig. 3A). These results suggest that certain Toll pathway components are partly required for the induction of Drs upon pinching. We next investigated IMD pathway mutants and found that larvae of the pgrp-le and pgrp-lc double mutant, imd mutant or relish mutant exhibited normal Drs induction after pinching (Fig. 3B). Furthermore, the level of induction of Drs in the double mutant larvae for imd and spz, or for relish and spz was almost the same as that in the spz single mutant (Fig. 3D), suggesting that the Toll and IMD pathways did not have a redundant role in pinching-induced Drs expression. We further investigated the involvement of the JAK-STAT, JNK, p38, dFOXO and pro-PO pathways, all of which suggested a role for AMP induction or host defense under certain conditions (Kim et al., 2002; Buchon et al., 2009a; Becker et al., 2010; Chen et al., 2010; Binggeli et al., 2014; Parisi et al., 2014). Inhibition of the JAK-STAT pathway by using an upd2 and upd3 double mutant did not reduce the induction of Drs in pinched larvae (Fig. 3C), implying that the JAK-STAT pathway may be dispensable for Drs induction. Similarly, the normal Drs induction observed upon pinching in larvae with an eiger mutation or c564-GAL4-driven expression of a dominant negative form of Bsk implied that there was no requirement of the JNK pathway in Drs induction (Fig. 3E). The level of Drs induction was also same in wild-type larvae as in the larvae of p38a, p38b and p38c (Fig. 3F), dfoxo (Fig. 3G) and PPO (Fig. 3H) mutants, indicating that the p38, dFOXO and pro-PO pathways played no role in the induction of Drs following larval pinching with forceps.
Fig. 3.

Drs induction in several mutant larvae upon pinching with forceps. (A-H) Real-time qPCR analysis of Drs expression after pinching larvae with forceps at 4 h with larvae of Oregon R, w1118, spzrm7, spzΔ8-1, psh1;;modSPKO, Difnmc, Dif1 and dMyd88kra1 (A); larvae of y w, pgrp-le and pgrp-lc double mutant (LE112;;LCΔ), imd1, and relishE20 (B); larvae of y w and upd2Δ upd3Δ (C); larvae of Oregon R, y w, imd1, spzrm7, imd1;spzrm7, reslihE20, and spzrm7 relishE20 (D); larvae of Oregon R, y w, bsk1/CyO, eiger1, GPF-IR, or bskDN driven by c564-GAL4 tubP-GAL80ts (E); larvae of y w, p38a13, p38b156A, p38c7B1, p38a13 p38b156A, and p38b156A/CyO ; p38c7B1 (F); larvae of y w, dFoxo21, and dFoxo21/dFoxow24 (G); and larvae of y w, PPO1Δ, PPO2Δ, and PPO1Δ PPO2Δ PPO3Δ (H). Data are representative of at least two independent experiments and were analyzed by Student's t-test; values represent the means±s.e. of triplicate samples with 10 larvae each. *P<0.05; n.s., not significant.
Sensory neurons and hemocytes are dispensable for the induction of Drs upon pinching larvae with forceps
When pinching larvae with forceps, we touched the larval cuticle under which the web of sensory neurons exists, prompting us to examine the role of sensory neurons in pinching-induced Drs expression. We first ablated sensory neurons by expressing the apoptosis-inducing genes reaper and hid with a pan-sensory neuron GAL4109(2)80 driver (Fig. 4A, Fig. S5) or class IV sensory neuron ppk-GAL4 driver. Fig. 4A shows that the level of induction of Drs in larvae with sensory neurons ablated by either driver was the same as that in the wild type. We next inhibited the function of sensory neurons with the same drivers by expressing temperature-sensitive Shibire (Kitamoto, 2001). However, no effect was again observed on Drs induction when neurotransmission was suppressed (Fig. 4B). Conversely, we then monitored Drs expression using larvae in which the sensory neurons were artificially activated by expressing the dTrpA1 ion channel (Hamada et al., 2008). Fig. 4C shows that Drs was not induced in the activated larvae without pinching. These results collectively suggest that sensory neurons are not involved in the induction of Drs upon larval pinching with forceps.
Fig. 4.

Sensory neurons and hemocytes are dispensable for the induction of Drs upon pinching. (A) Fluorescence microscopy (right) of larval sensory neurons around the mouth hook. Larvae of +/+; GAL4109(2)80 UAS-mCD8::GFP/+ and UAS-rpr UAS-hid/+; GAL4109(2)80 UAS-mCD8::GFP/+ were used. Green indicates the GFP signal and white dotted lines indicate the outlines of each larva. Scale bar: 100 µm. (A-C) Real-time qPCR analysis of Drs expression after pinching larvae with forceps for larvae of y w, UAS-rpr UAS-hid ; GAL4109(2)80 UAS-mCD8::GFP/+, UAS-rpr UAS-hid; ppk-GAL4/+ (second instar stage), UAS-rpr UAS-hid ;; ppk-GAL4/+ (third instar stage) at 0 or 4 h (A, left); larvae of UAS-lacZ/GAL4109(2)80 UAS-mCD8::GFP, UAS-Shits/GAL4109(2)80 UAS-mCD8::GFP, with or without pinching and with or without heat shock at 30°C for 4 h (hs) (B); larvae 4 h after treatment in UAS-lacZ/GAL4109(2)80 UAS-mCD8::GFP, UAS-Shits/GAL4109(2)80 UAS-mCD8::GFP, with or without pinching or heat shock treatment at 37°C for 2 min (hs) (C). Data are representative of at least two independent experiments and were analyzed by the Student's t-test; values represent the means±s.e. of triplicate samples with 10 larvae each. (D) Fluorescence microscopy observation of larval hemocytes. Larvae of hmlΔ-GAL4 UAS-2×EGFP/UAS-lacZ, and UAS-rpr UAS-hid/+; hmlΔ-GAL4 UAS-2×EGFP/+ were used. Green indicates the GFP signal and white dotted lines indicate the outlines of each larva. Scale bar: 200 µm. (E) Real-time qPCR analysis of Drs expression at 4 h after pinching larvae with forceps of hmlΔ-GAL4 UAS-2×EGFP/UAS-lacZ, UAS-rpr UAS-hid/+; hmlΔ-GAL4 UAS-2×EGFP/+, hmlΔ-GAL4 UAS-2×EGFP/+; UAS-Shits/+, with or without pinching or treatment at 29°C for 2 days (hs). Data are representative of at least two independent experiments and were analyzed by the Student's t-test; values represent the means±s.e. of triplicate samples with 10 larvae each.
As Spz has been suggested to be secreted from hemocytes (Shia et al., 2009), we tested the role of hemocytes in the induction of Drs upon pinching. We observed that hemocyte-specific expression of reaper and hid mediated by using a hmlΔ-GAL4 driver effectively ablated hemocytes in larvae (Fig. 4D). Using these larvae, we next examined the induction of Drs upon pinching and found that no difference could be detected between wild-type and hemocyte-ablated larvae with respect to the level of Drs induction (Fig. 4E). Consistent with this, inhibition of the phagocytic function of hemocytes by expressing Shibire (Awasaki and Ito, 2004) had no effect on the induction of Drs upon pinching (Fig. 4E). These results suggest that hemocytes are dispensable for the induction of Drs after pinching with forceps.
DISCUSSION
In this study, we present a method by which systemic bacterial infection can be performed easily and consistently in Drosophila larvae, thus providing another genetically tractable model to decipher infectious diseases. In this model, we show that the role of the Toll pathway in resistance against systemic infection in Drosophila larvae differs from that in adults to a certain extent. Specifically, the Toll pathway is likely to be required for tolerance (Ayres and Schneider, 2012) but not for resistance against Gram-positive bacteria, and exhibits partial involvement in the induction of AMPs; the latter being consistent with suggestions from a previous study (Manfruelli et al., 1999). Drs, an antimicrobial peptide whose expression is under the control of the Toll pathway, is strongly induced upon Gram-positive bacterial infection, although Drs is only active against fungi but not bacteria (Fehlbaum et al., 1994). One possibility to explain the former discrepancy might be that certain genes induced upon infection, including Drs, might function in conjunction to confer resistance and tolerance to adults and larvae, albeit with as-yet unknown mechanisms. This point should be elucidated in future research.
While performing these infection studies, we serendipitously found that the humoral innate immune response is activated in Drosophila larvae by modest mechanical stimuli; i.e. by pinching larvae with forceps, as they are commonly handled. AMP expression induced by pinching in larvae is sterile and partially independent from known innate immune signaling; these conclusions are supported by the following evidence: (1) the induction of Drs was observed in germ-free larvae upon pinching; (2) substantial induction of Drs remained in double mutants for the Toll and IMD pathways; (3) normal induction of Drs occurred after pinching in p38, JNK, JAK-STAT, dFOXO and pro-PO pathway mutants; and (4) hemocytes were dispensable for the induction of Drs upon pinching. These observations support the assertion that Drosophila larvae possess a novel mode of induction of the humoral innate immune response that might represent a good model for studying the mechanisms underlying sterile inflammation. Although we demonstrated that Dif, a Drosophila NF-κB essential for the induction of Drs upon systemic infection in adults, was not involved in pinching-induced Drs expression in larvae, we could not rule out the possible involvement of NF-κB in transactivating Drs expression, as we were unable to examine the redundant role of the other NF-κB proteins, Dorsal and Relish, because of the unavailability of viable lines. The dependency on NF-κB remains a question to be solved in future studies. Furthermore, we were unable to identify the essential genes required for the induction of Drs upon pinching. Unbiased genetic screening might therefore be necessary to unravel the molecular mechanism underlying this phenomenon.
Sterile inflammation is believed to contribute to many pathological conditions such as chronic inflammatory diseases including cancer (Rock et al., 2010). In Drosophila, several studies have established a sterile inflammation model in larvae (Shaukat et al., 2015). Ming et al. (2014) established a larval model for sterile AMP induction using a caspase mutant. They showed that the induction of Drs is solely dependent on Spz and Persephone, suggesting that the molecular mechanism of Drs induction in this mutant is different from our ‘pinching’ model. In addition, Hauling et al. (2014) and Parisi et al. (2014) reported that tumors can induce the expression of AMPs. Parisi et al. (2014) demonstrated that the induction of Drs is dependent on eiger and spz, both of which are not essential for our pinching-induced expression of Drs. Furthermore, Kanoh et al. (2015a) showed that Drosophila larvae possess another intrinsic ligand for the Toll receptor in addition to Spz, although its molecular nature has not yet been identified. Together, these reports suggest that Drosophila larvae possess multiple modes of induction of AMPs in response to various sterile stimuli that activate innate immunity.
In the current study, we showed that pinching stimuli can induce AMP expression; however, the physiological relevance of this phenomenon has not yet been elucidated. The larvae of Drosophila melanogaster in the wild are expected to be exposed to serious likelihood of attack by parasitoid wasps. Thus, mechanical stimuli might be considered as a potential infectious danger, suggesting that even small injuries resulting from oviposition might be able to activate AMP expression. Consistent with this, Schmid et al. (2014) recently showed that overactivation of Toll signaling could provoke a cellular immune defense that has potential importance in the response to wasp infection.
In conclusion, we demonstrate in this study that Drosophila larvae represent a suitable model in which to perform microbial infection by using a fine and uniform tungsten needle and to assess sterile induction of the humoral immune response by pinching larvae with forceps. In particular, because pinching-induced AMP expression is likely to be dependent on an as-yet uncharacterized molecular mechanism, our model might be useful to decipher the complex mechanisms that regulate sterile inflammation, which has considerable importance for the treatment of inflammatory diseases in humans such as ischemia, atherosclerosis and cancer.
MATERIALS AND METHODS
Fly stocks and maintenance
Drosophila stocks were maintained in standard corn meal-yeast agar medium vials at 25°C. Oregon R, w1118, and y w flies were used as wild-type controls. As Toll pathway mutants, spzrm7 (Morisato and Anderson, 1994), spzΔ8-1 (described below), modSPKO, psh1;;modSPKO (a gift from Dr Bruno Lemaitre) (Chamy et al., 2008; Buchon et al., 2009b), forR Difnmc (described below), Dif1 (a gift from Drs Jean-Marc Reichhart and Dominique Ferrandon) (Rutschmann et al., 2000) and dMyd88[kra1] (a gift from Dr Jean-Luc Imler) (Charatsi et al., 2003) were used. As IMD pathway mutants, PGRP-LE112;;PGRP-LCE12 (Gao et al., 1999; Takehana et al., 2004), imd1 (a gift from Dr. Bruno Lemaitre) and RelishE20 (Lemaitre et al., 1995; Hedengren et al., 1999) were used. The following mutants were used: bsk1 [Bloomington Drosophila Stock Center (BDSC), 3088], eiger1 (a gift from Dr Masayuki Miura) (Igaki et al., 2002), upd2Δ upd3Δ (a gift from Dr Bruno Lemaitre) (Osman et al., 2013), PPO1Δ PPO2Δ PPO3Δ (a gift from Dr Bruno Lemaitre) (Binggeli et al., 2014; Dudzic et al., 2015), dFoxo21 and dFoxow24 (a gift from Dr Marc Tatar) (Jünger et al., 2003; Weber et al., 2005), as well as p3813, p38b156A and p38c7B1 (a gift from Dr Bruno Lemaitre) (Davis et al., 2008; Chen et al., 2010; Chakrabarti, et al., 2014). The following transgenic flies were used: Drs-GFP Dpt-lacZ (a gift from Dr Dominique Ferrandon) (Jung et al., 2001), c564-GAL4, GAL4109(2)80 UAS-mCD8::GFP (a gift from Dr Tadashi Uemura) (Gao et al., 1999), ppk-GAL4 on 2nd instar stage (BDSC, 32078), ppk-GAL4 on 3rd instar stage (BDSC, 32079), hmlΔ-GAL4 UAS-2×EGFP (Sinenko and Mathey-Prevot, 2004), UAS-lacZ (a gift from Dr Manabu Ote), UAS-shits (a gift from Dr Takeshi Awasaki) (Kitamoto, 2001), UAS-rpr UAS-hid (a gift from Dr Shigeo Hayashi) (Zhou et al., 1997), UAS-dTrpA1 (a gift from Dr Paul Garrity) (Hamada et al., 2008), tubP-GAL80ts (BDSC), UAS-GFP-IR (BDSC, 9330), and UAS-bskDN (BDSC, 6409).
Difnmc allele
When using an allele of the foraging (for) gene, forR (a gift from Dr Shireen-Anne Davies) (Osborne et al., 1997) we happened to find that the homozygous flies of forR were susceptible to systemic infection with S. saprophyticus. However, trans-heterozygotes of the deficiency line that covers the for locus, forR/Df(2L)ED243, were not sensitive to the infection (Fig. S1A). Consistent with this, Drs induction upon S. saprophyticus in adults of forR homozygotes were severely impaired, although forR/Df(2L)ED243 trans-heterozygotes were perfectly normal (Fig. S1B). These results suggest that our forR line contains a second point mutation that is involved in the Toll pathway. Therefore, we named this mutation ‘nmc’ (natural immunity-mediating component) and performed mapping with several deficiency lines on the second chromosome. We found that deficiencies that covered the Dif and Dorsal locus, such as Df(2L)ED1161 or Df(J4), could not recuperate the immune phenotype of forR (Fig. S1C). Because Dorsal is reported not to be required for Drs induction in adults upon infection, we investigated the Dif locus by PCR. We found that our forR line contains foreign sequence of approximately 5 kb, possibly representing an accidental transposable element insertion at the third exon of Dif-RC (Fig. S1D and E), using several primers as shown in Fig. S1D or as follows: Dif-genome-PCR-F1-6: 5′-CGT ATC CAC TCC ACC AG-3′; Dif-genome-PCR-F1-7: 5′-ATT GGA AGG GTA GAC ACA TT-3′; Dif-genome-PCR-F1-8: 5′-GGT AGG ACT ACA GCC GTT TA-3′; Dif-genome-PCR-F1-9: 5′-ATG CCA AAC CCT TCC-3′; Dif-genome-PCR-F1-10: 5′-AGA TTG CCG ACC TTA AGA C-3′; Dif-genome-PCR-F1-11: 5′-GAA GGA GGT TGA ATC TCG-3′; Dif-genome-PCR-F1-12: 5′-TTC ATG GGT TCA TCT CAG T-3′; and Dif-genome-PCR-R1-6: 5′-AGC CTA AGC TCC AAT AGA ACT-3′. These results indicate that nmc represents a serendipitous mutation of the Dif gene. As the induction levels of Drs upon systemic infection with S. saprophyticus in forR hemocytes and forR/Df(2L)ED1161 are comparable (Fig. 1C), the Difnmc allele might be considered as a null or strong hypomorphic allele. We used this forR Difnmc line as the Dif mutant line in this study (note that forR Difnmc is denoted Difnmc in this manuscript).
spzΔ8-1 and DrsΔ7-17 mutants
spz or Drs mutants were generated using the CRISPR/Cas9 system as described in Kondo and Ueda (2013). A double-gRNA vector was constructed using pBFv-U6.2, pBFv-U6.2B, and the following primers: spz_in1_1_F: 5′-CTT CGT GCT TGT CTT AAG AAG ACA-3′; spz_in1_1_R: 5′-AAA CTG TCT TCT TAA GAC AAG CAC-3′; spz_ex1_1_F: 5′-CTT CGC AGG TGA TTG GCG GAT CGG-3′; spz_ex1_1_R: 5′-AAA CCC GAT CCG CCA ATC ACC TGC-3′; Drs-int1-1-F; 5′-CTT CGA AAA GGT TCT CAC GGA GCT-3′; Drs-int1-1-R: 5′-AAA CAG CTC CGT GAG AAC CTT TTC-3′; Drs-ex1-1-F: 5′-CTT CGC AGC CCC AGT CTG AAG TGC-3′; Drs-ex1-1-R: 5′-AAA CGC ACT TCA GAC TGG GGC TGC-3′. The constructed vector was used to generate the U6-spz-gRNA line, by using the y1 w67c23; P{CaryP}attP2 (BDSC, 8622) line (performed in BestGene). The U6-spz-gRNA line was crossed to nos-Cas9 (National Institute of Genetics, CAS-0001) as described in Kondo (2014) to generate candidate deletion lines. Genomic DNAs of each candidate mutant were screened by PCR to check for the deletion (Fig. S2) using the following primes: spz-Fw: 5′-GGA ACT GCT AGA ACA ACT ATG GA-3′; spz-Rv: 5′-CAG TAA CAC CAG CTA CCA GT-3′; Drs-Fw: 5′-GTG ACT GCA CAT GTA TCA TCA TAA TTT G-3′; and Drs-Rv: 5′-GTA GGT CGG GAA CAT TAG GG-3′. One line, spzΔ8-1, was found to harbor a 350 bp deletion that includes the start codon (Fig. S2); in addition, the line, DrsΔ7-17 was found to have a 210 bp deletion that includes the start codon (Fig. S2); thus, we used these lines as spz or Drs null mutants, respectively.
To provoke neural activity, ppk-GAL4 or GAL4109(2)80 UAS-mCD8::GFP was crossed to UAS-dTrpA1 and maintained at 18°C until they had developed into third instar larvae. In ppk-GAL4, third instar larvae were incubated at 29°C prior to assessment. In GAL4109(2)80, third instar larvae were incubated in a water bath at 37°C twice for 2 min (10 min intervals at 25°C), then maintained at 25°C for 4 h and used for assays.
To inhibit neural activity, GAL4109(2)80 UAS-mCD8: GFP was crossed to UAS-shits and maintained at 18°C. Third instar larvae were incubated in a water bath at 32°C for 5 min and experiments were performed at 30°C with warmed equipment. Pinched larvae were moved to agar plates and maintained at 25°C.
To remove sensory neurons or hemocytes, ppk-GAL4, GAL4109(2)80 UAS-mCD8::GFP, or hmlΔ-GAL4 UAS-2×EGFP were crossed to UAS-rpr UAS-hid and maintained at 18°C. Third instar larvae were incubated at 29°C for two days to induce apoptosis and observed under a stereo fluorescence microscope (M205FA, Leica, Wetzlar, Germany) to check the decrease of GFP signal, or used for the assays.
To inhibit exocytosis and phagocytic activity of hemocytes, hmlΔ-GAL4 UAS-2×EGFP was crossed to UAS-shits and maintained at 18°C. Third instar larvae were incubated at 29°C for two days, the experiments were performed at 25°C, and the pinched larvae were soon moved to agar plates and maintained at 29°C.
Microbial infection and pinching
The following pathogens were used for infection: Ecc15 (IFO3830) and S. saprophyticus (GTC0205). For larval infection, overnight bacterial cultures were concentrated by centrifugation, the pellet was washed with phosphate-buffered saline (PBS), and the larvae were then placed on a cold agar plate and pricked with a fine tungsten needle until complete penetration was achieved (Seimi, Sendai, Japan; total length: 43.5 mm; diameter: 0.2 mm; taper length: 2.5 mm) (Fig. S3A) that had been dipped in a pellet of concentrated bacteria (Fig. S3B) and moved to sealed Petri dishes containing apple juice agar. The needle was frequently changed before it became dull.
To monitor survival, 60 larvae of each genotype were incubated at 29°C after infection and the surviving larvae were every 2 h during transfer to fresh apple juice plates.
To assess the bacterial load in larvae, a colony-forming unit (CFU) assay was performed. Larvae were collected and their surfaces were sterilized with 70% ethanol. At total of 20 larvae of each genotype were homogenized in 500 μl nutrient broth (NB) bacterial medium, serially diluted, and plated onto NB medium plates.
For pinching larvae, MilliQ water was poured into Drosophila vials and the water and larvae were moved to Petri dishes. The middle part of third instar larvae were gently (0.2-0.25 MPa, Prescale, Fujifilm, Tokyo, Japan) pinched by forceps (Dumont, 0108-5-PO) for about 1 s (Fig. S3C), and then the larvae were moved to sealed Petri dishes containing apple juice agar. Melanization spots and GFP signals in larvae after pinching were observed using a fluorescent stereo microscope.
Rearing the axenic fly line
To obtain germ-free larvae, embryos were washed with bleach as described in Broderick et al. (2014). Briefly, embryos were rinsed in 70% ethanol for 1 min, placed in a 2.5% solution of sodium hypochlorite for 2 min, and then washed with 70% ethanol for 2 min. Embryos were then rinsed in sterile MilliQ water. Embryos were transferred to sterile foods and developed to larvae.
To check the axenic state, bacterial DNA was extracted from whole larvae and assessed by real-time qPCR using 16S rRNA primers (Suau et al., 1999).
Total RNA isolation, real-time qPCR, and microarray analysis
Larvae infected with bacteria or pinched by forceps were collected. Total RNA (1 µg), isolated from around 10 larvae using TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA), was used for cDNA synthesis with ReverTra Ace reverse transcriptase (Toyobo Ltd., Osaka, Japan) and oligo (dT) 15 primers (Promega, Madison, WI, USA). Using first-strand cDNA (0.5 μl), real-time qPCR was performed using a LightCycler (Roche Diagnostics, Roswell, GA, USA). rpL32 was used as the internal control. The following primers were used for real-time qPCR (F=forward, R=reverse): rpL32: 5′-AGA TCG TGA AGA AGC GCA CCA AG-3′ (F), 5′-CAC CAG GAA CTT CTT GAA TCC GG-3′ (R); Drs: 5′-TTG TTC GCC CTC TTC GCT GTC CT-3′ (F), 5′-GCA TCC TTC GCA CCA GCA CTT CA-3′ (R); Dpt: 5′-GTT CAC CAT TGC CGT CGC CTT AC-3′ (F), 5′-CCA AGT GCT GTC CAT ATC CTC C-3′ (R); 12S rRNA: 5'-TGG CGG TAT TTT AGT CTA TCT AGA GG-3′ (F), 5′-TAA GCT ACA CCT TGA TCT GA-3′ (R); and 16S rRNA: 5′-CAG GAT TAG ATA CGG TGG TAG T-3′ (F), 5′-TAA CCA CAT GCT CCA CCG CTT-3′ (R).
For microarray analysis, total RNA from Drosophila larvae homogenized in TRIzol was isolated using an RNeasy kit (Qiagen, Venlo, The Netherlands). The RNA quality was checked using an Agilent Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA). Total RNA (1 μg) was amplified and labeled as complementary RNA (cRNA) using an IVT Labeling Kit (Affymetrix, Santa Clara, CA, USA). Affymetrix Drosophila Genome 2.0 arrays were hybridized with 30 μg labeled cRNA, washed, stained and scanned (Goto et al., 2010). Data were analyzed by R software (https://www.r-project.org/).
Statistical analysis
Statistical analyses were performed using the Student's t-test or log-rank test, and P<0.05 was considered significant.
Acknowledgements
We are grateful to Drs Bruno Lemaitre, Jean-Marc Reichhart, Dominique Ferrandon, Masayuki Miura, Marc Tatar, Tadashi Uemura, Manabu Ote, Takeshi Awasaki, Shigeo Hayashi and Shireen-Anne Davies, as well as the Bloomington Stock Center, the Genetic Strain Research Center of National Institute of Genetics, and the Vienna Drosophila RNAi Center for fly stocks. We thank Drs Kenya Honda, Koji Atarashi, Hirotaka Kanoh, Nichole Broderick, Takeshi Awasaki, Hirofumi Furuhashi and Masayuki Miura for discussions and suggestions. We thank Fumi Shishido and Ryo Watanabe for technical support. We would also like to thank Editage (www.editage.jp) for English language editing.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
H.K., A.H., and T.K. conceived this study; H.K, A.H., and T.K. designed the experiments; A.H. and T.K. found that pinching larvae with forceps induced Drosomycin expression; A.H. and T.K. performed the experiments in Figs 1 and 2; H.K. performed the experiments in Figs 2-4; H.K., A.H., and T.K. analyzed the data; T.K. wrote the draft; H.K. and A.H. prepared the figures; all authors finalized the manuscript; T.K. led the entire project; and S.K. oversaw the study.
Funding
This work was supported by the Japan Science and Technology Agency (JST) of PRESTO ‘Chronic Inflammation’, and by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (MEXT).
Data availability
Microarray data have been deposited in GEO under accession number GSE94668 (available at: www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE94668).
Supplementary information
Supplementary information available online at http://dmm.biologists.org/lookup/doi/10.1242/dmm.027102.supplemental
References
- Anderson K. V., Jürgens G. and Nüsslein-Volhard C. (1985a). Establishment of dorsal-ventral polarity in the Drosophila embryo: genetic studies on the role of the Toll gene product. Cell 42, 779-789. 10.1016/0092-8674(85)90274-0 [DOI] [PubMed] [Google Scholar]
- Anderson K. V., Bokla L. and Nüsslein-Volhard C. (1985b). Establishment of dorsal-ventral polarity in the Drosophila embryo: the induction of polarity by the Toll gene product. Cell 42, 791-798. 10.1016/0092-8674(85)90275-2 [DOI] [PubMed] [Google Scholar]
- Arefin B., Kucerova L., Krautz R., Kranenburg H., Parvin F. and Theopold U. (2015). Apoptosis in hemocytes induces a shift in effector mechanisms in the Drosophila immune system and leads to a pro-inflammatory state . PLoS ONE 10, e0136593 10.1371/journal.pone.0136593 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Awasaki T. and Ito K. (2004). Engulfing action of glial cells is required for programmed axon pruning during Drosophila metamorphosis. Curr. Biol. 14, 668-677. 10.1016/j.cub.2004.04.001 [DOI] [PubMed] [Google Scholar]
- Ayres J. S. and Schneider D. S. (2012). Tolerance of infections. Annu. Rev. Immunol. 30, 271-294. 10.1146/annurev-immunol-020711-075030 [DOI] [PubMed] [Google Scholar]
- Becker T., Loch G., Beyer M., Zinke I., Aschenbrenner A. C., Carrera P., Inhester T., Schultze J. L. and Hoch M. (2010). FOXO-dependent regulation of innate immune homeostasis. Nature 463, 369-373. 10.1038/nature08698 [DOI] [PubMed] [Google Scholar]
- Binggeli O., Neyen C., Poidevin M. and Lemaitre B. (2014). Prophenoloxidase activation is required for survival to microbial infections in Drosophila. PLoS Pathog. 10, e1004067 10.1371/journal.ppat.1004067 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Broderick N. A., Buchon N. and Lemaitre B. (2014). Microbiota-induced changes in Drosophila melanogaster host gene expression and gut morphology. MBio 5, e01117-e01114 10.1128/mBio.01117-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchon N., Broderick N. A., Poidevin M., Pradervand S. and Lemaitre B. (2009a). Drosophila intestinal response to bacterial infection: activation of host defense and stem cell proliferation. Cell Host Microbe 5, 200-211. 10.1016/j.chom.2009.01.003 [DOI] [PubMed] [Google Scholar]
- Buchon N., Poidevin M., Kwon H.-M., Guillou A., Sottas V., Lee B.-L. and Lemaitre B. (2009b). A single modular serine protease integrates signals from pattern-recognition receptors upstream of the Drosophila Toll pathway. Proc. Natl. Acad. Sci. USA 106, 12442-12447. 10.1073/pnas.0901924106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buchon N., Silverman N. and Cherry S. (2014). Immunity in Drosophila melanogaster -- from microbial recognition to whole-organism physiology. Nat. Rev. Immunol. 14, 796-810. 10.1038/nri3763 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chakrabarti S., Poidevin M. and Lemaitre B. (2014). The Drosophila MAPK p38c regulates oxidative stress and lipid homeostasis in the intestine. PLoS Genet. 10, e1004659 10.1371/journal.pgen.1004659 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chamy L. E., Leclerc V., Caldelari I. and Reichhart J.-M. (2008). Sensing of ‘danger signals’ and pathogen-associated molecular patterns defines binary signaling pathways “upstream” of Toll. Nat. Immunol. 9, 1165-1170. 10.1038/ni.1643 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charatsi I., Luschnig S., Bartoszewski S., Nüsslein-Volhard C. and Moussian B. (2003). Krapfen/dMyd88 is required for the establishment of dorsoventral pattern in the Drosophila embryo. Mech. Dev. 120, 219-226. 10.1016/S0925-4773(02)00410-0 [DOI] [PubMed] [Google Scholar]
- Chen J., Xie C., Tian L., Hong L., Wu X. and Han J. (2010). Participation of the p38 pathway in Drosophila host defense against pathogenic bacteria and fungi. Proc. Natl. Acad. Sci. USA 107, 20774-20779. 10.1073/pnas.1009223107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davis M. M., Primrose D. A. and Hodgetts R. B. (2008). A member of the p38 mitogen-activated protein kinase family is responsible for transcriptional induction of dopa decarboxylase in the epidermis of Drosophila melanogaster during the innate immune response. Mol. Cell.Biol. 28, 4883-4895. 10.1128/MCB.02074-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Gregorio E., Spellman P. T., Rubin G. M. and Lemaitre B. (2001). Genome-wide analysis of the Drosophila immune response by using oligonucleotide microarrays. Proc. Natl. Acad. Sci. USA 98, 12590-12595. 10.1073/pnas.221458698 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Gregorio E., Spellman P. T., Tzou P., Rubin G. M. and Lemaitre B. (2002). The Toll and Imd pathways are the major regulators of the immune response in Drosophila. EMBO J. 21, 2568-2579. 10.1093/emboj/21.11.2568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dudzic J. P., Kondo S., Ueda R., Bergman C. M. and Lemaitre B. (2015). Drosophila innate immunity: regional and functional specialization of prophenoloxidases. BMC Biol. 13, 81 10.1186/s12915-015-0193-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fehlbaum P., Bulet P., Michaut L., Lagueux M., Broekaert W. F., Hetru C. and Hoffmann J. A. (1994). Insect immunity. Septic injury of Drosophila induces the synthesis of a potent antifungal peptide with sequence homology to plant antifungal peptides. J. Biol. Chem. 269, 33159-33163. [PubMed] [Google Scholar]
- Ferrandon D., Jung A. C., Criqui M., Lemaitre B., Uttenweiler-Joseph S., Michaut L., Reichhart J. and Hoffmann J. A. (1998). A drosomycin-GFP reporter transgene reveals a local immune response in Drosophila that is not dependent on the Toll pathway. EMBO J. 17, 1217-1227. 10.1093/emboj/17.5.1217 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrandon D., Imler J.-L., Hetru C. and Hoffmann J. A. (2007). The Drosophila systemic immune response: sensing and signalling during bacterial and fungal infections. Nat. Rev. Immunol. 7, 862-874. 10.1038/nri2194 [DOI] [PubMed] [Google Scholar]
- Gao F.-B., Brenman J. E., Jan L. Y. and Jan Y. N. (1999). Genes regulating dendritic outgrowth, branching, and routing in Drosophila. Genes Dev. 13, 2549-2561. 10.1101/gad.13.19.2549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gobert V., Gottar M., Matskevich A. A., Rutschmann S., Royet J., Belvin M., Hoffman J. A. and Ferrandon D. (2003). Dual activation of the Drosophila toll pathway by two pattern recognition receptors. Science 302, 2126-2130. 10.1126/science.1085432 [DOI] [PubMed] [Google Scholar]
- Gold K. S. and Brückner K. (2015). Macrophages and cellular immunity in Drosophila melanogaster. Semin. Immunol. 27, 357-368. 10.1016/j.smim.2016.03.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goto A., Yano T., Terashima J., Iwashita S., Oshima Y. and Kurata S. (2010). Cooperative regulation of the induction of the novel antibacterial listericin by peptidoglycan recognition protein LE and the JAK-STAT pathway. J. Biol. Chem. 285, 15731-15738. 10.1074/jbc.M109.082115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gottar M., Gobert V., Michel T., Belvin M., Duyk G., Hoffmann J. A., Ferrandon D. and Royet J. (2002). The Drosophila immune response against Gram-negative bacteria is mediated by a peptidoglycan recognition protein. Nature 416, 640-644. 10.1038/nature734 [DOI] [PubMed] [Google Scholar]
- Gottar M., Gobert V., Matskevich A. A., Reichhart J.-M., Wang C., Butt T. M., Belvin M., Hoffman J. A. and Ferrandon D. (2006). Dual detection of fungal infections in Drosophila via recognition of glucans and sensing of virulence factors. Cell 127, 1425-1437. 10.1016/j.cell.2006.10.046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamada F. N., Rosenzweig M., Kang K., Pulver S. R., Ghezzi A., Jegla T. J. and Garrity P. A. (2008). An internal thermal sensor controlling temperature preference in Drosophila. Nature 454, 217-220. 10.1038/nature07001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hauling T., Krautz R., Markus R., Volkenhoff A., Kucerova L. and Theopold U. (2014). A Drosophila immune response against Ras-induced overgrowth. Biol. Open 3, 250-260. 10.1242/bio.20146494 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hedengren M., Åsling B., Dushay M. S., Ando I., Ekengren S., Wihlborg M. and Hultmark D. (1999). Relish, a central factor in the control of humoral but not cellular immunity in Drosophila. Mol. Cell 4, 827-837. 10.1016/S1097-2765(00)80392-5 [DOI] [PubMed] [Google Scholar]
- Hillyer J. F. (2016). Insect immunology and hematopoiesis. Dev. Comp. Immunol. 58, 102-118. 10.1016/j.dci.2015.12.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honti V., Csordás G., Kurucz É., Márkus R. and Andó I. (2014). The cell-mediated immunity of Drosophila melanogaster: Hemocyte lineages, immune compartments, microanatomy and regulation. Dev. Comp. Immunol. 42, 47-56. 10.1016/j.dci.2013.06.005 [DOI] [PubMed] [Google Scholar]
- Igaki T., Kanda H., Yamamoto-Goto Y., Kanuka H., Kuranaga E., Aigaki T. and Miura M. (2002). Eiger, a TNF superfamily ligand that triggers the Drosophila JNK pathway. EMBO J. 21, 3009-3018. 10.1093/emboj/cdf306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang I.-H., Chosa N., Kim S.-H., Nam H.-J., Lemaitre B., Ochiai M., Kambris Z., Brun S., Hashimoto C., Ashida M. et al. (2006). A Spätzle-processing enzyme required for toll signaling activation in Drosophila innate immunity. Dev. Cell 10, 45-55. 10.1016/j.devcel.2005.11.013 [DOI] [PubMed] [Google Scholar]
- Jung A. C., Criqui M. C., Rutschmann S., Hoffmann J. A. and Ferrandon D. (2001). Microfluorometer assay to measure the expression of beta-galactosidase and green fluorescent protein reporter genes in single Drosophila flies. BioTechniques 30, 594-598. [DOI] [PubMed] [Google Scholar]
- Jünger M. A., Rintelen F., Stocker H., Wasserman J. D., Végh M., Radimerski T., Greenberg M. E. and Hafen E. (2003). The Drosophila forkhead transcription factor FOXO mediates the reduction in cell number associated with reduced insulin signaling. J. Biol. 2, 20 10.1186/1475-4924-2-20 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanoh H., Kuraishi T., Tong L.-L., Watanabe R., Nagata S. and Kurata S. (2015a). Ex vivo genome-wide RNAi screening of the Drosophila Toll signaling pathway elicited by a larva-derived tissue extract. Biochem. Biophys. Res. Commun. 467, 400-406. 10.1016/j.bbrc.2015.09.138 [DOI] [PubMed] [Google Scholar]
- Kanoh H., Tong L.-L., Kuraishi T., Suda Y., Momiuchi Y., Shishido F. and Kurata S. (2015b). Genome-wide RNAi screening implicates the E3 ubiquitin ligase Sherpa in mediating innate immune signaling by Toll in Drosophila adults. Sci. Signal. 8, ra107 10.1126/scisignal.2005971 [DOI] [PubMed] [Google Scholar]
- Kari B., Csordás G., Honti V., Cinege G., Williams M. J., Andó I. and Kurucz É. (2016). The raspberry gene is involved in the regulation of the cellular immune response in Drosophila melanogaster. PLoS ONE 11, e0150910 10.1371/journal.pone.0150910 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim D. H., Feinbaum R., Alloing G., Emerson F. E., Garsin D. A., Inoue H., Tanaka-Hino M., Hisamoto N., Matsumoto K., Tan M. W. et al. (2002). A conserved p38 MAP kinase pathway in Caenorhabditis elegans innate immunity. Science 297, 623-626. 10.1126/science.1073759 [DOI] [PubMed] [Google Scholar]
- Kitamoto T. (2001). Conditional modification of behavior in Drosophila by targeted expression of a temperature-sensitive shibire allele in defined neurons. J. Neurobiol. 47, 81-92. 10.1002/neu.1018 [DOI] [PubMed] [Google Scholar]
- Kleino A. and Silverman N. (2014). The Drosophila IMD pathway in the activation of the humoral immune response. Dev. Comp. Immunol. 42, 25-35. 10.1016/j.dci.2013.05.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kondo S. (2014). New horizons in genome engineering of Drosophila melanogaster. Genes Genet. Syst. 89, 3-8. 10.1266/ggs.89.3 [DOI] [PubMed] [Google Scholar]
- Kondo S. and Ueda R. (2013). Highly improved gene targeting by germline-specific Cas9 expression in Drosophila. Genetics 195, 715-721. 10.1534/genetics.113.156737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kucerova L., Broz V., Arefin B., Maaroufi H. O., Hurychova J., Strnad H., Zurovec M. and Theopold U. (2015). The Drosophila chitinase-like protein IDGF3 is involved in protection against nematodes and in wound healing. J. Innate Immun. 8, 199-210. 10.1159/000442351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuraishi T., Hori A. and Kurata S. (2013). Host-microbe interactions in the gut of Drosophila melanogaster. Front. Physiol. 4, 375 10.3389/fphys.2013.00375 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemaitre B. and Hoffmann J. (2007). The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 25, 697-743. 10.1146/annurev.immunol.25.022106.141615 [DOI] [PubMed] [Google Scholar]
- Lemaitre B., Kromer-Metzger E., Michaut L., Nicolas E., Meister M., Georgel P., Reichhart J. M. and Hoffman J. A. (1995). A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defense. Proc. Natl. Acad. Sci. USA 92, 9465-9469. 10.1073/pnas.92.21.9465 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lemaitre B., Nicolas E., Michaut L., Reichhart J.-M. and Hoffmann J. A. (1996). The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 86, 973-983. 10.1016/S0092-8674(00)80172-5 [DOI] [PubMed] [Google Scholar]
- Ligoxygakis P., Bulet P. and Reichhart J.-M. (2002). Critical evaluation of the role of the Toll-like receptor 18-Wheeler in the host defense of Drosophila. EMBO Rep. 3, 666-673. 10.1093/embo-reports/kvf130 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lindsay S. A. and Wasserman S. A. (2014). Conventional and non-conventional Drosophila Toll signaling. Dev. Comp. Immunol. 42, 16-24. 10.1016/j.dci.2013.04.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manfruelli P., Reichhart J.-M., Steward R., Hoffmann J. A. and Lemaitre B. (1999). A mosaic analysis in Drosophila fat body cells of the control of antimicrobial peptide genes by the Rel proteins Dorsal and DIF. EMBO J. 18, 3380-3391. 10.1093/emboj/18.12.3380 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ming M., Obata F., Kuranaga E. and Miura M. (2014). Persephone/Spätzle pathogen sensors mediate the activation of Toll receptor signaling in response to endogenous danger signals in apoptosis-deficient Drosophila. J. Biol. Chem. 289, 7558-7568. 10.1074/jbc.M113.543884 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morisato D. and Anderson K. V. (1994). The spätzle gene encodes a component of the extracellular signaling pathway establishing the dorsal-ventral pattern of the Drosophila embryo. Cell 76, 677-688. 10.1016/0092-8674(94)90507-X [DOI] [PubMed] [Google Scholar]
- Myllymäki H., Valanne S. and Rämet M. (2014). The Drosophila imd signaling pathway. J. Immunol. 192, 3455-3462. 10.4049/jimmunol.1303309 [DOI] [PubMed] [Google Scholar]
- Obata F., Kuranaga E., Tomioka K., Ming M., Takeishi A., Chen C.-H., Soga T. and Miura M. (2014). Necrosis-driven systemic immune response alters SAM metabolism through the FOXO-GNMT Axis. Cell Rep. 7, 821-833. 10.1016/j.celrep.2014.03.046 [DOI] [PubMed] [Google Scholar]
- Osborne K. A., Robichon A., Burgess E., Butland S., Shaw R. A., Coulthard A., Pereira H. S., Greenspan R. J. and Sokolowski M. B. (1997). Natural behavior polymorphism due to a cGMP-dependent protein kinase of Drosophila. Science 277, 834-836. 10.1126/science.277.5327.834 [DOI] [PubMed] [Google Scholar]
- Osman D., Buchon N., Chakrabarti S., Huang Y.-T., Su W.-C., Poidevin M., Tsai Y. C. and Lemaitre B. (2013). Autocrine and paracrine unpaired signaling regulate intestinal stem cell maintenance and division. J. Cell Sci. 125, 5944-5949. 10.1242/jcs.113100 [DOI] [PubMed] [Google Scholar]
- Paddibhatla I., Lee M. J., Kalamarz M. E., Ferrarese R. and Govind S. (2010). Role for sumoylation in systemic inflammation and immune homeostasis in Drosophila larvae. PLoS Pathog. 6, e1001234 10.1371/journal.ppat.1001234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paquette N., Broemer M., Aggarwal K., Chen L., Husson M., Ertürk-Hasdemir D., Reichhart J.-M., Meier P. and Silverman N. (2010). Caspase-mediated cleavage, IAP binding, and ubiquitination: linking three mechanisms crucial for Drosophila NF-kappaB signaling. Mol. Cell 37, 172-182. 10.1016/j.molcel.2009.12.036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parisi F., Stefanatos R. K., Strathdee K., Yu Y. and Vidal M. (2014). Transformed epithelia trigger non-tissue-autonomous tumor suppressor response by adipocytes via activation of Toll and Eiger/TNF signaling. Cell Rep. 6, 855-867. 10.1016/j.celrep.2014.01.039 [DOI] [PubMed] [Google Scholar]
- Parsons B. and Foley E. (2016). Cellular immune defenses of Drosophila melanogaster. Dev. Comp. Immunol. 58, 95-101. 10.1016/j.dci.2015.12.019 [DOI] [PubMed] [Google Scholar]
- Rämet M. and Hultmark D. (2014). Drosophila immunity – glorious past, dynamic present and exciting future. Dev. Comp. Immunol. 42, 1-2. 10.1016/j.dci.2013.07.013 [DOI] [PubMed] [Google Scholar]
- Rock K. L., Latz E., Ontiveros F. and Kono H. (2010). The sterile inflammatory response. Ann. Rev. Immunol. 28, 321-342. 10.1146/annurev-immunol-030409-101311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Romeo Y. and Lemaitre B. (2008). Drosophila immunity: methods for monitoring the activity of Toll and Imd signaling pathways. Methods Mol. Biol. 415, 379-394. 10.1007/978-1-59745-570-1_22 [DOI] [PubMed] [Google Scholar]
- Rutschmann S., Jung A. C., Hetru C., Reichhart J.-M., Hoffmann J. A. and Ferrandon D. (2000). The Rel protein DIF mediates the antifungal but not the antibacterial host defense in Drosophila. Immunity 12, 569-580. 10.1016/S1074-7613(00)80208-3 [DOI] [PubMed] [Google Scholar]
- Schmid M. R., Anderl I., Vesala L., Vanha-aho L.-M., Deng X.-J., Rämet M. and Hultmark D. (2014). Control of Drosophila blood cell activation via Toll signaling in the fat body. PLoS ONE 9, e102568 10.1371/journal.pone.0102568 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaukat Z., Liu D. and Gregory S. (2015). Sterile inflammation in Drosophila. Mediators Inflamm. 2015, 369286 10.1155/2015/369286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shia A. K. H., Glittenberg M., Thompson G., Weber A. N., Reichhart J.-M. and Ligoxygakis P. (2009). Toll-dependent antimicrobial responses in Drosophila larval fat body require Spätzle secreted by haemocytes. J. Cell Sci. 122, 4505-4515. 10.1242/jcs.049155 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinenko S. A. and Mathey-Prevot B. (2004). Increased expression of Drosophila tetraspanin, Tsp68C, suppresses the abnormal proliferation of ytr-deficient and Ras/Raf-activated Oncogene 23, 9120-9128. 10.1038/sj.onc.1208156 [DOI] [PubMed] [Google Scholar]
- Suau A., Bonnet R., Sutren M., Godon J. J., Gibson G. R., Collins M. D. and Doré J. (1999). Direct analysis of genes encoding 16S rRNA from complex communities reveals many novel molecular species within the human gut. Appl. Environ. Microbiol. 65, 4799-4807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takehana A., Yano T., Mita S., Kotani A., Oshima Y. and Kurata S. (2004). Peptidoglycan recognition protein (PGRP)-LE and PGRP-LC act synergistically in Drosophila immunity. EMBO J. 23, 4690-4700. 10.1038/sj.emboj.7600466 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valanne S., Wang J.-H. and Rämet M. (2011). The Drosophila Toll signaling pathway. J. Immunol. 186, 649-656. 10.4049/jimmunol.1002302 [DOI] [PubMed] [Google Scholar]
- Vanha-aho L.-M., Anderl I., Vesala L., Hultmark D., Valanne S. and Rämet M. (2015). Edin expression in the fat body is required in the defense against parasitic wasps in Drosophila melanogaster. PLoS Pathog. 11, e1004895 10.1371/journal.ppat.1004895 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber K., Johnson N., Champlin D. and Patty A. (2005). Many P-element insertions affect wing shape in Drosophila melanogaster. Genetics 169, 1461-1475. 10.1534/genetics.104.027748 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamamoto-Hino M. and Goto S. (2016). Spätzle-processing enzyme-independent activation of the Toll pathway in Drosophila innate immunity. Cell Struct. Funct. 41, 55-60. 10.1247/csf.16002 [DOI] [PubMed] [Google Scholar]
- Yamamoto-Hino M., Muraoka M., Kondo S., Ueda R., Okano H. and Goto S. (2015). Dynamic regulation of innate immune responses in Drosophila by Senju-mediated glycosylation. Proc. Natl. Acad. Sci. USA 112, 5809-5814. 10.1073/pnas.1424514112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang H., Kronhamn J., Ekström J. O., Korkut G. G. and Hultmark D. (2015). JAK/STAT signaling in Drosophila muscles controls the cellular immune response against parasitoid infection. EMBO Rep. 16, 1664-1672. 10.15252/embr.201540277 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou L., Schnitzler A., Agapite J., Schwartz L. M., Steller H. and Nambu J. R. (1997). Cooperative functions of the reaper and head involution defective genes in the programmed cell death of Drosophila central nervous system midline cells. Proc. Natl. Acad. Sci. USA 94, 5131-5136. 10.1073/pnas.94.10.5131 [DOI] [PMC free article] [PubMed] [Google Scholar]