

Abstract
Krüppel-like factors (KLFs) are a family of zinc-finger transcription factors that are found in many species. Recent studies have shown that KLFs play a fundamental role in regulating diverse biological processes such as cell proliferation, differentiation, development and regeneration. Of note, several KLFs are also crucial for maintaining pluripotency and, hence, have been linked to reprogramming and regenerative medicine approaches. Here, we review the crucial functions of KLFs in mammalian embryogenesis, stem cell biology and regeneration, as revealed by studies of animal models. We also highlight how KLFs have been implicated in human diseases and outline potential avenues for future research.
KEY WORDS: Krüppel-like factors, Stem cells, Mammalian development, Cellular regeneration, Human disease
Summary: This Primer article provides an overview of Krüppel-like factors (KLFs), a family of zinc-finger transcription factors that play fundamental roles in development.
Introduction
Krüppel-like factors (KLFs) are evolutionarily conserved zinc-finger transcription factors that share homology with the Drosophila melanogaster protein Krüppel, which regulates body segmentation during fly embryogenesis (Preiss et al., 1985). Members of the KLF family also share structural homology and DNA-binding capacity with specificity proteins (SPs) and are thus often referred to as SP/KLF transcription factors. Recent studies have shown that specific KLFs are expressed across many species, spanning from single-celled to multicellular organisms (Presnell et al., 2015). Importantly, since their discovery, KLFs have been implicated in a variety of physiological processes, acting to regulate key cellular functions such as proliferation, differentiation and apoptosis. Furthermore, the identification of KLF4 as a crucial factor in reprogramming somatic cells into induced pluripotent cells has garnered great interest in KLFs in the stem cell biology and regeneration community.
Here, we describe the role of KLFs in mammalian development, highlighting how and when individual KLFs act during early embryogenesis and organogenesis. We also discuss the varied roles of KLFs in stem cell biology, reprogramming and regeneration, and review their roles in human diseases.
An introduction to KLFs: from structure to function
All KLFs possess three highly conserved C2H2 zinc finger domains in their carboxyl-terminal regions (Fig. 1) that mediate transcriptional activation and/or repression by interacting with GC-rich consensus, including 5′-CACCC-3′, DNA sequences (Lomberk and Urrutia, 2005; Suske et al., 2005). Conversely, the amino-terminal regions of KLFs are divergent and modulate the specificity of protein-protein and protein-DNA interactions (Nagai et al., 2009). Recently, comprehensive evolutionary studies of SP/KLF family members using zinc finger sequences as well as transactivation/repression domains for phylogenetic analysis confirmed the existence of this family in 48 species within Eukaryota (Presnell et al., 2015); the evolutionary relationships between KLFs of murine and human origin based on their zinc finger domains is outlined in Fig. 2
Fig. 1.
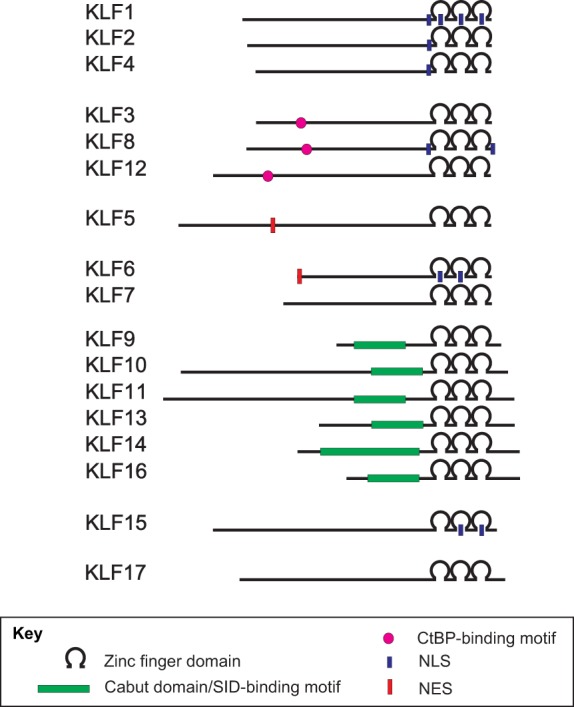
Schematic of the domain structure of human KLF proteins. All KLFs possess three highly conserved C2H2 zinc finger domains in their carboxyl-terminal regions that mediate transcriptional activation and/or repression. By contrast, their N-terminal regions are less conserved, harboring additional motifs, such as CtBP-motifs and Cabut domains/SID-binding motifs, that are implicated in protein-protein and protein-DNA interactions. Some KLFs also contain nuclear localization signals (NLSs) and nuclear export signals (NESs) that regulate their subcellular localization. Proteins are drawn to scale.
Fig. 2.

Phylogenetic relationship between human and mouse KLF family members. An amino acid alignment was produced from full-length mouse and human KLF family members using MAFFT (Katoh et al., 2002) employing the L-INS-i algorithm and BLOSUM45 scoring matrix. As KLF family members share little homology outside of the C-terminal ZNF region, the alignment was trimmed 22 residues N-terminal of the first ZNF domain and one residue C-terminal of the third ZNF domain. Phylogenetic analysis of the ZNF region was performed via RAxML 7.2.8 (Stamatakis, 2006) using the Gamma LG model, and node support in the ML tree was sampled via 100 bootstrap replicates. Both alignment and phylogenetic analyses were performed in Geneious version 8.1.8 (Kearse et al., 2012). Scale bar: 0.2 amino acid changes per site.
The specificity of KLF-mediated transcriptional activation is defined mostly by their N-terminal sequences. KLF1, 2, 4, 5, 6, 8 and 15 (the latter by homology only) possess a transactivation domain (TAD) within their N-terminal regions (Chen and Bieker, 1996; Conkright et al., 1999, 2001; Kojima et al., 1997; Koritschoner et al., 1997; Lahiri and Zhao, 2012; Mas et al., 2011; Ratziu et al., 1998; van Vliet et al., 2000; Wani et al., 1999a). KLF1 has a very well-defined and essential TAD in its first 100 amino acids (Chen and Bieker, 1996). It has been shown that the KLF1 TAD can be divided into two regions – TAD1 and TAD2 – and that the latter is also conserved in KLF2, 4, 5 and 15 (Mas et al., 2011). In addition, a repression domain adjacent to the activation domain has been identified in KLF2 and KLF4 (Conkright et al., 2001; Geiman et al., 2000; Wani et al., 1999a). Furthermore, two KLF subgroups have been identified based on their respective interacting partners – CtBP and Sin3: KLF3, 8 and 12 possess an N-terminal repression domain (Fig. 1) that contains a CtBP recognition motif (Lahiri and Zhao, 2012; Schuierer et al., 2001; Turner and Crossley, 1998; van Vliet et al., 2000), and KLF9, 10, 11, 13, 14 and 16 contain a Cabut domain in their N-terminal section (Fig. 1) that encompasses a Sin3 interaction domain (SID) and is referred to as a transcriptional regulatory repression domain (Cook et al., 1999; Daftary et al., 2012; Kaczynski et al., 2001, 2002; Lomberk et al., 2013; Truty et al., 2009; Zhang et al., 2001a). Nuclear localization signals (NLSs) have also been identified in several KLFs (Fig. 1). For example, KLF1 contains two NLSs: NLS1, which is adjacent to the first zinc finger domain, and NLS2, which spans through three zinc finger domains (Quadrini and Bieker, 2002). In comparison, KLF2 and KLF4 harbor NLSs adjacent to their first zinc finger domain (Shields and Yang, 1997), and KLF6 has a single NLS localized within its first and second zinc finger domains (Rodriguez et al., 2010). Furthermore, KLF15 has two NLSs located within its second and third zinc finger domains (Leenders et al., 2012), whereas the two NLSs of KLF8 flank the ends of its zinc finger domains (Lahiri and Zhao, 2012; Mehta et al., 2009). Finally, KLF7 has a putative NLS motif adjacent to the zinc finger domains (Matsumoto et al., 1998) and KLF13 possesses two putative NLSs, one adjacent to and one within its zinc finger domain (Song et al., 2002). Interestingly, two KLFs – KLF5 and KLF6 – also include nuclear export signal (NES) sequences (Fig. 1), which regulate their cellular localization (Du et al., 2008; Rodriguez et al., 2010). Although NLS or NES signals have not been identified within the KLF16 sequence, it has been shown in vitro that serum levels in the media regulate the nuclear-cytoplasm shuttling of KLF16 and that interactions with Sin3-HDAC complexes impact its chromatin localization (Daftary et al., 2012).
KLFs also display key differences in their temporal, spatial and cell-specific modes of expression, their post-translational modifications (phosphorylation, acetylation, methylation, SUMOlyation and ubiquitylation), as well as their interactions with co-activators or co-repressors, which add additional layers of specificity and governance over these factors (summarized in Table 1).
Table 1.
Pattern of expression, regulation and functions of KLFs

The role of KLFs in early embryogenesis
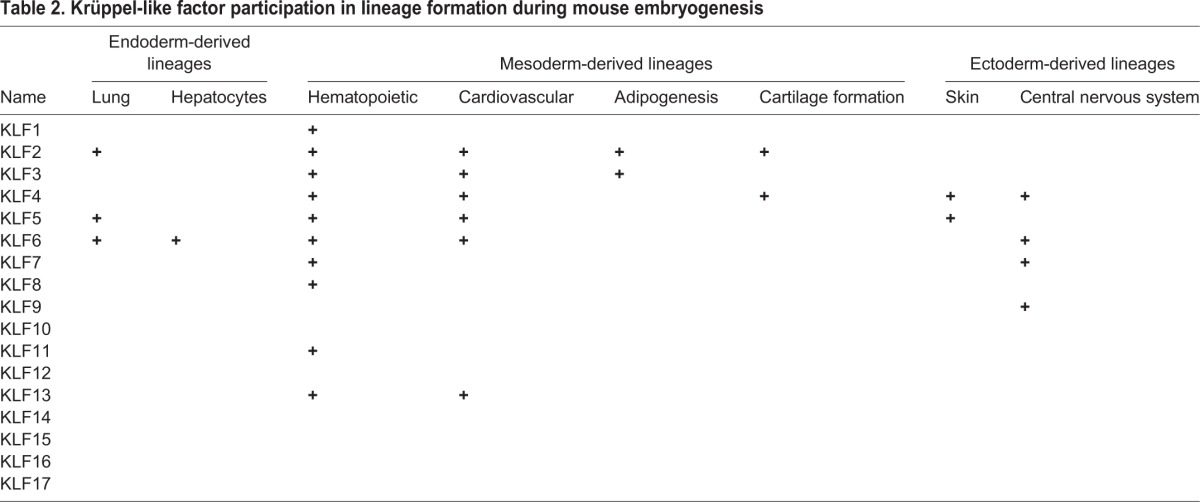
The early mammalian post-fertilization embryo undergoes cleavage divisions without growth, eventually giving rise to a structure known as a blastocyst, which contains a distinct inner cell mass (ICM) and an outer layer of trophectoderm (TE) cells. Notably, the ICM – which later separates out into epiblast (EPI) and primitive endoderm (PE) – is the source of mouse embryonic stem cells (ESCs; discussed in detail below), which are pluripotent, self-renewing and have the ability to give rise to all three types of primary germ layers (Keller, 2005). A large body of evidence has demonstrated crucial roles for various KLFs during early mouse embryonic development (Fig. 3). For instance, several laboratories have shown that Klf5−/− embryos die at embryonic day (E) 8.5, Klf2−/− at E12.5, Klf6−/− at E12.5, and Klf4−/− mice soon after birth (Katz et al., 2002; Kuo et al., 1997a; Matsumoto et al., 2006; Segre et al., 1999; Shindo et al., 2002). Further analyses showed that KLF5 is expressed early in the pre-implantation mouse embryo, starting at the two-cell stage, with persistent expression occurring during the morula and blastocyst stages (Lin et al., 2010). Although KLF5 is not detected at the epiblast stage between E5.5 and E6.5, it is present in primordial germ cells at E11.5 (Lin et al., 2010). Importantly, KLF5 regulates the formation of the three distinct cell lineages that arise during pre-implantation development, promoting TE and EPI formation, and repressing PE development (Lin et al., 2010). Abnormalities within all three lineages were also observed in Klf6−/− ESCs, suggesting some redundancy between KLFs in ESC differentiation (Matsumoto et al., 2006). In contrast, the expression of KLF2 and KLF4 is not required for the development of these cell lineages (Ema et al., 2008; Lin et al., 2010). Recently, comparative analyses of mouse and human blastocysts using single-cell RNA sequencing (Blakeley et al., 2015) demonstrated the expression pattern of KLF4 during pre-implantation development. It was observed that mouse embryos express Klf4 at the two-cell stage until the blastocyst stage, whereas KLF4 is upregulated at the eight-cell stage in human embryos. By contrast, Klf2 was highly expressed in the ICM and at very low levels in the TE in mouse embryos. Interestingly, KLF2 was completely absent from the EPI, PE and TE of human embryos. Finally, it was also demonstrated that KLF17 is highly expressed in EPI cells in human embryos, but is absent from the ICM and TE of mouse embryos (Blakeley et al., 2015). Combined, these findings highlight key roles for KLFs during early embryogenesis but also suggest that there are fundamental differences in the regulatory circuits that regulate lineage development in early mouse and human embryos. As we move on to discuss below, KLFs have also been implicated in later stages of development, in endoderm-, mesoderm- and ectoderm-derived lineages (as summarized in Table 2).
Fig. 3.

The expression of KLF2, KLF4 and KLF5 during mammalian pre-implantation embryo development. Diagram of mouse pre-implantation embryo development, highlighting the stages at which KLF2, KLF4 and KLF5 genes are expressed; the expression of these factors in human pre-implantation embryos is also indicated. Patterns of expression are based on findings reported by Blakeley et al. (2015) and Lin et al. (2010). EPI, epiblast; ICM, inner cell mass; PE, primitive ectoderm; TE, trophectoderm.
Table 2.
Krüppel-like factor participation in lineage formation during mouse embryogenesis
KLFs in the development of endoderm-derived lineages
Lung development
During mouse embryogenesis, normal lung development begins at E9.5 and can be divided into structural (growth) and functional (maturation) development with four histological stages: pseudoglandular, canalicular, terminal saccular and alveolar. The first two stages are restricted to embryonic development, whereas the terminal saccular stage extends from embryogenesis to the postnatal period, and the final alveolar maturation stage occurs in the postnatal (P) 5-P30 period (Warburton et al., 2010). Studies have shown that KLF2 is essential for lung development (Wani et al., 1999b). Because Klf2−/− mice die in utero at approximately E12.5, the role of KLF2 in lung development was studied using Klf2−/− ESCs in chimeric animals (Wani et al., 1999b). These Klf2−/− mice exhibit multiple abnormalities: lack of lung expansion with arrest in the late canalicular stage, with undilated acinar tubules and buds in peripheral regions of the lung. These findings suggest that KLF2 is vital for the later stages of embryonic lung development (Wani et al., 1999b).
KLF5 is also important for lung development. Immunohistochemistry analyses demonstrated that KLF5 is expressed in pulmonary epithelial cells throughout lung development in mice (Wan et al., 2008). At E18.5, KLF5 is expressed in a subset of cells in the alveolar compartment expressing surfactant protein C, a type II alveolar epithelial cell marker (Wan et al., 2008). KLF5 is also present in conducting airways, ciliated bronchiolar cells and non-ciliated cells expressing the bronchiolar cell marker CCSP (also known as SCGB1A1) (Wan et al., 2008). Because Klf5−/− mice are embryonically lethal at E8.5, well before lung development occurs, the conditional deletion of Klf5 in respiratory epithelial cells in the fetal lung was used to investigate KLF5 function in lung development (Wan et al., 2008). This study showed that ablation of Klf5 in respiratory (Lin et al., 2010) epithelial cells inhibits lung maturation during the saccular stage of development and causes death due to respiratory distress immediately after birth. Lung maturation and morphogenesis in the respiratory epithelium, the bronchiolar smooth muscle, and the pulmonary vasculature are also affected (Wan et al., 2008). Further analyses also demonstrated that KLF5 modulates the expression of key genes regulating surfactant lipid and protein homeostasis, vasculogenesis, and smooth muscle cell differentiation (Wan et al., 2008). In addition, Klf6 is also detected in the lung buds of mice at E12.5 (Laub et al., 2001b), but its function in lung development has not been explored. Nonetheless, these data demonstrate the crucial role of KLFs in lung development.
Gut development
Previous studies have demonstrated the expression of Klf4 and Klf5 in the intestine during mouse embryogenesis (Ohnishi et al., 2000). Klf4 is abundantly expressed at E15.5, but nearly absent by E17.5 (Ohnishi et al., 2000; Ton-That et al., 1997). Furthermore, the level of Klf4 expression in newborn mice is higher in the colon in comparison with the small intestine, but increases in both tissues with age (Ton-That et al., 1997). Klf5 transcripts are detected early in the primitive gut at E10.5, with increased expression observed at E15.5 and peaking at E16.5 until a decrease at E17.5. Interestingly, Klf5 expression is eventually confined to the crypt part of the mature intestinal epithelium (Ohnishi et al., 2000). Mice with intestinal-specific deletion of Klf5 (driven by the villin promoter) show high levels of mortality within the first 48 h after birth (McConnell et al., 2011). These mice, which display variegated expression of Klf5 in the gut, survive to 8 weeks of age but eventually exhibit defects in epithelial differentiation and migration resulting in impaired barrier function and inflammation (McConnell et al., 2011).
Other endoderm-derived organs
Unfortunately, the precise role of KLFs during the development of other endoderm-derived organs is not very well-defined. Previous studies have demonstrated that Klf6−/− mice exhibit failed hepatogenesis (Matsumoto et al., 2006). In addition, Klf13 is expressed in endothelial cells of the liver vasculature at E14.5 in mice (Lavallee et al., 2006). However, their function during hepatogenesis and development of liver vasculature remains unclear.
KLFs in the development of mesoderm-derived lineages
Development of the hematopoietic system
During embryogenesis, hematopoiesis is initiated independently at two sites: the extra-embryonic mesoderm of the yolk sac and the intra-embryonic site. The primitive hematopoiesis that occurs in the yolk sac generates erythrocytes that are necessary for embryo survival and distinct from fetal and adult erythrocytes (which express γ- and α-, and β-globin, respectively). By contrast, definitive hematopoiesis originating from the intra-embryonic site establishes myeloid, lymphoid and erythroid lineages as well as fetal and adult hematopoietic stem cells (HSCs). Several KLFs are expressed in these hematopoietic tissues and thus have been implicated in hematopoiesis. For example, the mouse yolk sac, fetal liver and adult spleen express ample levels of KLF1, 2, 3, 4, 5, 8, 11, 12 and 13 (Zhang et al., 2005). The expression pattern of these KLFs has also been confirmed in human erythroid cell line models (Zhang et al., 2005).
KLF1 is uniquely expressed in erythroid cells of the yolk sac, fetal liver, spleen and bone marrow in both mice and humans (Zhang et al., 2005). Klf1−/− mice develop fetal anemia during definitive erythropoiesis and die of profound β-thalassemia between E14 and E16 due to defects in red blood cell (RBC) maturation (Perkins et al., 1995; Siatecka and Bieker, 2011). This is mainly caused by defects in β-globin expression whereas α-globin levels are largely unaffected. Ultimately, KLF1 has been shown to play an essential role in embryonic and adult erythropoiesis by positively regulating β-globin (Siatecka and Bieker, 2011), ε-globin and γ-globin genes in the mouse embryo (Alhashem et al., 2011). KLF1 has also been shown to act as a master regulator of global erythroid gene expression by controlling the heme biosynthetic pathway, RBC membrane proteins, the cell cycle, transcription factor expression patterns (Drissen et al., 2005; Hodge et al., 2006), and long noncoding RNAs that regulate RBC maturation (Alvarez-Dominguez et al., 2014). Furthermore, during hematopoiesis, Klf1 represses megakaryocyte formation from the megakaryocyte-erythroid lineage (Frontelo et al., 2007). Finally, recent studies in mice demonstrate the existence of two KLF1 isoforms – full-length KLF1 and KLF1-SV – that are generated by alternative splicing, with the latter potentially impacting expression levels of the transcriptional targets of full-length KLF1 (Yien et al., 2015).
KLF2, by contrast, is required for the production of embryonic ε-globin in human and εy-globin and βh1-globin in mouse erythroid cells, but not adult β-globin (Alhashem et al., 2011; Basu et al., 2007). Consequently, Klf2−/− mice are embryonically lethal between E12.5 and E14.5 due to defects in the development of vascular endothelial cells, leading to intra-embryonic and intra-amniotic hemorrhage (Kuo et al., 1997a). Studies in mice show that KLF2 also regulates T-cell differentiation, quiescence and survival (Kuo et al., 1997b). Recently, it was also postulated that KLF2 alternatively regulates thymocyte and T-cell trafficking in mice (Carlson et al., 2006). Interestingly, Klf1/Klf2 double knockout mouse embryos die before E11.5 as the result of deficiencies in endothelial and erythroid development (Basu et al., 2007; Vinjamur et al., 2014). KLF3, which is expressed in the fetal liver, plays a role in normal B-cell development in the bone marrow, spleen and peritoneal cavity in mice; these processes are impaired in Klf3 null mice (Pearson et al., 2011; Turchinovich et al., 2011).
Although it was known originally for its expression in the skin and gut, Klf4 is induced in a stage-specific manner during embryogenesis as cells transition from HSCs into granulocyte/macrophage progenitors, and is reduced in megakaryocyte/erythrocyte progenitors. Interestingly, Klf4 deficiency within myeloid progenitors results in reduced monocyte and granulocyte formation, suggesting a potential role for KLF4 in primary myeloid differentiation (Feinberg et al., 2007; Kurotaki et al., 2013). KLF5 is also implicated in myeloid differentiation: Klf5 expression increases in granulocyte lineage cells during both mouse and human hematopoiesis, and the targeted, conditional ablation of Klf5 in the hematopoietic compartment of mice revealed its novel role as a myeloid transcription factor (Shahrin et al., 2016). In these studies, KLF5 contributed to neutrophil formation and inhibited eosinophil production. Moreover, KLF5 serves as a target of C/EBPα after the formation of granulocyte-macrophage progenitors and impacts the fate of myelomonocytic precursors (Shahrin et al., 2016).
Klf6 null mice also exhibit markedly reduced hematopoiesis and poorly organized yolk sac vascularization, which probably contribute to the embryonic lethality at E12.5 of these mice (Matsumoto et al., 2006). In contrast, Klf7 null mice do not exhibit changes in hematopoiesis despite the ubiquitous expression of Klf7 in all hematopoietic lineages and human pluripotent stem cell subsets (Schuettpelz et al., 2012). However, the overexpression of Klf7 was shown to inhibit the formation of all myeloid lineage cells (Schuettpelz et al., 2012), suggesting a deleterious role of excess Klf7 in development. Furthermore, recent transcript analysis of Klf8 demonstrated its expression in erythroid lineages: at E11.5 in the yolk sac, at E14.5 in the fetal liver, and finally in adult peripheral blood (Funnell et al., 2013). It was further shown that double mutants for Klf3 and Klf8 die at E14.5, whereas Klf3−/− and Klf3−/−/Klf8gt/gt mice exhibit de-repression of embryonic, but not adult, globin expression (Funnell et al., 2013). These observations suggest that KLF8 might play a compensatory role upon Klf3 ablation. KLF11 is also expressed during embryonic hematopoiesis, although Klf11−/− mice exhibit no changes in hematopoiesis during development (Song et al., 2005). Finally, studies have shown that Klf13 expression increases upon maturation of erythroid precursors in mice (Gordon et al., 2008). As such, Klf13−/− mice exhibit an increase in the number of immature erythrocytes in the spleen; however, the number of committed precursors is unchanged in the bone marrow (Gordon et al., 2008). Interestingly, these immature precursors accumulate at the proerythroblast stage of development and exhibit increased expression of Klf1 and Klf3 (Gordon et al., 2008). Collectively, these studies highlight the crucial role of KLFs in the development of the hematopoetic system.
Cardiovascular development
The vasculature is the first organ system to develop and its development is initiated early during embryogenesis. The vascular system is formed by the differentiation of primitive mesodermal cells into endothelial cells (vasculogenesis) and by endothelial cell proliferation and migration to colonize tissues (angiogenesis). In the final stage, with the onset of a heart beat and blood flow, vessels remodel and branch out to penetrate different organs. To date, six KLFs have been implicated in cardiovascular development: KLF2, 3, 4, 5, 6 and 13.
Early studies demonstrated that Klf2 is expressed in the mouse embryo as early as E7 in the developing vasculature, exhibits a decrease in expression at E11, but returns to high levels at E15 and remains elevated, specifically in the umbilical arteries and veins (Anderson et al., 1995). By contrast, recent experiments have highlighted that changes in Klf2 expression during cardiovascular development might correlate with variations in fluid shear stress (Lee et al., 2006). Namely, Klf2 expression begins at E8.5 within endothelial and endocardial compartments and rises by E10.5 as the heart tube loops and endocardial cushions form. By E11.5, Klf2 expression is higher in the arteries (compared with veins), in the endothelium overlying the endocardial cushions, and in the common ventricular outflow tract – sites predicted to experience high fluid shear forces (Lee et al., 2006). However, Klf2 expression becomes restricted to the flow side of developing valves as the cushions mature by E14.5 (Lee et al., 2006). In addition, Klf2 expression is also detectable in the endocardium lining the intraventricular papillary muscles. Surprisingly, no changes in vasculogenesis and angiogenesis are observed in Klf2−/− mice; however, these mice do exhibit impaired blood vessel maturation (Wu et al., 2008). More recent studies have shown that Klf2 is also expressed at E9.5 in endocardial cells of the atrioventricular canal cushion region and that its deletion results in embryonic heart failure, attributable to a high cardiac output state caused by a profound loss of peripheral vascular resistance (Chiplunkar et al., 2013b; Lee et al., 2006).
KLF3 is also implicated in cardiac development. Although early observations did not report an embryonic phenotype in Klf3−/− mice (Funnell et al., 2013), recent data show that homozygous mutant Klf3H275R/H275R mice have significant lethality at E14.5-E16.5 (Kelsey et al., 2013). Increased mortality in these mice is likely to be due to heart failure, as the ventricular myocardium and septum are notably thinned and disorganized in these mice (Kelsey et al., 2013). Furthermore, mice harboring heterozygous KLF3H275R point mutants exhibit increased perinatal lethality with numerous abnormalities in heart morphology at E12.5: marked biventricular myocardial hypertrophy and aortic valve leaflet thickening with adult survivors exhibiting hypotension, aortic valvular stenosis, aortic dilatation, myocardial hypertrophy and increased chamber size (Kelsey et al., 2013). It was suggested that the cardiovascular abnormalities in these mice could be attributed to blood pressure dysregulation, because KLF3 is also expressed in the renal vasculature during development (Kelsey et al., 2013).
KLF4 is expressed in extra-embryonic tissues at E4.5, and in mesenchymal and ectodermal tissues, and the endothelium by E10.5. However, vascular abnormalities have not been reported in Klf4 null mice (Ehlermann et al., 2003). A more recent study demonstrated that Klf4 is expressed in the heart (in the atrium and ventricle at E18.5, but not in smooth muscle) from late embryonic development to adulthood (Yoshida et al., 2010). These findings led to further investigations using the smooth and cardiac muscle-specific conditional deletion of Klf4 in mice (Yoshida et al., 2010). Although abnormalities were not observed within smooth muscle and vascular morphology or in the heart and great arteries at P14 and P28, five-week-old Klf4−/− mice demonstrated decreased cardiac output that was associated with abnormal postnatal growth (Yoshida et al., 2010). It was further shown that the lack of Klf4 in these mice decreased the expression of many cardiac genes, including myosin light polypeptide 1 (Myl1), cardiac/slow skeletal troponin C (Tnnc1), myosin binding protein C (Mybpc2), natriuretic peptide precursor type B (Nppb) and Gata4 (Yoshida et al., 2010). An independent study showed that Klf2/Klf4 double knockout mice die earlier than the single knockouts, possibly due to gross hemorrhaging of multiple vessels (Chiplunkar et al., 2013a). Thus, KLF4 might play a role in vasculogenesis and the maintenance of embryonic vascular integrity by interacting with KLF2.
KLF5 is also abundantly expressed in developing blood vessels, and its deletion leads to early embryonic lethality (Shindo et al., 2002). Similarly, KLF6 is important in specifying mesoderm to hematopoietic and vascular lineages, and for vasculogenesis (Matsumoto et al., 2006). As such, Klf6−/− mice have poorly organized yolk sac vasculature and decreased levels of Flk-1 (KDR)-positive cells (vascular progenitors) (Matsumoto et al., 2006). Furthermore, initial in situ hybridization studies revealed that Klf13 is highly expressed in the heart and cephalic mesenchyme: its expression in the primitive heart can be detected as early as E11.0 in the atria and ventricles, but decreases by E13.0-E16.0 (Martin et al., 2001). Further comprehensive immunohistochemistry of developing mouse embryos (Lavallee et al., 2006) demonstrated that, at E9.5, KLF13 is detected mostly in the heart and the epidermis. At E10.5, KLF13 is localized in the atrial myocardium and endocardial layer and by E12.5 in atria and ventricles with higher atrial expression (Lavallee et al., 2006). KLF13 staining is also present in the cushions of the atrioventricular region and the truncus arteriosus (Lavallee et al., 2006). Although KLF13 is subsequently downregulated postnatally, its expression is restricted to the atrial and ventricular myocardium, with the highest expression in the valves and interventricular septum (Lavallee et al., 2006). Mechanistically, KLF13 interacts with GATA4 to regulate atrial expression of Nppb, which is essential for cardiac development (Lavallee et al., 2006). Consequently, Klf13−/− mice exhibit an enlarged heart and increased susceptibility to cardiovascular injury, highlighting the importance of KLF13 in heart development (Gordon et al., 2008). Combined, these data emphasize the essential role of KLFs in cardiovascular development.
Adipogenesis
The adipocyte lineage originates from mesenchymal progenitors that form adipocyte precursor cells or preadipocytes, which then differentiate into mature, lipid-containing adipocytes. Several KLFs have been implicated in adipogenesis. For example, the analysis of embryoid bodies derived from Klf2−/− mouse ESCs showed that they retain the ability to form functional adipocyte precursors that differentiate upon adipogenic stimuli (Wu et al., 2005). As such, Klf2−/− cells are more prone to differentiate at the early stage of adipocyte differentiation, suggesting that KLF2 is a natural repressor of differentiation (Wu et al., 2005). In addition, Klf3−/− mice are smaller than their wild-type littermates and exhibit reduced levels of white adipose tissue (Sue et al., 2008). Although these studies highlight that KLF2 and KLF3 play a role in adipogenesis, further studies are required to delineate the mechanisms by which they act.
KLFs and the development of ectoderm-derived lineages
Skin development
Two KLFs, namely KLF4 and KLF5, are implicated in skin development. In mice, the expression of Klf4 and Klf5 is observed at E15.5 in the developing skin (Ohnishi et al., 2000). A high level of Klf4 expression is then observed in the epidermis at E16.5 with a slight reduction by E17.5 (Ohnishi et al., 2000). In contrast, Klf5 expression persists until E17.5 (Ohnishi et al., 2000). Importantly, Klf4−/− mice die within 24 h of birth due to improper late-stage differentiation of structures in the epidermis and loss of skin barrier function (Segre et al., 1999). Moreover, it was shown that Klf4 overexpression (under the control of the keratin 5 promoter) during development allows for the epidermal barrier to be formed in utero one day earlier than in wild-type mice (Jaubert et al., 2003). Similarly, it was shown that the ectopic expression of Klf5 during development in mice results in reduced numbers of hair follicles on the torso and a lack of hair follicles on the tail, as well as abnormal epidermal development and differentiation (Sur et al., 2006). Although KLF4 and KLF5 thus appear to be crucial for skin barrier function, future studies need to focus on their function in the formation of the epidermis.
Central nervous system development
Global gene expression analyses suggest that several KLFs are implicated in the development of the central nervous system (CNS): Klf2, 3, 4, 6, 7, 9, 11, 12 and 13 (Qin et al., 2011). However, only a few KLFs have been well characterized during CNS development. The expression of Klf4, for example, is enriched in fetal hypothalamic thyrotropin-releasing hormone neurons at E15.0, peaking at the neonate stage and P7, and then decreasing in adult (Qin et al., 2011). This downregulation of Klf4 during development and in differentiated neurons is necessary for brain development and homeostasis (Qin et al., 2011). Interestingly, the timing of Klf4 induction during embryogenesis coincides with an increase in the biosynthesis of thyrotropin-releasing hormone (TRH), and further experiments have characterized the role of KLF4 in regulating TRH expression in hypothalamic neurons (Pérez-Monter et al., 2011). Recently, KLF4 was also shown to play an important role in cerebellar development as well as in granule cell proliferation between E13.5 and E15.5 in mice (Zhang et al., 2015). In addition, analyses of Klf4−/− mice show that PAX6 – a marker for granule cells – is reduced at E13.5 and completely absent at E15.5, but is re-expressed at E16.5 and E18.5 (Zhang et al., 2015). KLF6, by contrast, is expressed strongly in the nervous system during embryogenesis and appears to play a role in neural tube development (Laub et al., 2001b). Consequently, one of the key anatomical anomalies in Klf6−/− mice is incomplete neural tube closure (Matsumoto et al., 2006).
The expression of KLF7 has also been closely examined in the developing nervous system of mouse embryos. One study (Lei et al., 2001) showed that Klf7 is expressed starting at E8.5 in the trigeminal ganglion, the VII-VIII neural crest complex, and the subventricular neuroepithelium of both forebrain and hindbrain regions. At E11.5, Klf7 expression is detected in the neural crest-derived sensory nervous system and, at E15.5, in the brain, spinal cord, retinal neuroepithelium and the inferior XI/X complex. This study also demonstrated that Klf7 is co-expressed with the receptor for nerve growth factor (TrkA; NTRK1) and directly regulates its expression in the peripheral nervous system. Subsequent studies demonstrated that Klf7 expression commences at about E9.5, is maximal at about E11.5, and declines in later stages of development (Laub et al., 2001a). At E9.5, Klf7 expression in the CNS and peripheral nervous system closely correlates with that of the neurospecific gene Neurod1. By E11.5, Klf7 is intensely expressed in the fore-, mid- and hindbrain, and in the trigeminal, geniculate, vestibulocochlear, petrosal, superior, jugular, nodose, accessory and dorsal root ganglia. In the neural tube at E11.5, Klf7 is expressed distinctly in the mantle zone, where postmitotic neuroblasts are located, and colocalizes with markers for neuronal differentiation [e.g. NeuN (RBFOX3), TuJ (TUBB3), SCG10 (STMN2)]. Between E11.5 and E18.5, Klf7 expression gradually decreases in the neural tube, while increasing in the dorsal root ganglia. Additionally, Klf7 is detected in the neural retina at E17.5 and the olfactory epithelium at E16.5 (Laub et al., 2001a). In postnatal and adult animals, Klf7 expression is confined to the cerebellum and dorsal root ganglia. Importantly, it was shown that Klf7−/− mice die within 3 days of birth and are characterized by an absence of milk in the stomach, hypopnea, cyanosis and failed response to clamp stimulation of their tails (Laub et al., 2005). Furthermore, Klf7−/− mice also exhibit severely hypoplastic olfactory bulbs (Laub et al., 2006), suggesting that KLF7 plays a role in the development of the olfactory system. Indeed, Klf7 was previously detected in the olfactory epithelium at E16.5 (Laub et al., 2001a), and more recent time-lapse analyses of wild-type mouse embryos show that Klf7 transcripts can be detected in the olfactory epithelium as early as E11.5. In addition, the loss of Klf7 activity leads to reduced formation of neurites at E11.5 and impaired axon projection in the olfactory and visual systems, cerebral cortex and hippocampus at E12.5 (Laub et al., 2005). Mechanistically, Klf7 deletion correlates with a significant downregulation of p21WAF/CIP (CDKN1A) and a reduction in p27KIP1 (CDKN1B) protein levels in olfactory sensory neurons, suggesting that interaction between these factors is important for neuronal morphogenesis in the olfactory epithelium (Laub et al., 2005). Furthermore, expression arrays of olfactory sensory neurons from wild-type and Klf7−/− embryos revealed that KLF7 might regulate olfactory marker protein (OMP) and the neural cell adhesion protein L1 (L1CAM) (Kajimura et al., 2007), both of which are proteins that are crucial for olfactory development.
It has also been shown that Klf9 transcripts are upregulated by mature thyroid hormone in primary neurons and astrocytes, but not oligodendrocytes, which are derived from embryonic and neonatal mice brains, respectively (Denver et al., 1999). In addition, the induction of Klf9 regulates thyroid hormone-mediated neurite extension and branching as well as neuronal differentiation (Denver et al., 1999). Further studies also showed that Klf9 expression is transiently induced during the early postnatal phase due to neuronal activity (Scobie et al., 2009). Finally, ablation of Klf9 impairs the maturation of dentate granule neurons during dentate gyrus development as well as during adult hippocampal neurogenesis (Scobie et al., 2009). Although further studies are required to ascertain the mechanism, these studies highlight that KLF9 is essential for neuronal development.
The role of KLFs in stem cells and regeneration
Klf2 was the first KLF transcript identified in mouse ESCs (Carlson et al., 2006). Soon after, Takahashi and Yamanaka reported that the overexpression of Klf4 along with c-Myc (Myc), Oct3/4 (Pou5f1) and Sox2 could reprogram somatic cells into induced pluripotent stem cells (iPSCs) (Takahashi et al., 2007; Takahashi and Yamanaka, 2006). Since this groundbreaking discovery, intense efforts have been focused on understanding how KLF4 as well as other KLFs function in stem cells and regeneration.
KLFs in ESC self-renewal and differentiation
A number of studies have shown that multiple KLFs regulate the expression of transcription factors involved in maintaining pluripotency (Fig. 4). KLF2, KLF4 and KLF5 are associated with the pluripotent state of mouse ESCs, as triple knockdown of these factors renders cells for differentiation (Jiang et al., 2008). Subsequent studies demonstrated that KLF2, KLF4 and KLF5 can maintain an undifferentiated state in mouse ESCs and can reprogram epiblast-derived stem cells (EpiSCs) into iPSCs when overexpressed (Jeon et al., 2016). In addition, KLF2, KLF4 and KLF5 were demonstrated to occupy the promoter regions of genes encoding key pluripotency factors (Oct4, Sox2 and Nanog) and, in turn, these pluripotency factors were shown to occupy the promoters of the genes encoding these KLFs, thereby suggesting an internal regulatory circuit within mouse ESCs (Jiang et al., 2008). Further analysis showed that Klf2 is regulated by OCT4, whereas Klf4 and Klf5 are regulated by OCT4 and the LIF/STAT3 pathway (Hall et al., 2009). In addition, KLF2, but not KLF4, exhibits clonogenic capacity, maintains undifferentiated ESCs in the genetic absence of STAT3, and confers resistance to bone morphogenetic protein-induced differentiation (Hall et al., 2009). In contrast, Klf5−/− ESCs can maintain pluripotency but show increased expression of differentiation-related genes and exhibit frequent and spontaneous differentiation. The overexpression of other KLFs, with the exception of KLF9 and KLF10, in ESCs demonstrated that they exhibit increased binding affinity to downstream regulatory genes similar to those bound by KLF2, KLF4 and KLF5 (Jeon et al., 2016). Further expression profiling performed during the differentiation of mouse ESCs under conditions that recapitulate differentiation in vivo allowed for the identification of three distinctly expressed KLF groups: (1) Klf2, Klf4, Klf5 and Klf9, which show high levels of expression in the undifferentiated state and are rapidly downregulated within 3 days of differentiation; (2) Klf1, Klf3 and Klf16, which are upregulated upon ESC differentiation; and (3) Klf6, Klf7, Klf8, Klf10-Klf15, the expression of which is unaffected by ESC differentiation (Bruce et al., 2007). Based on these data, it is postulated that competition among KLFs for occupancy of the promoter elements (CACCC boxes) probably determines self-renewal or progression towards differentiation. Interestingly, KLF1, KLF3, KLF6, KLF7 and KLF8 are not sufficient to maintain the self-renewal of mouse ESCs, despite their ability to bind to Nanog regulatory regions (Jeon et al., 2016).
Fig. 4.
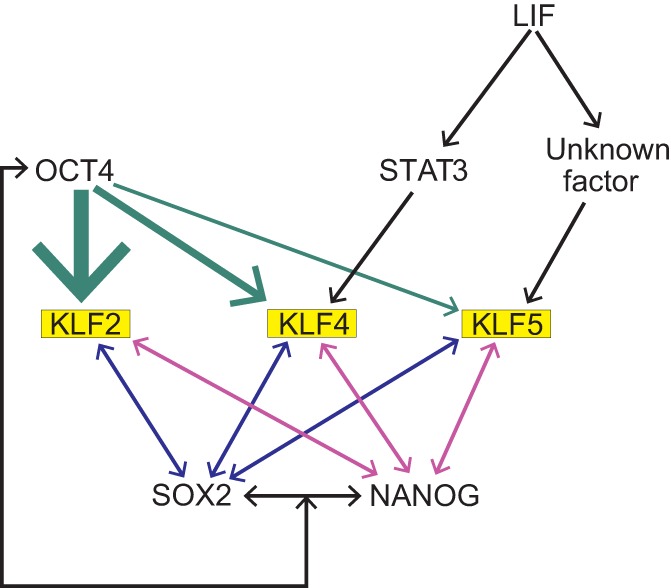
KLFs and pluripotency. Schematic summarizing some of the regulatory interactions between KLF2, KLF4, KLF5 and factors involved in pluripotency in iPSCs. Interactions are based on findings reported by Jiang et al. (2008) and Hall et al. (2009). Arrows indicate induction of expression, with the thickness of arrows indicating the level of induction.
Although most studies have focused on mouse ESCs, some approaches have attempted to understand the function of KLFs in human ESCs. In comparison with mouse ESCs, human hESCs exhibit low levels of KLF2, KLF4 and KLF5 expression that steadily increase upon differentiation (Cai et al., 2010), suggesting that mouse and human ESCs use distinct regulatory networks. This is potentially due to functional differences between these cells: human ESCs are more similar to mouse EpiSCs, which are derived from the late epiblast layer of post-implantation embryos, than to mouse ESCs, which originate from the ICM of blastocysts (Cai et al., 2010). Importantly, KLFs appear to play important roles in both the earlier ‘naive’ state and the ‘primed’ state (Kim et al., 2008). Naive mESCs are stabilized by the LIF/STAT3 pathway whereas basic fibroblast growth factor (bFGF) and TGFβ/activin signaling results in their destabilization. In comparison, bFGF and TGFβ/activin signaling are required for mouse EpiSC stabilization. Furthermore, in naive, mESCs, KLF2, KLF4 and KLF5 are expressed concurrently with NANOG and ESRRβ. However, their levels are downregulated during progression to the post-implantation epiblast (‘primed’) state (Hanna et al., 2010). Conversely, overexpression of Klf2 or Klf4 is sufficient for induction of naive pluripotency (Blakeley et al., 2015). Despite these differences between mouse and human ESCs, the generation of human iPSCs was accomplished using the same factors as for mouse iPSCs: Oct4, Sox2, c-Myc and Klf4 (Kim et al., 2008; Maffioletti et al., 2015; Moad et al., 2013; Park et al., 2008; Takahashi et al., 2007).
KLFs and reprogramming
The ability to reprogram numerous somatic cell types, from both mice and humans, into iPSCs has provided exciting opportunities for regenerative medicine, for studying diseases, and for drug discovery (Singh et al., 2015). Indeed, the Oct3/4, Sox2, Klf4 and c-Myc (OSKM) ‘cocktail’ or variations of it in multiple tissues of murine and human origin (fetal, neonatal and adult fibroblasts, primary hepatocytes, adult neuronal stem cells, menstrual blood stromal cell, prostate, bladder and ureter cells) can reprogram cells into iPSCs, with possible further differentiation into specialized cells such as gastric epithelial cells, lymphocytes, erythrocytes, myeloid cells, hepatocyte-like cells, retinal cells and melanocytes (Aoi et al., 2008; Kim et al., 2008; Kitada et al., 2012; Li et al., 2011; Park et al., 2008; Seiler et al., 2011; Takahashi et al., 2007; Takahashi and Yamanaka, 2006; Yang et al., 2011). As mentioned above, KLF4 has been identified as a factor that can reprogram mouse and human fibroblasts to induce their pluripotent state (Takahashi et al., 2007; Takahashi and Yamanaka, 2006). Initially, Takahashi and Yamanaka used a genetic approach to screen 24 pre-selected factors that had previously been shown to either participate in the maintenance of pluripotency at the embryonic stage or be upregulated in tumors and contribute to the ESC phenotype. The introduction of single factors into fibroblasts did not induce pluripotency, but by using a combination of the pre-selected factors they could induce a pluripotent state. By withdrawing factors individually from the combination, they were able to identify those factors that were essential for the induction of pluripotency in somatic cells. Using this approach, they determined that OCT4, SOX2, KLF4 and c-MYC play an important role in the generation of iPSCs from somatic cells.
Their genomic analyses of iPSCs also demonstrated that they expressed Myb, Kit, Gdf3, Zic3, Dppa3, Dppa4, Dppa5 (Dppa5a), Nanog, Sox2, Esrrb, Rex1 (Zfp42), Dnmt3a, Dnmt3b, Dnmt3l, Utf1, Tcl1, and the LIF receptor gene, all of which are commonly expressed in mESCs (Takahashi and Yamanaka, 2006). However, the levels of expression of these genes varied between iPSCs and mESCs. Additionally, Nishimura and colleagues showed that KLF4 dosage correlates with the level of pluripotency induction (Nishimura et al., 2014); however, the precise mechanism by which KLF4 induces pluripotency is not very well understood. It has been postulated that KLF4 plays a dual role in this process, by repressing markers of differentiation and facilitating the expression of pluripotency genes. Specifically, it has been shown that during the first phase, KLF4 regulates genes associated with differentiation, such as Tgfb1, Pdgfra and Col6a1 and, at the later stage of reprogramming, KLF4, upregulates genes associated with pluripotency (Oct4, Tdgf1 and Klf5) (Polo et al., 2012). For instance, in iPSCs, the levels of p53 (TRP53) – a known repressor of Nanog – are lower in comparison with those in somatic cells, and previous studies showed that KLF4 negatively regulates p53 during differentiation. It has also been shown that KLF4 induces the expression of Nanog by binding to its promoter, thus preventing ESC differentiation (Zhang et al., 2010). Another study demonstrated that UTX (KDM6A; a H3K27 demethylase enzyme) interacts with OCT4, SOX2 and KLF4, and that this results in removal of the repressive H3K27me3 mark from early activated pluripotency genes such as Fgf4, Sall4, Sall1 and Utf1 (Mansour et al., 2012). Finally, it was demonstrated that during the reprogramming of human fibroblasts to iPSCs, OCT4, SOX2 and KLF4 bind closed chromatin with c-Myc enhancing these interactions (Soufi et al., 2012). Together, these studies highlight the importance of KLF4 in the reprogramming process and the versatility of this system for understanding KLF function.
KLFs and regeneration
Given their roles in controlling self-renewal and/or cell differentiation, it is not surprising that KLFs have been implicated in the regenerative processes and, hence, are being targeted for use in regenerative medicine. For example, the prospect of overexpressing KLF4, SOX9 and c-MYC to direct the reprogramming of adult dermal fibroblasts into polygonal chondrogenic cells, independent of iPSCs, for cartilage regeneration has been previously proposed (Hiramatsu et al., 2011; Outani et al., 2011). In addition, it has been shown in a mouse model that epidermal multipotent stem cells express Klf4 and that deletion of Klf4 reduces the stem cell population and self-renewal efficiency of these stem cells (Li et al., 2012). For instance, KLF4 has been shown to contribute to skin integrity and Klf4−/− mice have significantly delayed wound healing post-injury compared with wild-type mice, suggesting a crucial role for KLF4 in cutaneous wound healing (Li et al., 2012). Another factor, KLF6, also known as tissue remodeling factor, was shown in mice to promote vascular repair after bilateral endoluminal injury to the common femoral artery. This enhanced repair process was accompanied by increased activity of matrix metallopeptidase 14, endoglin and activin receptor-like kinase 1 (ACVRL1), which are involved in angiogenesis and vascular remodeling (Gallardo-Vara et al., 2016; Garrido-Martin et al., 2013). The potential role of KLFs in axonal regeneration has also attracted a lot of attention. For instance, studies in rats have shown that KLF4 and KLF9 are repressors of axon growth in retinal ganglion cells (RGCs), whereas KLF6 and KLF7 are capable of stimulating neurite growth in these cells (Moore et al., 2009). Furthermore, KLF7 induction in the adult rat corticospinal tract improves axon regeneration in conjunction with upregulation of Trk neurotrophin receptors (Blackmore et al., 2012), and KLF9 was revealed as a factor that contributes to myelin regeneration in the mouse cortex following cuprizone-mediated demyelination, acting via thyroid hormone pathway induction (Dugas et al., 2012). Overall, these various studies highlight the therapeutic potential of KLFs in regenerative medicine, although further studies are needed to dissect their precise roles and mechanisms of action.
The role of KLFs in mammalian diseases
Accumulating evidence in the last decade has demonstrated that KLFs play a vital role in the etiology and progression of many mammalian diseases (summarized in Fig. 5), ranging from cardiovascular disease to various types of neuronal disorders. The function of KLFs in cancer biology (Box 1) has also been well studied (reviewed by Tetreault et al., 2013), and therapeutic strategies to modulate the expression of KLFs and/or its downstream effectors are the focus of current studies. Below, we discuss how KLFs have been implicated in some of these diseases and disorders, focusing on those that are linked to the aforementioned developmental roles for KLFs.
Fig. 5.

Summary of the roles of KLFs in human disease. Diagram summarizing the specific roles reported for KLFs in neuronal disorders, cardiovascular diseases, kidney diseases, gastrointestinal disorders and hematopoietic disorders. It should be noted that the involvement of KLFs in various types of cancer is not included in this schematic.
Box 1. KLFs and cancer.
As KLFs play a vital role in mammalian development, stem cell biology, and cellular regeneration, it is not surprising that they have also been implicated in cancer biology. In fact, KLFs are associated with various malignancies (reviewed by Tetreault et al., 2013). Mechanistically, KLFs have a direct impact on almost every aspect of cancer biology: regulation of the cell cycle, apoptosis, Wnt/β-catenin signaling, RAS signaling, NOTCH signaling, oncogenic transformation, tumor metastasis, and microenvironment (Tetreault et al., 2013). For instance, KLF4, a key regulator of normal cell proliferation, inhibits tumor growth in various cancers such as pancreatic, colorectal, neuroblastoma and lung cancers by inducing the expression of CDK inhibitors and inhibiting cyclin D1 and FOXM1 expression (Kong et al., 2013; Shum et al., 2013; Wei et al., 2008; Zammarchi et al., 2011). Conversely, KLF5 promotes tumor cell growth by upregulating CCNA2, CDT1 and E2F3, all of which are key cell cycle genes (Liu et al., 2011; Takagi et al., 2012). Furthermore, loss of KLF5 expression in colon cancer cells reduces activation of Wnt/β-catenin signaling, a key pathway in tumor cell proliferation (McConnell et al., 2009; Nandan et al., 2008). Interestingly, splice variants of KLF6 have also been implicated in the progression of non-small cell lung cancer as well as prostate cancer by reducing the expression of CDKN1A (Narla et al., 2005). Collectively, these studies suggest that specific KLFs might potentially serve as therapeutic targets when treating the progression of cancer.
KLFs in hematopoietic disorders
KLF1 is essential for erythropoiesis and, as such, has been implicated in disorders that involve RBC generation in adults. For example, it was demonstrated that KLF1-mediated erythrocyte differentiation is essential for RBC generation, and that the suppression of Klf1 expression by inhibiting IL-2 (IL2) signaling in mice contributes to failed erythropoiesis and eventual anemia in mice (Chopra et al., 2015). In addition, dominant mutants and haploinsufficiency in mouse and human KLF1 contribute to altered erythropoiesis, leading to several hematological disorders such as neonatal anemia and congenital dyserythropoietic anemia (Siatecka and Bieker, 2011). Because β-globin levels are reduced and embryonic globin levels are increased in Klf1−/− mice (Bieker, 2010), it has been postulated that the induction of human γ-globin levels by knockdown of KLF1 could serve as a viable therapeutic strategy for individuals with sickle-cell anemia or β-thalassemia (Bieker, 2010).
KLFs in cardiovascular disease, obesity, and metabolic syndrome
As mentioned above, KLF2 is vital for endothelial cell homeostasis: it regulates leukocyte adhesion to the endothelium, thrombotic function, and endothelial proliferation, migration and angiogenesis. Based on these diverse roles, loss of Klf2 expression has been implicated in atherosclerosis and thrombotic angiopathy (Agustian et al., 2013; Atkins et al., 2008; Boon et al., 2011). Furthermore, the pleiotropic effects of statins in preventing atherosclerosis might be directly mediated by KLF2 in endothelial cells (Tuuminen et al., 2013). Interestingly, the endothelial lineage-specific expression of Klf2 in bone marrow-derived mononuclear cells induces neovascularization in ischemic murine models, suggesting that KLF2 might have a therapeutic role in diseases with impaired neovascularization such as atherosclerosis (Boon et al., 2011). In addition, KLF2 was reported as an atheroprotective factor that regulates primary macrophage foam cell formation (Atkins et al., 2008).
A number of recent studies have also highlighted anti-inflammatory and anti-thrombotic roles for KLF4 in vascular biology (Hamik et al., 2007), which have direct implications for understanding the progression of atherosclerosis. The conditional knockout of Klf4 in smooth muscle cells (SMCs), for example, specifically increases the stability of atherosclerotic plaques and fibrous cap thickness in murine models of atherosclerosis (Shankman et al., 2015), although further studies are needed to decipher the role of KLF4 in mediating potential crosstalk between SMCs and endothelial cells in atherosclerosis. Atherosclerosis and peripheral vascular disease can also lead to neovascularization due to a chronic ischemic state, and recent studies demonstrate that KLF10 mediates the TGFβ1-induced generation of bone marrow-derived proangiogenic cells in the setting of ischemic injury (Wara et al., 2011), again highlighting a potential target for therapy in these conditions.
A number of KLFs have also been associated with heart failure. For example, reduction of myocardial Klf4 increases susceptibility to heart failure (Liao et al., 2010), which mechanistically might be due to a loss of transcriptional control of mitochondrial biogenesis (Liao et al., 2015). Furthermore, the loss of Klf15 in mice increases susceptibility to heart failure and aortic aneurysmal formation in a p53- and p300 acetyltransferase-dependent manner (Haldar et al., 2010). In addition, recent findings have revealed that KLF15 inhibits vascular smooth muscle activation by inhibiting NF-κB activity (Lu et al., 2013). Finally, recent studies demonstrate that KLF15 is a key regulator of cardiac lipid metabolism (Prosdocimo et al., 2014) and circadian mediated arrhythmogenesis (Jeyaraj et al., 2012).
Several KLFs have recently been linked to metabolic syndromes. KLF7 inhibits transcriptional inducers of human adipogenesis (Kanazawa et al., 2005) and modulates insulin sensitivity in human pancreatic β cells and skeletal muscle cells (Kawamura et al., 2006). Furthermore, it has been reported that variants in KLF7 are protective against type 2 diabetes mellitus in Japanese and Danish patient cohorts (Kanazawa et al., 2005; Zobel et al., 2009). In addition, multiple laboratories have reported that KLF15 induces mouse adipocyte differentiation by increasing the expression of PPARγ (Mori et al., 2005), which has direct implications when correlated with the glucocorticoid-induced effects on mouse adipogenesis (Asada et al., 2011). Interestingly, all of these findings are based on tissue-specific constitutive knockout murine models, highlighting specific and important roles for KLFs in development and disease.
KLFs in gastrointestinal disorders
The role of KLF4 and KLF5 in intestinal disorders has been well studied (McConnell and Yang, 2010). In addition, Klf9 null mice exhibit dysregulated intestinal crypt cell proliferation and villus cell migration, which contribute to loss of intestinal barrier function in mice (Simmen et al., 2007). Furthermore, the predominant expression of KLF9 occurs in mouse intestinal smooth muscles, thereby suggesting potential crosstalk between intestinal smooth muscle and epithelial cells that might contribute to maintenance of the mouse intestinal barrier (Simmen et al., 2007).
KLFs have also been implicated in aberrant liver function. For instance, KLF6-mediated upregulation of the TGFβ pathway is linked to the progression of mouse liver fibrosis (Friedman, 2000; Kawada, 2011). Furthermore, functional polymorphisms in KLF6 are associated with progression of nonalcoholic fatty liver disease in humans (Miele et al., 2008). The modulation of PPARα activity by KLF6 in mice contributes to hepatic glucose and lipid metabolism in fatty liver disease (Bechmann et al., 2013). In addition, KLF11 regulates collagen gene expression by silencing heterochromatin protein 1 expression and, consequently, Klf11 null mice exhibit increased liver fibrosis due to unchecked collagen deposition (Mathison et al., 2013). Combined, these data suggest an essential role for KLFs in gastrointestinal disorders.
KLFs in kidney disease
In the last decade, several studies of the functional role of KLFs in kidney disease have been reported. Damage to the glomerular filtration barrier is the predominant consequence of kidney disease, and several recent studies demonstrate the significance of KLFs in filtration barrier maintenance in the setting of injury. For instance, the loss of endothelial-specific Klf2 exacerbates the progression of diabetic nephropathy and chronic kidney disease (Zhong et al., 2015, 2016). In addition, the constitutive knockdown of Klf15 in visceral epithelial cells (podocytes) of the glomerulus leads to the loss of stable differentiation markers and key actin cytoskeleton elements leading to podocyte injury and eventual kidney disease in mice (Mallipattu et al., 2016, 2012; Zhong et al., 2015). Interestingly, it was also reported that salutary benefits of glucocorticoids in the kidney are mediated by KLF15 (Mallipattu et al., 2016). Furthermore, it was recently reported that transcriptional regulation of KLF6 enhances mitochondrial cytochrome oxidase activity in the setting of podocyte stress (Mallipattu et al., 2015), and KLF4 increases the expression of genes involved in maintenance of the renal filtration barrier by reducing methylation in their promoter region (Hayashi et al., 2014). Finally, haploinsufficiency of Klf5 in collecting duct cells exacerbates the inflammatory response in murine models of kidney fibrosis (Fujiu et al., 2011). Although the role of KLFs in kidney disease were interrogated in the adult phase, the utilization of constitutive knockout models suggests that changes in KLF expression during kidney development might have a potential impact on the kidney phenotype in adults.
KLFs in neuronal disorders
As mentioned earlier, KLF9 is implicated in neuronal development, and recent studies have also determined that KLF9 regulates neuronal plasticity by mediating the stress-induced effects of glucocorticoids in hippocampal neurons (Bagamasbad et al., 2012; Bonett et al., 2009). In addition, thyroid hormone induced oligodendrocyte differentiation and myelin regeneration are regulated by KLF9, which has direct implications in demyelinating disorders such as multiple sclerosis (Dugas et al., 2012). Furthermore, STAT3-induced axon regeneration is crucial for promoting repair in certain neuronal disorders and, interestingly, it was shown that the loss of KLF4 results in the activation of JAK/STAT3 signaling, thereby leading to regeneration of axons in the setting of injury (Qin et al., 2013). Collectively, these studies suggest that modulating KLF expression contributes to neuronal plasticity, a finding that has implications for furthering our understanding of neuronal disorders and diseases.
Conclusions
In the last decade, there has been a dramatic rise in the number of publications that are centered on KLFs. Genetic knockouts of select members of the KLF family in various model organisms have demonstrated many interesting developmental phenotypes. In addition, several groups have highlighted essential roles for KLFs in cellular regeneration in many tissue types. Finally, as we have summarized here, KLFs have a direct impact on human diseases due to their key roles in development, highlighting potential areas that are ripe for future studies.
Although the field of KLFs has advanced dramatically in the last decade, much more work lies ahead. For instance, the mechanisms by which specific KLFs impact embryogenesis require further exploration. Moreover, the impact of changes in KLF expression during development and in the adult remains a mystery in many organ systems. Finally, as a field we need to clarify the redundant as well as the competitive functions of KLFs, both during development as well as for maintaining a differentiated phenotype in the adult state. In particular, the mechanism by which KLFs contribute to pluripotency and regeneration mandate further studies, especially for neurons and kidney cells that harbor low regenerative potential after injury. Finally, the field remains unclear on whether these transcription factors with diverse roles can serve as a viable target for therapy with minimal off-target effects. To address this, we believe efforts should focus on a system-based interdisciplinary approach combining our understanding of specific KLF functions during normal development with our knowledge of how these functions could be implicated in homeostasis, disease and the prevention of disease.
Acknowledgements
We thank Dr William E. Diehl (University of Massachusetts Medical School) for the analysis of the phylogenetic relationship between human and mouse KLF family members provided in Fig. 2.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Funding
This work was supported by funds from the National Institutes of Health (V.W.Y. and S.K.M.), the American Heart Association (S.K.M.), and Dialysis Clinic, Inc. (S.K.M.). Deposited in PMC for release after 12 months.
References
- Agustian P. A., Bockmeyer C. L., Modde F., Wittig J., Heinemann F. M., Brundiers S., Dämmrich M. E., Schwarz A., Birschmann I., Suwelack B. et al. (2013). Glomerular mRNA expression of prothrombotic and antithrombotic factors in renal transplants with thrombotic microangiopathy. Transplantation 95, 1242-1248. 10.1097/TP.0b013e318291a298 [DOI] [PubMed] [Google Scholar]
- Alhashem Y. N., Vinjamur D. S., Basu M., Klingmuller U., Gaensler K. M. L. and Lloyd J. A. (2011). Transcription factors KLF1 and KLF2 positively regulate embryonic and fetal beta-globin genes through direct promoter binding. J. Biol. Chem. 286, 24819-24827. 10.1074/jbc.M111.247536 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alvarez-Dominguez J. R., Hu W., Yuan B., Shi J., Park S. S., Gromatzky A. A., van Oudenaarden A. and Lodish H. F. (2014). Global discovery of erythroid long noncoding RNAs reveals novel regulators of red cell maturation. Blood 123, 570-581. 10.1182/blood-2013-10-530683 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anderson K. P., Kern C. B., Crable S. C. and Lingrel J. B. (1995). Isolation of a gene encoding a functional zinc finger protein homologous to erythroid Krüppel-like factor: identification of a new multigene family. Mol. Cell. Biol. 15, 5957-5965. 10.1128/MCB.15.11.5957 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aoi T., Yae K., Nakagawa M., Ichisaka T., Okita K., Takahashi K., Chiba T. and Yamanaka S. (2008). Generation of pluripotent stem cells from adult mouse liver and stomach cells. Science 321, 699-702. 10.1126/science.1154884 [DOI] [PubMed] [Google Scholar]
- Asada M., Rauch A., Shimizu H., Maruyama H., Miyaki S., Shibamori M., Kawasome H., Ishiyama H., Tuckermann J. and Asahara H. (2011). DNA binding-dependent glucocorticoid receptor activity promotes adipogenesis via Krüppel-like factor 15 gene expression. Lab. Invest. 91, 203-215. 10.1038/labinvest.2010.170 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asano H., Li X. S. and Stamatoyannopoulos G. (1999). FKLF, a novel Krüppel-like factor that activates human embryonic and fetal beta-like globin genes. Mol. Cell. Biol. 19, 3571-3579. 10.1128/MCB.19.5.3571 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Atkins G. B. and Jain M. K. (2007). Role of Kruppel-like transcription factors in endothelial biology. Circ. Res. 100, 1686-1695. 10.1161/01.RES.0000267856.00713.0a [DOI] [PubMed] [Google Scholar]
- Atkins G. B., Wang Y., Mahabeleshwar G. H., Shi H., Gao H., Kawanami D., Natesan V., Lin Z., Simon D. I. and Jain M. K. (2008). Hemizygous deficiency of Kruppel-like factor 2 augments experimental atherosclerosis. Circ. Res. 103, 690-693. 10.1161/CIRCRESAHA.108.184663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bagamasbad P., Ziera T., Borden S. A., Bonett R. M., Rozeboom A. M., Seasholtz A. and Denver R. J. (2012). Molecular basis for glucocorticoid induction of the Krüppel-like factor 9 gene in hippocampal neurons. Endocrinology 153, 5334-5345. 10.1210/en.2012-1303 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Banck M. S., Beaven S. W., Narla G., Walsh M. J., Friedman S. L. and Beutler A. S. (2006). KLF6 degradation after apoptotic DNA damage. FEBS Lett. 580, 6981-6986. 10.1016/j.febslet.2006.10.077 [DOI] [PubMed] [Google Scholar]
- Basu P., Morris P. E., Haar J. L., Wani M. A., Lingrel J. B., Gaensler K. M. L. and Lloyd J. A. (2005). KLF2 is essential for primitive erythropoiesis and regulates the human and murine embryonic beta-like globin genes in vivo. Blood 106, 2566-2571. 10.1182/blood-2005-02-0674 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basu P., Lung T. K., Lemsaddek W., Sargent T. G., Williams D. C. Jr, Basu M., Redmond L. C., Lingrel J. B., Haar J. L. and Lloyd J. A. (2007). EKLF and KLF2 have compensatory roles in embryonic beta-globin gene expression and primitive erythropoiesis. Blood 110, 3417-3425. 10.1182/blood-2006-11-057307 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechmann L. P., Vetter D., Ishida J., Hannivoort R. A., Lang U. E., Kocabayoglu P., Fiel M. I., Muñoz U., Patman G. L., Ge F. et al. (2013). Post-transcriptional activation of PPAR alpha by KLF6 in hepatic steatosis. J. Hepatol. 58, 1000-1006. 10.1016/j.jhep.2013.01.020 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bensamoun S. F., Hawse J. R., Subramaniam M., Ilharreborde B., Bassillais A., Benhamou C. L., Fraser D. G., Oursler M. J., Amadio P. C., An K.-N. et al. (2006). TGFbeta inducible early gene-1 knockout mice display defects in bone strength and microarchitecture. Bone 39, 1244-1251. 10.1016/j.bone.2006.05.021 [DOI] [PubMed] [Google Scholar]
- Bieker J. J. (2010). Putting a finger on the switch. Nat. Genet. 42, 733-734. 10.1038/ng0910-733 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blackmore M. G., Wang Z., Lerch J. K., Motti D., Zhang Y. P., Shields C. B., Lee J. K., Goldberg J. L., Lemmon V. P. and Bixby J. L. (2012). Kruppel-like Factor 7 engineered for transcriptional activation promotes axon regeneration in the adult corticospinal tract. Proc. Natl. Acad. Sci. USA 109, 7517-7522. 10.1073/pnas.1120684109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blakeley P., Fogarty N. M. E., Del Valle I., Wamaitha S. E., Hu T. X., Elder K., Snell P., Christie L., Robson P. and Niakan K. K. (2015). Defining the three cell lineages of the human blastocyst by single-cell RNA-seq. Development 142, 3613 10.1242/dev.131235 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonett R. M., Hu F., Bagamasbad P. and Denver R. J. (2009). Stressor and glucocorticoid-dependent induction of the immediate early gene Krüppel-like factor 9: implications for neural development and plasticity. Endocrinology 150, 1757-1765. 10.1210/en.2008-1441 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boon R. A., Urbich C., Fischer A., Fontijn R. D., Seeger F. H., Koyanagi M., Horrevoets A. J. G. and Dimmeler S. (2011). Kruppel-like factor 2 improves neovascularization capacity of aged proangiogenic cells. Eur. Heart J. 32, 371-377. 10.1093/eurheartj/ehq137 [DOI] [PubMed] [Google Scholar]
- Bruce S. J., Gardiner B. B., Burke L. J., Gongora M. M., Grimmond S. M. and Perkins A. C. (2007). Dynamic transcription programs during ES cell differentiation towards mesoderm in serum versus serum-freeBMP4 culture. BMC Genomics 8, 365 10.1186/1471-2164-8-365 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buck A., Buchholz M., Wagner M., Adler G., Gress T. and Ellenrieder V. (2006). The tumor suppressor KLF11 mediates a novel mechanism in transforming growth factor beta-induced growth inhibition that is inactivated in pancreatic cancer. Mol. Cancer Res. 4, 861-872. 10.1158/1541-7786.MCR-06-0081 [DOI] [PubMed] [Google Scholar]
- Cai J., Xie D., Fan Z., Chipperfield H., Marden J., Wong W. H. and Zhong S. (2010). Modeling co-expression across species for complex traits: insights to the difference of human and mouse embryonic stem cells. PLoS Comput. Biol. 6, e1000707 10.1371/journal.pcbi.1000707 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cao Z., Wara A. K., Icli B., Sun X., Packard R. R. S., Esen F., Stapleton C. J., Subramaniam M., Kretschmer K., Apostolou I. et al. (2009). Kruppel-like factor KLF10 targets transforming growth factor-beta1 to regulate CD4(+)CD25(−) T cells and T regulatory cells. J. Biol. Chem. 284, 24914-24924. 10.1074/jbc.M109.000059 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carlson C. M., Endrizzi B. T., Wu J., Ding X., Weinreich M. A., Walsh E. R., Wani M. A., Lingrel J. B., Hogquist K. A. and Jameson S. C. (2006). Kruppel-like factor 2 regulates thymocyte and T-cell migration. Nature 442, 299-302. 10.1038/nature04882 [DOI] [PubMed] [Google Scholar]
- Chaib H., Cockrell E. K., Rubin M. A. and Macoska J. A. (2001). Profiling and verification of gene expression patterns in normal and malignant human prostate tissues by cDNA microarray analysis. Neoplasia 3, 43-52. 10.1038/sj.neo.7900126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen X. and Bieker J. J. (1996). Erythroid Kruppel-like factor (EKLF) contains a multifunctional transcriptional activation domain important for inter- and intramolecular interactions. EMBO J. 15, 5888-5896. [PMC free article] [PubMed] [Google Scholar]
- Chen C., Bhalala H. V., Qiao H. and Dong J.-T. (2002). A possible tumor suppressor role of the KLF5 transcription factor in human breast cancer. Oncogene 21, 6567-6572. 10.1038/sj.onc.1205817 [DOI] [PubMed] [Google Scholar]
- Chen C., Hyytinen E.-R., Sun X., Helin H. J., Koivisto P. A., Frierson H. F. Jr, Vessella R. L. and Dong J.-T. (2003). Deletion, mutation, and loss of expression of KLF6 in human prostate cancer. Am. J. Pathol. 162, 1349-1354. 10.1016/S0002-9440(10)63930-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen C., Sun X., Ran Q., Wilkinson K. D., Murphy T. J., Simons J. W. and Dong J.-T. (2005a). Ubiquitin-proteasome degradation of KLF5 transcription factor in cancer and untransformed epithelial cells. Oncogene 24, 3319-3327. 10.1038/sj.onc.1208497 [DOI] [PubMed] [Google Scholar]
- Chen Z. Y., Wang X., Zhou Y., Offner G. and Tseng C.-C. (2005b). Destabilization of Kruppel-like factor 4 protein in response to serum stimulation involves the ubiquitin-proteasome pathway. Cancer Res. 65, 10394-10400. 10.1158/0008-5472.CAN-05-2059 [DOI] [PubMed] [Google Scholar]
- Chiplunkar A. R., Curtis B. C., Eades G. L., Kane M. S., Fox S. J., Haar J. L. and Lloyd J. A. (2013a). The Krüppel-like factor 2 and Krüppel-like factor 4 genes interact to maintain endothelial integrity in mouse embryonic vasculogenesis. BMC Dev. Biol. 13, 40 10.1186/1471-213X-13-40 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiplunkar A. R., Lung T. K., Alhashem Y., Koppenhaver B. A., Salloum F. N., Kukreja R. C., Haar J. L. and Lloyd J. A. (2013b). Krüppel-like factor 2 is required for normal mouse cardiac development. PLoS ONE 8, e54891 10.1371/journal.pone.0054891 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chopra M., Langenhorst D., Beilhack A., Serfling E. and Patra A. K. (2015). Interleukin-2 critically regulates bone marrow erythropoiesis and prevents anemia development. Eur. J. Immunol. 45, 3362-3374. 10.1002/eji.201545596 [DOI] [PubMed] [Google Scholar]
- Conkright M. D., Wani M. A., Anderson K. P. and Lingrel J. B. (1999). A gene encoding an intestinal-enriched member of the Kruppel-like factor family expressed in intestinal epithelial cells. Nucleic Acids Res. 27, 1263-1270. 10.1093/nar/27.5.1263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conkright M. D., Wani M. A. and Lingrel J. B. (2001). Lung Kruppel-like factor contains an autoinhibitory domain that regulates its transcriptional activation by binding WWP1, an E3 ubiquitin ligase. J. Biol. Chem. 276, 29299-29306. 10.1074/jbc.M103670200 [DOI] [PubMed] [Google Scholar]
- Cook T., Gebelein B., Mesa K., Mladek A. and Urrutia R. (1998). Molecular cloning and characterization of TIEG2 reveals a new subfamily of transforming growth factor-beta-inducible Sp1-like zinc finger-encoding genes involved in the regulation of cell growth. J. Biol. Chem. 273, 25929-25936. 10.1074/jbc.273.40.25929 [DOI] [PubMed] [Google Scholar]
- Cook T., Gebelein B., Belal M., Mesa K. and Urrutia R. (1999). Three conserved transcriptional repressor domains are a defining feature of the TIEG subfamily of Sp1-like zinc finger proteins. J. Biol. Chem. 274, 29500-29504. 10.1074/jbc.274.41.29500 [DOI] [PubMed] [Google Scholar]
- Crossley M., Whitelaw E., Perkins A., Williams G., Fujiwara Y. and Orkin S. H. (1996). Isolation and characterization of the cDNA encoding BKLF/TEF-2, a major CACCC-box-binding protein in erythroid cells and selected other cells. Mol. Cell. Biol. 16, 1695-1705. 10.1128/MCB.16.4.1695 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daftary G. S., Lomberk G. A., Buttar N. S., Allen T. W., Grzenda A., Zhang J., Zheng Y., Mathison A. J., Gada R. P., Calvo E. et al. (2012). Detailed structural-functional analysis of the Kruppel-like factor 16 (KLF16) transcription factor reveals novel mechanisms for silencing Sp/KLF sites involved in metabolism and endocrinology. J. Biol. Chem. 287, 7010-7025. 10.1074/jbc.M111.266007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denver R. J., Ouellet L., Furling D., Kobayashi A., Fujii-Kuriyama Y. and Puymirat J. (1999). Basic transcription element-binding protein (BTEB) is a thyroid hormone-regulated gene in the developing central nervous system. Evidence for a role in neurite outgrowth. J. Biol. Chem. 274, 23128-23134. 10.1074/jbc.274.33.23128 [DOI] [PubMed] [Google Scholar]
- Dewi V., Kwok A., Lee S., Lee M. M., Tan Y. M., Nicholas H. R., Isono K.-I., Wienert B., Mak K. S., Knights A. J. et al. (2015). Phosphorylation of Krüppel-like factor 3 (KLF3/BKLF) and C-terminal binding protein 2 (CtBP2) by homeodomain-interacting protein kinase 2 (HIPK2) modulates KLF3 DNA binding and activity. J. Biol. Chem. 290, 8591-8605. 10.1074/jbc.M115.638338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Difeo A., Huang F., Sangodkar J., Terzo E. A., Leake D., Narla G. and Martignetti J. A. (2009). KLF6-SV1 is a novel antiapoptotic protein that targets the BH3-only protein NOXA for degradation and whose inhibition extends survival in an ovarian cancer model. Cancer Res. 69, 4733-4741. 10.1158/0008-5472.CAN-08-4282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drissen R., von Lindern M., Kolbus A., Driegen S., Steinlein P., Beug H., Grosveld F. and Philipsen S. (2005). The erythroid phenotype of EKLF-null mice: defects in hemoglobin metabolism and membrane stability. Mol. Cell. Biol. 25, 5205-5214. 10.1128/MCB.25.12.5205-5214.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Souza U. M., Lammers C.-H., Hwang C. K., Yajima S. and Mouradian M. M. (2002). Developmental expression of the zinc finger transcription factor DRRF (dopamine receptor regulating factor). Mech. Dev. 110, 197-201. 10.1016/S0925-4773(01)00564-0 [DOI] [PubMed] [Google Scholar]
- Du J. X., Yun C. C., Bialkowska A. and Yang V. W. (2007). Protein inhibitor of activated STAT1 interacts with and up-regulates activities of the pro-proliferative transcription factor Kruppel-like factor 5. J. Biol. Chem. 282, 4782-4793. 10.1074/jbc.M603413200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du J. X., Bialkowska A. B., McConnell B. B. and Yang V. W. (2008). SUMOylation regulates nuclear localization of Kruppel-like factor 5. J. Biol. Chem. 283, 31991-32002. 10.1074/jbc.M803612200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dugas J. C., Ibrahim A. and Barres B. A. (2012). The T3-induced gene KLF9 regulates oligodendrocyte differentiation and myelin regeneration. Mol. Cell. Neurosci. 50, 45-57. 10.1016/j.mcn.2012.03.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehlermann J., Pfisterer P. and Schorle H. (2003). Dynamic expression of Krüppel-like factor 4 (Klf4), a target of transcription factor AP-2alpha during murine mid-embryogenesis. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 273A, 677-680. 10.1002/ar.a.10089 [DOI] [PubMed] [Google Scholar]
- Ellenrieder V., Buck A., Harth A., Jungert K., Buchholz M., Adler G., Urrutia R. and Gress T. M. (2004). KLF11 mediates a critical mechanism in TGF-beta signaling that is inactivated by Erk-MAPK in pancreatic cancer cells. Gastroenterology 127, 607-620. 10.1053/j.gastro.2004.05.018 [DOI] [PubMed] [Google Scholar]
- Ema M., Mori D., Niwa H., Hasegawa Y., Yamanaka Y., Hitoshi S., Mimura J., Kawabe Y.-I., Hosoya T., Morita M. et al. (2008). Krüppel-like factor 5 is essential for blastocyst development and the normal self-renewal of mouse ESCs. Cell Stem Cell 3, 555-567. 10.1016/j.stem.2008.09.003 [DOI] [PubMed] [Google Scholar]
- Evans P. M., Zhang W., Chen X., Yang J., Bhakat K. K. and Liu C. (2007). Kruppel-like factor 4 is acetylated by p300 and regulates gene transcription via modulation of histone acetylation. J. Biol. Chem. 282, 33994-34002. 10.1074/jbc.M701847200 [DOI] [PubMed] [Google Scholar]
- Feinberg M. W., Wara A. K., Cao Z., Lebedeva M. A., Rosenbauer F., Iwasaki H., Hirai H., Katz J. P., Haspel R. L., Gray S. et al. (2007). The Kruppel-like factor KLF4 is a critical regulator of monocyte differentiation. EMBO J. 26, 4138-4148. 10.1038/sj.emboj.7601824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisch S., Gray S., Heymans S., Haldar S. M., Wang B., Pfister O., Cui L., Kumar A., Lin Z., Sen-Banerjee S. et al. (2007). Kruppel-like factor 15 is a regulator of cardiomyocyte hypertrophy. Proc. Natl. Acad. Sci. USA 104, 7074-7079. 10.1073/pnas.0701981104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer E. A., Verpont M. C., Garrett-Sinha L. A., Ronco P. M. and Rossert J. A. (2001). Klf6 is a zinc finger protein expressed in a cell-specific manner during kidney development. J. Am. Soc. Nephrol. 12, 726-735. [DOI] [PubMed] [Google Scholar]
- Friedman S. L. (2000). Molecular regulation of hepatic fibrosis, an integrated cellular response to tissue injury. J. Biol. Chem. 275, 2247-2250. 10.1074/jbc.275.4.2247 [DOI] [PubMed] [Google Scholar]
- Frontelo P., Manwani D., Galdass M., Karsunky H., Lohmann F., Gallagher P. G. and Bieker J. J. (2007). Novel role for EKLF in megakaryocyte lineage commitment. Blood 110, 3871-3880. 10.1182/blood-2007-03-082065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujiu K., Manabe I. and Nagai R. (2011). Renal collecting duct epithelial cells regulate inflammation in tubulointerstitial damage in mice. J. Clin. Invest. 121, 3425-3441. 10.1172/JCI57582 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Funnell A. P. W., Mak K. S., Twine N. A., Pelka G. J., Norton L. J., Radziewic T., Power M., Wilkins M. R., Bell-Anderson K. S., Fraser S. T. et al. (2013). Generation of mice deficient in both KLF3/BKLF and KLF8 reveals a genetic interaction and a role for these factors in embryonic globin gene silencing. Mol. Cell. Biol. 33, 2976-2987. 10.1128/MCB.00074-13 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gallardo-Vara E., Blanco F. J., Roqué M., Friedman S. L., Suzuki T., Botella L. M. and Bernabeu C. (2016). Transcription factor KLF6 upregulates expression of metalloprotease MMP14 and subsequent release of soluble endoglin during vascular injury. Angiogenesis 19, 155-171. 10.1007/s10456-016-9495-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrido-Martin E. M., Blanco F. J., Roque M., Novensa L., Tarocchi M., Lang U. E., Suzuki T., Friedman S. L., Botella L. M. and Bernabeu C. (2013). Vascular injury triggers Kruppel-like factor 6 mobilization and cooperation with specificity protein 1 to promote endothelial activation through upregulation of the activin receptor-like kinase 1 gene. Circ. Res. 112, 113-127. 10.1161/CIRCRESAHA.112.275586 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ge F., Chen W., Qin J., Zhou Z., Liu R., Liu L., Tan J., Zou T., Li H., Ren G. et al. (2015). Ataxin-3 like (ATXN3L), a member of the Josephin family of deubiquitinating enzymes, promotes breast cancer proliferation by deubiquitinating Krüppel-like factor 5 (KLF5). Oncotarget 6, 21369-21378. 10.18632/oncotarget.4128 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Geiman D. E., Ton-That H., Johnson J. M. and Yang V. W. (2000). Transactivation and growth suppression by the gut-enriched Kruppel-like factor (Kruppel-like factor 4) are dependent on acidic amino acid residues and protein-protein interaction. Nucleic Acids Res. 28, 1106-1113. 10.1093/nar/28.5.1106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghaleb A. M., Nandan M. O., Chanchevalap S., Dalton W. B., Hisamuddin I. M. and Yang V. W. (2005). Krüppel-like factors 4 and 5: the yin and yang regulators of cellular proliferation. Cell Res. 15, 92-96. 10.1038/sj.cr.7290271 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gordon A. R., Outram S. V., Keramatipour M., Goddard C. A., Colledge W. H., Metcalfe J. C., Hager-Theodorides A. L., Crompton T. and Kemp P. R. (2008). Splenomegaly and modified erythropoiesis in KLF13−/− mice. J. Biol. Chem. 283, 11897-11904. 10.1074/jbc.M709569200 [DOI] [PubMed] [Google Scholar]
- Gray S., Wang B., Orihuela Y., Hong E.-G., Fisch S., Haldar S., Cline G. W., Kim J. K., Peroni O. D., Kahn B. B. et al. (2007). Regulation of gluconeogenesis by Krüppel-like factor 15. Cell Metab. 5, 305-312. 10.1016/j.cmet.2007.03.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haldar S. M., Lu Y., Jeyaraj D., Kawanami D., Cui Y., Eapen S. J., Hao C., Li Y., Doughman Y.-Q., Watanabe M. et al. (2010). Klf15 deficiency is a molecular link between heart failure and aortic aneurysm formation. Sci. Transl. Med. 2, 26ra26 10.1126/scitranslmed.3000502 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall J., Guo G., Wray J., Eyres I., Nichols J., Grotewold L., Morfopoulou S., Humphreys P., Mansfield W., Walker R. et al. (2009). Oct4 and LIF/Stat3 additively induce Krüppel factors to sustain embryonic stem cell self-renewal. Cell Stem Cell 5, 597-609. 10.1016/j.stem.2009.11.003 [DOI] [PubMed] [Google Scholar]
- Hamik A., Lin Z., Kumar A., Balcells M., Sinha S., Katz J., Feinberg M. W., Gerszten R. E., Edelman E. R. and Jain M. K. (2007). Kruppel-like factor 4 regulates endothelial inflammation. J. Biol. Chem. 282, 13769-13779. 10.1074/jbc.M700078200 [DOI] [PubMed] [Google Scholar]
- Hanna J., Cheng A. W., Saha K., Kim J., Lengner C. J., Soldner F., Cassady J. P., Muffat J., Carey B. W. and Jaenisch R. (2010). Human embryonic stem cells with biological and epigenetic characteristics similar to those of mouse ESCs. Proc. Natl. Acad. Sci. USA 107, 9222-9227. 10.1073/pnas.1004584107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawse J. R., Iwaniec U. T., Bensamoun S. F., Monroe D. G., Peters K. D., Ilharreborde B., Rajamannan N. M., Oursler M. J., Turner R. T., Spelsberg T. C. et al. (2008). TIEG-null mice display an osteopenic gender-specific phenotype. Bone 42, 1025-1031. 10.1016/j.bone.2008.02.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi K., Sasamura H., Nakamura M., Azegami T., Oguchi H., Sakamaki Y. and Itoh H. (2014). KLF4-dependent epigenetic remodeling modulates podocyte phenotypes and attenuates proteinuria. J. Clin. Invest. 124, 2523-2537. 10.1172/JCI69557 [DOI] [PMC free article] [PubMed] [Google Scholar]
- He M., Han M., Zheng B., Shu Y.-N. and Wen J.-K. (2009). Angiotensin II stimulates KLF5 phosphorylation and its interaction with c-Jun leading to suppression of p21 expression in vascular smooth muscle cells. J. Biochem. 146, 683-691. 10.1093/jb/mvp115 [DOI] [PubMed] [Google Scholar]
- Hiramatsu K., Sasagawa S., Outani H., Nakagawa K., Yoshikawa H. and Tsumaki N. (2011). Generation of hyaline cartilaginous tissue from mouse adult dermal fibroblast culture by defined factors. J. Clin. Invest. 121, 640-657. 10.1172/JCI44605 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hodge D., Coghill E., Keys J., Maguire T., Hartmann B., McDowall A., Weiss M., Grimmond S. and Perkins A. (2006). A global role for EKLF in definitive and primitive erythropoiesis. Blood 107, 3359-3370. 10.1182/blood-2005-07-2888 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu D., Gur M., Zhou Z., Gamper A., Hung M.-C., Fujita N., Lan L., Bahar I. and Wan Y. (2015). Interplay between arginine methylation and ubiquitylation regulates KLF4-mediated genome stability and carcinogenesis. Nat. Commun. 6, 8419 10.1038/ncomms9419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang B., Ahn Y.-T., McPherson L., Clayberger C. and Krensky A. M. (2007). Interaction of PRP4 with Kruppel-like factor 13 regulates CCL5 transcription. J. Immunol. 178, 7081-7087. 10.4049/jimmunol.178.11.7081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang C. K., D'Souza U. M., Eisch A. J., Yajima S., Lammers C.-H., Yang Y., Lee S.-H., Kim Y.-M., Nestler E. J. and Mouradian M. M. (2001). Dopamine receptor regulating factor, DRRF: a zinc finger transcription factor. Proc. Natl. Acad. Sci. USA 98, 7558-7563. 10.1073/pnas.121635798 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hwang Y.-C., Yang C.-H., Lin C.-H., Ch'ang H.-J., Chang V. H. S. and Yu W. C. Y. (2013). Destabilization of KLF10, a tumor suppressor, relies on thr93 phosphorylation and isomerase association. Biochim. Biophys. Acta 1833, 3035-3045. 10.1016/j.bbamcr.2013.08.010 [DOI] [PubMed] [Google Scholar]
- Imhof A., Schuierer M., Werner O., Moser M., Roth C., Bauer R. and Buettner R. (1999). Transcriptional regulation of the AP-2alpha promoter by BTEB-1 and AP-2rep, a novel wt-1/egr-related zinc finger repressor. Mol. Cell. Biol. 19, 194-204. 10.1128/MCB.19.1.194 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaubert J., Cheng J. and Segre J. A. (2003). Ectopic expression of kruppel like factor 4 (Klf4) accelerates formation of the epidermal permeability barrier. Development 130, 2767-2777. 10.1242/dev.00477 [DOI] [PubMed] [Google Scholar]
- Jeon H., Waku T., Azami T., Khoa L. T. P., Yanagisawa J., Takahashi S. and Ema M. (2016). Comprehensive identification of Krüppel-like factor family members contributing to the self-renewal of mouse embryonic stem cells and cellular reprogramming. PLoS ONE 11, e0150715 10.1371/journal.pone.0150715 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeyaraj D., Haldar S. M., Wan X., McCauley M. D., Ripperger J. A., Hu K., Lu Y., Eapen B. L., Sharma N., Ficker E. et al. (2012). Circadian rhythms govern cardiac repolarization and arrhythmogenesis. Nature 483, 96-99. 10.1038/nature10852 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang J., Chan Y.-S., Loh Y.-H., Cai J., Tong G.-Q., Lim C.-A., Robson P., Zhong S. and Ng H.-H. (2008). A core Klf circuitry regulates self-renewal of embryonic stem cells. Nat. Cell Biol. 10, 353-360. 10.1038/ncb1698 [DOI] [PubMed] [Google Scholar]
- Kaczynski J., Zhang J.-S., Ellenrieder V., Conley A., Duenes T., Kester H., van Der Burg B. and Urrutia R. (2001). The Sp1-like protein BTEB3 inhibits transcription via the basic transcription element box by interacting with mSin3A and HDAC-1 co-repressors and competing with Sp1. J. Biol. Chem. 276, 36749-36756. 10.1074/jbc.M105831200 [DOI] [PubMed] [Google Scholar]
- Kaczynski J. A., Conley A. A., Fernandez Zapico M., Delgado S. M., Zhang J.-S. and Urrutia R. (2002). Functional analysis of basic transcription element (BTE)-binding protein (BTEB) 3 and BTEB4, a novel Sp1-like protein, reveals a subfamily of transcriptional repressors for the BTE site of the cytochrome P4501A1 gene promoter. Biochem. J. 366, 873-882. 10.1042/bj20020388 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kajimura D., Dragomir C., Ramirez F. and Laub F. (2007). Identification of genes regulated by transcription factor KLF7 in differentiating olfactory sensory neurons. Gene 388, 34-42. 10.1016/j.gene.2006.09.027 [DOI] [PubMed] [Google Scholar]
- Kanazawa A., Kawamura Y., Sekine A., Iida A., Tsunoda T., Kashiwagi A., Tanaka Y., Babazono T., Matsuda M., Kawai K. et al. (2005). Single nucleotide polymorphisms in the gene encoding Krüppel-like factor 7 are associated with type 2 diabetes. Diabetologia 48, 1315-1322. 10.1007/s00125-005-1797-0 [DOI] [PubMed] [Google Scholar]
- Katoh K., Misawa K., Kuma K.-I. and Miyata T. (2002). MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 30, 3059-3066. 10.1093/nar/gkf436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz J. P., Perreault N., Goldstein B. G., Lee C. S., Labosky P. A., Yang V. W. and Kaestner K. H. (2002). The zinc-finger transcription factor Klf4 is required for terminal differentiation of goblet cells in the colon. Development 129, 2619-2628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawada N. (2011). Evolution of hepatic fibrosis research. Hepatol. Res. 41, 199-208. 10.1111/j.1872-034X.2011.00776.x [DOI] [PubMed] [Google Scholar]
- Kawai-Kowase K., Kumar M. S., Hoofnagle M. H., Yoshida T. and Owens G. K. (2005). PIAS1 activates the expression of smooth muscle cell differentiation marker genes by interacting with serum response factor and class I basic helix-loop-helix proteins. Mol. Cell. Biol. 25, 8009-8023. 10.1128/MCB.25.18.8009-8023.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kawamura Y., Tanaka Y., Kawamori R. and Maeda S. (2006). Overexpression of Kruppel-like factor 7 regulates adipocytokine gene expressions in human adipocytes and inhibits glucose-induced insulin secretion in pancreatic beta-cell line. Mol. Endocrinol. 20, 844-856. 10.1210/me.2005-0138 [DOI] [PubMed] [Google Scholar]
- Kearse M., Moir R., Wilson A., Stones-Havas S., Cheung M., Sturrock S., Buxton S., Cooper A., Markowitz S., Duran C. et al. (2012). Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28, 1647-1649. 10.1093/bioinformatics/bts199 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keller G. (2005). Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes Dev. 19, 1129-1155. 10.1101/gad.1303605 [DOI] [PubMed] [Google Scholar]
- Kelsey L., Flenniken A. M., Qu D., Funnell A. P. W., Pearson R., Zhou Y.-Q., Voronina I., Berberovic Z., Wood G., Newbigging S. et al. (2013). ENU-induced mutation in the DNA-binding domain of KLF3 reveals important roles for KLF3 in cardiovascular development and function in mice. PLoS Genet. 9, e1003612 10.1371/journal.pgen.1003612 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim J. B., Zaehres H., Wu G., Gentile L., Ko K., Sebastiano V., Araúzo-Bravo M. J., Ruau D., Han D. W., Zenke M. et al. (2008). Pluripotent stem cells induced from adult neural stem cells by reprogramming with two factors. Nature 454, 646-650. 10.1038/nature07061 [DOI] [PubMed] [Google Scholar]
- Kim D. S., Zhang W., Millman S. E., Hwang B. J., Kwon S. J., Clayberger C., Pagano M. and Krensky A. M. (2012a). Fbw7gamma-mediated degradation of KLF13 prevents RANTES expression in resting human but not murine T lymphocytes. Blood 120, 1658-1667. 10.1182/blood-2012-03-415968 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim M. O., Kim S.-H., Cho Y.-Y., Nadas J., Jeong C.-H., Yao K., Kim D. J., Yu D.-H., Keum Y.-S., Lee K.-Y. et al. (2012b). ERK1 and ERK2 regulate embryonic stem cell self-renewal through phosphorylation of Klf4. Nat. Struct. Mol. Biol. 19, 283-290. 10.1038/nsmb.2217 [DOI] [PubMed] [Google Scholar]
- Kitada M., Wakao S. and Dezawa M. (2012). Muse cells and induced pluripotent stem cell: implication of the elite model. Cell. Mol. Life Sci. 69, 3739-3750. 10.1007/s00018-012-0994-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kojima S., Kobayashi A., Gotoh O., Ohkuma Y., Fujii-Kuriyama Y. and Sogawa K. (1997). Transcriptional activation domain of human BTEB2, a GC box-binding factor. J. Biochem. 121, 389-396. 10.1093/oxfordjournals.jbchem.a021600 [DOI] [PubMed] [Google Scholar]
- Kong X., Li L., Li Z., Le X., Huang C., Jia Z., Cui J., Huang S., Wang L. and Xie K. (2013). Dysregulated expression of FOXM1 isoforms drives progression of pancreatic cancer. Cancer Res. 73, 3987-3996. 10.1158/0008-5472.CAN-12-3859 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koritschoner N. P., Bocco J. L., Panzetta-Dutari G. M., Dumur C. I., Flury A. and Patrito L. C. (1997). A novel human zinc finger protein that interacts with the core promoter element of a TATA box-less gene. J. Biol. Chem. 272, 9573-9580. 10.1074/jbc.272.14.9573 [DOI] [PubMed] [Google Scholar]
- Kuo C. T., Veselits M. L., Barton K. P., Lu M. M., Clendenin C. and Leiden J. M. (1997a). The LKLF transcription factor is required for normal tunica media formation and blood vessel stabilization during murine embryogenesis. Genes Dev. 11, 2996-3006. 10.1101/gad.11.22.2996 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuo C. T., Veselits M. L. and Leiden J. M. (1997b). LKLF: a transcriptional regulator of single-positive T cell quiescence and survival. Science 277, 1986-1990. 10.1126/science.277.5334.1986 [DOI] [PubMed] [Google Scholar]
- Kurotaki D., Osato N., Nishiyama A., Yamamoto M., Ban T., Sato H., Nakabayashi J., Umehara M., Miyake N., Matsumoto N. et al. (2013). Essential role of the IRF8-KLF4 transcription factor cascade in murine monocyte differentiation. Blood 121, 1839-1849. 10.1182/blood-2012-06-437863 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lahiri S. K. and Zhao J. (2012). Kruppel-like factor 8 emerges as an important regulator of cancer. Am. J. Transl. Res. 4, 357-363. [PMC free article] [PubMed] [Google Scholar]
- Lahiri S. K., Lu H., Mukherjee D., Yu L. and Zhao J. (2016). ERK2 phosphorylates Kruppel-like factor 8 protein at serine 48 to maintain its stability. Am. J. Cancer Res. 6, 910-923. [PMC free article] [PubMed] [Google Scholar]
- Lang U. E., Kocabayoglu P., Cheng G. Z., Ghiassi-Nejad Z., Muñoz U., Vetter D., Eckstein D. A., Hannivoort R. A., Walsh M. J. and Friedman S. L. (2013). GSK3beta phosphorylation of the KLF6 tumor suppressor promotes its transactivation of p21. Oncogene 32, 4557-4564. 10.1038/onc.2012.457 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laub F., Aldabe R., Friedrich V. Jr, Ohnishi S., Yoshida T. and Ramirez F. (2001a). Developmental expression of mouse Krüppel-like transcription factor KLF7 suggests a potential role in neurogenesis. Dev. Biol. 233, 305-318. 10.1006/dbio.2001.0243 [DOI] [PubMed] [Google Scholar]
- Laub F., Aldabe R., Ramirez F. and Friedman S. (2001b). Embryonic expression of Krüppel-like factor 6 in neural and non-neural tissues. Mech. Dev. 106, 167-170. 10.1016/S0925-4773(01)00419-1 [DOI] [PubMed] [Google Scholar]
- Laub F., Lei L., Sumiyoshi H., Kajimura D., Dragomir C., Smaldone S., Puche A. C., Petros T. J., Mason C., Parada L. F. et al. (2005). Transcription factor KLF7 is important for neuronal morphogenesis in selected regions of the nervous system. Mol. Cell. Biol. 25, 5699-5711. 10.1128/MCB.25.13.5699-5711.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laub F., Dragomir C. and Ramirez F. (2006). Mice without transcription factor KLF7 provide new insight into olfactory bulb development. Brain Res. 1103, 108-113. 10.1016/j.brainres.2006.05.065 [DOI] [PubMed] [Google Scholar]
- Lavallée G., Andelfinger G., Nadeau M., Lefebvre C., Nemer G., Horb M. E. and Nemer M. (2006). The Kruppel-like transcription factor KLF13 is a novel regulator of heart development. EMBO J. 25, 5201-5213. 10.1038/sj.emboj.7601379 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee J. S., Yu Q., Shin J. T., Sebzda E., Bertozzi C., Chen M., Mericko P., Stadtfeld M., Zhou D., Cheng L. et al. (2006). Klf2 is an essential regulator of vascular hemodynamic forces in vivo. Dev. Cell 11, 845-857. 10.1016/j.devcel.2006.09.006 [DOI] [PubMed] [Google Scholar]
- Leenders J. J., Wijnen W. J., van der Made I., Hiller M., Swinnen M., Vandendriessche T., Chuah M., Pinto Y. M. and Creemers E. E. (2012). Repression of cardiac hypertrophy by KLF15: underlying mechanisms and therapeutic implications. PLoS ONE 7, e36754 10.1371/journal.pone.0036754 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lei L., Ma L., Nef S., Thai T. and Parada L. F. (2001). mKlf7, a potential transcriptional regulator of TrkA nerve growth factor receptor expression in sensory and sympathetic neurons. Development 128, 1147-1158. [DOI] [PubMed] [Google Scholar]
- Li D., Yea S., Dolios G., Martignetti J. A., Narla G., Wang R., Walsh M. J. and Friedman S. L. (2005). Regulation of Kruppel-like factor 6 tumor suppressor activity by acetylation. Cancer Res. 65, 9216-9225. 10.1158/0008-5472.CAN-05-1040 [DOI] [PubMed] [Google Scholar]
- Li H.-Y., Chien Y., Chen Y.-J., Chen S.-F., Chang Y.-L., Chiang C.-H., Jeng S.-Y., Chang C.-M., Wang M.-L., Chen L.-K. et al. (2011). Reprogramming induced pluripotent stem cells in the absence of c-Myc for differentiation into hepatocyte-like cells. Biomaterials 32, 5994-6005. 10.1016/j.biomaterials.2011.05.009 [DOI] [PubMed] [Google Scholar]
- Li J., Zheng H., Wang J., Yu F., Morris R. J., Wang T. C., Huang S. and Ai W. (2012). Expression of Kruppel-like factor KLF4 in mouse hair follicle stem cells contributes to cutaneous wound healing. PLoS ONE 7, e39663 10.1371/journal.pone.0039663 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao X., Haldar S. M., Lu Y., Jeyaraj D., Paruchuri K., Nahori M., Cui Y., Kaestner K. H. and Jain M. K. (2010). Krüppel-like factor 4 regulates pressure-induced cardiac hypertrophy. J. Mol. Cell. Cardiol. 49, 334-338. 10.1016/j.yjmcc.2010.04.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liao X., Zhang R., Lu Y., Prosdocimo D. A., Sangwung P., Zhang L., Zhou G., Anand P., Lai L., Leone T. C. et al. (2015). Kruppel-like factor 4 is critical for transcriptional control of cardiac mitochondrial homeostasis. J. Clin. Invest. 125, 3461-3476. 10.1172/JCI79964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin S.-C. J., Wani M. A., Whitsett J. A. and Wells J. M. (2010). Klf5 regulates lineage formation in the pre-implantation mouse embryo. Development 137, 3953-3963. 10.1242/dev.054775 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu N., Li H., Li S., Shen M., Xiao N., Chen Y., Wang Y., Wang W., Wang R., Wang Q. et al. (2010). The Fbw7/human CDC4 tumor suppressor targets proproliferative factor KLF5 for ubiquitination and degradation through multiple phosphodegron motifs. J. Biol. Chem. 285, 18858-18867. 10.1074/jbc.M109.099440 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu R., Zhou Z., Zhao D. and Chen C. (2011). The induction of KLF5 transcription factor by progesterone contributes to progesterone-induced breast cancer cell proliferation and dedifferentiation. Mol. Endocrinol. 25, 1137-1144. 10.1210/me.2010-0497 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lomberk G. and Urrutia R. (2005). The family feud: turning off Sp1 by Sp1-like KLF proteins. Biochem. J. 392, 1-11. 10.1042/BJ20051234 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lomberk G., Grzenda A., Mathison A., Escande C., Zhang J.-S., Calvo E., Miller L. J., Iovanna J., Chini E. N., Fernandez-Zapico M. E. et al. (2013). Kruppel-like factor 11 regulates the expression of metabolic genes via an evolutionarily conserved protein interaction domain functionally disrupted in maturity onset diabetes of the young. J. Biol. Chem. 288, 17745-17758. 10.1074/jbc.M112.434670 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu Y., Zhang L., Liao X., Sangwung P., Prosdocimo D. A., Zhou G., Votruba A. R., Brian L., Han Y. J., Gao H. et al. (2013). Kruppel-like factor 15 is critical for vascular inflammation. J. Clin. Invest. 123, 4232-4241. 10.1172/JCI68552 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maffioletti S. M., Gerli M. F. M., Ragazzi M., Dastidar S., Benedetti S., Loperfido M., VandenDriessche T., Chuah M. K. and Tedesco F. S. (2015). Efficient derivation and inducible differentiation of expandable skeletal myogenic cells from human ES and patient-specific iPS cells. Nat. Protoc. 10, 941-958. 10.1038/nprot.2015.057 [DOI] [PubMed] [Google Scholar]
- Mallipattu S. K., Liu R., Zheng F., Narla G., Ma'ayan A., Dikman S., Jain M. K., Saleem M., D'Agati V., Klotman P. et al. (2012). Krüppel-Like factor 15 (KLF15) is a key regulator of podocyte differentiation. J. Biol. Chem. 287, 19122-19135. 10.1074/jbc.m112.345983 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallipattu S. K., Horne S. J., D'Agati V., Narla G., Liu R., Frohman M. A., Dickman K., Chen E. Y., Ma'ayan A., Bialkowska A. B. et al. (2015). Krüppel-like factor 6 regulates mitochondrial function in the kidney. J. Clin. Invest. 125, 1347-1361. 10.1172/JCI77084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallipattu S. K., Guo Y., Revelo M. P., Roa-Pena L., Miller T., Ling J., Shankland S. J., Bialkowska A. B., Ly V., Estrada C. et al. (2016). Kruppel-like factor 15 mediates glucocorticoid-induced restoration of podocyte differentiation markers. J. Am. Soc. Nephrol. 28, 166-184. 10.1681/ASN.2015060672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansour A. A. F., Gafni O., Weinberger L., Zviran A., Ayyash M., Rais Y., Krupalnik V., Zerbib M., Amann-Zalcenstein D., Maza I. et al. (2012). The H3K27 demethylase Utx regulates somatic and germ cell epigenetic reprogramming. Nature 488, 409-413. 10.1038/nature11272 [DOI] [PubMed] [Google Scholar]
- Martin K. M., Metcalfe J. C. and Kemp P. R. (2001). Expression of Klf9 and Klf13 in mouse development. Mech. Dev. 103, 149-151. 10.1016/S0925-4773(01)00343-4 [DOI] [PubMed] [Google Scholar]
- Mas C., Lussier-Price M., Soni S., Morse T., Arseneault G., Di Lello P., Lafrance-Vanasse J., Bieker J. J. and Omichinski J. G. (2011). Structural and functional characterization of an atypical activation domain in erythroid Kruppel-like factor (EKLF). Proc. Natl. Acad. Sci. USA 108, 10484-10489. 10.1073/pnas.1017029108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathison A., Grzenda A., Lomberk G., Velez G., Buttar N., Tietz P., Hendrickson H., Liebl A., Xiong Y. Y., Gores G. et al. (2013). Role for Krüppel-like transcription factor 11 in mesenchymal cell function and fibrosis. PLoS ONE 8, e75311 10.1371/journal.pone.0075311 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsumoto N., Laub F., Aldabe R., Zhang W., Ramirez F., Yoshida T. and Terada M. (1998). Cloning the cDNA for a new human zinc finger protein defines a group of closely related Kruppel-like transcription factors. J. Biol. Chem. 273, 28229-28237. 10.1074/jbc.273.43.28229 [DOI] [PubMed] [Google Scholar]
- Matsumoto N., Kubo A., Liu H., Akita K., Laub F., Ramirez F., Keller G. and Friedman S. L. (2006). Developmental regulation of yolk sac hematopoiesis by Kruppel-like factor 6. Blood 107, 1357-1365. 10.1182/blood-2005-05-1916 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell B. B. and Yang V. W. (2010). Mammalian Kruppel-like factors in health and diseases. Physiol. Rev. 90, 1337-1381. 10.1152/physrev.00058.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell B. B., Ghaleb A. M., Nandan M. O. and Yang V. W. (2007). The diverse functions of Krüppel-like factors 4 and 5 in epithelial biology and pathobiology. Bioessays 29, 549-557. 10.1002/bies.20581 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell B. B., Bialkowska A. B., Nandan M. O., Ghaleb A. M., Gordon F. J. and Yang V. W. (2009). Haploinsufficiency of Kruppel-like factor 5 rescues the tumor-initiating effect of the Apc(Min) mutation in the intestine. Cancer Res. 69, 4125-4133. 10.1158/0008-5472.CAN-08-4402 [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell B. B., Kim S. S., Yu K., Ghaleb A. M., Takeda N., Manabe I., Nusrat A., Nagai R. and Yang V. W. (2011). Kruppel-like factor 5 is important for maintenance of crypt architecture and barrier function in mouse intestine. Gastroenterology 141, 1302-1313.e6. 10.1053/j.gastro.2011.06.086 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mehta T. S., Lu H., Wang X., Urvalek A. M., Nguyen K.-H. H., Monzur F., Hammond J. D., Ma J. Q. and Zhao J. (2009). A unique sequence in the N-terminal regulatory region controls the nuclear localization of KLF8 by cooperating with the C-terminal zinc-fingers. Cell Res. 19, 1098-1109. 10.1038/cr.2009.64 [DOI] [PubMed] [Google Scholar]
- Miele L., Beale G., Patman G., Nobili V., Leathart J., Grieco A., Abate M., Friedman S. L., Narla G., Bugianesi E. et al. (2008). The Kruppel-like factor 6 genotype is associated with fibrosis in nonalcoholic fatty liver disease. Gastroenterology 135, 282-291.e281. 10.1053/j.gastro.2008.04.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyamoto S., Suzuki T., Muto S., Aizawa K., Kimura A., Mizuno Y., Nagino T., Imai Y., Adachi N., Horikoshi M. et al. (2003). Positive and negative regulation of the cardiovascular transcription factor KLF5 by p300 and the oncogenic regulator SET through interaction and acetylation on the DNA-binding domain. Mol. Cell. Biol. 23, 8528-8541. 10.1128/MCB.23.23.8528-8541.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moad M., Pal D., Hepburn A. C., Williamson S. C., Wilson L., Lako M., Armstrong L., Hayward S. W., Franco O. E., Cates J. M. et al. (2013). A novel model of urinary tract differentiation, tissue regeneration, and disease: reprogramming human prostate and bladder cells into induced pluripotent stem cells. Eur. Urol. 64, 753-761. 10.1016/j.eururo.2013.03.054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore D. L., Blackmore M. G., Hu Y., Kaestner K. H., Bixby J. L., Lemmon V. P. and Goldberg J. L. (2009). KLF family members regulate intrinsic axon regeneration ability. Science 326, 298-301. 10.1126/science.1175737 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mori T., Sakaue H., Iguchi H., Gomi H., Okada Y., Takashima Y., Nakamura K., Nakamura T., Yamauchi T., Kubota N. et al. (2005). Role of Kruppel-like factor 15 (KLF15) in transcriptional regulation of adipogenesis. J. Biol. Chem. 280, 12867-12875. 10.1074/jbc.M410515200 [DOI] [PubMed] [Google Scholar]
- Morita M., Kobayashi A., Yamashita T., Shimanuki T., Nakajima O., Takahashi S., Ikegami S., Inokuchi K., Yamashita K., Yamamoto M. et al. (2003). Functional analysis of basic transcription element binding protein by gene targeting technology. Mol. Cell. Biol. 23, 2489-2500. 10.1128/MCB.23.7.2489-2500.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagai R. Z., Friedman S. L. and Kasuga M. (2009). The Biology of Krüppel-like Factors. Tokyo: Springer. [Google Scholar]
- Nakagawa M., Koyanagi M., Tanabe K., Takahashi K., Ichisaka T., Aoi T., Okita K., Mochiduki Y., Takizawa N. and Yamanaka S. (2008). Generation of induced pluripotent stem cells without Myc from mouse and human fibroblasts. Nat. Biotechnol. 26, 101-106. 10.1038/nbt1374 [DOI] [PubMed] [Google Scholar]
- Nakamura Y., Migita T., Hosoda F., Okada N., Gotoh M., Arai Y., Fukushima M., Ohki M., Miyata S., Takeuchi K. et al. (2009). Krüppel-like factor 12 plays a significant role in poorly differentiated gastric cancer progression. Int. J. Cancer 125, 1859-1867. 10.1002/ijc.24538 [DOI] [PubMed] [Google Scholar]
- Nandan M. O., McConnell B. B., Ghaleb A. M., Bialkowska A. B., Sheng H., Shao J., Babbin B. A., Robine S. and Yang V. W. (2008). Krüppel-like factor 5 mediates cellular transformation during oncogenic KRAS-induced intestinal tumorigenesis. Gastroenterology 134, 120-130. 10.1053/j.gastro.2007.10.023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nandan M. O., Ghaleb A. M., McConnell B. B., Patel N. V., Robine S. and Yang V. W. (2010). Krüppel-like factor 5 is a crucial mediator of intestinal tumorigenesis in mice harboring combined ApcMin and KRASV12 mutations. Mol. Cancer 9, 63 10.1186/1476-4598-9-63 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Narla G., Heath K. E., Reeves H. L., Li D., Giono L. E., Kimmelman A. C., Glucksman M. J., Narla J., Eng F. J., Chan A. M. et al. (2001). KLF6, a candidate tumor suppressor gene mutated in prostate cancer. Science 294, 2563-2566. 10.1126/science.1066326 [DOI] [PubMed] [Google Scholar]
- Narla G., DiFeo A., Yao S., Banno A., Hod E., Reeves H. L., Qiao R. F., Camacho-Vanegas O., Levine A., Kirschenbaum A. et al. (2005). Targeted inhibition of the KLF6 splice variant, KLF6 SV1, suppresses prostate cancer cell growth and spread. Cancer Res. 65, 5761-5768. 10.1158/0008-5472.CAN-05-0217 [DOI] [PubMed] [Google Scholar]
- Neve B., Fernandez-Zapico M. E., Ashkenazi-Katalan V., Dina C., Hamid Y. H., Joly E., Vaillant E., Benmezroua Y., Durand E., Bakaher N. et al. (2005). Role of transcription factor KLF11 and its diabetes-associated gene variants in pancreatic beta cell function. Proc. Natl. Acad. Sci. USA 102, 4807-4812. 10.1073/pnas.0409177102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishimura K., Kato T., Chen C., Oinam L., Shiomitsu E., Ayakawa D., Ohtaka M., Fukuda A., Nakanishi M. and Hisatake K. (2014). Manipulation of KLF4 expression generates iPSCs paused at successive stages of reprogramming. Stem Cell Rep. 3, 915-929. 10.1016/j.stemcr.2014.08.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuez B., Michalovich D., Bygrave A., Ploemacher R. and Grosveld F. (1995). Defective haematopoiesis in fetal liver resulting from inactivation of the EKLF gene. Nature 375, 316-318. 10.1038/375316a0 [DOI] [PubMed] [Google Scholar]
- Ohnishi S., Laub F., Matsumoto N., Asaka M., Ramirez F., Yoshida T. and Terada M. (2000). Developmental expression of the mouse gene coding for the Krüppel-like transcription factor KLF5. Dev. Dyn. 217, 421-429. [DOI] [PubMed] [Google Scholar]
- Oishi Y., Manabe I., Tobe K., Tsushima K., Shindo T., Fujiu K., Nishimura G., Maemura K., Yamauchi T., Kubota N. et al. (2005). Krüppel-like transcription factor KLF5 is a key regulator of adipocyte differentiation. Cell Metab. 1, 27-39. 10.1016/j.cmet.2004.11.005 [DOI] [PubMed] [Google Scholar]
- Oishi Y., Manabe I., Tobe K., Ohsugi M., Kubota T., Fujiu K., Maemura K., Kubota N., Kadowaki T. and Nagai R. (2008). SUMOylation of Krüppel-like transcription factor 5 acts as a molecular switch in transcriptional programs of lipid metabolism involving PPAR-delta. Nat. Med. 14, 656-666. 10.1038/nm1756 [DOI] [PubMed] [Google Scholar]
- Outani H., Okada M., Hiramatsu K., Yoshikawa H. and Tsumaki N. (2011). Induction of chondrogenic cells from dermal fibroblast culture by defined factors does not involve a pluripotent state. Biochem. Biophys. Res. Commun. 411, 607-612. 10.1016/j.bbrc.2011.06.194 [DOI] [PubMed] [Google Scholar]
- Ouyang L., Chen X. and Bieker J. J. (1998). Regulation of erythroid Kruppel-like factor (EKLF) transcriptional activity by phosphorylation of a protein kinase casein kinase II site within its interaction domain. J. Biol. Chem. 273, 23019-23025. 10.1074/jbc.273.36.23019 [DOI] [PubMed] [Google Scholar]
- Park I.-H., Zhao R., West J. A., Yabuuchi A., Huo H., Ince T. A., Lerou P. H., Lensch M. W. and Daley G. Q. (2008). Reprogramming of human somatic cells to pluripotency with defined factors. Nature 451, 141-146. 10.1038/nature06534 [DOI] [PubMed] [Google Scholar]
- Parker-Katiraee L., Carson A. R., Yamada T., Arnaud P., Feil R., Abu-Amero S. N., Moore G. E., Kaneda M., Perry G. H., Stone A. C. et al. (2007). Identification of the imprinted KLF14 transcription factor undergoing human-specific accelerated evolution. PLoS Genet. 3, e65 10.1371/journal.pgen.0030065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pearson R. C., Funnell A. P. and Crossley M. (2011). The mammalian zinc finger transcription factor Kruppel-like factor 3 (KLF3/BKLF). IUBMB Life 63, 86-93. 10.1002/iub.422 [DOI] [PubMed] [Google Scholar]
- Perdomo J., Verger A., Turner J. and Crossley M. (2005). Role for SUMO modification in facilitating transcriptional repression by BKLF. Mol. Cell. Biol. 25, 1549-1559. 10.1128/MCB.25.4.1549-1559.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pérez-Monter C., Martínez-Armenta M., Miquelajauregui A., Furlan-Magaril M., Varela-Echavarría A., Recillas-Targa F., May V., Charli J.-L. and Pérez-Martínez L. (2011). The Krüppel-like factor 4 controls biosynthesis of thyrotropin-releasing hormone during hypothalamus development. Mol. Cell. Endocrinol. 333, 127-133. 10.1016/j.mce.2010.12.020 [DOI] [PubMed] [Google Scholar]
- Perkins A. C., Sharpe A. H. and Orkin S. H. (1995). Lethal beta-thalassaemia in mice lacking the erythroid CACCC-transcription factor EKLF. Nature 375, 318-322. 10.1038/375318a0 [DOI] [PubMed] [Google Scholar]
- Polo J. M., Anderssen E., Walsh R. M., Schwarz B. A., Nefzger C. M., Lim S. M., Borkent M., Apostolou E., Alaei S., Cloutier J. et al. (2012). A molecular roadmap of reprogramming somatic cells into iPS cells. Cell 151, 1617-1632. 10.1016/j.cell.2012.11.039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preiss A., Rosenberg U. B., Kienlin A., Seifert E. and Jäckle H. (1985). Molecular genetics of Krüppel, a gene required for segmentation of the Drosophila embryo. Nature 313, 27-32. 10.1038/313027a0 [DOI] [PubMed] [Google Scholar]
- Presnell J. S., Schnitzler C. E. and Browne W. E. (2015). KLF/SP transcription factor family evolution: expansion, diversification, and innovation in eukaryotes. Genome Biol. Evol. 7, 2289-2309. 10.1093/gbe/evv141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prosdocimo D. A., Anand P., Liao X., Zhu H., Shelkay S., Artero-Calderon P., Zhang L., Kirsh J., Moore D. V., Rosca M. G. et al. (2014). Kruppel-like factor 15 is a critical regulator of cardiac lipid metabolism. J. Biol. Chem. 289, 5914-5924. 10.1074/jbc.M113.531384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin S., Liu M., Niu W. and Zhang C.-L. (2011). Dysregulation of Kruppel-like factor 4 during brain development leads to hydrocephalus in mice. Proc. Natl. Acad. Sci. USA 108, 21117-21121. 10.1073/pnas.1112351109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin S., Zou Y. and Zhang C. L. (2013). Cross-talk between KLF4 and STAT3 regulates axon regeneration. Nat. Commun. 4, 2633 10.1038/ncomms3633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qin J., Zhou Z., Chen W., Wang C., Zhang H., Ge G., Shao M., You D., Fan Z., Xia H. et al. (2015). BAP1 promotes breast cancer cell proliferation and metastasis by deubiquitinating KLF5. Nat. Commun. 6, 8471 10.1038/ncomms9471 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quadrini K. J. and Bieker J. J. (2002). Kruppel-like zinc fingers bind to nuclear import proteins and are required for efficient nuclear localization of erythroid Kruppel-like factor. J. Biol. Chem. 277, 32243-32252. 10.1074/jbc.M205677200 [DOI] [PubMed] [Google Scholar]
- Rajamannan N. M., Subramaniam M., Abraham T. P., Vasile V. C., Ackerman M. J., Monroe D. G., Chew T.-L. and Spelsberg T. C. (2007). TGFbeta inducible early gene-1 (TIEG1) and cardiac hypertrophy: discovery and characterization of a novel signaling pathway. J. Cell. Biochem. 100, 315-325. 10.1002/jcb.21049 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ratziu V., Lalazar A., Wong L., Dang Q., Collins C., Shaulian E., Jensen S. and Friedman S. L. (1998). Zf9, a Kruppel-like transcription factor up-regulated in vivo during early hepatic fibrosis. Proc. Natl. Acad. Sci. USA 95, 9500-9505. 10.1073/pnas.95.16.9500 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodriguez E., Aburjania N., Priedigkeit N. M., DiFeo A. and Martignetti J. A. (2010). Nucleo-cytoplasmic localization domains regulate Krüppel-like factor 6 (KLF6) protein stability and tumor suppressor function. PLoS ONE 5, e12639 10.1371/journal.pone.0012639 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rozenblum E., Vahteristo P., Sandberg T., Bergthorsson J., Syrjakoski K., Weaver D., Haraldsson K., Johannsdottir H., Vehmanen P., Nigam S. et al. (2002). A genomic map of a 6-Mb region at 13q21-q22 implicated in cancer development: identification and characterization of candidate genes. Hum. Genet. 110, 111-121. 10.1007/s00439-001-0646-6 [DOI] [PubMed] [Google Scholar]
- Schuettpelz L. G., Gopalan P. K., Giuste F. O., Romine M. P., van Os R. and Link D. C. (2012). Kruppel-like factor 7 overexpression suppresses hematopoietic stem and progenitor cell function. Blood 120, 2981-2989. 10.1182/blood-2012-02-409839 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuierer M., Hilger-Eversheim K., Dobner T., Bosserhoff A.-K., Moser M., Turner J., Crossley M. and Buettner R. (2001). Induction of AP-2alpha expression by adenoviral infection involves inactivation of the AP-2rep transcriptional corepressor CtBP1. J. Biol. Chem. 276, 27944-27949. 10.1074/jbc.M100070200 [DOI] [PubMed] [Google Scholar]
- Scobie K. N., Hall B. J., Wilke S. A., Klemenhagen K. C., Fujii-Kuriyama Y., Ghosh A., Hen R. and Sahay A. (2009). Kruppel-like factor 9 is necessary for late-phase neuronal maturation in the developing dentate gyrus and during adult hippocampal neurogenesis. J. Neurosci. 29, 9875-9887. 10.1523/JNEUROSCI.2260-09.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scohy S., Gabant P., Van Reeth T., Hertveldt V., Drèze P.-L., Van Vooren P., Rivière M., Szpirer J. and Szpirer C. (2000). Identification of KLF13 and KLF14 (SP6), novel members of the SP/XKLF transcription factor family. Genomics 70, 93-101. 10.1006/geno.2000.6362 [DOI] [PubMed] [Google Scholar]
- Segre J. A., Bauer C. and Fuchs E. (1999). Klf4 is a transcription factor required for establishing the barrier function of the skin. Nat. Genet. 22, 356-400. 10.1038/11926 [DOI] [PubMed] [Google Scholar]
- Seiler K., Soroush Noghabi M., Karjalainen K., Hummel M., Melchers F. and Tsuneto M. (2011). Induced pluripotent stem cells expressing elevated levels of sox-2, oct-4, and klf-4 are severely reduced in their differentiation from mesodermal to hematopoietic progenitor cells. Stem Cells Dev. 20, 1131-1142. 10.1089/scd.2010.0391 [DOI] [PubMed] [Google Scholar]
- Shahrin N. H., Diakiw S., Dent L. A., Brown A. L. and D'Andrea R. J. (2016). Conditional knockout mice demonstrate function of Klf5 as a myeloid transcription factor. Blood 128, 55-59. 10.1182/blood-2015-12-684514 [DOI] [PubMed] [Google Scholar]
- Shankman L. S., Gomez D., Cherepanova O. A., Salmon M., Alencar G. F., Haskins R. M., Swiatlowska P., Newman A. A. C., Greene E. S., Straub A. C. et al. (2015). KLF4-dependent phenotypic modulation of smooth muscle cells has a key role in atherosclerotic plaque pathogenesis. Nat. Med. 21, 628-637. 10.1038/nm.3866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shields J. M. and Yang V. W. (1997). Two potent nuclear localization signals in the gut-enriched Kruppel-like factor define a subfamily of closely related Kruppel proteins. J. Biol. Chem. 272, 18504-18507. 10.1074/jbc.272.29.18504 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shindo T., Manabe I., Fukushima Y., Tobe K., Aizawa K., Miyamoto S., Kawai-Kowase K., Moriyama N., Imai Y., Kawakami H. et al. (2002). Kruppel-like zinc-finger transcription factor KLF5/BTEB2 is a target for angiotensin II signaling and an essential regulator of cardiovascular remodeling. Nat. Med. 8, 856-863. 10.1038/nm738 [DOI] [PubMed] [Google Scholar]
- Shinoda Y., Ogata N., Higashikawa A., Manabe I., Shindo T., Yamada T., Kugimiya F., Ikeda T., Kawamura N., Kawasaki Y. et al. (2008). Kruppel-like factor 5 causes cartilage degradation through transactivation of matrix metalloproteinase 9. J. Biol. Chem. 283, 24682-24689. 10.1074/jbc.M709857200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shum C. K. Y., Lau S. T., Tsoi L. L. S., Chan L. K., Yam J. W. P., Ohira M., Nakagawara A., Tam P. K. H. and Ngan E. S. W. (2013). Krüppel-like factor 4 (KLF4) suppresses neuroblastoma cell growth and determines non-tumorigenic lineage differentiation. Oncogene 32, 4086-4099. 10.1038/onc.2012.437 [DOI] [PubMed] [Google Scholar]
- Siatecka M. and Bieker J. J. (2011). The multifunctional role of EKLF/KLF1 during erythropoiesis. Blood 118, 2044-2054. 10.1182/blood-2011-03-331371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siatecka M., Xue L. and Bieker J. J. (2007). Sumoylation of EKLF promotes transcriptional repression and is involved in inhibition of megakaryopoiesis. Mol. Cell. Biol. 27, 8547-8560. 10.1128/MCB.00589-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmen R. C. M., Eason R. R., McQuown J. R., Linz A. L., Kang T.-J., Chatman L. Jr., Till S. R., Fujii-Kuriyama Y., Simmen F. A. and Oh S. P. (2004). Subfertility, uterine hypoplasia, and partial progesterone resistance in mice lacking the Kruppel-like factor 9/basic transcription element-binding protein-1 (Bteb1) gene. J. Biol. Chem. 279, 29286-29294. 10.1074/jbc.M403139200 [DOI] [PubMed] [Google Scholar]
- Simmen F. A., Xiao R., Velarde M. C., Nicholson R. D., Bowman M. T., Fujii-Kuriyama Y., Oh S. P. and Simmen R. C. M. (2007). Dysregulation of intestinal crypt cell proliferation and villus cell migration in mice lacking Kruppel-like factor 9. Am. J. Physiol. Gastrointest. Liver Physiol. 292, G1757-G1769. 10.1152/ajpgi.00013.2007 [DOI] [PubMed] [Google Scholar]
- Simmen R. C. M., Pabona J. M. P., Velarde M. C., Simmons C., Rahal O. and Simmen F. A. (2010). The emerging role of Kruppel-like factors in endocrine-responsive cancers of female reproductive tissues. J. Endocrinol. 204, 223-231. 10.1677/JOE-09-0329 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh V. K., Kalsan M., Kumar N., Saini A. and Chandra R. (2015). Induced pluripotent stem cells: applications in regenerative medicine, disease modeling, and drug discovery. Front. Cell Dev. Biol. 3, 1276 10.3389/fcell.2015.00002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Slavin D., Sapin V., Lopez-Diaz F., Jacquemin P., Koritschoner N., Dastugue B., Davidson I., Chatton B. and Bocco J. L. (1999). The Kruppel-like core promoter binding protein gene is primarily expressed in placenta during mouse development. Biol. Reprod. 61, 1586-1591. 10.1095/biolreprod61.6.1586 [DOI] [PubMed] [Google Scholar]
- Song A., Patel A., Thamatrakoln K., Liu C., Feng D., Clayberger C. and Krensky A. M. (2002). Functional domains and DNA-binding sequences of RFLAT-1/KLF13, a Kruppel-like transcription factor of activated T lymphocytes. J. Biol. Chem. 277, 30055-30065. 10.1074/jbc.M204278200 [DOI] [PubMed] [Google Scholar]
- Song C.-Z., Gavriilidis G., Asano H. and Stamatoyannopoulos G. (2005). Functional study of transcription factor KLF11 by targeted gene inactivation. Blood Cells Mol. Dis. 34, 53-59. 10.1016/j.bcmd.2004.08.027 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soufi A., Donahue G. and Zaret K. S. (2012). Facilitators and impediments of the pluripotency reprogramming factors’ initial engagement with the genome. Cell 151, 994-1004. 10.1016/j.cell.2012.09.045 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stacey S. N., Sulem P., Masson G., Gudjonsson S. A., Thorleifsson G., Jakobsdottir M., Sigurdsson A., Gudbjartsson D. F., Sigurgeirsson B., Benediktsdottir K. R. et al. (2009). New common variants affecting susceptibility to basal cell carcinoma. Nat. Genet. 41, 909-914. 10.1038/ng.412 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stamatakis A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22, 2688-2690. 10.1093/bioinformatics/btl446 [DOI] [PubMed] [Google Scholar]
- Subramaniam M., Harris S. A., Oursler M. J., Rasmussen K., Riggs B. L. and Spelsberg T. C. (1995). Identification of a novel TGF-beta-regulated gene encoding a putative zinc finger protein in human osteoblasts. Nucleic Acids Res. 23, 4907-4912. 10.1093/nar/23.23.4907 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramaniam M., Gorny G., Johnsen S. A., Monroe D. G., Evans G. L., Fraser D. G., Rickard D. J., Rasmussen K., van Deursen J. M. A., Turner R. T. et al. (2005). TIEG1 null mouse-derived osteoblasts are defective in mineralization and in support of osteoclast differentiation in vitro. Mol. Cell. Biol. 25, 1191-1199. 10.1128/MCB.25.3.1191-1199.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suda S., Rai T., Sohara E., Sasaki S. and Uchida S. (2006). Postnatal expression of KLF12 in the inner medullary collecting ducts of kidney and its trans-activation of UT-A1 urea transporter promoter. Biochem. Biophys. Res. Commun. 344, 246-252. 10.1016/j.bbrc.2006.03.138 [DOI] [PubMed] [Google Scholar]
- Sue N., Jack B. H. A., Eaton S. A., Pearson R. C. M., Funnell A. P. W., Turner J., Czolij R., Denyer G., Bao S., Molero-Navajas J. C. et al. (2008). Targeted disruption of the basic Kruppel-like factor gene (Klf3) reveals a role in adipogenesis. Mol. Cell. Biol. 28, 3967-3978. 10.1128/MCB.01942-07 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sur I., Rozell B., Jaks V., Bergström A. and Toftgård R. (2006). Epidermal and craniofacial defects in mice overexpressing Klf5 in the basal layer of the epidermis. J. Cell Sci. 119, 3593-3601. 10.1242/jcs.03070 [DOI] [PubMed] [Google Scholar]
- Suske G., Bruford E. and Philipsen S. (2005). Mammalian SP/KLF transcription factors: bring in the family. Genomics 85, 551-556. 10.1016/j.ygeno.2005.01.005 [DOI] [PubMed] [Google Scholar]
- Takagi K., Miki Y., Onodera Y., Nakamura Y., Ishida T., Watanabe M., Inoue S., Sasano H. and Suzuki T. (2012). Kruppel-like factor 5 in human breast carcinoma: a potent prognostic factor induced by androgens. Endocr. Relat. Cancer 19, 741-750. 10.1530/ERC-12-0017 [DOI] [PubMed] [Google Scholar]
- Takahashi K. and Yamanaka S. (2006). Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 126, 663-676. 10.1016/j.cell.2006.07.024 [DOI] [PubMed] [Google Scholar]
- Takahashi K., Tanabe K., Ohnuki M., Narita M., Ichisaka T., Tomoda K. and Yamanaka S. (2007). Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 131, 861-872. 10.1016/j.cell.2007.11.019 [DOI] [PubMed] [Google Scholar]
- Takeda N., Manabe I., Uchino Y., Eguchi K., Matsumoto S., Nishimura S., Shindo T., Sano M., Otsu K., Snider P. et al. (2010). Cardiac fibroblasts are essential for the adaptive response of the murine heart to pressure overload. J. Clin. Invest. 120, 254-265. 10.1172/JCI40295 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tetreault M.-P., Yang Y. and Katz J. P. (2013). Krüppel-like factors in cancer. Nat. Rev. Cancer 13, 701-713. 10.1038/nrc3582 [DOI] [PubMed] [Google Scholar]
- Ton-That H., Kaestner K. H., Shields J. M., Mahatanankoon C. S. and Yang V. W. (1997). Expression of the gut-enriched Krüppel-like factor gene during development and intestinal tumorigenesis. FEBS Lett. 419, 239-243. 10.1016/S0014-5793(97)01465-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tong D., Czerwenka K., Heinze G., Ryffel M., Schuster E., Witt A., Leodolter S. and Zeillinger R. (2006). Expression of KLF5 is a prognostic factor for disease-free survival and overall survival in patients with breast cancer. Clin. Cancer Res. 12, 2442-2448. 10.1158/1078-0432.CCR-05-0964 [DOI] [PubMed] [Google Scholar]
- Truty M. J., Lomberk G., Fernandez-Zapico M. E. and Urrutia R. (2009). Silencing of the transforming growth factor-beta (TGFbeta) receptor II by Kruppel-like factor 14 underscores the importance of a negative feedback mechanism in TGFbeta signaling. J. Biol. Chem. 284, 6291-6300. 10.1074/jbc.M807791200 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turchinovich G., Vu T. T., Frommer F., Kranich J., Schmid S., Alles M., Loubert J.-B., Goulet J.-P., Zimber-Strobl U., Schneider P. et al. (2011). Programming of marginal zone B-cell fate by basic Kruppel-like factor (BKLF/KLF3). Blood 117, 3780-3792. 10.1182/blood-2010-09-308742 [DOI] [PubMed] [Google Scholar]
- Turner J. and Crossley M. (1998). Cloning and characterization of mCtBP2, a co-repressor that associates with basic Kruppel-like factor and other mammalian transcriptional regulators. EMBO J. 17, 5129-5140. 10.1093/emboj/17.17.5129 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuuminen R., Nykänen A. I., Saharinen P., Gautam P., Keränen M. A. I., Arnaudova R., Rouvinen E., Helin H., Tammi R., Rilla K. et al. (2013). Donor simvastatin treatment prevents ischemia-reperfusion and acute kidney injury by preserving microvascular barrier function. Am. J. Transplant. 13, 2019-2034. 10.1111/ajt.12315 [DOI] [PubMed] [Google Scholar]
- Uchida S., Tanaka Y., Ito H., Saitoh-Ohara F., Inazawa J., Yokoyama K. K., Sasaki S. and Marumo F. (2000). Transcriptional regulation of the CLC-K1 promoter by myc-associated zinc finger protein and kidney-enriched Kruppel-like factor, a novel zinc finger repressor. Mol. Cell. Biol. 20, 7319-7331. 10.1128/MCB.20.19.7319-7331.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Vliet J., Turner J. and Crossley M. (2000). Human Kruppel-like factor 8: a CACCC-box binding protein that associates with CtBP and represses transcription. Nucleic Acids Res. 28, 1955-1962. 10.1093/nar/28.9.1955 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Vliet J., Crofts L. A., Quinlan K. G. R., Czolij R., Perkins A. C. and Crossley M. (2006). Human KLF17 is a new member of the Sp/KLF family of transcription factors. Genomics 87, 474-482. 10.1016/j.ygeno.2005.12.011 [DOI] [PubMed] [Google Scholar]
- van Zelm M. C., van der Burg M., de Ridder D., Barendregt B. H., de Haas E. F. E., Reinders M. J. T., Lankester A. C., Revesz T., Staal F. J. T. and van Dongen J. J. M. (2005). Ig gene rearrangement steps are initiated in early human precursor B cell subsets and correlate with specific transcription factor expression. J. Immunol. 175, 5912-5922. 10.4049/jimmunol.175.9.5912 [DOI] [PubMed] [Google Scholar]
- Venuprasad K., Huang H., Harada Y., Elly C., Subramaniam M., Spelsberg T., Su J. and Liu Y.-C. (2008). The E3 ubiquitin ligase Itch regulates expression of transcription factor Foxp3 and airway inflammation by enhancing the function of transcription factor TIEG1. Nat. Immunol. 9, 245-253. 10.1038/ni1564 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vinjamur D. S., Wade K. J., Mohamad S. F., Haar J. L., Sawyer S. T. and Lloyd J. A. (2014). Kruppel-like transcription factors KLF1 and KLF2 have unique and coordinate roles in regulating embryonic erythroid precursor maturation. Haematologica 99, 1565-1573. 10.3324/haematol.2014.104943 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan H., Luo F., Wert S. E., Zhang L., Xu Y., Ikegami M., Maeda Y., Bell S. M. and Whitsett J. A. (2008). Kruppel-like factor 5 is required for perinatal lung morphogenesis and function. Development 135, 2563-2572. 10.1242/dev.021964 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X. and Zhao J. (2007). KLF8 transcription factor participates in oncogenic transformation. Oncogene 26, 456-461. 10.1038/sj.onc.1209796 [DOI] [PubMed] [Google Scholar]
- Wang B., Haldar S. M., Lu Y., Ibrahim O. A., Fisch S., Gray S., Leask A. and Jain M. K. (2008). The Kruppel-like factor KLF15 inhibits connective tissue growth factor (CTGF) expression in cardiac fibroblasts. J. Mol. Cell. Cardiol. 45, 193-197. 10.1016/j.yjmcc.2008.05.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang R., Wang Y., Liu N., Ren C., Jiang C., Zhang K., Yu S., Chen Y., Tang H., Deng Q. et al. (2013). FBW7 regulates endothelial functions by targeting KLF2 for ubiquitination and degradation. Cell Res. 23, 803-819. 10.1038/cr.2013.42 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang X., Shen Q. W., Wang J., Zhang Z., Feng F., Chen T., Zhang Y., Wei H., Li Z., Wang X. et al. (2016). KLF7 regulates satellite cell quiescence in response to extracellular signaling. Stem Cells 34, 1310-1320. 10.1002/stem.2346 [DOI] [PubMed] [Google Scholar]
- Wani M. A., Conkright M. D., Jeffries S., Hughes M. J. and Lingrel J. B. (1999a). cDNA isolation, genomic structure, regulation, and chromosomal localization of human lung Kruppel-like factor. Genomics 60, 78-86. 10.1006/geno.1999.5888 [DOI] [PubMed] [Google Scholar]
- Wani M. A., Wert S. E. and Lingrel J. B. (1999b). Lung Kruppel-like factor, a zinc finger transcription factor, is essential for normal lung development. J. Biol. Chem. 274, 21180-21185. 10.1074/jbc.274.30.21180 [DOI] [PubMed] [Google Scholar]
- Wara A. K., Foo S., Croce K., Sun X., Icli B., Tesmenitsky Y., Esen F., Lee J.-S., Subramaniam M., Spelsberg T. C. et al. (2011). TGF-beta1 signaling and Kruppel-like factor 10 regulate bone marrow-derived proangiogenic cell differentiation, function, and neovascularization. Blood 118, 6450-6460. 10.1182/blood-2011-06-363713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warburton D., El-Hashash A., Carraro G., Tiozzo C., Sala F., Rogers O., De Langhe S., Kemp P. J., Riccardi D., Torday J. et al. (2010). Lung organogenesis. Curr. Top. Dev. Biol. 90, 73-158. 10.1016/S0070-2153(10)90003-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wei H., Wang X., Gan B., Urvalek A. M., Melkoumian Z. K., Guan J.-L. and Zhao J. (2006). Sumoylation delimits KLF8 transcriptional activity associated with the cell cycle regulation. J. Biol. Chem. 281, 16664-16671. 10.1074/jbc.M513135200 [DOI] [PubMed] [Google Scholar]
- Wei D., Kanai M., Jia Z., Le X. and Xie K. (2008). Kruppel-like factor 4 induces p27Kip1 expression in and suppresses the growth and metastasis of human pancreatic cancer cells. Cancer Res. 68, 4631-4639. 10.1158/0008-5472.CAN-07-5953 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J., Srinivasan S. V., Neumann J. C. and Lingrel J. B. (2005). The KLF2 transcription factor does not affect the formation of preadipocytes but inhibits their differentiation into adipocytes. Biochemistry 44, 11098-11105. 10.1021/bi050166i [DOI] [PubMed] [Google Scholar]
- Wu J., Bohanan C. S., Neumann J. C. and Lingrel J. B. (2008). KLF2 transcription factor modulates blood vessel maturation through smooth muscle cell migration. J. Biol. Chem. 283, 3942-3950. 10.1074/jbc.M707882200 [DOI] [PubMed] [Google Scholar]
- Xie P., Tang Y., Shen S., Wang Y., Xing G., Yin Y., He F. and Zhang L. (2011). Smurf1 ubiquitin ligase targets Kruppel-like factor KLF2 for ubiquitination and degradation in human lung cancer H1299 cells. Biochem. Biophys. Res. Commun. 407, 254-259. 10.1016/j.bbrc.2011.03.016 [DOI] [PubMed] [Google Scholar]
- Yajima S., Lammers C. H., Lee S. H., Hara Y., Mizuno K. and Mouradian M. M. (1997). Cloning and characterization of murine glial cell-derived neurotrophic factor inducible transcription factor (MGIF). J. Neurosci. 17, 8657-8666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan W., Burns K. H., Ma L. and Matzuk M. M. (2002). Identification of Zfp393, a germ cell-specific gene encoding a novel zinc finger protein. Mech. Dev. 118, 233-239. 10.1016/S0925-4773(02)00258-7 [DOI] [PubMed] [Google Scholar]
- Yang R., Jiang M., Kumar S. M., Xu T., Wang F., Xiang L. and Xu X. (2011). Generation of melanocytes from induced pluripotent stem cells. J. Invest. Dermatol. 131, 2458-2466. 10.1038/jid.2011.242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yerges L. M., Klei L., Cauley J. A., Roeder K., Kammerer C. M., Ensrud K. E., Nestlerode C. S., Lewis C., Lang T. F., Barrett-Connor E. et al. (2010). Candidate gene analysis of femoral neck trabecular and cortical volumetric bone mineral density in older men. J. Bone Miner. Res. 25, 330-338. 10.1359/jbmr.090729 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yien Y. Y. and Bieker J. J. (2012). Functional interactions between erythroid Kruppel-like factor (EKLF/KLF1) and protein phosphatase PPM1B/PP2Cbeta. J. Biol. Chem. 287, 15193-15204. 10.1074/jbc.M112.350496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yien Y. Y., Gnanapragasam M. N., Gupta R., Rivella S. and Bieker J. J. (2015). Alternative splicing of EKLF/KLF1 in murine primary erythroid tissues. Exp. Hematol. 43, 65-70. 10.1016/j.exphem.2014.08.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yoshida T., Gan Q., Franke A. S., Ho R., Zhang J., Chen Y. E., Hayashi M., Majesky M. W., Somlyo A. V. and Owens G. K. (2010). Smooth and cardiac muscle-selective knock-out of Krüppel-like factor 4 causes postnatal death and growth retardation. J. Biol. Chem. 285, 21175-21184. 10.1074/jbc.M110.112482 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zammarchi F., Morelli M., Menicagli M., Di Cristofano C., Zavaglia K., Paolucci A., Campani D., Aretini P., Boggi U., Mosca F. et al. (2011). KLF4 is a novel candidate tumor suppressor gene in pancreatic ductal carcinoma. Am. J. Pathol. 178, 361-372. 10.1016/j.ajpath.2010.11.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z. and Teng C. T. (2003). Phosphorylation of Kruppel-like factor 5 (KLF5/IKLF) at the CBP interaction region enhances its transactivation function. Nucleic Acids Res. 31, 2196-2208. 10.1093/nar/gkg310 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang J.-S., Moncrieffe M. C., Kaczynski J., Ellenrieder V., Prendergast F. G. and Urrutia R. (2001a). A conserved alpha-helical motif mediates the interaction of Sp1-like transcriptional repressors with the corepressor mSin3A. Mol. Cell. Biol. 21, 5041-5049. 10.1128/MCB.21.15.5041-5049.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang W., Kadam S., Emerson B. M. and Bieker J. J. (2001b). Site-specific acetylation by p300 or CREB binding protein regulates erythroid Kruppel-like factor transcriptional activity via its interaction with the SWI-SNF complex. Mol. Cell. Biol. 21, 2413-2422. 10.1128/MCB.21.7.2413-2422.2001 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang X., Srinivasan S. V. and Lingrel J. B. (2004). WWP1-dependent ubiquitination and degradation of the lung Krüppel-like factor, KLF2. Biochem. Biophys. Res. Commun. 316, 139-148. 10.1016/j.bbrc.2004.02.033 [DOI] [PubMed] [Google Scholar]
- Zhang P., Basu P., Redmond L. C., Morris P. E., Rupon J. W., Ginder G. D. and Lloyd J. A. (2005). A functional screen for Krüppel-like factors that regulate the human gamma-globin gene through the CACCC promoter element. Blood Cells Mol. Dis. 35, 227-235. 10.1016/j.bcmd.2005.04.009 [DOI] [PubMed] [Google Scholar]
- Zhang P., Andrianakos R., Yang Y., Liu C. and Lu W. (2010). Kruppel-like factor 4 (Klf4) prevents embryonic stem (ES) cell differentiation by regulating Nanog gene expression. J. Biol. Chem. 285, 9180-9189. 10.1074/jbc.M109.077958 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang P., Ha T., Larouche M., Swanson D. and Goldowitz D. (2015). Kruppel-like factor 4 regulates granule cell Pax6 expression and cell proliferation in early cerebellar development. PLoS ONE 10, e0134390 10.1371/journal.pone.0134390 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao D., Zheng H.-Q., Zhou Z. and Chen C. (2010). The Fbw7 tumor suppressor targets KLF5 for ubiquitin-mediated degradation and suppresses breast cell proliferation. Cancer Res. 70, 4728-4738. 10.1158/0008-5472.CAN-10-0040 [DOI] [PubMed] [Google Scholar]
- Zhao T., Liu C. and Chen L. (2015). Roles of Klf5 acetylation in the self-renewal and the differentiation of mouse embryonic stem cells. PLoS ONE 10, e0138168 10.1371/journal.pone.0138168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong F., Chen H., Wei C., Zhang W., Li Z., Jain M. K., Chuang P. Y., Chen H., Wang Y., Mallipattu S. K. et al. (2015). Reduced Krüppel-like factor 2 expression may aggravate the endothelial injury of diabetic nephropathy. Kidney Int. 87, 382-395. 10.1038/ki.2014.286 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong F., Mallipattu S. K., Estrada C., Menon M., Salem F., Jain M. K., Chen H., Wang Y., Lee K. and He J. C. (2016). Reduced Krüppel-like factor 2 aggravates glomerular endothelial cell injury and kidney disease in mice with unilateral nephrectomy. Am. J. Pathol. 186, 2021-2031. 10.1016/j.ajpath.2016.03.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou M., McPherson L., Feng D., Song A., Dong C., Lyu S.-C., Zhou L., Shi X., Ahn Y.-T., Wang D. et al. (2007). Kruppel-like transcription factor 13 regulates T lymphocyte survival in vivo. J. Immunol. 178, 5496-5504. 10.4049/jimmunol.178.9.5496 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zobel D. P., Andreasen C. H., Burgdorf K. S., Andersson E. A., Sandbaek A., Lauritzen T., Borch-Johnsen K., Jorgensen T., Maeda S., Nakamura Y. et al. (2009). Variation in the gene encoding Kruppel-like factor 7 influences body fat: studies of 14 818 Danes. Eur. J. Endocrinol. 160, 603-609. 10.1530/EJE-08-0688 [DOI] [PubMed] [Google Scholar]